
Abstract
Introduction
This study aimed to investigate changes in renal function and the AGE-RAGE axis in the kidney of a non-genetic animal model of metabolic syndrome (MetS) induced by high-fat diet (HFD). Additionally, we evaluated the protective effect of pyridoxamine (PM), a vitamin B6 analog with anti-AGE effects, in the context of diet-related renal endothelial dysfunction.
Methodology
In Wistar rats, the MetS animal model was induced by 20 or 28 weeks of HFD feeding. When indicated, a subgroup of animals was treated daily with PM (60 mg/kg) for 2 months. Tissue perfusion in renal microcirculation was examined by laser speckle contrast imaging. Oxidative stress was analyzed by thiobarbituric acid reactive species and the inflammatory markers by ELISA (TNF-α and IL-1β). Reverse transcription polymerase chain reaction was used to analyze eNOs, IL-6, vascular cell adhesion molecule (VCAM), NADPH oxidase subunit 47 (N47), catalase, and receptor for AGE (RAGE) gene expression.
Results
Wistar rats fed a HFD showed negligible alteration in renal function, decrease in catalase mRNA transcripts and catalase enzyme activity compared to control (CTL) animals. Increased levels of IL-1β were observed in the kidney of MetS-induced rats. HFD-fed rats exhibited kidney endothelial dysfunction, with no significant differences in basal microvascular blood flow. PM significantly improved kidney vasorelaxation in HFD-fed rats. eNOS, VCAM, and RAGE gene expression and AGE content were not altered in kidneys of HFD-induced MetS rats in comparison to CTLs.
Conclusions
Our findings suggest that HFD-induced microvascular dysfunction precedes the decline in renal function, and could be related to antioxidant machinery defects and inflammation activation in the kidney. PM showed a vasoprotective effect, and thus, could be an important contributory factor in ameliorating diet-induced renal damage.
Introduction
The metabolic syndrome (MetS) is a combination of metabolic conditions, comprising of elevated blood pressure, obesity, atherogenic dyslipidemia, and dysglycemia, associated with increased risk of type 2 diabetes and atherosclerotic cardiovascular disease.Citation1,Citation2 MetS is a recurrent public health problem with increasing prevalence during the last three decades, mainly due to the global spread of the Western lifestyle.Citation3,Citation4 Individuals with MetS are at high risk of developing chronic kidney disease (CKD), which is an independent risk factor for cardiovascular morbidity and mortality.Citation5–Citation8 Also, one of the most frequent microvascular complications of diabetes is the diabetic nephropathy, which progresses to end-stage renal disease in one-third of patients with type 1 diabetes (T1DM) and 20% of the patients with type 2 diabetes (T2DM).Citation9 Therefore, the prevention of renal alterations and/or CKD is critical to improve cardio-renal outcomes in MetS and diabetes patients. To develop a preventative strategy against CKD in these patients, it is essential to clarify renal alterations in MetS, including oxidative, inflammatory, and endothelial function status. However, these have not been yet thoroughly evaluated, and the mechanisms by which MetS may influence kidney dysfunction remain unclear.
Currently established therapeutic approaches for diabetic nephropathy are primarily antihypertensives that target the renin angiotensin aldosterone system, including angiotensin-converting enzyme inhibitors (iECA) and angiotensin II receptor blockers.Citation10 These interventions appear to reduce or delay the proteinuria but do not prevent the establishment of end-stage renal disease.Citation11,Citation12 Therefore, the primary challenge is to identify pharmacological and non-pharmacological treatments that are able to prevent the progression of renal dysfunction.
Pyridoxamine (PM) is a vitamin B6 analog that exerts antiglycative effect. It has been proposed as supplementary approach in patients with renal diseases as it has demonstrated effectiveness in impairing the development of diabetic nephropathy, in both patients and experimental models of diabetes.Citation11–Citation15 PM is effective in inhibiting advanced glycation end products (AGE) or advanced lipoxidation end products (ALE) formation from Amadori adducts by trapping pathogenic reactive carbonyl intermediates of AGE/ALE formation.Citation16–Citation18 AGE/ALE are irreversible products expressed ubiquitously and overproduced in pathologies such as diabetes, CKD, Alzheimer disease, and non-alcoholic fatty liver disease.Citation19–Citation22 Additionally, PM has anti-inflammatory, antioxidant, and metabolic effects, which makes it a promising candidate for the management of MetS related renal alterations.Citation23 However, the effect of PM on renal endothelial dysfunction due to high-fat diet (HFD)-induced MetS remains unveiled.
