
Abstract
Background
Interactions between environmental factors, such as diet and lifestyle, and metabolic pathways are pivotal in understanding aging mechanisms. hNAA40, Nicotinamide phosphoribosyltransferase (NAMPT), and NAD-dependent protein deacetylase sirtuin-1 (SIRT-1) have been shown to exert important biological processes, including stress response and aging.
Methods
hNAA40, NAMPT, and SIRT-1 mRNA expression in peripheral blood mononuclear cells (PBMC) were quantitated in 30 lean adult volunteers of normal weight, 30 obese, 20 drug-naïve obese Type 2 diabetes mellitus (T2DM), and 30 obese T2DM on Metformin. Similarly, hNAA40, NAMPT, and SIRT-1 expression in PBMC were quantitated in 36 normal healthy adults randomly assigned to three different groups (Glucose or Whey proteins or lipids; 300 kcal). Blood samples were obtained at 1, 2, and 3 hrs after the macronutrient intake.
Results
There was an increase in hNAA40 and a decrease in NAMPT and SIRT-1 expression in PBMC from T2DM. Metformin treatment reverted hNAA40, NAMPT, and SIRT-1 expression levels to normal levels. Glucose intake resulted in a significant increase in expression of hNAA40 at 1 hr and decreased significantly at 3 hrs post intake. Lipid intake resulted in an increase in expression of hNAA40 at 2 hr post intake and returned to normal levels at 3 hrs. Neither glucose nor lipid intake resulted in a significant change in NAMPT or SIRT-1 expression. Whey proteins resulted in significantly lower expression of NAMPT at 3 hrs and did not alter the expression levels of SIRT-1 significantly.
Conclusion
hNAA40, NAMPT, and SIRT-1 pathway could play a role in the determination of the healthy life-span. Metformin modulates this pathway.
Introduction
Uncovering interactions between environmental factors, such as diet and lifestyle, and metabolic pathways is crucial to elucidate aging mechanisms. Some of the most promising candidate genes appear to be involved in regulatory pathways such as stress resistance, immune/inflammatory response, insulin signalling, or cardiovascular function.Citation1 The survival and maintenance of the internal environment in many cells depend largely on their ability to respond to cellular stressors. In yeast, it has been demonstrated that Nat4 (the yeast ortholog of hNAA40) levels are mediated and controlled by caloric restriction (CR) and that one of the main effects of decreasing Nat4 levels is increasing the lifespan of yeast cells through a pathway that upregulates the host’s stress-response genes, PNC1 (the yeast ortholog of nicotinamide phosphoribosyltransferase (NAMPT)), Glycogen phosphorylase (GPH1), Hexokinase (HXK), 1,4-alpha-glucan-branching enzyme (GLC3), Trehalose-phosphatase (TPS2), Glycogen [starch] synthase isoform 1 (GSY1), and Neutral trehalase (NTH1).Citation2 Of these genes, PNC1 is known to encode a nicotinamidase that lowers the accumulation of nicotinamide (Nam) intracellularly which would result in stimulating the activity of Sir2 (the yeast ortholog of Sirtuins)Citation3 and thus enhancing the cell’s response to stressors. NAMPT is the rate-limiting enzyme in mammalian NAD synthesis, catalyzing nicotinamide mononucleotide (NMN) formation from Nam and 5-phosphoribosyl 1-pyrophosphate (PRPP). Thus, NAMPT is thought to be involved in many important biological processes, including metabolism, stress response, and aging.
It can be assumed that hNAA40 (human ortholog of Nat4) is similarly controlled and has similar effects in humans. Accordingly, high caloric intake, the abundance of macronutrients in the body, and high amounts of body fat could increase the levels of hNAA40 resulting in the reduction of stress-response proteins, NAMPT and Sirtuins. This would result in reduced cell’s response to stressors and decreased life span. Increasing cellular lifespan can be helpful in age-related diseases such as Alzheimer’s or cancer treatment where increasing longevity and activity of immune cells may boost their efficacy in killing tumor cells.Citation4 Histone modification changes represent an attractive model through which several environmental signals, such as diet, could play a pivotal role in regulating cell lifespan.Citation3 Several reports show that diet manipulation, such as CR, extends the cellular lifespan and hinders age-related chronic diseases of many species, such as rats, fish, mites, and yeast.Citation5 CR is also known to regulate insulin secretion and prevents insulin resistance. In contrast, obesity, a state of overeating, is usually associated with inappropriate secretion of insulin and is considered as a risk factor for T2DM, which results in hyperglycemia, insulin resistance, and hyperinsulinemia.Citation6,Citation7 Therefore, CR is considered the most conserved and well‐studied extracellular intervention that prolongs lifespan.Citation8,Citation9
Histone modification changes represent an attractive target of therapeutic intervention in relation to aging. The use of PBMC may provide a useful tool to study human aging as it has been accepted to represent changes in the whole bodyCitation10 and thus has been suggested to represent the biological processes occurring in the body relevant to aging and longevity. Therefore, this study was conducted to determine the differential expression of hNaa40 and its associated stress-response genes, NAMPT and SIRT-1, in PBMC isolated from lean, obese, and drug-Naïve obese T2DM humans. We also examined their levels in T2DM on Metformin which has anti-aging properties and has been shown to be a direct SIRT1-activating compound.Citation11 In addition, the association between the source of substrates for energy production and the expression levels of hNAA40, NAMPT, and SIRT-1 in PBMC was examined following a caloric intake of 300 kcalories of glucose or Whey proteins or lipids.
