Abstract
Purpose
Obesity is a disease with genetic susceptibility characterized by an increase in storage and irregular distribution of body fat. In obese patients, the decrease in the Adiponectin gene (ADIPOQ) expression has been associated with a systemic low-grade inflammatory state. Our aim was to investigate the relationship between ADIPOQ +45T>G gene simple nucleotide polymorphism (SNP rs2241766) with serum adiponectin (sAdiponectin), distribution of body fat storage, and inflammation markers.
Subjects and methods
In this cross-sectional study, 242 individuals from Western Mexico characterized as Mexican-Mestizo and classified by body mass index (BMI), were included. Anthropometrics, body composition, body fat distribution, and inflammation markers were measured by routine methods. Genotypes were characterized using the polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) technique and sAdiponectin by the ELISA method. A P-value <0.05 was considered the statistically significant threshold.
Results
sAdiponectin is associated with BMI (P < 0.001) and the genotypes (P < 0.001 to 0.0046) GG (8169 ± 1162 ng/mL), TG (5189 ± 501 ng/mL), and TT (3741 ± 323 ng/mL), but the SNP ADIPOQ +45T>G is not associated with BMI. However, the detailed analysis showed association of this SNP with a pattern of fat distribution and correlations (P < 0.05) with inflammation markers and distribution of body fat storage (Pearson’s r = −0.169 to −0.465) were found.
Conclusion
In this study, we have suggested that the ADIPOQ +45G allele could be associated with distribution of body fat storage in obesity. On the other hand, as no association was observed between ADIPOQ +45T>G gene polymorphism and obesity, it cannot be concluded that the ADIPOQ +45G allele is responsible for the increase of adiponectin levels.
Introduction
Obesity is a systemic and chronic disease with genetic susceptibility.Citation1,Citation2 It is accompanied by metabolic changes that predispose the presentation of disorders in the health status and is associated in most cases with endocrine, cardiovascular, and orthopedic disorders relating to diverse biological, sociocultural, and psychological factors. In Mexico, the combined prevalence of overweight and obesity in adults is 70.9%.Citation3,Citation4
Obesity is characterized by an increase of the body mass index (BMI) and disproportion of body composition leading to a rise of fat mass.Citation5,Citation6 This increase, represented by an expansion of white adipose tissue (WAT) with an irregular distribution, is the most consistent pathological process in obesity. This WAT increase also promotes a unique microenvironment with a persistent abnormal inflammatory response that leads to a systemic low-grade subacute inflammatory state.Citation7–Citation9
An early phase of this inflammatory response occurs when the adipocytes and infiltrated macrophages (M1) in the WAT increase the release of inflammation markers and decrease adiponectin.Citation10 Adiponectin belongs to the adipokine family and is secreted almost exclusively by adipose tissue. Its function is to maintain stability in glucose homeostasis and insulin sensitivity,Citation11 and is down-regulated in subjects with obesity related comorbidity. Low serum levels of adiponectin (sAdiponectin) were associated with unfavorable serum lipid profiles and high inflammatory markers.Citation12
On the other hand, the susceptibility for development of obesity with an irregular accumulation of WAT in the body depends on the interaction of two factors: (1) exposure to environmental risk factors, and (2) variation in genes (characterized as “obesogenes”) involved in body weight regulation and obesity pathogenesis such as ADIPOQ.Citation1,Citation2 The ADIPOQ gene, linked to the regulation and expansion of WAT, has been investigated as a possible factor associated with obesity and a synonymous (15Gly > Gly) single nucleotide polymorphism (SNP) +45T > G in the exon 2 was identified. The genotype frequencies of this polymorphism show ethnic variation and the +45G polymorphic allele has been associated with insulin resistance and type 2 diabetes (T2D) in several populations;Citation13,Citation14 however, their functional effects are undefined.
In this context, we investigated the distribution of ADIPOQ +45T>G polymorphism in a Mexican-Mestizo population from Western Mexico. This study evaluated the probable association of ADIPOQ genetic variants with blood levels of adiponectin, inflammation markers, and the distribution of body fat storage in obesity.
