
Abstract
Obesity has become a widespread disease that is harmful to human health. Fat homeostasis is essentially maintained by fat accumulation and energy expenditure. Studies on brown adipose tissue (BAT) represent a promising opportunity to identify a pharmaceutical intervention against obesity through increased energy expenditure. Long non-coding RNAs (lncRNAs) were thought to be critical regulators in a variety of biological processes. Recent studies have revealed that lncRNAs, including ones that are BAT-specific, conserved, and located at key protein-coding genes, function in brown adipogenesis, white adipose browning (ie, beige adipogenesis), and brown thermogenesis. In this review, we describe lncRNA properties and highlight functional lncRNAs in these biological processes, with the goal of establishing links between lncRNAs and BAT. Based on the advances of lncRNAs in the regulation of BAT, we discussed the advantages of potential lncRNA-based obesity drugs. Further BAT lncRNA-based drug development may provide new exciting approaches to defend obesity by regulation of fat homeostasis.
Keywords:
Introduction
In the last several decades, obesity has become a widespread public disease due to a global increase in the number of obese individuals,Citation1,Citation2 and has been clinically evidenced as a high-risk factor for several serious chronic diseases including type 2 diabetes, cardiovascular diseases, and certain fatal cancers.Citation3–Citation5 Obesity develops primarily due to an excessive accumulation of white adipose tissue (WAT), which mainly gets distributed in the subcutaneous and visceral regions.Citation6 While the process is complicated, fat homeostasis is essentially maintained by fat accumulation and energy expenditure. Combating obesity requires either a decrease in fat accumulation or an increase in energy expenditure. In this regard, tremendous progress has been made to understand the mechanism of WAT development in order to identify a method to decrease fat accumulation.Citation7–Citation9 Furthermore, strategies based on brown adipose tissue (BAT), another type of adipose tissue specialized for energy expenditure, have been conceived in recent years.
Studies on brown adipose tissue (BAT) have been considered a promising opportunity to find a pharmaceutical intervention for the treatment of obesity with energy dissipation, and therefore, have attracted considerable attention.Citation10–Citation12 BAT is a type of adipose tissue that had originally been observed in the interscapular region of rodents and newborn infants, and could uniquely induce non-shivering thermogenesis via un-coupling respiration licensed through un-coupling protein 1 (UCP1) on the mitochondrial membrane under cold exposure.Citation13,Citation14 BAT is innervated by the sympathetic system and histological studies have shown that nerve fibers distribute both at the parenchymal space and around blood vessels of BAT.Citation15,Citation16 In contrast to white adipocytes that contain few mitochondria and unilocular lipid droplets, brown adipocytes contain abundant mitochondria and multilocular lipid droplets, which matches BAT function of energy expenditure. Interestingly, a BAT-like fat (referred to beige fat thereafter) has been found to be experimentally induced in WAT under the condition of constant cold exposureCitation17 or β-adrenergic receptor activation,Citation18 and shares common characteristics with bona fide BAT including high expression of UCP1 and the capacity of uncoupling respiration.
The presence of BAT in humans was reported to positively correlate with resting metabolic rate and negatively correlate with BMI, and genetic ablation of BAT in mice increased susceptibility to obesity.Citation19–Citation21 Hence, increased attention has been placed on orchestrated genetic mechanisms of brown adipogenesis, the transition from WAT to BAT (referred to as ‘beige adipogenesis’ thereafter), and brown thermogenesis. Some transcription factors (TFs) and cofactors, including peroxisome proliferator-activated receptor α (PPARα), PPARγ, CCAAT/enhancer-binding protein β (C/EBPβ), transcription factor early B-cell factor 2 (EBF2), PR domain containing 16 (PRMD16), zinc finger and BTB domain-containing 7b (Zbtb7b), and PPARγ coactivator 1α (PCG1α) have been identified to modulate different BAT biological processes.Citation22–Citation25
As a class of regulatory RNA molecules, long non-coding RNAs (lncRNAs) have attracted increased attention in the field of genetics. LncRNAs continue to be increasingly identified in several different types of tissue among vertebrates and have been found to be involved in many biological processes including cell proliferation and differentiation,Citation26,Citation27 cell aging,Citation28 and disease development.Citation29 On the other hand, recent studies have highlighted an important role for lncRNAs in regulating fat metabolism.Citation30 However, the relationship between lncRNAs and the BAT biological processes has not yet been fully elucidated. Recent studies have indicated that many different lncRNAs participate in the regulatory network of brown adipogenesis, beige adipogenesis, and brown thermogenesis.Citation11,Citation12,Citation31–Citation37 Herein, we depict lncRNA characteristics and summarize recent advances that have been made on the most prominent lncRNAs in the regulation of biological processes in brown/beige fat, and thus may facilitate a better understanding of lncRNAs and their regulatory mechanisms in BAT.
An Overview of LncRNA
Canonically, the genetic central dogma indicates that messenger RNAs (mRNAs) are the intermediary molecules that serve to deliver information from DNA to the production of a protein.Citation38 However, with the rapid development of high-throughput sequencing technology, increasingly more RNA molecules that are not able to code for protein production have been demonstrated to play important roles in a variety of biological processes, despite the fact that they were historically thought to be junk RNAs or transcriptional noise.Citation39 Since these RNAs cannot be translated into proteins, they were termed as non-coding RNAs (ncRNAs). The Human Genome Project revealed that only a very small portion of the entire genome can actually be transcribed into RNAs, and a substantial part of the total transcripts were ncRNAs.Citation40,Citation41 Generally, ncRNAs can be classified into transfer RNAs (tRNAs), ribosomal RNAs (rRNAs), PIWI-interacting RNAs (piRNAs), microRNAs (miRNAs) and lncRNAs.
