
Abstract
Purpose
Carbapenemases-producing Klebsiella pneumoniae are challenging antimicrobial therapy of hospitalised patients, which is further complicated by colistin resistance. This study describes molecular epidemiological insights into colistin-resistant and carbapenemases-producing clinical K. pneumoniae.
Patients and methods
Cultures collected from 26 hospitalised patients during 2014–2017 in the main hospital in Molise Region, central Italy, were characterized. The minimum inhibitory concentration for 19 antibiotics was determined, including carbapenems and colistin. Prevalence of resistance-associated genes was investigated through PCR, detecting blaKPC, blaGES, blaVIM, blaIMP, blaNDM, blaOXA-48, blaCTX-M, blaTEM, blaSHV, and mcr-1,2,3,4,5,6,7,8. The mgrB gene was also analysed in colistin-resistant strains by PCR and sequencing assays. K. pneumoniae were typed by pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST).
Results
Twenty out of 26 K. pneumoniae were phenotypically resistant to carbapenems and 19 were resistant to colistin. All isolates harbored blaKPC, and blaSHV, blaTEM and blaVIM were further the most common resistance-associated genes. In colistin-resistant strains, mcr-1,2,3,4,5,6,7,8 variants were not detected, while mutations and insertion elements in mgrB were observed in 68.4% (n=13) in 31.6% (n=6) isolates, respectively. PFGE revealed 12 clusters and 18 pulsotypes at 85% and 95% cut-off, while the Sequence Types ST512 (n=13, 50%), ST101 (n=10, 38.5%), ST307 (n=2, 7.7%) plus a novel ST were detected using MLST.
Conclusion
All K. pneumoniae showed a multidrug-resistant phenotype, particularly to carbapenems and colistin. According to national data, blaKPC was the prevailing carbapenemase, followed by blaVIM, while blaTEM and blaSHV were among the most frequent beta-lactamases. Consistent with previous reports in Italy, ST512 was the most common clone, particularly during 2014–15, whilst ST101 became dominant in 2016–17. Colistin resistance was mainly associated with deleterious mutations and transposon in the mgrB gene. Improvements of surveillance, compliance with infection prevention procedures and antimicrobial stewardship are essential to limit the spread of resistant K. pneumoniae.
Introduction
Klebsiella pneumoniae is the most clinically relevant Klebsiella species.Citation1 The 2011–2012 Point Prevalence study of the European Centre for Disease Prevention and Control identified K. pneumoniae causing 6.8% of hospital-acquired infections (HAIs), which represented the second most frequent Enterobacteriaceae after Escherichia coli.Citation2 In 2005, almost all European regions were carbapenem-resistant K. pneumoniae (CR-Kp) free. From 2005 to 2015, CR-Kp has emerged in several countries, reaching rates of 40–60%.Citation3 In the TOTEM study, CR-Kp was classified as the most critical antimicrobially resistant pathogen and a leading cause of nosocomial infections, mainly in intensive care units (ICUs).Citation4 Furthermore, among 5331 bacteremia cases due to carbapenemase-producing Enterobacteriaceae, 96.8% was attributed to K. pneumoniae.Citation5 At EU/EEA level, in 2017, 34% of K. pneumoniae notified to the European Antimicrobial Resistance Surveillance Network were resistant to at least one of the antimicrobial groups under surveillance, including carbapenems.Citation6
Carbapenemases production, particularly KPC, represents the most prevalent mechanism for carbapenems resistance in K. pneumoniae,Citation5,Citation7 but the carbapenemases GES, NDM, IMP, VIM, and OXA-48 can also be involved.Citation8 K. pneumoniae carrying the gene blaKPC are endemic in Italy since nearly 90% of CR-Kp are KPC producers, followed by blaVIM (9.2%) and blaOXA-48 (1.3%).Citation3,Citation9,Citation10 In K. pneumoniae, extended spectrum beta-lactamases (ESBLs) have been also detected, being involved in oximino-cephalosporin resistance and able to hydrolyze beta-lactams.Citation11 During 1990–2000, K. pneumoniae has become the major ESBL-carrying pathogen in hospital outbreaks, mostly carrying blaTEM and blaSHV.Citation12