Subjects and methods
Subjects
In this cross-sectional study, beginning February 2009 and ending January 2011, we included a total of 242 nonrelated adults, aged 24 to 69 years, recruited from a population of Western Mexico. Individuals were classified according to BMI, with World Health Organization (WHO) categories, as (1) overweight: BMI of 25.0 to 29.9 kg/m2, obese: BMI > 30.0 kg/m2, lean: BMI of 18.5 to 24.9 kg/m2; and (2) individuals with excess body weight BMI > 25.0 kg/m2.Citation15
All individuals were Mexican-Mestizos, in accordance with the National Institute of Anthropology classification: they were born in Mexico, with a family last name of Spanish origin, and a family history of Mexican ancestors for at least three generations.Citation16
The group was composed of healthy adult volunteers (ie, without medication and glucose intolerance according to clinical history, and with a stable weight for at least three weeks); individuals with infectious diseases, hypertension, history of cardiovascular disease, malignancy, or renal and metabolic diseases such as T2D, were not included. Before enrollment in this study, informed participants gave their signature in a consent document, in keeping with the Helsinki declaration guidelinesCitation17 and the Institutional (University of Guadalajara) Review Committees to ensure appropriate ethical and biosecurity conduct (record No 70/ UG-JAL/2011).
Methods
According to the inclusion criteria, all subjects had a complete medical history. Distribution of body fat storage measurements included: height, which was measured to the nearest 1 mm by using a stadiometer (Seca GmbH and Co, KG, Hamburg, Germany), weight and body composition (that included total muscle and fat mass and body fat percentage), which were measured by using bio-electrical impedance analysis (TBF-304; Tokyo, Japan) to the nearest 0.1 kg, and BMI according to Lukaski’s technique.Citation18,Citation19 Waist, hip, and arm circumferences were measured using an anthropometric fiberglass tape (GULICK®, length 0–180 cm, accuracy ± 1 mm; North Coast Medical, Inc., Gilroy, CA) and following the procedures recommended by the anthropometric indicators measurement guide.Citation20–Citation22 Four measures of skinfold thickness (ie, biceps, triceps, subscapular, and suprailiac) were obtained on the right side of the body by using a caliper skinfold Harpenden (opening 80 mm and precision of ±0.2 mm, constant pressure of 10 g/mm2; Holtain Ltd, Crosswell, UK) in accordance with the procedures recommended by Ness-Abramof and Apovian,Citation22 Durnin and Rahaman,Citation23 and Lukaski.Citation24 We calculated the sum of the four skinfold thicknesses as an indicator of subcutaneous fatness, body fat ratio as an indicator of adiposity, and waist-hip ratio (WHR) as an indicator of preferential accumulation of fat in the abdomen rather than on extremities.Citation22,Citation25
We confirmed an overnight fast of 12 hours in all subjects, and obtained venous blood samples. After allowing these to clot at room temperature they were centrifuged at 1509 RCF (Rotanta 460R; Andreas Hettich GmbH and Co, KG, Tuttlingen, Germany) for 10 minutes and serum samples were separated and stored at −70°C until analysis. Serum concentrations of sAdiponectin were determined using a commercial kit of enzyme-linked immune-absorbent assay (R&D Systems Inc, Minneapolis, MN), with a limit of detection of 1 ng/mL. The profile of inflammation markers included high-sensitivity C reactive protein (CRP) with a limit of detection of 0.15 mg/L (Random Access Analyzer Clinical Chemistry-Turbidimetry A25 Immunochemistry; Biosystems, Barcelona, Spain); erythrocyte sedimentation rate was measured by the Wintrobe method; and a complete blood count, including platelet count and white blood cell count, were measured with the Cell-Dyn 3700 (Abbott Diagnostics, Abbott Park, IL).