Large number of associative studies between MetS and renal ED are available; however, a causal relationship between them is not proven.Citation24 The major factors contributing to endothelial dysfunction and/or renal damage in MetS and related diseases are increases in oxidative stress and inflammatory cytokines which could be due to (a) enlargement of white adipose tissue;Citation25–Citation27 (b) insulin resistance; (c) increase in TGL and FFA; (d) activation of AGE-RAGE axis;Citation28 (e) increased angiotensin II production,Citation29 and (f) hyperuricemia.Citation30 Dissecting out the relative contribution of each of those mechanisms to MetS pathophysiology, however, is difficult. Since kidney endothelial dysfunction is a hallmark of MetS and has its physiopathology is not fully understood. Our study aims to investigate (a) selective inflammatory and pro-oxidant signaling pathways as possible participants in the physiopathology of MetS induced renal alterations and (b) whether PM treatment would exert renoprotective effects in renal endothelial dysfunction triggered by HFD consumption.
Materials and methods
Animals and experimental protocol
Twenty male Wistar rats (4 weeks of age) from the central animal facility of the Oswaldo Cruz Foundation, Brazil were housed in standard cages in a controlled environment (22±1°C temperature and 12 hrs, light/dark). MetS was induced in 10 animals by 20 weeks of feeding with a HFD consisted of a standard rat diet modified to contain 30% fat (lard), 56% carbohydrate, and 14% protein (% g) ().Citation20 The non-MetS control (CTL) animals received standard rat diet (n=10) for 20 weeks. For the study of PM effect on endothelial dysfunction, twenty-four (24) Wistar rats were used, divided into three experimental groups: the non-MetS CTL animals, which received standard rat diet for 28 weeks (n=8), the HFD untreated animals, which received HFD for 28 weeks and were treated with vehicle between weeks 20–28 (HFD, n=9), and treated HFD-fed animals, which received PM (60 mg/kg/day, by gavage) also between weeks 20–28 (HFD + PM, n=7) (). Rats were provided with diet and water ad libitum. Animals under anesthesia were subjected to laser speckle contrast imaging (LSCI) evaluations, and then subjected to euthanasia by cardiac puncture. Total blood and kidney samples were collected and aliquots were stored at −80°C until measurement. Rats in each dietary group were placed in metabolic balance cages (Nalgene; Nalge Company, Rochester, NY, USA) for 24-hr urine collection to measure microalbuminuria and urine volume. All experiments were conducted according to the internationally accepted principles for the Care and Use of Laboratory Animals. Experimental protocols were approved by the Oswaldo Cruz Foundation Animal Welfare Committee (CEUA license L-034/2016).
Figure 1 Schematic representation of the experimental protocol used in the present study to assess the effect of pyridoxamine (PM) treatment on endothelial function.
Laser speckle contrast imaging (LSCI)
Kidney microvascular blood flow was measured by LSCI apparatus (Pericam PSI system, Perimed, Sweden).Citation20,Citation31 Overnight-fasted animals were anesthetized with ketamine (100 mg/kg, i.p.) and xylazine (10 mg/kg, i.p). Body temperature was continuously measured and maintained at 37.5°C by placing the animals on an electric heating pad (Harvard Apparatus, MA, United States) which adjust the body temperature in response to measured rectal temperatures. Left Kidney was exposed to laser light system for continuous measurement of tissue perfusion. Acetylcholine (Ach)(100 µL, 10-1 M) was applied directly on the kidney surface after 5 mins of microcirculatory flux stabilization. Relative kidney blood flow was expressed as arbitrary perfusion units.
Oxidative stress parameters
Lipid peroxidation in the kidney was assessed by measuring thiobarbituric acid reactive species as previously described.Citation20 Catalase activity was determined in the kidneys by measuring the enzymatic decomposition of hydrogen peroxide at 240 nm.Citation20
TNF-α and IL-1β measurements
TNF-α and IL-1β levels were measured in kidney samples with ELISA assay (R&D Systems, Minneapolis, USA).