Materials and Methods
Subjects
Lean/Obese/T2DM/T2DM+Metformin Subjects
Fasting blood samples were obtained from 30 lean adult volunteers of normal weight, 30 obese adult volunteers, 20 drug-naïve obese T2DM newly diagnosed volunteers, and 30 obese T2DM on Metformin volunteers were recruited for the study (). The majority of T2DM on Metformin subjects were on Metformin for at least 9 months and up to 15 years. Most of the subjects on Metformin regiments comprised dosages which varied from 1000 to 2000 mg, 5 had dosages ranging from 100 to 500 daily, and 3 had dosages ranging from 3000 to 4500 mg daily. Five T2DM on Metformin subjects were on insulin therapy as well. The study has been approved by the Institutional Review Board (IRB; approval no. SP17/380/R) of King Abdullah International Medical Research Center (KAIMRC) and all subjects gave their written informed consent.
Table 1 Demographic Data of the Subjected Participated in the Study. Results are Presented as Mean ± S.E.M
Caloric (300 Kcal) Intake Study
Thirty-six normal healthy adult volunteers were recruited into the study. All were normotensive, had a normal lipid profile, normal renal and liver function tests, and were not on any medications. Subjects were randomly assigned to three different groups; each received one type of macronutrient (Glucose or Whey proteins or lipids). Following an overnight fast, a baseline blood sample was taken. Subjects were then given either 300 kcalories of glucose (NERL Trutol 75) or lipids (90 grams whipping cream, 31.5 grams fat, 1.7 grams protein, and 2.25 grams carbohydrate) or protein (Isopure unflavored Whey proteins isolate (The Isopure Company, California, USA) powder containing 26 grams per serving of 100% WPI, stripped of fat, carbs, fillers, sugars, and lactose) solution over 5 mins. Cream and protein preparations were diluted with water up to 300 mL solutions. Blood samples were then obtained at 1, 2, and 3 hrs after the macronutrient intake. Subjects, either 1 week before or after the macronutrient challenge, were given 300 mL of water to drink in the fasting state. Blood samples were obtained before and at 1, 2, and 3 hrs after water intake as well. Each subject served as his/her own control. The study was approved by the IRB of King Abdulaziz Medical City. Subjects gave their written informed consent.
Isolation of PBMC
Blood was collected in EDTA containing vacutainers. Blood was carefully layered over 15 mL of Ficol-Hypaque (50 mL Leucosep Tubes, Greiner Bio-One North America Inc, North Carolina, USA) following dilution with an equal volume of Phosphate Buffered Saline (PBS). Tubes were then centrifuged at 450 xg, in a swing out rotor for 30 mins at 22°C and the PBMC layer was harvested with a pipette. PBMC were repeatedly washed with PBS. Fifty microliters of Qiagen RNALater were added to the pellet and samples were then frozen at −80ºC.
Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR) Analysis
Total RNA was isolated using the Ambion Aqueous kit (Ambion). All isolated RNA samples were treated with DNase I to remove contaminating genomic DNA. The quality and quantity of the isolated RNA were determined using Agilent Bioanalyzer 2100. One μg of total RNA was reverse-transcribed using first-strand cDNA synthesis Kit (Millipore, USA) followed by RT-qPCR. RT-qPCR was performed with a 7900HT Fast Real-Time PCR System (Applied Biosystems, USA), using 2 µL cDNA, 10 µL 2X Sybergreen Master mix (150 mM Tris, pH 9.2, 40 mM (NH4)2SO4, 5 mM MgCl2, 0.02% Tween-20, 0.4 mM dNTPs, 1.25 Units Taq Polymerase, 1X Sybergreen) and 0.5 µL of 20 μM gene-specific primers (). All primers were purchased from Bio-Basic Canada Inc. (Ontario, Canada). The specificity and size of the PCR products were tested by adding a melt curve at the end of the amplifications, analysis on a 2% agarose gel of the bands. Amplicon Bands were isolated and sequenced. The reaction protocol consisted of one activation cycle of 50°C for 2 mins followed by 95°C for 15 s. Thereafter, 40 cycles of denaturation at 95°C for 15 s, and at 60°C annealing/extension for 2 mins were performed. Although normalization to Cyclophilin A, RPL13, and GAPDH showed similar trends, all values were normalized to Cyclophilin A. Results were calculated using Q-Gene Core module 1.2.Citation12
Table 2 Primer Sequences for All Primers Used in RT-qPCR
Statistical Analysis
Statistical analysis was carried out using SigmaStat software ver. 3.0 (Jandel Scientific, San Rafael, CA). Normalized mRNA expression was calculated for RT-qPCR results and analysis was carried out using one-way ANOVA on Ranks followed by Dunn’s test for pairwise comparisons, and comparison against the lean group was used to compare data for lean, obese, T2DM, and T2DM with Metformin study. One-way repeated measures ANOVA on Ranks test was used for caloric intake challenge study. Two-factor ANOVA was used to evaluate the interaction between treatment (water vs caloric intake) and time. P-value < 0.05 was used to assess significance for all statistical analyses. Results are presented as mean ± S.E.M.
Results
Demographic Data of Subjects
There were significant differences in the age among the four groups included in the study (). However, there was no significant correlation between the expression levels of hNAA40, nor NAMPT nor SIRT-1 and age. On the other hand, there were no significant differences in age, BMI, glucose, lipid profile, HbA1c, and insulin between T2DM and T2DM on Metformin ().
mRNA Expression of hNAA40, NAMPT, and SIRT-1 in PBMC of Lean, Obese, T2DM, and T2DM on Metformin
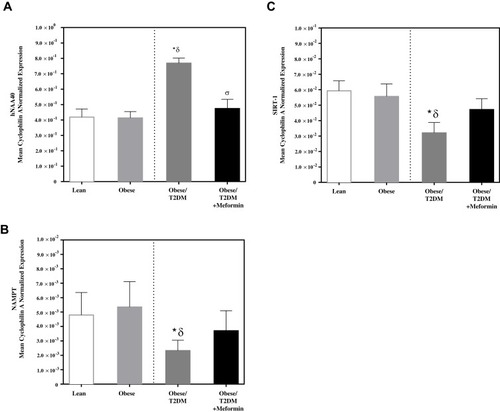
There was a significant increase in mRNA expression of hNAA40 in T2DM when compared to lean and obese subjects. T2DM on Metformin subjects had lower mRNA expression of hNAA40 PBMC when compared to T2DM (, P<0.05). Similarly, NAMPT expression was significantly lower in PBMC of T2DM (, P<0.05) when compared to normal lean and obese subjects and Metformin treatment reverted NAMPT expression levels to normal levels. SIRT-1 expression in PBMC of T2DM was also significantly inhibited (, P<0.05) when compared to lean and obese subjected. T2DM on Metformin subjects had similar mRNA expression of SIRT-1 in PBMC of lean and obese subjects.
Figure 1 mRNA expression in PBMC of lean, obese, T2DM, and T2DM on Metformin of (A) hNAA40; (B) NAMPT; and (C) SIRT-1. Results are presented as Mean ± S.E.M. *P<0.05 vs lean subjects; δP<0.05 vs obese; σP<0.05 vs T2DM.