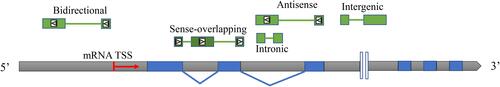
LncRNAs are a type of endogenous RNA molecule that contains >200 nucleotide residues and lacks functional open reading frames (ORFs).Citation42 Based on the location of lncRNAs in the genome, they can be separated into five types including intergenic lncRNAs (lincRNAs), antisense lncRNAs (AS lncRNAs), intronic lncRNAs, sense-overlapping lncRNAs, and bidirectional lncRNAsCitation43,Citation44 (). In contrast to the functional role of other ncRNAs, such as miRNAs that repress mRNA translation by binding to the target mRNA 3ʹ-untranslated region (UTR),Citation45 the functional role of lncRNAs in the biological process is significantly complex. LncRNAs can act as competing endogenous RNAs (ceRNAs) to neutralize miRNAs to protect mRNA translation from miRNA-repression in the cytosol, thus leading to positive regulation of mRNA expression.Citation46,Citation47 Interestingly, lncRNAs that act as ceRNAs competitively bind to not only miRNAs but also proteins, which negatively regulates its corresponding protein.Citation48 LncRNAs that act as molecular vehicles recruit DNA methyltransferases or histone modifiers to the genome or chromatin to change properties of the epigenome and regulate genomic transcription.Citation11,Citation49 LncRNAs that act as imprinted genes specifically modulate the expression of monoallelic genes.Citation11,Citation50 LncRNAs that act as precursors of small RNAs generate regulatory miRNAs that exert additional function of repressing relevant mRNAs.Citation51 On the other hand, lncRNAs can bind to specific proteins in order to influence pre-mRNA processing, thus regulating the diversity of gene isoforms.Citation52
Figure 1 Classification of lncRNAs. The red arrow represents mRNA transcriptional start site (TSS) and transcriptional direction. The blue blocks represent coding gene exons and the grey blocks represent introns or intergenic regions of the genome. The green blocks represent lncRNA exons and their relative position to mRNA exons or TSS. The triangles in the green blocks represent the relative direction of lncRNAs to mRNA. The lncRNA without a triangle indicates that it could be a transcript from a positive-strand and negative-strand of the genome.
LncRNAs in BAT
BAT Tissue-Specific LncRNAs: Essential or Positive LncRNA Roles in BAT Biological Processes
Definition of tissue-specific RNAs from different types of tissue can help identify potential functional BAT genes. Recent studies have revealed a considerable number of lncRNAs with BAT specificity among different types of tissue through de novo reconstruction of transcriptomes or meta-analysis of published datasets.Citation31–Citation33 Among these, three lncRNAs were validated to be involved in BAT biological processes. Thus, we present these three BAT-specific lncRNAs and their functions.
LncRNA-BATE1
Alvarez-Dominguez et al designed a pipeline for tissue-specific RNA-seq analysis to identify potential regulatory lncRNA candidates in BAT.Citation31 Transcriptome profiling analysis was utilized to comprehensively detect a total of 1535 polyadenylated lncRNAs in BAT, epididymal WAT (eWAT), and inguinal WAT (iWAT), among which 127 lncRNAs exhibited BAT-specificity across 30 types of primary tissue datasets retrieved from the mouse ENCODE project.Citation31 Overall, approximately a third of the 127 lncRNAs were found to be up-regulated during brown adipogenesis and targeted by key TFs (PPARγ, C/EBPα, and C/EBPβ).Citation31,Citation53 In total, 40 lncRNAs were selected based on three criteria of degree of BAT specificity, degree of up-regulation during brown adipogenesis, and abundance of expression. These 40 lncRNAs were therefore called lncRNA-BATEs (s = 1–40).
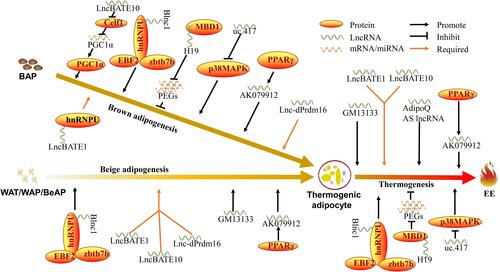
Among the lncRNA-BATEs, LncRNA-BATE1 was found to be a lincRNA containing 2 exons and almost averagely distributed across both the nucleus and cytosol of mature brown adipocytes.Citation31 Knockdown (KD) of lncRNA-BATE1 during brown preadipocytes differentiation led to a limited change in lipid accumulation but significant down-regulation of BAT marker genes including Cidea, C/EBPβ, PGC1α, PRDM16, UCP1 and PPARα, indicating that lncRNA-BATE1 has a limited influence on lipogenesis but a great impact on brown adipogenesis.Citation31 KD of lncRNA-BATE1 was found to hinder UCP1 and PGC1α expression during induction of white preadipocyte brown differentiation, suggesting that lncRNA-BATE1 has a function in the beige adipogenesis.Citation31 Oxygen consumption assay showed lower oxygen consumption with KD of lncRNA-BATE1 in brown adipocytes, indicating an essential function in the thermogenesis of brown adipocytes.Citation31 Moreover, global gene expression analysis conducted by RNA-seq in mouse mature brown adipocytes revealed that depletion of lnc-BATE1 led to down-regulation of genes that were related to brown adipogenesis and mitochondrial biogenesis and function.Citation31 A nuclear matrix factor of heterogeneous nuclear ribonucleoprotein U (hnRNPU) was previously reported to guide lncRNA-Firre, an essential lncRNA for white adipogenesis, to its target to play role.Citation53,Citation54 RNA immunoprecipitation (RIP) showed that lncRNA-BATE1 interacts with hnRNPU to form a functional ribonucleoprotein complex to regulate brown adipocyte development (). This comprehensive data clearly sheds light onto a functional lncRNA that is required for BAT brown adipogenesis, beige adipogenesis, and thermogenesis.
Figure 2 LncRNAs involved in BAT biological processes.
LncRNA-BATE10
Similar to lncRNA-BATE1, lncRNA-BATE10 is also a member of the lncRNA-BATEs family that manifests BAT-specificity. Another systematic transcriptome study on adipose tissue indicated that lncRNA-BATE10 was dramatically induced in iWAT by cold exposure, β-adrenergic agonist stimulation, and extensive physical exercise, respectively.Citation32 Furthermore, lncRNA-BATE10 was found to be up-regulated during activation in cold exposure for 6 hours and down-regulated during inactivation at 30°C for 7 days in BAT, indicating opposite directions of the gene expression program upon the dual stimulations of temperature.Citation32 These lncRNA transcriptome analyses indicate a potential function of lncRNA-BATE10 in the response to a variety of stimulations in both BAT and WAT. LncRNA-BATE10 is approximately 1.7 kb in length, contains 4 exons, and is located in an intergenic region of mouse chromosome 18.Citation32 Similar to lncRNA-BATE1, KD of lncRNA-BATE10 did not cause a significant difference in cell morphology, lipid accumulation, and expression of common lipid markers including PPARγ, CEBPα, and FABP4 during differentiation of brown preadipocytes.Citation32 However, KD of lncRNA-BATE10 led to significant impairment in BAT-selective marker genes including UCP1 and PGC1α, and down-regulation of respiratory electron transport at the genome-wide level, as detected by gene-set enrichment analysis (GSEA),Citation32,Citation55 a knowledge-based approach for interpreting genome-wide expression profiles. On the other hand, lncRNA-BATE10 is required for beige adipogenesis as KD of lncRNA-BATE10 led to down-regulation in the expression of BAT-selective genes during both processes of white primary adipocytes browning and iWAT browning.Citation32 Further analysis of the lncRNA-BATE10 on thermogenic activation of brown adipocytes stimulated by norepinephrine (NE) in the differentiated brown adipocytes showed that KD of the lncRNA-BATE10 hindered the up-regulation of UCP1 and PGC1α, suggesting an essential role for BAT-selective gene expression program in thermogenesis.Citation32 Celf1, an RNA-binding protein (RBP), binds the 3ʹ UTR of its target mRNAs to promote RNA degeneration and repressing translation.Citation56 The promoter binding site and truncated promoter transfection assay determined that the lncRNA-BATE10 is regulated by the cAMP-Creb axis and further experiments validated that it mechanistically competes with PGC1α mRNA for Celf1 binding during differentiation of brown preadipocytes,Citation32 as it is a ceRNA in brown adipogenesis ().