The increasing prevalence of multidrug-resistant (MDR) Gram-negative bacteria has led to re-introduction of colistin, especially for infections sustained by K. pneumoniae.Citation4 Nevertheless, in the last years, colistin resistance has also emerged in CR-Kp with rates as high as 36%.Citation3,Citation13,Citation14 In this case, resistance is due to structural modifications of lipopolysaccharide (LPS) that is the target for colistin. Resistance could be attributed to mgrB inactivation by down-regulation of the Pmr system involved in LPS modification, neutralizing the negative charge and decreasing colistin binding,Citation15,Citation16 or through plasmid-mediated mcr (mcr-1,2,3,4,5,6,7,8 variants).Citation17–Citation23
Management of CR-Kp infections is associated with long hospitalizations and poor outcomesCitation1,Citation14 and complicated by MDR emergence, which severely limits antimicrobial treatment options.Citation14 Since K. pneumoniae is among the most frequent agents in nosocomial settings,Citation8 identification of outbreaks due to MDR strains is crucial. In this scenario, molecular typing enabling strains comparison is required for detecting epidemics and tracking infection sources and factors involved in the transmission.Citation24 For K. pneumoniae molecular characterization, the foremost approaches rely on pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST) systems.Citation25
In this study, clinical K. pneumoniae isolated in the main hospital in Molise Region, central Italy, were characterized to evaluate MDR patterns, genetic differences and relationships, and prevalence of carbapenem resistance determinants, as well as to elucidate the mechanisms involved in colistin resistance.
Materials and Methods
K. pneumoniae Cultures and DNA Extraction
Twenty-six K. pneumoniae cultures isolated within the “Alert Organism” surveillance system during 2014–2017 were collected from the main hospital for acute care in Molise Region, central Italy. The hospital at the time of the study had a total of 336 beds, 320 for acute care and 16 for ICU.Citation26 Additionally, there were 19 total wards: ten of medicine and surgery specialties, four of pediatrics and two of ICU specialties, followed by single wards of gynecology/obstetrics, geriatrics, psychiatry, rehabilitation, and mixed specialties.
The selection criteria for the tested strains were non-replicates cultures, and a KPC phenotype evaluated with the Matrix-Assisted Laser Desorption Ionization-Time-Of-Flight Mass Spectrometry (MALDI-TOF) assay, as reported by the hospital laboratory. K. pneumoniae cultures were mostly recovered from aspirated bronchial (n=9, 35.0%) samples, urine (n=6, 23%), rectal swab (n=4, 15.4%), and blood cultures (n=3, 11.5%) (). Fifteen (57.7%) isolates were from patients admitted to ICU: 60% male, overall mean age 73±12.6 years (median 78.5, range 44–89 years). Clinical specimens were cultured and purified on McConkey agar plates (Biolife, Milan, Italy) incubated at 37°C overnight. DNA was extracted using Maxwell® 16 Cell DNA Purification Kit (Promega, Milan, Italy).
Table 1 Patient's Clinical Data and Characteristics of Analyzed Strains
Antimicrobial Susceptibility Testing
The susceptibility to nineteen antimicrobials was evaluated by the hospital Microbiology laboratory using BD Phoenix™ Automated Microbiology System (Becton Dickinson Diagnostic Systems, Sparks, United States). The minimum inhibitory concentrations (MICs) were calculated for imipenem, ertapenem and meropenem (carbapenems); ampicillin, amoxicillin-clavulanate, piperacillin and piperacillin-tazobactam; ceftazidime, cefuroxime and cefotaxime; amikacin and gentamicin; ciprofloxacin and levofloxacin; fosfomycin, trimethoprim-sulfamethoxazole, tigecycline, and tobramycin. The microdilution method was used for colistin (polymyxin) MIC determination. Results were interpreted according to the European Committee on Antimicrobial Susceptibility Testing breakpoints.Citation27
Detection of Resistance-Associated Genes
Genes involved in carbapenems resistance were detected through single PCR assays, targeting blaKPC, blaGES, blaIMP, blaVIM, blaOXA-48 and blaNDM-1 genes.Citation28,Citation29 Amplifications were performed in 25 μL volume with 2 μL DNA template, 1X PCR Master Mix (Promega Corporation) and 1 µM of each primer. Target genes were amplified at specific conditions: 94°C 2 mins; 35 cycles: 94°C 1 min, 45°C (blaIMP)/52°C (blaKPC)/54°C (blaGES)/56°C (blaVIM/OXA-48)/60°C (blaNDM-1) 40 sec, 72°C 1 min; 72°C 5 mins. PCR amplicons were electrophoretically separated (1.0–1.5% m/v concentration, 1X TAE buffer at 100 V for 1 hr), including 100 bp DNA ladder (Promega). Positive and negative control were used in each batch of reactions.