Quantitative real-time polymerase chain reaction
Total RNA was extracted from the kidney using RNeasy Mini Kit (Qiagen), and reverse-transcribed to cDNA using a high capacity cDNA reverse transcription Kit (Applied Biosystems). The primers used for PCR amplification are described in . Real-time PCR was performed with the power SYBR Green PCR Master Mix (Applied Biosystems) using a 7500 real-time PCR system (Applied Biosystems). The expression of target genes was normalized to the expression of β-actin, and the ΔΔCt method was used for gene expression determination. Expressions of target genes are reported relative to endogenous beta-actin CTL.
Table 1 Primers used for PCR amplification
Quantification of advanced glycation end products (AGE)
Fluorescent AGEs were determined at an emission and excitation wavelength of 440 and 370 nm, respectively on a SpectraMax M5 ELISA Microplate Reader (Molecular Devices, CA, USA).Citation32 AGE-specific fluorescence of the samples was measured at a protein concentration of 1 mg/mL and are expressed in AU compared with a native BSA solution (1 mg/mL, 0.1 N NaOH).
Statistical analysis
Results are expressed as the mean ± standard deviation (SD) for each group. Statistical analyses were performed with the Prism package. Student’s t-test was used for comparison of data of two groups. One-way ANOVA with Bonferroni’s post hoc test were used to determine the differences among three or more groups. Differences were considered significant at P<0.05.
Results
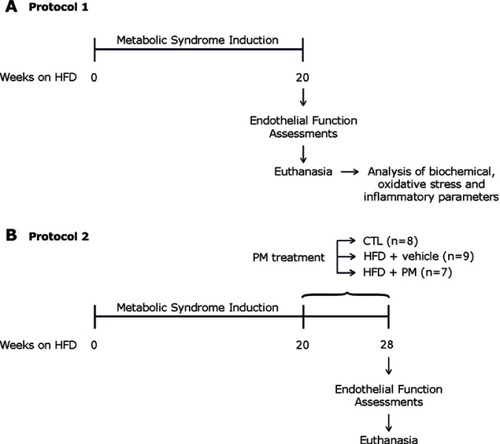
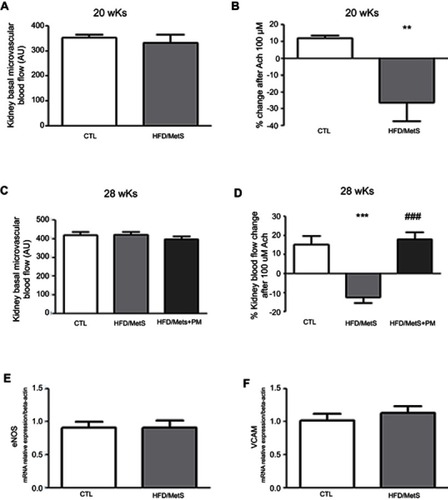
Metabolic, hemodynamic, and biochemical characterization of HFD-induced MetS rats were previously published in Pereira et al.Citation20 Presently, we observed that HFD induced small but statistically significant alteration in renal function, as shown by increased levels of serum creatinine and decreased urine volume ( and , respectively). The HFD-fed rats had higher serum uric acid levels than the CTL group (). However, biochemical analyses of 24-hr-urine did not show evidence of microalbuminuria (data not shown).
Figure 2 Renal function parameters in control (CTL) and HFD-fed rats (HFD/MetS) for 20 weeks. Serum creatinine (A), urinary volume (B), and serum uric acid (C). *P<0.05 vs control.
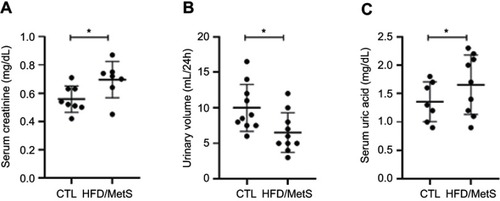
Evaluation of oxidative stress parameters showed that HFD-fed rats presented decreased catalase mRNA transcripts and catalase enzyme activity in comparison to CTL animals ( and , respectively). Analysis of NADPH gene expression or MDA content in kidneys revealed no differences between groups ( and , respectively).