AK079912
The lncRNA AK079912 was originally identified in mouse using a full-length cDNA functional annotation.Citation57 Xiong et al found that AK079912 was a brown adipose enriched lncRNA using bioinformatics analysis of published RNA-seq datasets, which indicated that AK079912 expression was up-regulated 100-fold in BAT compared to eWAT.Citation33 Cold exposure induced up-regulation of AK079912, Blnc1 (a lncRNA gene related to BAT development), and UCP1 in both BAT and iWAT, indicated a potential role of AK079912 in thermogenesis.Citation33 Additionally, KD and overexpression of AK079912 revealed that AK079912 is a positive regulator of brown adipogenesis, WAT browning, and brown adipocyte mitochondrial biogenesis.Citation33 Mechanistically, the transcription of AK079912 was found to be regulated by PPARγ binding to the AK079912 promoterCitation33 (). These results reported a PPARγ-regulated positive modulator of lncRNA expression involved in brown adipogenesis, beige adipogenesis, and brown thermogenesis.
Conserved LncRNAs Between Human and Mouse: Most Potential LncRNAs of Preclinical Significance for Humans Fighting Against Obesity
For a long time, it was generally accepted that lncRNAs were poorly conserved across species,Citation58–Citation60 which hindered the exploration of lncRNAs among different species. However, recent studies have demonstrated several sequences and functionally conserved BAT lncRNAs between humans and mice. These conserved lncRNAs might have the most potential and preclinical significance for the fight against obesity.
Blnc1
Blnc1 is a lncRNA that contains a single exon that is approximately 965 bp in length. The function and mechanism of Blnc1 during brown adipogenesis was initially illustrated by Zhao et al.Citation34 Blnc1 is transcribed from the intergenic region and was found to be up-regulated during brown adipocyte differentiation and exhibits specificity of expression in BAT compared to eWAT.Citation34 Additionally, Blnc1 expression was induced in eWAT when treated with a β3-adrenergic agonist of CL-316,243.Citation34 This lncRNA is thought to be essential for brown adipogenesis as KD of this lncRNA significantly impaired brown adipogenesis.Citation34 In vivo, gain-of-function transplantation experiment strongly implicated Blnc1 act as a lncRNA activator for brown adipogenesis and brown fat formation as fad pad formed from exogenous preadipocytes that had transduced Blnc1 retroviral vectors expressed higher levels of UCP1, Cidea, and FABP4.Citation34 Furthermore, gain- and loss-of-function during differentiation of beige adipocyte progenitor reveal that Blnc1 is sufficient and required for beige adipogenesis.Citation34 Additionally, further binding event study revealed that Blnc1 exerts its function by forming a ribonucleoprotein complex with EBF2.Citation34 These discoveries on the murine model exhibited a powerful positive lncRNA regulator of the BAT biological processes.
To investigate whether Blnc1 is functionally conserved across humans and mice, as well as Blnc1’s molecular mechanism in humans, Mi et al analyzed genomic DNA sequences adjacent to mouse and human Paqr9 gene and found mouse Blnc1 and human Blnc1 were both transcribed near from Paqr9 gene of their own genome.Citation61 Obtaining full-length cDNA sequence and alignment analysis indicated that mouse and human Blnc1 contain two highly conserved segments as they share high sequence identity, which suggests sequence conservation between mouse Blnc1 and human Blnc1.Citation61 Interestingly, overexpression of human Blnc1 during mouse brown preadipocytes differentiation exhibited functional conservation in induced thermogenic genes and human Blnc1 was able to rescue KD of mouse Blnc1, indicating functional conservation between the human and mouse Blnc1.Citation61 The ability of lncRNAs to form secondary structures probably contributes to its complicated function.Citation62,Citation63 Blnc1 was predicted to contain three different domains, with one domain being responsible for transcriptional activation of brown adipocytes genes including UCP1, Elvol3, Pcg1α, and Dio2.Citation61 Blnc1 has been shown to physically interact with hnRNPU which, in turn, physically interacts with EBF2.Citation61 Thus, hnRNPU acts as a vehicle between Blnc1 and EBF2. Intriguingly, hnRNPU also serves as a medium to catenate Blnc1 to Zbtb7b (), another transcription cofactor that reportedly plays a crucial role in brown fat development.Citation25 Collectively, these studies reveal Blnc1 acts as a positive conserved lncRNA in the regulation of brown adipogenesis, beige adipogenesis, and brown thermogenesis.
H19
As it was first identified in imprinted lncRNA, H19 has been shown to play a role in multiple biological processes including cancer formation,Citation64 cell proliferation and differentiation,Citation65,Citation66 and immune response.Citation67 This lncRNA is one of the transcripts transcribed from the imprinted H19/IGF2 locus. H19 is conserved on the human 11p15.5 and mouse chromosome 7, and is expressed exclusively from the maternal allele.Citation64,Citation68 A recent study revealed that H19 was highly expressed in BAT in comparison to subcutaneous WAT (scWAT) and visceral WAT (vWAT).Citation11 H19 was found to be, respectively, up-regulated and decreased in cold exposure and dietary-induced obesity mice BAT, but did not show a similar manner of expression in scWAT and vWAT in the two conditions,Citation11 which argues for selective involvement in brown adipogenesis over beige adipogenesis. During differentiation of the BAT stromal-vascular fraction (SVF) adipocyte precursor cell, RNA interference (RNAi) led to hindered lipid accumulation, impaired expression of BAT markers (ie, UCP1 and Cidea) and several common adipose tissue markers (ie, PPARγ, FABP4, and CEBPα), indicating that H19 is required for commitment of BAT.Citation11 In vivo, H19 transgenic (TG) mice strongly prevent diets-induced weight gain with mounting energy expenditure, and H19 expression was found to be negatively related to BMI in human population, which suggests that it may function as a potential therapeutic molecule for energy dissipation and counteraction of obesity.Citation11 Further interrogation of the underlying mechanism on H19 regulating BAT revealed that H19 improved insulin sensitivity and mitochondrial biogenesis, and managed imprinted gene networks that consisted of paternally expressed genes (PEGs) including GPR1, IGF2, Peg10, Peg3, Plagl1, miR184, miR298, and miR335 by recruiting MBD1 (a chromatin modifier)Citation11 (). These data clearly indicate an important role for H19 in BAT protection from dietary obesity by constraining monoallelic gene expression to promote brown adipogenesis and thermogenesis.