K. pneumoniae isolates were also screened for blaSHV, blaTEM, and blaCTX-M genes by Multiplex PCR assays, using previously described oligonucleotides and specific cycling conditions.Citation11 The amplified products were resolved by agarose gel electrophoresis (1.5% m/v concentration, 1X TAE buffer at 100 V for 1 hr) including a 100 bp DNA ladder (Promega) and controls in each batch of reactions.
Molecular Analysis of Colistin Resistance
The colistin-resistant (col-R) isolates were screened by singleplex PCRs for the presence of mcr-1,Citation17 mcr-2,Citation18 mcr-3,Citation19 mcr-4,Citation20 mcr-5,Citation21 mcr-6,Citation22 mcr-7,Citation23 mcr-7.1Citation23 and mcr-8.Citation22 Amplifications were performed in 25 μL volume using 2 μL DNA, 1X PCR master mix (Promega Corporation) and primers at 1 µM. Amplicons were characterized after agarose gel electrophoresis (1.5% m/v concentration, 1X TAE buffer at 100 V for 1 hr) including a 100 bp DNA ladder (Promega).
PCR analysis of mgrB was performed using mgrB_Ext_F and mgrB_Ext_R primers targeting mgrB coding sequence and some flanking regions, as previously reported.Citation13 Amplifications were carried out in 25 μL volume using 5 μL DNA template, 1X PCR master mix (Promega) and oligonucleotides at 2 µM.
Colistin-sensitive (col-S) strains were used as a negative control carrying wild-type mgrB. PCR products were characterised using agarose gel electrophoresis (1.5% m/v concentration, 1X TAE buffer at 100 V for 1 hr) with a 100 bp DNA ladder (Promega). Amplicons longer than the expected molecular weight (253 bp) suggested the presence of an Insertion Sequence (IS), and were analyzed by Sanger sequencing (Eurofins Genomics, Germany GmbH, Ebersberg, Germany), including col-S strains as control. Sequences were analyzed with Basic Local Alignment Search Tool (BLAST; blast.ncbi.nlm.nih.gov/Blast.cgi) and processed with BioEdit v7.0.5.
The nucleotide sequences of wild-type mgrB in KP25 and KP42 isolates (GenBank Accession numbers MN389772 and MN389773, respectively), as well as those of col-R strains without ISs (KP5, KP6, KP7, KP9, KP10, KP28, KP31, KP34, KP36, KP39, KP40, KP41 and KP43) were deposited at BankIt/GenBank (Accession numbers: MN389775, MN389774, MN389776, MN389777, MN389778, MN389779, MN389780, MN389781, MN389782, MN389783, MN389784, MN389785, and MN389786, respectively).
To translate DNA sequences, EMBOSS Transeq (https://www.ebi.ac.uk/Tools/st/emboss) tool was used. The amino acid sequences were analyzed with Protein Variation Effect Analyzer (PROVEAN, http://provean.jcvi.org/index.php) allowing prediction by algorithm of the functional impact for all classes of sequence variations.Citation30 The change in the alignment score was considered as a measure of change in similarity caused by variation and thus to protein functionality.