mRNA Expression of hNAA40, NAMPT, and SIRT-1 in PBMC Following Macronutrient Intake
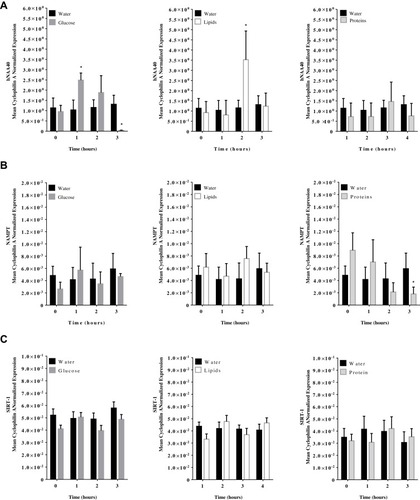
Caloric intake (300 kcal) of either glucose or proteins or lipids in normal volunteers changed insulin and blood glucose significantly when compared to water intake (). Whey protein intake increased insulin concentrations at 1–3 hrs while glucose intake increased insulin concentrations at 1 and 2 hrs (*P<0.05; ). Lipid intake did not change insulin concentrations significantly. On the other hand, Whey protein intake decreased blood glucose significantly at 1, 2, and 3 hrs when compared to water intake while glucose concentrations decreased at 2 and 3 hrs following glucose or lipid intake (*P<0.05; ). Glucose intake resulted in a significant increase in mRNA expression of hNAA40 in PBMC at 1 hr and decreased significantly at 3 hrs post intake (*P<0.05; ). Lipid intake resulted in an increase in mRNA expression of hNAA40 in PBMC at 2 hr post intake when compared to water intake (*P<0.05; ) and returned to normal levels at 3 hrs. Neither glucose nor lipids intake resulted in a significant change in NAMPT or SIRT-1 expression in PBMC ( and C). Whey proteins, on the other hand, resulted in significantly lower mRNA expression of NAMPT in PBMC at 3 hrs post protein intake when compared to the baseline expression levels or water intake (). Whey protein intake did not alter expression levels of SIRT-1 in PBMC significantly.
Table 3 Demographic Data of the Subjects Participated in the Study. Results are Presented as Mean ± S.E.M
Figure 2 hNAA40, NAMPT, and SIRT-1 mRNA expression levels in PBMC following (A) glucose; (B), Whey proteins; and (C) lipids at 0, 1, 2, and 3 hrs post-caloric challenge. Results are presented as Mean ± S.E.M. *P<0.05 vs water intake; n=12 per group.
Discussion
The use of PBMC as a tool to study human aging has been proposed to be a useful tool to represent changes in the whole body.Citation10 Increased age-associated changes in gene expression of PBMC have been reported, including increased capacity for apoptosis (Bcl-2 homologous antagonist/killer; BAK1), cell cycle regulation (Cyclin-dependent kinase inhibitor 1B; CDKN1B), metabolic process (Alpha-2-macroglobulin receptor-associated protein; LRPAP1), insulin action (Cation-independent mannose-6-phosphate receptor; IGF2R), and increased immune and inflammatory response (Interleukin-27 receptor subunit alpha; IL27RA). On the other hand, response to stress (Heat shock cognate 71 kDa protein; HSPA8), damage stimulus (X-ray repair cross-complementing protein 6; XRCC6), and chromatin remodeling (TERF1-interacting nuclear factor 2; TINF2) pathways have been shown to be downregulated in PBMC of old people.Citation1 Aging is associated with overactive immune cells that contribute to autoimmune diseases and downregulation of certain metabolic pathways which increase the risk of infection. This process is known as immunosenescence and is related to mitochondrial function and energy balance.Citation13 NAD+ levels markedly decline with age, creating an energy deficit that decreases the body’s ability to retain key functions.Citation14 Intracellular levels of NAD+ regulate immune and inflammatory pathways, including interleukin-6 (IL-6) and tumor necrosis factor-alpha (TNFα).Citation15,Citation16 The rate-limiting step within the NAD+ salvage pathway required for converting nicotinamide to NAD+ is catalyzed by NAMPT which has been recently labeled as a novel mediator of innate immunity. The silencing of NAMPT expression in monocytes decreased activation and infiltration of monocytes/macrophages and neutrophils in arthritic joints. The decreased monocyte activation was associated with reduced IL-6 production, Th17 cells, and autoantibody titers.Citation17 On the other hand, NAMPT upregulation blocked macrophage apoptosis by activating IL-6 and the prosurvival signal transducer and activator of transcription 3 (STAT3).Citation18 However, the ability of NAMPT to trigger IL-6/STAT3 cell survival pathway did not depend on the presence of nicotinamide and was not blocked by the NAMPT enzyme inhibitor FK866.Citation18 Thus, inhibition of NAMPT expression in PBMC and the higher expression levels of IL-6 previously reportedCitation19 in our study support the notion that these pathways are regulated independently.