Figure 3 Oxidative stress parameters in the kidney of control (CTL) and HFD-fed rats (HFD/MetS) for 20 weeks. Real-time PCR analyses of mRNA transcript levels of genes coding for catalase (A) and NADPH oxidase (B). Levels of malondialdehyde (MDA) indicating lipid peroxidation assessed by thiobarbituric acid reactive species (TBARs) (C) and catalase enzyme activity (D). **P<0.01 vs control, ***P<0.001 vs control. For this analysis, at least six animals of each group were analyzed out of three independent experiments performed.
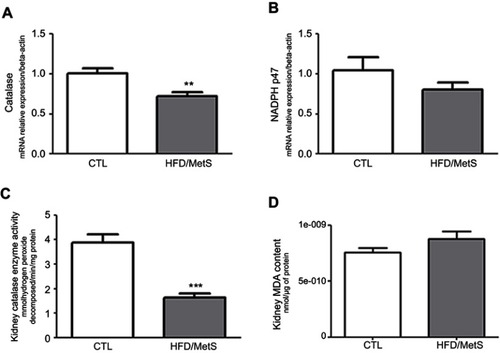
As shown in , kidneys of HFD-fed rats exhibited inflammation, as demonstrated by markedly increased IL-1β. No significant changes were observed between groups in TNF-α cytokine levels and IL-6 mRNA transcripts in kidneys ( and , respectively).
Figure 4 Inflammatory parameters assessed in the kidney of control group (CTL) and HFD-fed rats (HFD/MetS) for 20 weeks. Kidney levels of IL-1β and TNF-α (A and B, respectively) assessed by ELISA quantification and mRNA transcript levels of gene coding for IL-6 evaluated by RT-PCR (C). *P<0.05 vs control.
In order to evaluate if kidneys of HFD-induced MetS rats presented endothelial dysfunction, the basal and % of change in kidney microvascular flux after Ach administration were compared between groups (). As shown in and , kidneys of HFD-fed or HFD/MetS PM-treated animals exhibited no significant differences in basal microvascular blood flow, irrespective of HFD duration. HFD-fed rats exhibited kidney endothelial dysfunction, as demonstrated by paradoxical vasoconstriction in response to Ach, while kidneys of control animals exhibited vasorelaxation at the same dose of Ach ( and , 20 and 28 weeks of HFD, respectively). Importantly, kidney vasorelaxation was marked and significantly improved when rats were treated with PM ( and , 20 and 28 weeks of HFD, respectively). Both eNOS and vascular cell adhesion molecule (VCAM) gene expression were not altered in the kidneys of HFD-induced MetS rats in comparison to CTLs ( and , respectively).
Figure 5 Kidney endothelial function of control group (CTL), HFD-induced MetS rats (HFD/MetS) and MetS rats treated with 60 mg/kg/day pyridoxamine (PM) during the last 8 weeks of HFD feeding. Laser speckle contrast imaging (LSCI) assessment of basal microvascular flux (A and C, 20 and 28 weeks of HFD, respectively) and percentage (%) of change in kidney microvascular flux after topic acetylcholine (Ach) administration (B and D, 20 and 28 weeks of HFD, respectively). Real-time PCR analyses of mRNA transcript levels of genes coding for eNOS (E) and VCAM (F). **P<0.001, ***P<0.001 vs control and P<0.001 vs HFD/MetS.
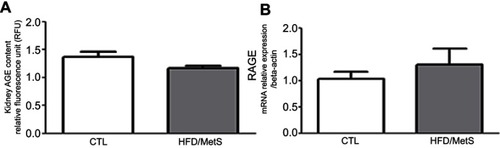
The participation of AGE-RAGE axis was evaluated in kidneys of HFD-induced MetS rats, and we have observed that there was no difference in the kidney AGE content and RAGE mRNA expression between the groups ( and , respectively).
Figure 6 AGE deposition in the kidney assessed by fluorescent spectroscopy (A) and RAGE mRNA levels evaluated by RT-PCR in control (CTL) and HFD-induced MetS rats (HFD/MetS) for 20 weeks.