Lnc-Uc.417
Aging is accompanied by a decline in the regenerative capacity of brown adipocyte in humans.Citation19,Citation69 In order to investigate age-related lncRNAs in BAT, the ultra-conserved lncRNA lnc-uc.417 that is primarily distributed in the nucleus and increases with age in BAT was selected to further research.Citation35 This lncRNA was found to share highly conserved sequence similarity among mouse, human, dog, elephant, chicken, African clawed fogs, and zebrafish. Furthermore, lnc-uc.417 shares 100% sequence identity between human and mouse.Citation35 Cold induction and agonist stimulation (CL-316,243, forskolin, or db-cAMP) reduced lnc-uc.417 expression in BAT and differentiated interscapular BAT primary SVF cells, respectively.Citation35 Overexpression of lnc-uc.417 mediated by lentivirus transduction during brown preadipocyte differentiation led to limited changes in lipid accumulation but significant a decreased in expression of brown adipogenic genes including PRDM16, PPARγ2, C/EBPβ, EBF2, and Cidea.Citation35 Moreover, overexpression of lnc-uc.417 led to not only a lower respiration rate in the mitochondria but also decreased expression of related mitochondrial genes, such as UCP1, PGC1α, and CytC.Citation35 Further bioinformatics analysis and protein detection have demonstrated that lnc-uc.417 moderately suppresses the phosphorylation of p38MAPK without affecting the total protein levels of p38MAPKCitation35 (). These data indicate that lnc-uc.417 functions as a conserved negative lncRNA regulator of brown adipogenesis and thermogenesis.
Lnc-dPrdm16
LncRNA expression is generally specific to certain cell types and most of the detectable lncRNAs can be significantly differentially expressed, even among closely related cell type,Citation70,Citation71 which hinders the understanding of human adipose lncRNAs. In order to address this issue, Ding et al constructed a comprehensive human adipose lncRNA catalog from human fetal BAT, adult omental WAT (oWAT), and adult scWAT. Lnc-dPrdm16, also known as LINC00982, is one of the lncRNAs from the catalog that was found to be syntenically conserved between humans and mice.Citation12 Lnc-dPrdm16 contains 2 exons, is mainly expressing in the cytosol, and was found to be a bidirectional lncRNA that is located divergently from Prdm16, a master regulator for brown adipocyte biology.Citation12 Lnc-dPrdm16 is known to positively regulate Prdm16 expression at the beginning of Ding’s research and was predicted to exert canonical functions including lipid catabolic processes, cellular lipid metabolic processes, and fatty acid catabolic processes using Gene Ontology (GO) enrichment based on its highly co-expressed protein-coding genes.Citation12,Citation72 Furthermore, RT-qPCR results showed lnc-dPrdm16 is significantly higher expressed in BAT than oWAT and scWAT, indicating a potential regulatory function in BAT. KD of lnc-dPrdm16 during primary brown adipocytes differentiation led to a reduction in lipid accumulation, down-regulation of general adipogenic markers, and down-regulation of brown fat markers (ie, UCP1, C/EBPβ, and lnc-BATE1), suggesting that it functions as a required regulator in brown adipogenesis. Additionally, KD of lnc-dPrdm16 led to a reduction of both BAT-selective and pan-adipogenic marker expression during induction of iWAT browning in vivo, indicating that lnc-dPrdm16 is required for maintaining a mature adipocyte program in WAT and BAT marker induction during iWAT browningCitation12 ().
Anti-Sense LncRNAs Located at Key Protein-Coding Genes: Positive Regulators in Beige Adipogenesis and Thermogenesis
LncRNAs can regulate genes proximal to them, which is known as cis-regulation.Citation73,Citation74 As a secretory organ, WAT is reported to secrete hormones in an autocrine, paracrine, and endocrine manner.Citation75 LncRNAs that are located on the locus of a hormone gene might regulate their corresponding hormone gene, which helps expand our understanding of the metabolic functions of BAT. On the other hand, some known critical BAT TF genes might be regulated by certain lncRNAs. However, whether these key protein-coding genes are regulated by neighboring lncRNAs in a cis-regulatory manner might need further investigation.
AdipoQ as LncRNA
Previous studies have shown that AS lncRNAs regulate their neighboring genes both positively or negatively.Citation76,Citation77 AdipoQ AS lncRNA is translated from the opposite strand to AdipoQ mRNA transcript. The AdipoQ protein is a selectively expressed hormone in adipocytes that modulates glucose and lipid metabolism in order to maintain energy homeostasis.Citation78 A recent study demonstrated that overexpression of AdipoQ AS lncRNA by intraperitoneal injection of adenoviruses into mice that consumed a high-fat diet (HFD) led to an up-regulation of BAT thermogenic genes including UCP1, PGC1α, and PRDM16, and decreased BAT triglycerides (TG) content.Citation36 Further interrogation of AdipoQ AS lncRNA regulation during adipogenesis revealed that it is bound to the AdipoQ mRNA to form an RNA duplex (). In addition, the distribution of AdipoQ AS lncRNAs in adipocytes has been reported to exhibit dynamic change during differentiation, and is distributed approximately equally into nucleus and cytoplasm in preadipocytes, and almost completely distributed in the cytoplasm in mature adipocytes,Citation36 which indicates a transferring process during preadipocyte differentiation. These findings provide a potential novel therapeutic target of AS lncRNA located at a hormone gene, and might protect patients from obesity-related metabolism diseases. AdipoQ AS lncRNA acts as a ceRNA to attenuate AdipoQ gene post-transcriptional translation. However, whether this lncRNA regulates an AdipoQ genomic element to influence AdipoQ transcription in cis-manner might need further investigation.