Endothelial dysfunction is a characteristic of aging‐related vascular disease and is worsened during diabetes. Overexpression of NAMPT increased proliferation and extended replicative lifespan and did so preferentially during glucose overload. Additionally, upregulation of NAMPT in endothelial cells delayed markers of senescence and oxidative stress in high glucose and tube formation were more extensive and glucose-resistant.Citation20 Obesity and T2DM are both associated with insulin resistance and hyperinsulinemia while T2DM is associated with hyperglycemia as well. Since hNAA40, NAMPT, and SIRT-1 expression in PBMC from T2DM were modulated significantly only in T2DM, this suggests that chronic hyperglycemia could be the modulator of these proteins. Although glucose intake induced hNAA40 in PBMC of lean subjects, there was no change in the expression of NAMPT and SIRT-1. Chronic hyperglycemia may be required to modulate NAMPT and SIRT-1. Moreover, the anti-inflammatory effect of insulin,Citation21 stimulated by glucose intake, could inhibit this pathway. Glucose intake in normal subjects induced inflammatory mediators expression in PBMCCitation22 while insulin infusion reduced expression of these inflammatory mediators in PBMC.Citation23 Whey protein intake did not alter the expression levels of hNAA40 as with either glucose or lipid intake. Whey proteins intake induced high levels of insulin secretion that persisted for more than 3 hrs.Citation24 This was associated with a lack of hNAA40 expression level changes and inhibition of NAMPT at 3 hrs. Further studies are needed to examine the insulin modulatory effect of this pathway which may play a role in inflammaging and age-related inflammatory conditions.
Synthesis of NAD+ by NAMPT has been shown to be highly sensitive to nutritional changes in adipose tissue. Mice-fed hyper-calorific diets have decreased NAD+ levels, primarily as a result of a reduction in NAMPT.Citation25,Citation26 In addition, circulating extracellular-secreted form of NAMPT (eNAMPT), also known as pre-B cell colony-enhancing factor or visfatin, has been reported to be associated with several metabolic and inflammatory disorders such as obesity.Citation27 However, the correlation of plasma eNAMPT levels in obesity and other metabolic disorders still unclear. eNAMPT was not lowered following Metformin treatment, low caloric diets or after gastric surgery in obese patients.Citation28 The lack of correlation could be attributed to the lack of eNAMPT plasma assay sensitivity.Citation29 In our study, Metformin lowered cellular NAMPT expression in PBMC in addition to SIRT-1. Metformin has been proposed to operate as an agonist of SIRT-1.Citation11 Through activation of SIRT-1, whole-body insulin resistance is prevented by increased Adiponectin release. SIRT-1 also inhibits the activation of Peroxisome proliferator-activated receptor-γ (PPARγ)-dependent insulin-resistance genes. Whether Metformin exerts its effect on NAMPT-SIRT-1- PPARγ pathway by modulating hNAA40 is a possibility and requires further investigation.
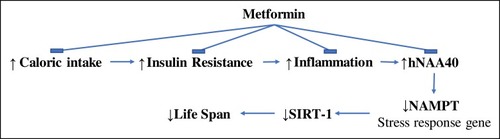
Human primary monocytes comprise a heterogeneous population that can be classified into three subsets based on CD14 and CD16 expression.Citation30,Citation31 A dichotomy has been proposed for macrophage activation: classic (M1) vs alternative (M2). The proinflammatory and phagocytic M1 macrophages are classically activated, typically by IFN-γ or lipopolysaccharide (LPS), whereas M2 macrophages decrease inflammation and encourage tissue repair. The highly pro-inflammatory nature of the non-classical monocytes could be a manifestation of the senescence-associated secretory phenotype (SASP). Interestingly, an accumulation of the non-classical monocytes, in conjunction with higher levels of plasma TNFα and interleukin-8 (IL-8), has been observed in the elderly.Citation32 Our previous study has shown that PBMC from T2DM subjects have increased expression of IL-6, inducible nitric oxide synthase (iNOS), TNFα, CD36, and CD206 and reduced expression of CD11b, CD11c, CD169, and CD163 (19). Other studies have demonstrated an increased inflammatory state in obesity and T2DM.Citation33–Citation35 Similarly, caloric intake has been shown to induce an acute inflammatory response in PBMC of healthy subjectsCitation22,Citation36,Citation37 while insulin infusion reduced inflammation.Citation23,Citation38 These factors may contribute to inflammaging and age-related inflammatory conditions, such as atherosclerosis observed in T2DM. In this study, PBMC present in T2DM has increased hNAA40 and a decreased NAMPT and SIRT-1 expression. These data suggest that PBMC are senescent cells since NAMPT and SIRT-1 expression is inhibited. Interestingly, caloric restriction has been shown to reduce inflammation and oxidative stress and results in prolonged lifespan.Citation5,Citation39–Citation41 Metformin, on the other hand, reduced the expression of hNAA40 and reverted the expression levels of NAMPT and SIRT-1 in PBMC to normal levels. Metformin has been shown previously to reduce caloric intake.Citation42 In addition, Metformin not only improves chronic inflammation through the improvement of metabolic parameters such as hyperglycemia, insulin resistance, and atherogenic dyslipidemia but also has a direct anti-inflammatory action.Citation43–Citation45 These effects could explain the modulatory effect of Metformin on hNAA40/NAMPT/SIRT-1 pathway () but do not exclude a direct effect of Metformin on hNAA40/NAMPT/SIRT-1 pathway. Further studies are needed to explore this effect.