Molecular Typing by PFGE and MLST
For PFGE, bacterial DNA was digested with XbaI (Fermentas, Milan, Italy) according to PulseNet protocol using conditions of pulse times from 5 to 40 sec over 24 hrs at 6.0V/cm and at 14°C.Citation31 Pulsotypes were analyzed through BioNumerics software (Applied Maths, Sint-Martens-Latem, Belgium), and dendrograms were generated using Dice coefficient and unweighted pair group method with arithmetic mean (UPGMA).Citation9 The similarity band patterns interpretation was performed according to Tenover criteria,Citation24,Citation32 setting 85% and 95% similarity cut-off for identifying similar restriction patterns and clusters, respectively. A validated MLST scheme was used,Citation33 and PCR products were sequenced by Sanger method (Eurofins Genomics) to identify allelic profiles and assign the Sequence Type (ST). The allelic combination was analysed on Pasteur platform (http://bigsdb.pasteur.fr/perl/bigsdb/bigsdb.pl?db=pubmlst_klebsiella_seqdef_public).
Results
Antimicrobial Resistance Profiles in K. pneumoniae Cultures
K. pneumoniae cultures showed a multi-carbapenem-resistant phenotype, with all resistant to ertapenem, 22 (84%) to meropenem, and 20 (77%) to imipenem. Twenty isolates were resistant to all carbapenems tested. In addition, nineteen (73%) strains were col-R. No isolates were susceptible to ampicillin, amoxicillin-clavulanate, ceftazidime, ciprofloxacin, cefotaxime, cefuroxime, levofloxacin, piperacillin and piperacillin-tazobactam. Moreover, 81% (n=21) isolates showed resistance to tobramycin, while 16 (62%) were resistant to trimethoprim, 16 (61.5%) to amikacin, 8 (31%) to tigecycline, 5 to gentamicin, and 2 to fosfomycin.
Prevalence of Resistance-Associated Genes
All K. pneumoniae harbored blaKPC, and 69.2% (n=18) were blaVIM positive. A high proportion of isolates also carried ESBLs. The blaSHV and blaTEM were found in 96.2% (n=25) and 88.4% (n=23), respectively, and 84.6% (n=22) harbored both genes. The blaCTX-M was only found in two strains. None of the strains carried blaGES, blaNDM-1 or blaOXA-48.
Prevalence of mcr Variants and mgrB Analysis
None of the 19 col-R isolates showed plasmid mcr-1,2,3,4,5,6,7,8 variants. Colistin resistance was also investigated through mgrB analysis, and the initial evaluation using agarose gel analysis, considering that the expected amplicon for wild-type mgrB has a 253 bp size.Citation13 PCR products were sequenced to identify IS or mutations involved in colistin resistance.
Amplicons longer than the expected size were observed in six (31.6%) out of the 19 col-R isolates, and sequencing confirmed the presence of transposon. The most common insertion element detected in five cultures belonged to IS5-like family (1056 bp), while ISKpn14 element was found in KP11 (). Furthermore, KP8, KP12, KP14, KP18 and KP19 isolates with IS5-like elements were all grouped in the PFGE cluster VIII.
In col-R strains with 253 bp amplicon, an identical deletion ∆g19 causing frameshift mutation and premature MgrB termination was found in KP9 and KP10 (n=2, 10.5%) isolates; missense mutations t95→g translated into V32G were found in 42.1% (n=8; KP5, KP31, KP34, KP36, KP40, KP41 and KP43) isolates; missense mutations c62→a translated as T21N occurred in KP28, as well as missense mutation g60→a translated into W20Stop was found in KP31 ( and ). After open reading frame identification, sequence analysis with PROVEAN was reported in .
Table 2 PROVEAN Analysis of the Single Amminoacid Change of MgrB Protein
K. pneumoniae Molecular Epidemiology
PFGE revealed 12 clusters at 85% cut-off similarity (): cluster VIII was the most common, grouping 9 (34.6%) isolates, followed by cluster V with three isolates, and clusters III, IV, and VI, each including two cultures. Dendrogram analysis at 95% similarity revealed 18 pulsotypes (PTs), with PT12 as the prevalent (n=4 isolates, 15.3%), followed by PT1, 3, 8 and PT10, each associated with two strains. PFGE discriminatory power was of 96%, as calculated by Simpson’s Index of Diversity.Citation34
Figure 1 PFGE dendrogram (Dice coefficient) and MLST results for 26 clinical K. pneumoniae isolates.
Note: The new ST is highlighted in blue.
Abbreviation: ST, Sequence Type.