Discussion
We have previously shown that Wistar rats fed a HFD for 20 weeks presented features of MetS, liver steatosis, and liver endothelial dysfunction.Citation20 At present, we observed that HFD feeding induced paradoxical vasoconstriction in renal arteries at early-stages of renal disease, which to the best of our knowledge have not been previously explored. Paradoxical vasoconstriction, i.e., Ach mediated vasoconstriction, is a feature of patients with nonobstructive coronary atherosclerosis. It is believed that in atherosclerosis the lack of NO bioavailability triggers smooth muscle cell constriction leading to vasospasm in nonobstructive coronary arteries, which leads to myocardial infarction.Citation33–Citation35 Classically, neutrophils and monocytes infiltrations, pro-inflammatory mediators as well as elevated oxidative stress are implicated the physiopathology of vascular damage,Citation36 nonetheless, these cascades are still not proven participants of the paradoxical vasoconstriction phenomenon.
The increase in abdominal fat deposition, insulin resistance, and a pro-inflammatory and pro-oxidative environmental are well-accepted participants of the pathogenesis of endothelial dysfunction and contributes to the worsening of cardio-renal diseases.Citation37 We demonstrated that HFD-induced MetS animals presented increased fat content and insulin resistance,Citation20 as well as decreased antioxidant defense, evidenced by decrease in kidney catalase mRNA transcripts and activity, and increased pro-inflammatory cytokine IL-1β. The increase in the inflammatory mediator IL-1β in the kidneys is an indicator of tissue injury and danger signaling which triggers local and systemic inflammation through the activation of inflammasome.Citation38 IL-1β has been implicated in the stimulation of leukocyte recruitment by activation of adhesion molecules, including intercellular adhesion molecule -1 and VCAM-1, which are implicated in the etiology of microcirculatory disturbances, finally leading to endothelial dysfunction.Citation38
Interestingly, despite having negligible changes in renal function, animals with MetS showed pronounced kidney microvascular endothelial dysfunction. Studies have shown that renal lesion (i.e., glomerulosclerosis) is present in obese patients with normal renal function evaluated by serum levels of plasma proteins, serum creatinine, and 24-hr proteinuria.Citation39,Citation40 Previously Pasarin et al, also showed the occurrence of endothelial dysfunction before the appearance of inflammation and fibrosis.Citation41 In streptozotocin-induced diabetic rats, Maric-Bilkan and colleges showed that microvascular changes preceded decline in renal function.Citation42 Therefore, endothelial dysfunction could be an early-stage manifestation of MetS and related diseases and could have a causal role in the development of organ failure and/or CKD.Citation43
Increases in AGE levels were not observed in the kidney of 20 weeks HFD-induced MetS rats. Since we have previously shown that serum and liver AGEs levels are increased in animals with MetSCitation20 it is plausible to think that macrophages and kupffer cells from the liver are actively participating in AGEs detoxification, generating low molecular weight soluble peptides which are further excreted by the kidneys.Citation28 Over time, the loss of capacity of AGE detoxification and increased production of different types of AGE lead to a self-perpetuating cycle of AGE formation and oxidative stress which progressively increase renal injury.Citation44
PM has been recently proposed as a supplementary approach in diabetic nephropathy, due to its effect on reducing serum creatinine and/or urinary albumin/creatinine ratio in initial diabetic nephropathy.Citation12,Citation14,Citation15 The therapeutic effects of PM to revert the vascular complication in MetS presently shown do not seem to depend on AGE-RAGE modulation and could be due to its antioxidant and metabolic effects.Citation15 Jain and Lim reported that PM inhibits protein glycation, superoxide production, and lipid peroxidation.Citation45 Improvement of body weight gain and glucose intolerance in HFD C57BL6/J or diabetic KK-Ay/Ta mice was observed after PM treatment.Citation46,Citation47 Indeed, metabolic improvements associated with PM treatment in HFD-induced MetS rats were evidenced by our group including body weight (HFD 569.8 g±75.5 vs HFD + PM 492.7 g±31.6, P≤0.01), fasting blood glucose (HFD 5.1 mmol/L±0.6 vs HFD + PM 4.2 mmol/L±0.4, P≤0.01), and visceral fat recovery (HFD 18.8 g±5.1 vs HFD + PM 13.4±1.5, P≤0.05)(submitted paper). Mechanistically, the protective effect of PM could be due to increased expression of kidney and adipose tissue CD36, a receptor/transporter of fatty acids.Citation47
Our findings suggested that HFD-induced kidney microvascular dysfunction is an early manifestation of MetS and is related to antioxidant machinery defects and inflammation activation. Additionally, PM is a promissory agent in the management of MetS related kidney endothelial dysfunction, probably due to its metabolic and antioxidant effects. Our results reinforce the view that therapeutic strategies aimed at minimizing HFD-induced renal damage are especially necessary for MetS patients.