Figure 3 A proposed mechanism by which Metformin could be exerting its anti-aging properties. Insulin resistance and chronic food intake may lead to an increased inflammatory state present in T2DM which results in an increase in hNAA40 and reduction in NAMPT and SIRT-1. Metformin could be exerting its effect by reducing caloric intake, insulin resistance, and inflammation.
Ethical Approval
The protocol of this study was approved by the institutional review board (IRB) for KAIMRC at the Ministry of National Guard Health Affairs (MNG-HA) and all subjects gave their written informed consent. All procedures performed in studies involving human participants were in accordance with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Author Contributions
All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
All authors declare no conflicts of interest in relation to the work reported in this manuscript.
Additional information
Funding
References
- Abdul Rahman A , Abdul Karim N , Abdul Hamid NA , Harun R , Ngah WZ . Senescence-related changes in gene expression of peripheral blood mononuclear cells from octo/nonagenarians compared to their offspring. Oxid Med Cell Longev . 2013;2013:189129. doi:10.1155/2013/189129 24381713
- Yang H , Lavu S , Sinclair DA . Nampt/PBEF/Visfatin: a regulator of mammalian health and longevity? Exp Gerontol . 2006;41(8):718–726. doi:10.1016/j.exger.2006.06.003 16842957
- Molina‐Serrano D , Schiza V , Demosthenous C , et al. Loss of Nat4 and its associated histone H4 N-terminal acetylation mediates calorie restriction-induced longevity. EMBO Rep . 2016;17(12):1829–1843. doi:10.15252/embr.201642540 27799288
- Nayar S , Dasgupta P , Galustian C . Extending the lifespan and efficacies of immune cells used in adoptive transfer for cancer immunotherapies-A review. Oncoimmunology . 2015;4(4):e1002720. doi:10.1080/2162402X.2014.1002720 26155387
- Heilbronn LK , Ravussin E . Calorie restriction and aging: review of the literature and implications for studies in humans. Am J Clin Nutr . 2003;78(3):361–369. doi:10.1093/ajcn/78.3.361 12936916
- Bornfeldt KE , Tabas I . Insulin resistance, hyperglycemia, and atherosclerosis. Cell Metab . 2011;14(5):575–585. doi:10.1016/j.cmet.2011.07.015 22055501
- Madonna R , De Caterina R . Atherogenesis and diabetes: focus on insulin resistance and hyperinsulinemia. Rev Esp Cardiol . 2012;65(4):309–313. doi:10.1016/j.recesp.2011.11.010 22357362
- Mair W , Dillin A . Aging and survival: the genetics of life span extension by dietary restriction. Annu Rev Biochem . 2008;77:727–754. doi:10.1146/annurev.biochem.77.061206.171059 18373439
- Koubova J . How does calorie restriction work? Genes Dev . 2003;17(3):313–321. doi:10.1101/gad.1052903 12569120
- Passtoors WM , Beekman M , Gunn D , et al. Genomic studies in ageing research: the need to integrate genetic and gene expression approaches. J Intern Med . 2008;263(2):153–166. doi:10.1111/jim.2008.263.issue-2 18226093
- Cuyas E , Verdura S , Llorach-Pares L , et al. Metformin is a direct SIRT1-activating compound: computational modeling and experimental validation. Front Endocrinol (Lausanne) . 2018;9:657. doi:10.3389/fendo.2018.00657 30459716
- Muller PY , Janovjak H , Miserez AR , Dobbie Z . Processing of gene expression data generated by quantitative real-time RT-PCR. BioTechniques . 2002;32(6):1372–4, 6, 8–9.