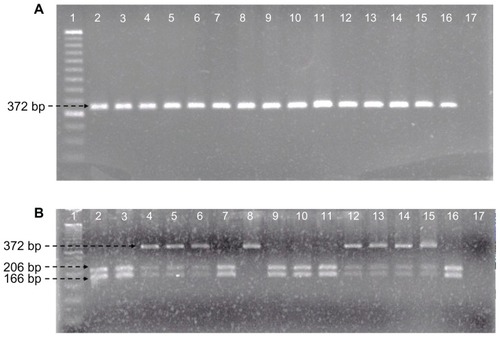
To identify ADIPOQ +45T>G gene polymorphism, genomic DNA was obtained from total blood using a standard protocol for extraction with the modified Miller methodCitation26 and was stored at −20°C until use for genotyping. The polymorphic region was amplified by the polymerase chain reaction (PCR) methodCitation27 to analyze the SNP +45T>G in ADIPOQ (rs2241766) in the locus 3q27. Primers were: forward 5′-GCAGCTCCTAGAAGTAGACTCTGCTG-3′, reverse 5′-GGAGGTCTGTGATGAAAGAGGCC-3′, and annealing temperature of 50°C. The PCR product was 372 base pairs (bp, ); and was performed in a 25 μL total volume mixture (100 ng of DNA, 2 nM of each primer, 0.20 mM of each dNTP, 2.5 mM MgCl2, 0.25 U Taq polymerase, and 1X PCR buffer, Invitrogen®); followed by digestion of PCR products with Bsp HI restriction enzyme (New England Biolabs© Inc, Ipswich, MA). The following digestion fragments were obtained: allele T, 206, and 166 bp; allele G, 372 bp (). Electrophoresis was performed at a constant voltage of 80 volts on 3% agarose gels stained with ethidium bromide 0.01 mg. For quality control, a blank and samples previously confirmed as positive for each genotype were used as controls. To ensure the accuracy of genotype data, we used internal controls and repetitive experiments. Any sample that yielded a weak signal was repeated. In addition, all samples were repeated at random to verify the reproducibility with positive controls in each experiment and we confirmed that the positive controls were genotyped correctly. The genotyping success rate was 100%.
Figure 1 Identification of ADIPOQ +45T>G polymorphism. (A) PCR amplification fragments. Line 1, molecular weight marker of 50 bp; lines 2–16, samples; line 17, negative control. (B) Bsp HI restriction fragments. Line 1, molecular weight marker of 50 bp; lines 2, 3, 7, 9–11, and 16, TT genotype; lines 4–6 and 12–15, TG genotype; line 8, GG genotype; line 17, negative control.
Statistical analysis
Data were analyzed with the PASW Statistics program (version 18.0; IBM Corporation, Armonk, NY). Results are given as percentages or mean ± standard deviation or standard error of the mean. The distribution of body fat storage measurements, sAdiponectin levels, and profile of inflammation markers were analyzed with the unpaired Student’s t-test, one-way ANOVA, analysis of covariance, the multifactorial ANOVA model corrected by sex among carriers of the three genotypes, and the additive model with the post hoc Bonferroni test.
Data from serum concentrations of sAdiponectin, inflammation markers, and distribution of body fat storage variables were subjected to Pearson correlation tests. After analysis we performed a multiple linear regression analysis with the positive correlation variables. The test for Hardy– Weinberg equilibrium for ADIPOQ loci was performed. Contingency tables with χ2 trend test or Fisher’s exact test, as appropriate, were used for testing the differences of genotype distribution and allele frequencies between all groups. Two models were used for these analyses: (1) the dominant model where the SNP was modeled categorically and separated into three categories, one for each genotype, and (2) the additive model where the SNP was modeled into two categories with two genotypes combined into one category (TG + GG) choosing one genotype (TT) as the reference group for both models. A P-value less than 0.05 was considered statistically significant.
Results
In our study, we found an overweight and obesity prevalence of 40.4% and 23.8%, respectively. shows data of anthropometrics measurements in our study group. The main anthropometric characteristics in females were higher percentage of body fat mass and skinfold thicknesses and lower total muscle mass and WHR than males.
Table 1 Demographics and distribution of body fat mass in study group according to WHO BMI categories
Intergroup comparison according to BMI categories (lean versus overweight versus obesity) displayed higher levels of CRP and erythrocyte sedimentation rate in female than male individuals (). While sAdiponectin levels were higher in the lean group than the overweight and obesity groups (), this difference was also represented in the males (). A negative correlation was found between sAdiponectin with inflammation markers and distribution of body fat storage. We performed a multiple linear regression analysis and found that 10.6% of the variation of sAdiponectin is explained by fat percentage, age, and sex (P = 0.001). Other correlations of inflammation markers with adiposity are shown in .
Figure 2 Soluble levels of sAdiponectin. Study group (A); sex-specific study group (B). Group comparisons between genotypes of ADIPOQ +45T>G polymorphism (C), which included: (1) the dominant model modeled into three categories, with TT genotype chosen as the reference group; and (2) the additive model modeled into two categories with TT genotype as the reference group and the other two genotypes combined into one category (TG + GG).