Disclosure
The authors report no conflicts of interest in this work.
References
- Grundy SM , Cleeman JI , Daniels SR , et al. Diagnosis and management of the metabolic syndrome: an American Heart Association/National Heart, Lung, and Blood Institute Scientific Statement. Circulation . 2005;112(17):2735–2752. doi:10.1161/CIRCULATIONAHA.105.169404 16157765
- Grundy SM . Metabolic syndrome update. Trends Cardiovasc Med . 2016;26(4):364–373. doi:10.1016/j.tcm.2015.10.004 26654259
- Saklayen MG . The global epidemic of the metabolic syndrome. Curr Hypertens Rep . 2018;20(2):12. doi:10.1007/s11906-018-0812-z 29480368
- Engin A . The definition and prevalence of obesity and metabolic syndrome. Adv Exp Med Biol . 2017;960:1–17. doi:10.1007/978-3-319-48382-5_1 28585193
- Nashar K , Egan BM . Relationship between chronic kidney disease and metabolic syndrome: current perspectives. Diabetes Metab Syndr Obes . 2014;7:421–435. doi:10.2147/DMSO.S45183 25258547
- Kurella M , Lo JC , Chertow GM . Metabolic syndrome and the risk for chronic kidney disease among nondiabetic adults. J Am Soc Nephrol . 2005;16(7):2134–2140. doi:10.1681/ASN.2005010106 15901764
- Guo Y , Cui L , Ye P , Li J , Wu S , Luo Y . Change of kidney function is associated with all-cause mortality and cardiovascular diseases: results from the Kailuan study. J Am Heart Assoc . 2018;7(21):e010596. doi:10.1161/JAHA.118.008528 30608199
- Zhang X , Lerman LO . The metabolic syndrome and chronic kidney disease. Transl Res . 2017;183:14–25. doi:10.1016/j.trsl.2016.12.004 28025032
- Ritz E , Orth SR . Nephropathy in patients with type 2 diabetes mellitus. N Engl J Med . 1999;341(15):1127–1133. doi:10.1056/NEJM199910073411506 10511612
- Fernandez-Fernandez B , Ortiz A , Gomez-Guerrero C , Egido J . Therapeutic approaches to diabetic nephropathy–beyond the RAS. Nat Rev Nephrol . 2014;10(6):325–346. doi:10.1038/nrneph.2014.74 24802062
- Williams ME , Bolton WK , Khalifah RG , Degenhardt TP , Schotzinger RJ , McGill JB . Effects of pyridoxamine in combined phase 2 studies of patients with type 1 and type 2 diabetes and overt nephropathy. Am J Nephrol . 2007;27(6):605–614. doi:10.1159/000108104 17823506
- Dwyer JP , Greco BA , Umanath K , et al. Pyridoxamine dihydrochloride in diabetic nephropathy (PIONEER-CSG-17): lessons learned from a pilot study. Nephron . 2015;129(1):22–28. doi:10.1159/000369310 25532068
- Tanimoto M , Gohda T , Kaneko S , et al. Effect of pyridoxamine (K-163), an inhibitor of advanced glycation end products, on type 2 diabetic nephropathy in KK-A(y)/Ta mice. Metabolism . 2007;56(2):160–167. doi:10.1016/j.metabol.2006.08.026 17224327
- Degenhardt TP , Alderson NL , Arrington DD , et al. Pyridoxamine inhibits early renal disease and dyslipidemia in the streptozotocin-diabetic rat. Kidney Int . 2002;61(3):939–950. doi:10.1046/j.1523-1755.2002.00207.x 11849448
- Chiazza F , Cento AS , Collotta D , et al. Protective effects of pyridoxamine supplementation in the early stages of diet-induced kidney dysfunction. Biomed Res Int . 2017;2017:2682861. doi:10.1155/2017/2682861 29214163
- Voziyan PA , Hudson BG . Pyridoxamine as a multifunctional pharmaceutical: targeting pathogenic glycation and oxidative damage. Cell Mol Life Sci . 2005;62(15):1671–1681. doi:10.1007/s00018-005-5082-7 15905958