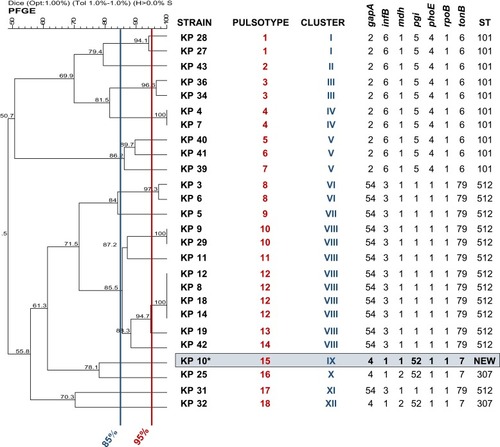
Figure 2 Antimicrobials resistance phenotypes and antimicrobial resistance genes profiles for 26 K. pneumoniae isolates.
Abbreviations: col-S, colistin-sensitive; Δg19, deletion of guanine in position 19; V32G, Valine in position 32 is mutated in Glycine; T21N, Threonine in position 21 is mutated in Asparagine; W20stop, Tryptophan is mutated in stop codon; unknown, no mutation in mgrB gene.
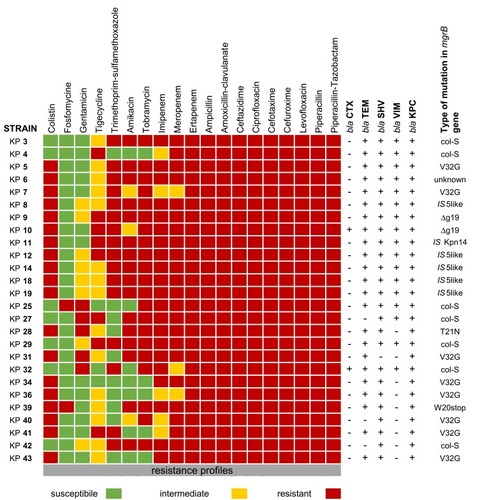
Figure 3 Alignment of FASTA mgrB sequence in col-R K. pneumoniae without ISs compared with wild-type (WT) sequences in col-S isolates.
Notes: WT strains in blue; mutation highlighted in red.
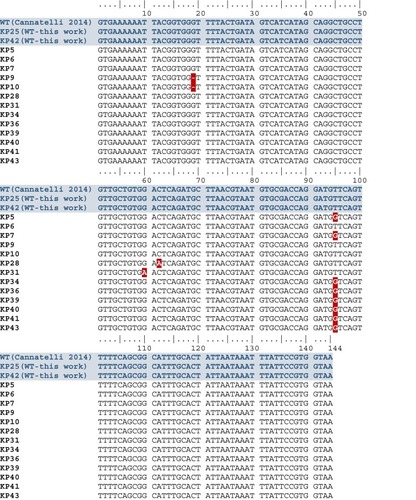
Figure 4 FASTA alignment of MgrB amino acid sequence in col-R K. pneumoniae without ISs compared with wild-type (WT) sequence of col-S strains.
Notes: WT MgrB in col-S strains in light blue; non-functional MgrB without ISs in col-R strains in green; truncated MgrB in col-R strains in yellow.
Three STs were identified (discriminatory power=0.61): the ST512 as the most common (n=13, 50%), followed by ST101 (n=10), and ST307 (n=2). It was not possible to define ST for KP10. ST512 was the most frequently detected during 2014–2015 (84.6%), while ST101 was the predominant during 2016–2017 (61.5%).