GM13133
You et al identified GM13133, an AS lncRNA that is 736 bp in length and overlaps with PRDM16, which is significantly induced in BAT during canonical adipose browning protocol of cold exposure, β-adrenergic agonist, and cAMP.Citation37 Overexpression of GM13133 during white adipocyte differentiation causes an increased number of mitochondria, up-regulation of UCP1 expression, slightly increased oxygen consumption rate and gives rise to the characteristics of brown adipocytes.Citation37 Further investigation of differentially expressed genes during white-to-brown adipocytes transition with the interference of GM13133 revealed that GM13133 regulates the thermogenic program through cAMP signaling pathway.Citation37 Overexpression of GM13133 moderately but not significantly up-regulates its neighboring gene PRDM16 expression.Citation37 On the other hand, whether GM13133 is required for PRDM16 expression is unknown. Therefore, whether PRDM16 is cis-regulated by GM13133 might need further validation. Overall, these results highlight an AS lncRNA that plays an important role in beige adipogenesis and brown thermogenesis ().
Conclusions and Future Perspectives
BAT has been considered a potential target for tackling obesity due to its unique function of energy expenditure. LncRNAs were a class of key regulators that are involved in many biological processes including BAT development. LncRNAs are defined by their transcript length and protein-coding ability. The length of a lncRNA transcript is measured from exon start and end site. On the other hand, the protein-coding ability of a lncRNA cannot easily be calculated precisely. In this review, most of the lncRNAs were predicted using multiple coding software in the analysis pipeline in order to ensure that they were indeed non-coding.Citation12,Citation31,Citation32,Citation34,Citation35 Notably, Blnc1Citation34 and uc.417Citation35 were identified using extra mass hypothetical protein expression and spectrometry analysis in vitro, respectively, which improved the lncRNA reliability that these are non-coding RNAs. The combination of bioinformatics prediction and in vitro protein analysis is worthy of being adopted in the identification of novel lncRNAs in future studies. The transcripts with a single exon generally become discarded in the data processing of lncRNA identification in several studies.Citation12,Citation31,Citation79 However, the single exon lncRNA Blnc1 was proven to play an important role in BAT biological processes,Citation34 which suggests more attention should be paid to single exon lncRNA identification. RNAi was employed to explore BAT functional lncRNAs and lncRNAs including lnc-BATE1Citation31 and Blnc1Citation34 were verified by the rescue experiments, which was able to verify their robust functional role in BAT biological processes. Despite the fact that AS lncRNAs are generally considered to function in cis-manner, there has been no evidence that AS lncRNAs only play role in cis. Hence, further research should be conducted to indicate both the cis- and trans-regulation of AS lncRNAs in BAT. Among different types of ncRNAs, binding to specific protein is one of the hallmarks of lncRNAs. Experiments such as RNA immunoprecipitation, RNA pull down, and immunoblotting have confirmed that some lncRNAs, including lnc-BATE1, lnc-BATE10, Blnc1 and H19, interact with certain proteins in BAT biological processes. However, the protein partners of several lncRNA, such as lnc-dPrdm16, GM13133, and AdipoQ AS lncRNA, have yet to be elucidated. On the other hand, the lncRNA functions, such as neutralizing miRNA to protect mRNA translation from miRNA binding, have been found in WAT or other tissue development,Citation80–Citation83 though it, remains to be explored in BAT.
Despite the fact that many lncRNAs have been identified to play potential roles in BAT development by de novo reconstruction of transcriptome,Citation12,Citation31,Citation32 only a small portion of lncRNAs including BAT-specific ones, sequence and functional conserved ones, and those located at key protein-coding genes have been verified to play crucial roles in the brown adipogenesis, beige adipogenesis, or brown thermogenesis in vitro or in vivo (, ). Most lncRNAs play roles that depend on the regulation of certain TFs or transcription cofactors, such as EBF2, PPARγ, and Celf1 (). Despite the complexity of lncRNA mechanisms in regulating the BAT biological network, emerging technologies such as single-cell RNA-seq,Citation84 chromatin accessibility analysis,Citation85 and multi-omics studyCitation86 have shown the powerful capacity to reveal deeper insight into RNA regulatory mechanisms and may contribute to the precise interrogation of the mechanisms by which lncRNA regulates BAT biological processes. Inversely, efficient genetic editing of lncRNA gene might help to validate lncRNA functions in vivo, which can speed up the preclinical research of lncRNA-based drugs. In contrast to other types of RNA molecules (such as mRNAs), lncRNAs are expressed at relatively lower levels in tissues, which suggests that most lncRNAs have a powerful regulatory function at even a low expression level. Hence, possible lncRNA-based drugs can be more efficient at low doses in treatment. LncRNAs, for instance Blnc1, exhibit the ability to directly recruit certain TFs to exert transcriptional functions (), thus implicating the efficiency of potential lncRNA-based drugs. However, to date, there is not an approved BAT lncRNA-based drug for the treatment of obesity. Recent studies have shown that small-molecule drugs can help regulate lncRNA expression,Citation87 which indicates that small-molecule drugs that target lncRNA genes can modulate BAT lncRNA expression, and thus might be a novel treatment method for human obesity treatment. On the other hand, with the development of precision medicine being applied to the treatment of obesity, specific oligonucleotides that are based on precise base complementarity could be designed to target BAT lncRNAs,Citation88 thus regulating the expression of BAT lncRNAs in the fight against obesity.
Table 1 Characterization of LncRNAs in the Regulating BAT Biological Processes
To date, although limited lncRNAs have been identified to play important roles in BAT biological processes, many BAT lncRNAs and their complex regulatory mechanisms remain to be determined. Further lncRNA studies including in vitro and in vivo experiments may help develop novel exciting approaches to defend obesity by regulating fat homeostasis.
Ethics Statement
The authors declare no funding and no conflicts of interest for this work. All figures in this review are original.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Acknowledgment
This work was supported by the Earmarked Fund for China Agriculture Research System (No. CARS-44-A-2).
Disclosure
The authors report no conflicts of interest for this work.