Abbreviations: sAdiponectin, serum adiponectin; SEM, standard error of the mean.
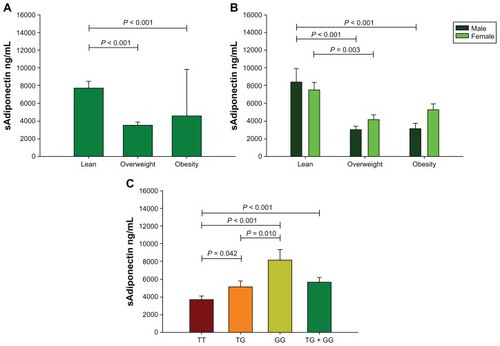
Table 2 Inflammation markers in study group according to WHO BMI categories
Table 3 sAdiponectin correlations with inflammation markers and distribution of body fat mass in study group
In a different analysis of ADIPOQ +45T>G SNP we found that the independent segregation of the alleles tested in this Mexican-Mestizo group kept with the Hardy–Weinberg equilibrium (P = 0.377). The association of the genotypes and alleles with overweight and obesity was analyzed: no significant association was found between lean, overweight, obesity, or BMI > 25.0 kg/m2 individuals ().
Table 4 Distribution of ADIPOQ +45T>G gene polymorphism in Mexican-Mestizo population
We detected different sAdiponectin levels in genotype carriers as follows: GG (8169 ± 1162 ng/mL), TG (5189 ± 501 ng/mL), and TT (3741 ± 323 ng/mL) (). In TG genotype carriers we detected lower measures of hip circumference and biceps skinfold thickness than TT genotype carriers. However, in the additive model the TG + GG genotype carriers showed lower measures of total body fat mass, hip circumference, biceps and triceps skinfold thicknesses, and sum of four skinfold thicknesses than the TT genotype carriers (). Additionally, the females carrying the G allele (additive model) with an excess of body weight (BMI > 25.0 kg/m2) had low measures of total body fat mass, hip circumference, arm circumference, and skin fold thickness and a high WHR as compared to TT genotype carriers with similar BMI. In lean female individuals carrying the G allele we observed a lower BMI and body fat ratio than TT genotype carriers ().
Table 5 Comparison of ADIPOQ +45T>G gene polymorphism with distribution and storage of body fat
Table 6 Comparisons of the distribution of body fat mass between females ADIPOQ +45T>G genotype carriers
Finally, we independently analyzed the female obesity group (BMI > 30 kg/m2), finding the following differences: (1) body fat mass (P = 0.009) in carriers of TT genotype (43.8% ± 3.09%) was lower as compared to GG genotype carriers (37.5% ± 1.65%), and (2) WHR (P = 0.005) in carriers of TT genotype (0.8625 ± 0.07) was lower as compared to carriers of TG genotype (0.9711 ± 0.11).
Discussion
We investigated the distribution of ADIPOQ +45T>G polymorphism in a Mexican-Mestizo population from Western Mexico. This study evaluated the probable association of ADIPOQ genetic polymorphism with blood levels of adiponectin, inflammation markers, and the distribution of body fat storage in obesity. We found a high prevalence of overweight (40.4%) and obesity (23.8%) in our study group, classified by the BMI according to WHO categories. These findings are in accordance with those reported by the National Survey of Health and Nutrition 2006 for Western Mexico. Obesity is considered a public health problem in Mexico, worsened in the early 1990s. The National Institute of Public Health reported a 47% increase in the prevalence of obesity in adults (older than 20 years) from 1994 through 2006. This increased tendency is not exclusive to Mexico, mainly because obesity is considered a pandemic disease.Citation5,Citation28
The difference between overweight and obese individuals for the anthropometric measurements and body composition, with the exception of the BMI and arm circumference, suggests the following: (1) physiologic factors are involved in obesity development, and (2) there does not exist an equal distribution of body fat storage during the transition from overweight to obesity.
In Mexico, the National Department of Health established a cut-off value of the waist circumference measurement (80.0 cm for women and 90.0 cm for men) as a clinical criterion to estimate the risk of comorbidities in obesity; in this sense, all individuals with obesity in our study group had abdominal obesity, a state in which IL-6 production increases. This finding is important because the increase of IL-6 is a key inductor of the hepatic synthesis of CRP.Citation10,Citation29 Our CRP data coincides with previous reports where it was established that obesity is characterized by a systemic low-grade subacute inflammatory state,Citation7 also considered the clinical interphase between obesity and the appearance of multiple illnesses.