- Metz TO , Alderson NL , Thorpe SR , Baynes JW . Pyridoxamine, an inhibitor of advanced glycation and lipoxidation reactions: a novel therapy for treatment of diabetic complications. Arch Biochem Biophys . 2003;419(1):41–49. doi:10.1016/j.abb.2003.08.021 14568007
- Metz TO , Alderson NL , Chachich ME , Thorpe SR , Baynes JW . Pyridoxamine traps intermediates in lipid peroxidation reactions in vivo: evidence on the role of lipids in chemical modification of protein and development of diabetic complications. J Biol Chem . 2003;278(43):42012–42019. doi:10.1074/jbc.M304292200 12923193
- Brings S , Fleming T , Freichel M , Muckenthaler MU , Herzig S , Nawroth PP . Dicarbonyls and advanced glycation end-products in the development of diabetic complications and targets for intervention. Int J Mol Sci . 2017;18(5). doi:10.3390/ijms18050984
- Pereira E , Silvares RR , Flores EEI , et al. Hepatic microvascular dysfunction and increased advanced glycation end products are components of non-alcoholic fatty liver disease. PLoS One . 2017;12(6):e0179654. doi:10.1371/journal.pone.0179654 28628674
- Taniguchi N , Takahashi M , Kizuka Y , et al. Glycation vs. glycosylation: a tale of two different chemistries and biology in Alzheimer’s disease. Glycoconj J . 2016;33(4):487–497. doi:10.1007/s10719-016-9690-2 27325408
- Rabbani N , Thornalley PJ . Advanced glycation end products in the pathogenesis of chronic kidney disease. Kidney Int . 2018;93(4):803–813. doi:10.1016/j.kint.2017.11.034 29477239
- Maessen DE , Brouwers O , Gaens KH , et al. Delayed intervention with pyridoxamine improves metabolic function and prevents adipose tissue inflammation and insulin resistance in high-fat diet-induced obese mice. Diabetes . 2016;65(4):956–966. doi:10.2337/db15-1390 26718500
- Thomas G , Sehgal AR , Kashyap SR , Srinivas TR , Kirwan JP , Navaneethan SD . Metabolic syndrome and kidney disease: a systematic review and meta-analysis. Clin J Am Soc Nephrol . 2011;6(10):2364–2373. doi:10.2215/CJN.02180311 21852664
- Wisse BE . The inflammatory syndrome: the role of adipose tissue cytokines in metabolic disorders linked to obesity. J Am Soc Nephrol . 2004;15(11):2792–2800. doi:10.1097/01.ASN.0000141966.69934.21 15504932
- Dandona P , Aljada A , Chaudhuri A , Mohanty P , Garg R . Metabolic syndrome: a comprehensive perspective based on interactions between obesity, diabetes, and inflammation. Circulation . 2005;111(11):1448–1454. doi:10.1161/01.CIR.0000158483.13093.9D 15781756
- Prasad GV . Metabolic syndrome and chronic kidney disease: current status and future directions. World J Nephrol . 2014;3(4):210–219. doi:10.5527/wjn.v3.i4.210 25374814
- Stirban A , Gawlowski T , Roden M . Vascular effects of advanced glycation endproducts: clinical effects and molecular mechanisms. Mol Metab . 2014;3(2):94–108. doi:10.1016/j.molmet.2013.11.006 24634815
- Cao Z , Cooper ME . Role of angiotensin II in tubulointerstitial injury. Semin Nephrol . 2001;21(6):554–562.11709803
- Wang H , Zhang H , Sun L , Guo W . Roles of hyperuricemia in metabolic syndrome and cardiac-kidney-vascular system diseases. Am J Transl Res . 2018;10(9):2749–2763.30323864
- Wu Y , Ren J , Zhou B , et al. Laser speckle contrast imaging for measurement of hepatic microcirculation during the sepsis: a novel tool for early detection of microcirculation dysfunction. Microvasc Res . 2015;97:137–146. doi:10.1016/j.mvr.2014.10.006 25446370