References
- You W, Henneberg M. Relaxed natural selection contributes to global obesity increase more in males than in females due to more environmental modifications in female body mass. PLoS One. 2018;13(7):e0199594. doi:10.1371/journal.pone.0199594
- Prentice AM. The emerging epidemic of obesity in developing countries. Int J Epidemiol. 2006;35(1):93–99. doi:10.1093/ije/dyi272
- Fox CS, Massaro JM, Hoffmann U, et al. Abdominal visceral and subcutaneous adipose tissue compartments: association with metabolic risk factors in the Framingham heart study. Circulation. 2007;116(1):39–48. doi:10.1161/CIRCULATIONAHA.106.675355
- Kim HJ, Kwon H, Jeong SM, Hwang SE, Park JH. Effects of abdominal visceral fat compared with those of subcutaneous fat on the association between PM10 and hypertension in Korean men: A cross-sectional study. Sci Rep. 2019;9(1):5951. doi:10.1038/s41598-019-42398-1
- Ogden CL, Carroll MD, Flegal KM. Prevalence of obesity in the United States. JAMA. 2014;312(2):189–190. doi:10.1001/jama.2014.6228
- Rosen ED, MacDougald OA. Adipocyte differentiation from the inside out. Nat Rev Mol Cell Biol. 2006;7(12):885–896. doi:10.1038/nrm2066
- Cai R, Tang G, Zhang Q, et al. A novel lnc-RNA, named lnc-ORA, is identified by RNA-Seq analysis, and its knockdown inhibits adipogenesis by regulating the PI3K/AKT/mTOR signaling pathway. Cells. 2019;8(5):E477. doi:10.3390/cells8050477
- Zhang L, Zhang D, Qin ZY, Li J, Shen ZY. The role and possible mechanism of long noncoding RNA PVT1 in modulating 3T3-L1 preadipocyte proliferation and differentiation. IUBMB Life. 2020;72(7):1460–1467. doi:10.1002/iub.2269
- Shen L, Han J, Wang H, et al. Cachexia-related long noncoding RNA, CAAlnc1, suppresses adipogenesis by blocking the binding of HuR to adipogenic transcription factor mRNAs. Int J Cancer. 2019;145(7):1809–1821. doi:10.1002/ijc.32236
- Bhatt PS, Dhillo WS, Salem V. Human brown adipose tissue-function and therapeutic potential in metabolic disease. Curr Opin Pharmacol. 2017;37:1–9. doi:10.1016/j.coph.2017.07.004
- Schmidt E, Dhaouadi I, Gaziano I, et al. LincRNA H19 protects from dietary obesity by constraining expression of monoallelic genes in brown fat. Nat Commun. 2018;9(1):3622. doi:10.1038/s41467-018-05933-8
- Ding C, Lim YC, Chia SY, et al. De novo reconstruction of human adipose transcriptome reveals conserved lncRNAs as regulators of brown adipogenesis. Nat Commun. 2018;9(1):1329. doi:10.1038/s41467-018-03754-3
- Peirce V, Carobbio S, Vidal-Puig A. The different shades of fat. Nature. 2014;510(7503):76–83. doi:10.1038/nature13477
- Roesler A, Kazak L. UCP1-independent thermogenesis. Biochem J. 2020;477(3):709–725. doi:10.1042/BCJ20190463
- Bargmann W, V. Hehn G, Lindner E. Über die Zellen des braunen Fettgewebes und ihre Innervation. Zeitschrift für Zellforschung und Mikroskopische Anatomie. 1968;85(4):601–613. doi:10.1007/BF00324749
- Zhu Q, Glazier BJ, Hinkel BC, et al. Neuroendocrine regulation of energy metabolism involving different types of adipose tissues. Int J Mol Sci. 2019;20(11):2707. doi:10.3390/ijms20112707
- Young P, Arch JR, Ashwell M. Brown adipose tissue in the parametrial fat pad of the mouse. FEBS Lett. 1984;167(1):10–14. doi:10.1016/0014-5793(84)80822-4
- Himms-Hagen J, Cui J, Danforth E Jr, et al. Effect of CL-316,243, a thermogenic beta 3-agonist, on energy balance and brown and white adipose tissues in rats. Am J Physiol. 1994;266(4 Pt 2):R1371–1382. doi:10.1152/ajpregu.1994.266.4.R1371
- Cypess AM, Lehman S, Williams G, et al. Identification and importance of brown adipose tissue in adult humans. N Engl J Med. 2009;360(15):1509–1517. doi:10.1056/NEJMoa0810780
- Feldmann HM, Golozoubova V, Cannon B, Nedergaard J. UCP1 ablation induces obesity and abolishes diet-induced thermogenesis in mice exempt from thermal stress by living at thermoneutrality. Cell Metab. 2009;9(2):203–209. doi:10.1016/j.cmet.2008.12.014
- Lowell BB, SS V, Hamann A, et al. Development of obesity in transgenic mice after genetic ablation of brown adipose tissue. Nature. 1993;366(6457):740–742. doi:10.1038/366740a0
- Farmer SR. Molecular determinants of brown adipocyte formation and function. Genes Dev. 2008;22(10):1269–1275. doi:10.1101/gad.1681308
- Kajimura S, Seale P, Spiegelman BM. Transcriptional control of brown fat development. Cell Metab. 2010;11(4):257–262. doi:10.1016/j.cmet.2010.03.005
- Wu J, Cohen P, Spiegelman BM. Adaptive thermogenesis in adipocytes: is beige the new brown? Genes Dev. 2013;27(3):234–250. doi:10.1101/gad.211649.112
- Li S, Mi L, Yu L, et al. Zbtb7b engages the long noncoding RNA Blnc1 to drive brown and beige fat development and thermogenesis. Proc Natl Acad Sci U S A. 2017;114(34):E7111–e7120. doi:10.1073/pnas.1703494114
- Xu B, Gerin I, Miao H, et al. Multiple roles for the non-coding RNA SRA in regulation of adipogenesis and insulin sensitivity. PLoS One. 2010;5(12):e14199. doi:10.1371/journal.pone.0014199
- Stamatoyannopoulos JA, Snyder M, Hardison R, et al. An encyclopedia of mouse DNA elements (Mouse ENCODE). Genome Biol. 2012;13(8):418. doi:10.1186/gb-2012-13-8-418
- Devaux Y, Zangrando J, Schroen B, et al. Long noncoding RNAs in cardiac development and ageing. Nat Rev Cardiol. 2015;12(7):415–425.
- Peng WX, Koirala P, Mo YY. LncRNA-mediated regulation of cell signaling in cancer. Oncogene. 2017;36(41):5661–5667. doi:10.1038/onc.2017.184
- Zeng Y, Ren K, Zhu X, Zheng Z, Yi G. Long noncoding RNAs: advances in lipid metabolism. Adv Clin Chem. 2018;87:1–36.