The present study shows that when sAdiponectin levels have decreased the BMI and body fat storage increase. Our data are consistent with previous findings: low sAdiponectin levels have been observed in overweight, obesity, and comorbidities.Citation12,Citation30
Low levels of sAdiponectin reflected the adiposity in our groups of study; this finding is underlined because we identified a negative correlation between sAdiponectin with body fat storage and inflammation markers. These findings therefore suggest that the relationship between low sAdiponectin concentrations and obesity may be in part mediated by the accumulation of adipose tissue. The possible explanation is that sAdiponectin is an exclusive product of the adipose tissue.
On the other hand, controversial results have been reported for ADIPOQ gene polymorphism in different populations. ADIPOQ +45T>G polymorphism has been associated with obesity, insulin resistance, metabolic syndrome, T2D, and variation in BMI and sAdiponectin in Chinese,Citation31,Citation32 Korean,Citation33,Citation34 Japanese,Citation27,Citation35 Canadian,Citation25 Spanish,Citation36 Iranian,Citation37 Finnish,Citation38 and GreekCitation39,Citation40 populations, while in Taiwanese,Citation41 French,Citation42 Swedish Caucasian,Citation43 Czech,Citation44 Malaysian,Citation45 Pima Indian,Citation14 and ItalianCitation46,Citation47 populations, there has been no association.Citation45
Significant associations between obesity and polymorphisms in the gene coding for ADIPOQ have been reported in other studies (). The genetic contribution to susceptibility of obesity is well accepted, although, in our study we did not find an association of the ADIPOQ +45T>G gene polymorphism with obesity, but we found in the additive model that the TT genotype carriers had a higher total body fat mass and biceps and triceps skinfold thickness than the TG + GG genotype carriers which also had higher sAdiponectin levels. The distribution of body fat storage can play different roles in adiponectin production and/or function during the maintenance of the inflammatory process in obesity. As a result of this association of the polymorphic allele with distribution of body fat storage, we suggest that genetic variability in the ADIPOQ gene may modulate the levels of sAdiponectin. However, a potential limitation of our study is the possibility that other variants in other ADIPOQ loci than in those we studied may play a role in obesity. Nevertheless, we chose +45T>G polymorphism that has been implicated functionally and associated with obesity comorbidities in previous studies. A limitation regarding the association analysis of our study is that our sample of only 242 individuals may reduce the ability to detect associations.
Table 7 Comparison of the ADIPOQ +45T>G gene polymorphism distribution in Mexican-Mestizo population with other studies
Conclusion
As the end point of this study, we have suggested that the ADIPOQ +45G allele could be associated with the distribution of body fat storage in obesity. On the other hand, as no association was observed between the ADIPOQ +45T>G gene polymorphism and obesity it cannot be concluded if the ADIPOQ +45G allele is responsible for the increase of adiponectin levels.
Acknowledgments
This work was supported by grant number PS-2009-552 to Rosa Elena Navarro of the State Council of Science and Technology (COECyTJal-University of Guadalajara). We thank Michele Brennan, MSc in Clinical Chemistry, for the review of the English language editing.
Disclosure
The authors report no conflicts of interest in this work.