- Rottenberg H , Wu S . Mitochondrial dysfunction in lymphocytes from old mice: enhanced activation of the permeability transition. Biochem Biophys Res Commun . 1997;240(1):68–74. doi:10.1006/bbrc.1997.7605 9367884
- Imai SI , Guarente L . It takes two to tango: NAD(+) and sirtuins in aging/longevity control. NPJ Aging Mech Dis . 2016;2:16017. doi:10.1038/npjamd.2016.17 28721271
- Van Gool F , Galli M , Gueydan C , et al. Intracellular NAD levels regulate tumor necrosis factor protein synthesis in a sirtuin-dependent manner. Nat Med . 2009;15(2):206–210. doi:10.1038/nm.1906 19151729
- Montecucco F , Cea M , Cagnetta A , et al. Nicotinamide phosphoribosyltransferase as a target in inflammation- related disorders. Curr Top Med Chem . 2013;13(23):2930–2938. doi:10.2174/15680266113136660208 24171767
- Presumey J , Courties G , Louis-Plence P , et al. Nicotinamide phosphoribosyltransferase/visfatin expression by inflammatory monocytes mediates arthritis pathogenesis. Ann Rheum Dis . 2013;72(10):1717–1724. doi:10.1136/annrheumdis-2012-202403 23313810
- Li Y , Zhang Y , Dorweiler B , et al. Extracellular Nampt promotes macrophage survival via a nonenzymatic interleukin-6/STAT3 signaling mechanism. J Biol Chem . 2008;283(50):34833–34843. doi:10.1074/jbc.M805866200 18945671
- Al Dubayee MS , Alayed H , Almansour R , et al. Differential expression of human peripheral mononuclear cells phenotype markers in type 2 diabetic patients and type 2 diabetic patients on metformin. Front Endocrinol (Lausanne) . 2018;9:537. doi:10.3389/fendo.2018.00537 30356719
- Borradaile NM , Pickering JG . Nicotinamide phosphoribosyltransferase imparts human endothelial cells with extended replicative lifespan and enhanced angiogenic capacity in a high glucose environment. Aging Cell . 2009;8(2):100–112. doi:10.1111/ace.2009.8.issue-2 19302375
- Dandona P , Aljada A , Dhindsa S , Garg R . Insulin as an anti-inflammatory and antiatherosclerotic hormone. Clin Cornerstone . 2003;Suppl 4:S13–20. doi:10.1016/S1098-3597(03)90062-7
- Aljada A , Ghanim H , Mohanty P , Syed T , Bandyopadhyay A , Dandona P . Glucose intake induces an increase in activator protein 1 and early growth response 1 binding activities, in the expression of tissue factor and matrix metalloproteinase in mononuclear cells, and in plasma tissue factor and matrix metalloproteinase concentrations. Am J Clin Nutr . 2004;80(1):51–57. doi:10.1093/ajcn/80.1.51 15213027
- Dandona P , Aljada A , Mohanty P , et al. Insulin inhibits intranuclear nuclear factor kappaB and stimulates IkappaB in mononuclear cells in obese subjects: evidence for an anti-inflammatory effect? J Clin Endocrinol Metab . 2001;86(7):3257–3265. doi:10.1210/jcem.86.7.7623 11443198
- Alshahrani A , Bin Khunayfir A , Al Rayih M , et al. Phenotypic characterization of human monocytes following macronutrient intake in healthy humans. Front Immunol . 2017;8:1293. doi:10.3389/fimmu.2017.01293 29109719
- Yoshino J , Mills KF , Yoon MJ , Imai S . Nicotinamide mononucleotide, a key NAD(+) intermediate, treats the pathophysiology of diet- and age-induced diabetes in mice. Cell Metab . 2011;14(4):528–536. doi:10.1016/j.cmet.2011.08.014 21982712
- Chalkiadaki A , Guarente L . High-fat diet triggers inflammation-induced cleavage of SIRT1 in adipose tissue to promote metabolic dysfunction. Cell Metab . 2012;16(2):180–188. doi:10.1016/j.cmet.2012.07.003 22883230
- Garten A , Petzold S , Schuster S , Korner A , Kratzsch J , Kiess W . Nampt and its potential role in inflammation and type 2 diabetes. Handb Exp Pharmacol . 2011;203:147–164.