- Alvarez-Dominguez JR, Bai Z, Xu D, et al. De novo reconstruction of adipose tissue transcriptomes reveals long non-coding RNA regulators of brown adipocyte development. Cell Metab. 2015;21(5):764–776. doi:10.1016/j.cmet.2015.04.003
- Bai Z, Chai XR, Yoon MJ, et al. Dynamic transcriptome changes during adipose tissue energy expenditure reveal critical roles for long noncoding RNA regulators. PLoS Biol. 2017;15(8):e2002176. doi:10.1371/journal.pbio.2002176
- Xiong Y, Yue F, Jia Z, et al. A novel brown adipocyte-enriched long non-coding RNA that is required for brown adipocyte differentiation and sufficient to drive thermogenic gene program in white adipocytes. Biochim Biophys Acta Mol Cell Biol Lipids. 2018;1863(4):409–419. doi:10.1016/j.bbalip.2018.01.008
- Zhao XY, Li S, Wang GX, Yu Q, Lin JD. A long noncoding RNA transcriptional regulatory circuit drives thermogenic adipocyte differentiation. Mol Cell. 2014;55(3):372–382. doi:10.1016/j.molcel.2014.06.004
- Cui X, You L, Li Y, et al. A transcribed ultraconserved noncoding RNA, uc.417, serves as a negative regulator of brown adipose tissue thermogenesis. FASEB J. 2016;30(12):4301–4312. doi:10.1096/fj.201600694R
- Cai R, Sun Y, Qimuge N, et al. Adiponectin AS lncRNA inhibits adipogenesis by transferring from nucleus to cytoplasm and attenuating Adiponectin mRNA translation. Biochim Biophys Acta Mol Cell Biol Lipids. 2018;1863(4):420–432. doi:10.1016/j.bbalip.2018.01.005
- You L, Zhou Y, Cui X, et al. GM13133 is a negative regulator in mouse white adipocytes differentiation and drives the characteristics of brown adipocytes. J Cell Physiol. 2018;233(1):313–324. doi:10.1002/jcp.25878
- Watson JD, Crick FH. Molecular structure of nucleic acids; a structure for deoxyribose nucleic acid. Nature. 1953;171(4356):737–738. doi:10.1038/171737a0
- Doolittle WF. Is junk DNA bunk? A critique of ENCODE. Proc Natl Acad Sci U S A. 2013;110(14):5294–5300. doi:10.1073/pnas.1221376110
- Stein LD. Human genome: end of the beginning. Nature. 2004;431(7011):915–916. doi:10.1038/431915a
- Siepel A. Finishing the euchromatic sequence of the human genome. Nature. 2005;50(2):931–945.
- Batista PJ, Chang HY. Long noncoding RNAs: cellular address codes in development and disease. Cell. 2013;152(6):1298–1307. doi:10.1016/j.cell.2013.02.012
- Tong C, Chen Q, Zhao L, Ma J, Ibeagha-Awemu EM, Zhao X. Identification and characterization of long intergenic noncoding RNAs in bovine mammary glands. BMC Genomics. 2017;18(1):468. doi:10.1186/s12864-017-3858-4
- Reik W, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs. Cell. 2009;136(4):629–641. doi:10.1016/j.cell.2009.02.006
- Ambros V. The functions of animal microRNAs. Nature. 2004;431(7006):350–355. doi:10.1038/nature02871
- Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP. A ceRNA hypothesis: the Rosetta stone of a hidden RNA language? Cell. 2011;146(3):353–358. doi:10.1016/j.cell.2011.07.014
- Cesana M, Cacchiarelli D, Legnini I, et al. A long noncoding RNA controls muscle differentiation by functioning as a competing endogenous RNA. Cell. 2011;147(2):358–369. doi:10.1016/j.cell.2011.09.028
- Gong C, Li Z, Ramanujan K, et al. A long non-coding RNA, LncMyoD, regulates skeletal muscle differentiation by blocking IMP2-mediated mRNA translation. Dev Cell. 2015;34(2):181–191. doi:10.1016/j.devcel.2015.05.009
- Zhou J, Yang L, Zhong T, et al. H19 lncRNA alters DNA methylation genome wide by regulating S-adenosylhomocysteine hydrolase. Nat Commun. 2015;6:10221. doi:10.1038/ncomms10221
- Lee JT, Bartolomei MS. X-inactivation, imprinting, and long noncoding RNAs in health and disease. Cell. 2013;152(6):1308–1323. doi:10.1016/j.cell.2013.02.016
- Dey BK, Pfeifer K, Dutta A. The H19 long noncoding RNA gives rise to microRNAs miR-675-3p and miR-675-5p to promote skeletal muscle differentiation and regeneration. Genes Dev. 2014;28(5):491–501. doi:10.1101/gad.234419.113
- Tripathi V, Ellis JD, Shen Z, et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Mol Cell. 2010;39(6):925–938. doi:10.1016/j.molcel.2010.08.011
- Sun L, Goff LA, Trapnell C, et al. Long noncoding RNAs regulate adipogenesis. Proc Natl Acad Sci U S A. 2013;110(9):3387–3392. doi:10.1073/pnas.1222643110
- Hacisuleyman E, Goff LA, Trapnell C, et al. Topological organization of multichromosomal regions by the long intergenic noncoding RNA Firre. Nat Struct Mol Biol. 2014;21(2):198–206. doi:10.1038/nsmb.2764
- Subramanian A, Tamayo P, Mootha VK, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A. 2005;102(43):15545–15550. doi:10.1073/pnas.0506580102
- Amaya Ramirez CC, Hubbe P, Mandel N, Béthune J. 4EHP-independent repression of endogenous mRNAs by the RNA-binding protein GIGYF2. Nucleic Acids Res. 2018;46(11):5792–5808. doi:10.1093/nar/gky198
- Okazaki Y, Furuno M, Kasukawa T, et al. Analysis of the mouse transcriptome based on functional annotation of 60,770 full-length cDNAs. Nature. 2002;420(6915):563–573.
- Cornelis G, Souquere S, Vernochet C, Heidmann T, Pierron G. Functional conservation of the lncRNA NEAT1 in the ancestrally diverged marsupial lineage: evidence for NEAT1 expression and associated paraspeckle assembly during late gestation in the opossum Monodelphis domestica. RNA Biol. 2016;13(9):826–836. doi:10.1080/15476286.2016.1197482
- Huang Y. The novel regulatory role of lncRNA-miRNA-mRNA axis in cardiovascular diseases. J Cell Mol Med. 2018;22(12):5768–5775. doi:10.1111/jcmm.13866
- Kutter C, Watt S, Stefflova K, et al. Rapid turnover of long noncoding RNAs and the evolution of gene expression. PLoS Genet. 2012;8(7):e1002841. doi:10.1371/journal.pgen.1002841
- Mi L, Zhao XY, Li S, Yang G, Lin JD. Conserved function of the long noncoding RNA Blnc1 in brown adipocyte differentiation. Mol Metab. 2017;6(1):101–110. doi:10.1016/j.molmet.2016.10.010
- Somarowthu S, Legiewicz M, Chillón I, Marcia M, Liu F, Pyle AM. HOTAIR forms an intricate and modular secondary structure. Mol Cell. 2015;58(2):353–361. doi:10.1016/j.molcel.2015.03.006
- Xue Z, Hennelly S, Doyle B, et al. A G-rich motif in the lncRNA braveheart interacts with a zinc-finger transcription factor to specify the cardiovascular lineage. Mol Cell. 2016;64(1):37–50. doi:10.1016/j.molcel.2016.08.010
- Ghafouri-Fard S, Esmaeili M, Taheri MH. 19 lncRNA: roles in tumorigenesis. Biomed Pharmacother. 2020;123:109774. doi:10.1016/j.biopha.2019.109774
- Xu X, Ji S, Li W, et al. LncRNA H19 promotes the differentiation of bovine skeletal muscle satellite cells by suppressing Sirt1/FoxO1. Cell Mol Biol Lett. 2017;22(1):10. doi:10.1186/s11658-017-0040-6
- Chen L, Wang Y, He J, Zhang C, Chen J, Shi D. Long non-coding RNA H19 promotes proliferation and invasion in human glioma cells by downregulating miR-152. Oncol Res. 2018;26:1419–1428.