References
- Blakemore AI Froguel P Is obesity our genetic legacy? J Clin Endocrinol Metab 2008 93 11 Suppl 1 S51 S56 18987270
- Das UN Obesity: genes, brain, gut, and environment Nutrition 2010 26 5 459 473 20022465
- Shamah-Levy TV-HS Rivera-Dommarco JA Nutrition Results Públic INdS National Health and Nutrition Survey, 2006 Cuernavaca, Mexico National Institute of Public Health 2007 Spanish
- Olaiz-Fernández G Rivera-Dommarco J Shamah-Levy T National Health and Nutrition Survey, 2006 Cuernavaca, México National Institute of Public Health 2006 Spanish
- Dixon JB The effect of obesity on health outcomes Mol Cell Endocrinol 2010 316 2 104 108 19628019
- Mutch DM Clément K Genetics of human obesity Best Pract Res Clin Endocrinol Metab 2006 20 4 647 664 17161337
- Mathis D Shoelson SE Immunometabolism: an emerging frontier Nat Rev Immunol 2011 11 2 81 21469396
- Galic S Oakhill JS Steinberg GR Adipose tissue as an endocrine organ Mol Cell Endocrinol 2010 316 2 129 139 19723556
- Poulos SP Hausman DB Hausman GJ The development and endocrine functions of adipose tissue Mol Cell Endocrinol 2010 323 1 20 34 20025936
- Ouchi N Parker JL Lugus JJ Walsh K Adipokines in inflammation and metabolic disease Nat Rev Immunol 2011 11 2 85 97 21252989
- Guerre-Millo M Adiponectin: an update Diabetes Metab 2008 34 1 12 18 18069030
- Chang LC Huang KC Wu YW The clinical implications of blood adiponectin in cardiometabolic disorders J Formos Med Assoc 2009 108 5 353 366 19443289
- Woo JG Dolan LM Deka R Interactions between noncontiguous haplotypes in the adiponectin gene ACDC are associated with plasma adiponectin Diabetes 2006 55 2 523 529 16443790
- Vozarova de Courten B Hanson RL Funahashi T Common polymorphisms in the adiponectin gene acdc are not associated with diabetes in Pima Indians Diabetes 2005 54 1 284 289 15616040
- The International Classification of adult underweight, overweight and obesity according to BMI WHO Technical Report Series Geneva World Health Organization 2004
- Gorodezky C Alaez C Vázquez-García MN The genetic structure of Mexican Mestizos of different locations: tracking back their origins through MHC genes, blood group systems, and microsatellites Hum Immunol 2001 62 9 979 991 11543900
- World Medical Association Declaration of Helsinki: Ethical Principles for Medical Research Involving Human Subjects (revised October 2008). Available from: http://www.wma.net/en/30publications/10policies/b3/ Accessed January 8, 2009
- Lukaski HC Body mass index, bioelectrical impedance, and body composition Nutrition 2001 17 1 55 56 11165891
- Lukaski HC Regional bioelectrical impedance analysis: applications in health and medicine Acta Diabetol 2003 40 Suppl 1 S196 S199 14618471
- van der Ploeg GE Gunn SM Withers RT Modra AC Use of anthropometric variables to predict relative body fat determined by a four-compartment body composition model Eur J Clin Nutr 2003 57 8 1009 1016 12879096
- Durnin JV Womersley J Body fat assessed from total body density and its estimation from skinfold thickness: measurements on 481 men and women aged from 16 to 72 years Br J Nutr 1974 32 1 77 97 4843734
- Ness-Abramof R Apovian CM Waist circumference measurement in clinical practice Nutr Clin Pract 2008 23 4 397 404 18682591
- Durnin JV Rahaman MM The assessment of the amount of fat in the human body from measurements of skinfold thickness Br J Nutr 1967 21 3 681 689 6052883
- Lukaski HC Methods for the assessment of human body composition: traditional and new Am J Clin Nutr 1987 46 4 537 556 3310598
- Loos RJ Ruchat S Rankinen T Tremblay A Pérusse L Bouchard C Adiponectin and adiponectin receptor gene variants in relation to resting metabolic rate, respiratory quotient, and adiposity-related phenotypes in the Quebec Family Study Am J Clin Nutr 2007 85 1 26 34 17209173
- Miller DN Bryant JE Madsen EL Ghiorse WC Evaluation and optimization of DNA extraction and purification procedures for soil and sediment samples Appl Environ Microbiol 1999 65 11 4715 4724 10543776
- Katsuda Y Asano A Murase Y Association of genetic variation of the adiponectin gene with body fat distribution and carotid atherosclerosis in Japanese obese subjects J Atheroscler Thromb 2007 14 1 19 26 17332688
- Rokholm B Baker JL Sørensen TI The levelling off of the obesity epidemic since the year 1999 – a review of evidence and perspectives Obes Rev 2010 11 12 835 846 20973911