- Stofkova A . Resistin and visfatin: regulators of insulin sensitivity, inflammation and immunity. Endocr Regul . 2010;44(1):25–36. doi:10.4149/endo_2010_01_25 20151765
- Imai S , Kiess W . Therapeutic potential of SIRT1 and NAMPT-mediated NAD biosynthesis in type 2 diabetes. Front Biosci . 2009;14:2983–2995. doi:10.2741/3428
- Mukherjee R , Kanti Barman P , Kumar Thatoi P , Tripathy R , Kumar Das B , Ravindran B . Non-classical monocytes display inflammatory features: validation in sepsis and systemic lupus erythematous. Sci Rep . 2015;5:13886. doi:10.1038/srep13886 26358827
- Ziegler-Heitbrock L , Ancuta P , Crowe S , et al. Nomenclature of monocytes and dendritic cells in blood. Blood . 2010;116(16):e74–80. doi:10.1182/blood-2010-02-258558 20628149
- Ong SM , Hadadi E , Dang TM , et al. The pro-inflammatory phenotype of the human non-classical monocyte subset is attributed to senescence. Cell Death Dis . 2018;9(3):266. doi:10.1038/s41419-018-0327-1 29449647
- Ghanim H , Aljada A , Hofmeyer D , Syed T , Mohanty P , Dandona P . Circulating mononuclear cells in the obese are in a proinflammatory state. Circulation . 2004;110(12):1564–1571. doi:10.1161/01.CIR.0000142055.53122.FA 15364812
- Dandona P , Aljada A , Bandyopadhyay A . Inflammation: the link between insulin resistance, obesity and diabetes. Trends Immunol . 2004;25(1):4–7. doi:10.1016/j.it.2003.10.013 14698276
- Aljada A , Garg R , Ghanim H , et al. Nuclear factor-kappaB suppressive and inhibitor-kappaB stimulatory effects of troglitazone in obese patients with type 2 diabetes: evidence of an antiinflammatory action? J Clin Endocrinol Metab . 2001;86(7):3250–3256. doi:10.1210/jcem.86.7.7564 11443197
- Aljada A , Friedman J , Ghanim H , et al. Glucose ingestion induces an increase in intranuclear nuclear factor kappaB, a fall in cellular inhibitor kappaB, and an increase in tumor necrosis factor alpha messenger RNA by mononuclear cells in healthy human subjects. Metabolism . 2006;55(9):1177–1185. doi:10.1016/j.metabol.2006.04.016 16919536
- Aljada A , Mohanty P , Ghanim H , et al. Increase in intranuclear nuclear factor kappaB and decrease in inhibitor kappaB in mononuclear cells after a mixed meal: evidence for a proinflammatory effect. Am J Clin Nutr . 2004;79(4):682–690. doi:10.1093/ajcn/79.4.682 15051615
- Aljada A , Ghanim H , Mohanty P , Kapur N , Dandona P . Insulin inhibits the pro-inflammatory transcription factor early growth response gene-1 (Egr)-1 expression in mononuclear cells (MNC) and reduces plasma tissue factor (TF) and plasminogen activator inhibitor-1 (PAI-1) concentrations. J Clin Endocrinol Metab . 2002;87(3):1419–1422. doi:10.1210/jcem.87.3.8462 11889219
- Dandona P , Mohanty P , Hamouda W , et al. Inhibitory effect of a two day fast on reactive oxygen species (ROS) generation by leucocytes and plasma ortho-tyrosine and meta-tyrosine concentrations. J Clin Endocrinol Metab . 2001;86(6):2899–2902. doi:10.1210/jcem.86.6.7745 11397907
- Pifferi F , Terrien J , Marchal J , et al. Caloric restriction increases lifespan but affects brain integrity in grey mouse lemur primates. Commun Biol . 2018;1:30. doi:10.1038/s42003-018-0024-8 30271916
- Heilbronn LK , de Jonge L , Frisard MI , et al. Effect of 6-month calorie restriction on biomarkers of longevity, metabolic adaptation, and oxidative stress in overweight individuals: a randomized controlled trial. JAMA . 2006;295(13):1539–1548. doi:10.1001/jama.295.13.1539 16595757
- Lee A , Morley JE . Metformin decreases food consumption and induces weight loss in subjects with obesity with type II non-insulin-dependent diabetes. Obes Res . 1998;6(1):47–53. doi:10.1002/j.1550-8528.1998.tb00314.x 9526970
- Giannarelli R , Aragona M , Coppelli A , Del Prato S . Reducing insulin resistance with metformin: the evidence today. Diabetes Metab . 2003;29(4 Pt 2):6S28–35. doi:10.1016/S1262-3636(03)72785-2 14502098
- Saisho Y . Metformin and inflammation: its potential beyond glucose-lowering effect. Endocr Metab Immune Disord Drug Targets . 2015;15(3):196–205. doi:10.2174/1871530315666150316124019 25772174
- Cameron AR , Morrison VL , Levin D , et al. Anti-inflammatory effects of metformin irrespective of diabetes status. Circ Res . 2016;119(5):652–665. doi:10.1161/CIRCRESAHA.116.308445 27418629