- Li X, Wang H, Zhang Y, et al. Overexpression of lncRNA H19 changes basic characteristics and affects immune response of bovine mammary epithelial cells. Peer J. 2019;7:e6715. doi:10.7717/peerj.6715
- Nordin M, Bergman D, Halje M, Engström W, Ward A. Epigenetic regulation of the Igf2/H19 gene cluster. Cell Prolif. 2014;47(3):189–199. doi:10.1111/cpr.12106
- Virtanen KA, Lidell ME, Orava J, et al. Functional brown adipose tissue in healthy adults. N Engl J Med. 2009;360(15):1518–1525. doi:10.1056/NEJMoa0808949
- Ranzani V, Rossetti G, Panzeri I, Arrigoni A, Pagani M. The long intergenic noncoding RNA landscape of human lymphocytes highlights the regulation of T cell differentiation by linc-MAF-4. Nat Immunol. 2015;16(3):318. doi:10.1038/ni.3093
- Amin V, Harris RA, Onuchic V, et al. Epigenomic footprints across 111 reference epigenomes reveal tissue-specific epigenetic regulation of lincRNAs. Nat Commun. 2015;6:6370. doi:10.1038/ncomms7370
- Huang DW, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4(1):44–57. doi:10.1038/nprot.2008.211
- Zhang B, Arun G, Mao YS, et al. The lncRNA Malat1 is dispensable for mouse development but its transcription plays a cis-regulatory role in the adult. Cell Rep. 2012;2(1):111–123. doi:10.1016/j.celrep.2012.06.003
- Yan P, Luo S, Lu JY, Shen X. Cis- and trans-acting lncRNAs in pluripotency and reprogramming. Curr Opin Genet Dev. 2017;46:170–178. doi:10.1016/j.gde.2017.07.009
- Wang GX, Zhao XY, Lin JD. The brown fat secretome: metabolic functions beyond thermogenesis. Trends Endocrinol Metab. 2015;26(5):231–237. doi:10.1016/j.tem.2015.03.002
- Magistri M, Faghihi MA, St Laurent G 3rd, Wahlestedt C. Regulation of chromatin structure by long noncoding RNAs: focus on natural antisense transcripts. Trends Genet. 2012;28(8):389–396. doi:10.1016/j.tig.2012.03.013
- Halley P, Kadakkuzha BM, Faghihi MA, et al. Regulation of the apolipoprotein gene cluster by a long noncoding RNA. Cell Rep. 2014;6(1):222–230. doi:10.1016/j.celrep.2013.12.015
- Stern JH, Rutkowski JM, Scherer PE. Adiponectin, leptin, and fatty acids in the maintenance of metabolic homeostasis through adipose tissue crosstalk. Cell Metab. 2016;23(5):770–784. doi:10.1016/j.cmet.2016.04.011
- Wang GZ, Du K, Hu SQ, et al. Genome-wide identification and characterization of long non-coding RNAs during postnatal development of rabbit adipose tissue. Lipids Health Dis. 2018;17(1):271. doi:10.1186/s12944-018-0915-1
- Huang X, Fu C, Liu W, et al. Chemerin-induced angiogenesis and adipogenesis in 3 T3-L1 preadipocytes is mediated by lncRNA Meg3 through regulating Dickkopf-3 by sponging miR-217. Toxicol Appl Pharmacol. 2019;385:114815. doi:10.1016/j.taap.2019.114815
- Liu Y, Wang Y, He X, et al. LncRNA TINCR/miR-31-5p/C/EBP-α feedback loop modulates the adipogenic differentiation process in human adipose tissue-derived mesenchymal stem cells. Stem Cell Res. 2018;32:35–42. doi:10.1016/j.scr.2018.08.016
- Liu H, Li H, Jin L, et al. Long noncoding RNA GAS5 suppresses 3T3-L1 cells adipogenesis through miR-21a-5p/PTEN signal pathway. DNA Cell Biol. 2018;37(9):767–777. doi:10.1089/dna.2018.4264
- Li FP, Lin DQ, Gao LY. LncRNA TUG1 promotes proliferation of vascular smooth muscle cell and atherosclerosis through regulating miRNA-21/PTEN axis. Eur Rev Med Pharmacol Sci. 2018;22(21):7439–7447. doi:10.26355/eurrev_201811_16284
- Burl RB, Ramseyer VD, Rondini EA, Pique-Regi R, Lee YH, Granneman JG. Deconstructing adipogenesis induced by β3-adrenergic receptor activation with single-cell expression profiling. Cell Metab. 2018;28(2):300–309.e304. doi:10.1016/j.cmet.2018.05.025
- Wang J, Zibetti C, Shang P, et al. ATAC-Seq analysis reveals a widespread decrease of chromatin accessibility in age-related macular degeneration. Nat Commun. 2018;9(1):1364. doi:10.1038/s41467-018-03856-y
- Suzuki A, Terao C, Yamamoto K. Linking of genetic risk variants to disease-specific gene expression via multi-omics studies in rheumatoid arthritis. Semin Arthritis Rheum. 2019;49(3s):S49–S53. doi:10.1016/j.semarthrit.2019.09.007
- Guo H, Liu J, Ben Q, et al. The aspirin-induced long non-coding RNA OLA1P2 blocks phosphorylated STAT3 homodimer formation. Genome Biol. 2016;17:24. doi:10.1186/s13059-016-0892-5
- Khvorova A, Watts JK. The chemical evolution of oligonucleotide therapies of clinical utility. Nat Biotechnol. 2017;35(3):238–248. doi:10.1038/nbt.3765
- van Heesch S, van Iterson M, Jacobi J, et al. Extensive localization of long noncoding RNAs to the cytosol and mono- and polyribosomal complexes. Genome Biol. 2014;15(1):R6. doi:10.1186/gb-2014-15-1-r6