- Nakayama H , Mitsuhashi T , Kuwajima S , et al. Immunochemical detection of advanced glycation end products in lens crystallins from streptozocin-induced diabetic rat. Diabetes . 1993;42(2):345–350. doi:10.2337/diab.42.2.345 8425672
- Ludmer PL , Selwyn AP , Shook TL , et al. Paradoxical vasoconstriction induced by acetylcholine in atherosclerotic coronary arteries. N Engl J Med . 1986;315(17):1046–1051. doi:10.1056/NEJM198610233151702 3093861
- Niccoli G , Scalone G , Crea F . Acute myocardial infarction with no obstructive coronary atherosclerosis: mechanisms and management. Eur Heart J . 2015;36(8):475–481. doi:10.1093/eurheartj/ehu469 25526726
- Nguyen MT , Pham I , Valensi P , et al. Flow-mediated-paradoxical vasoconstriction is independently associated with asymptomatic myocardial ischemia and coronary artery disease in type 2 diabetic patients. Cardiovasc Diabetol . 2014;13:20. doi:10.1186/1475-2840-13-80 24428877
- Sun CK , Shao PL , Wang CJ , Yip HK . Study of vascular injuries using endothelial denudation model and the therapeutic application of shock wave: a review. Am J Transl Res . 2011;3(3):259–268.21633631
- Engin A . Endothelial dysfunction in obesity. Adv Exp Med Biol . 2017;960:345–379. doi:10.1007/978-3-319-48382-5_15 28585207
- Libby P . Interleukin-1 beta as a target for atherosclerosis therapy: biological basis of CANTOS and beyond. J Am Coll Cardiol . 2017;70(18):2278–2289. doi:10.1016/j.jacc.2017.09.028 29073957
- Serra A , Romero R , Lopez D , et al. Renal injury in the extremely obese patients with normal renal function. Kidney Int . 2008;73(8):947–955. doi:10.1038/sj.ki.5002796 18216780
- Chang Y , Ryu S , Cho J , Pastor-Barriuso R , Guallar E . Metabolically healthy obesity and development of chronic kidney disease. Ann Intern Med . 2016;165(10):744–745. doi:10.7326/L16-0405 27842405
- Pasarin M , La Mura V , Gracia-Sancho J , et al. Sinusoidal endothelial dysfunction precedes inflammation and fibrosis in a model of NAFLD. PLoS One . 2012;7(4):e32785. doi:10.1371/journal.pone.0032785 22509248
- Maric-Bilkan C , Flynn ER , Chade AR . Microvascular disease precedes the decline in renal function in the streptozotocin-induced diabetic rat. Am J Physiol Renal Physiol . 2012;302(3):F308–F315. doi:10.1152/ajprenal.00421.2011 22031855
- Chade AR . Small vessels, big role: renal microcirculation and progression of renal injury. Hypertension . 2017;69(4):551–563. doi:10.1161/HYPERTENSIONAHA.116.08319 28193706
- Sanajou D , Ghorbani Haghjo A , Argani H , Aslani S . AGE-RAGE axis blockade in diabetic nephropathy: current status and future directions. Eur J Pharmacol . 2018;833:158–164. doi:10.1016/j.ejphar.2018.06.001 29883668
- Jain SK , Lim G . Pyridoxine and pyridoxamine inhibits superoxide radicals and prevents lipid peroxidation, protein glycosylation, and (Na+ + K+)-ATPase activity reduction in high glucose-treated human erythrocytes. Free Radic Biol Med . 2001;30(3):232–237.11165869
- Hagiwara S , Gohda T , Tanimoto M , et al. Effects of pyridoxamine (K-163) on glucose intolerance and obesity in high-fat diet C57BL/6J mice. Metabolism . 2009;58(7):934–945. doi:10.1016/j.metabol.2009.02.033 19427656
- Murakoshi M , Tanimoto M , Gohda T , et al. Pleiotropic effect of pyridoxamine on diabetic complications via CD36 expression in KK-Ay/Ta mice. Diabetes Res Clin Pract . 2009;83(2):183–189. doi:10.1016/j.diabres.2008.11.008 19144436