- Hodge AM Maple-Brown L Cunningham J Abdominal obesity and other risk factors largely explain the high CRP in indigenous Australians relative to the general population, but not gender differences: a cross-sectional study BMC Public Health 2010 10 700 21078191
- Staiger H Tschritter O Machann J Relationship of serum adiponectin and leptin concentrations with body fat distribution in humans Obes Res 2003 11 3 368 372 12634431
- Li LL Kang XL Ran XJ Associations between 45T/G polymorphism of the adiponectin gene and plasma adiponectin levels with type 2 diabetes Clin Exp Pharmacol Physiol 2007 34 12 1287 1290 17973869
- Tso AW Sham PC Wat NM Polymorphisms of the gene encoding adiponectin and glycaemic outcome of Chinese subjects with impaired glucose tolerance: a 5-year follow-up study Diabetologia 2006 49 8 1806 1815 16788799
- Park JW Park J Jee SH ADIPOQ gene variants associated with susceptibility to obesity and low serum adiponectin levels in healthy Koreans Epidemiol Health 2011 33 e2011003 21603224
- Jang Y Lee JH Kim OY The SNP276G > T polymorphism in the adiponectin (ACDC) gene is more strongly associated with insulin resistance and cardiovascular disease risk than SNP45T > G in nonobese/nondiabetic Korean men independent of abdominal adiposity and circulating plasma adiponectin Metabolism 2006 55 1 59 66 16324920
- Vendramini MF Pereira AC Ferreira SR Kasamatsu TS Moisés RS Japanese Brazilian Diabetes Study Group Association of genetic variants in the adiponectin encoding gene (ADIPOQ) with type 2 diabetes in Japanese Brazilians J Diabetes Complications 2010 24 2 115 120 19269196
- González-Sánchez JL Martínez-Calatrava MJ Martinez-Larrad MT Interaction of the −308G/A promoter polymorphism of the tumor necrosis factor-alpha gene with single-nucleotide polymorphism 45 of the adiponectin gene: effect on serum adiponectin concentrations in a Spanish population Clin Chem 2006 52 1 97 103 16254197
- Mohammadzadeh G Zarghami N Associations between single-nucleotide polymorphisms of the adiponectin gene, serum adiponectin levels and increased risk of type 2 diabetes mellitus in Iranian obese individuals Scand J Clin Lab Invest 2009 69 7 764 771 19929719
- Siitonen N Pulkkinen L Lindström J Association of ADIPOQ gene variants with body weight, type 2 diabetes and serum adiponectin concentrations: the Finnish Diabetes Prevention Study BMC Med Genet 2011 12 5 21219602
- Melistas L Mantzoros CS Kontogianni M Antonopoulou S Ordovas JM Yiannakouris N Association of the +45T > G and +276G > T polymorphisms in the adiponectin gene with insulin resistance in nondiabetic Greek women Eur J Endocrinol 2009 161 6 845 852 19755407
- Xita N Georgiou I Chatzikyriakidou A Effect of adiponectin gene polymorphisms on circulating adiponectin and insulin resistance indexes in women with polycystic ovary syndrome Clin Chem 2005 51 2 416 423 15590747
- Leu HB Chung CM Lin SJ Jong YS Pan WH Chen JW Adiponectin gene polymorphism is selectively associated with the concomitant presence of metabolic syndrome and essential hypertension PloS one 2011 6 5 e19999 21637762
- Vasseur F Helbecque N Dina C Single-nucleotide polymorphism haplotypes in the both proximal promoter and exon 3 of the APM1 gene modulate adipocyte-secreted adiponectin hormone levels and contribute to the genetic risk for type 2 diabetes in French Caucasians Hum Mol Genet 2002 11 21 2607 2614 12354786
- Gu HF Abulaiti A Ostenson CG Single nucleotide polymorphisms in the proximal promoter region of the adiponectin (APM1) gene are associated with type 2 diabetes in Swedish caucasians Diabetes 2004 53 Suppl 1 S31 S35 14749263
- Bienertova-Vasku J Bienert P Hlinomaz O Vasku A Common polymorphism +45T/G in adiponectin gene as potential modulator of in-stent restenosis development Int J Cardiol 2010 145 2 351 20045207
- Lau CH Muniandy S Adiponectin and resistin gene polymorphisms in association with their respective adipokine levels Ann Hum Genet 2011 75 3 370 382 21323646
- Menzaghi C Ercolino T Salvemini L Multigenic control of serum adiponectin levels: evidence for a role of the APM1 gene and a locus on 14q13 Physiological Genomics 2004 19 2 170 174 15252189
- Filippi E Sentinelli F Trischitta V Association of the human adiponectin gene and insulin resistance Eur J Hum Genet 2004 12 3 199 205 14673476