Discussion
The rapid spread of antibiotics resistance is nowadays a major concern causing untreatable infections in humans. A rising of MDR rate would lead to 10 million people dying every year by 2050, which exceeds the 8.2 million estimated deaths due to cancer.Citation35
This study describes the AMR profiles and molecular epidemiological insights concerning colistin-resistant CR-Kp isolated during 2014–2017. K. pneumoniae were most commonly isolated from patients aged ≥60 years who were treated in the ICU, where invasive procedures with devices at risk of generating biofilms formation play a crucial role in the occurrence of CR-Kp.Citation9,Citation36,Citation37
Twenty-one antimicrobial susceptibility patterns were found, underlining high inter-strain diversity. All cultures had an MDR pattern, with high percentages of carbapenem (76.9%) and colistin resistance (73%). Conversely, 92% of cultures were susceptible to fosfomycin, which has been recently evaluated for treating extensively drug-resistant (XDR) pathogens, although resistance associated to fosA gene is emerging and can be transferred between Enterobacteriaceae.Citation38
The increased application of colistin therapy for infections due to MDR Gram-negative bacteria has contributed to the spread of transmissible resistance, and may speed up the progression from XDR to Pan-drug Resistant (PDR) Enterobacteriaceae.Citation4,Citation14,Citation39 In our study, prevalence of col-R K. pneumoniae was higher than that reported in other Italian studies, ranging between 36.1% and 50%.Citation9,Citation40–Citation42
Results regarding ESBLs presence are in line with other studies, where plasmid-acquired blaTEM and blaSHV were frequently associated with Klebsiella spp. infections.Citation43,Citation44 Conversely, blaCTX-M enzymes have become the most prevalent in E. coli, with potential to spread beyond the hospital environment in other species.Citation45 The identification of ESBL-producing Klebsiella in hospital settings should be followed by infection control interventions, with reinforcement of hand hygiene of primary importance, followed by compliance with guidelines on antibiotic stewardship, and removal of contaminated devices.Citation46
Concerning carbapenemases encoding genes, blaVIM and blaKPC genes were detected in 70% and 100% isolates, respectively, which is consistent with the endemic KPC circulation reported in Italy.Citation5 Globally, the most worrying scenario is the increasing spread and dissemination of KPC-producing K. pneumoniae of clonal complex CC258 and CC512 being responsible for several outbreaks, unlike VIM carbapenemase, which is currently not widely diffused in Italy.Citation8,Citation47 Furthermore, K. pneumoniae producing NDM-1 or OXA-48 were not detected, similarly to IMP and GES, according to national data reporting sporadic cases.Citation45
The increasing occurrence of col-R strains is considered a global concern. In Italy, a retrospective study (from January 2010 to June 2014) reported a threefold increase of colistin-resistance rate in KPC-producing K. pneumoniae in blood isolates, and 51% mortality at 30 days due to bloodstream infections.Citation48
In our study, mcr-1,2,3,4,5,6,7,8 were not detected in col-R strains, which is consistent with previous reports,Citation42 being more frequently detected in E. coli than in K. pneumoniae.Citation49 Approximately 95% of col-R isolates carried alterations in mgrB, which is likely to be responsible for the colistin-resistant phenotypes. Inactivation of mgrB throughout ISs especially by IS5-like and ISKpn14 elements was detected in six out of 19 col-R strains. These mechanisms were reported elsewhere,Citation13,Citation16 and ISs transfer within genomes and plasmids has been considered a common driver of diversity and acquisition of antibiotic resistance.Citation50 Furthermore, it has been reported that plasmids transfer between strains within the gut is a potential mechanism of indirect acquisition of colistin resistance.Citation51 As assessed in vitro, IS interrupting mgrB and conferring colistin resistance was initially located on a plasmid.Citation52
In three isolates, a truncated MgrB due to one single nucleotide deletion causing frameshift mutation and premature termination was found, as reported elsewhere.Citation53 In addition, nine isolates had a non-functional MgrB due to one amino acid change (V32G, T21N and W20Stop), as observed in other studies.Citation1,Citation9,Citation53 Hence, colistin resistance was linked to alterations in mgrB because complementation studies with a wild-type mgrB demonstrated that susceptibility to colistin can be successfully restored.Citation53 In our study, col-R KP6 showed a wild-type mgrB, suggesting mutations in other colistin-resistance-related genes within Pmr signaling system or by alternative mechanism(s).
The presence of identical mgrB alterations in isolates from the same ward and assigned to the same ST and PFGE profile supports the clonal expansion and cross-transmission in hospital setting.Citation53
PFGE revealed high level of strains diversity, and results from MLST indicated the circulation of ST512, ST101 and ST307. In the tested isolates, ST512, a single-locus variant of ST258, the most frequently detected clone responsible for KPC global spread,Citation54 was the most common that is consistent with studies elsewhere in Italy.Citation9,Citation37,Citation39 For KP10 strain, the ST was not assigned, being found a monoallelic variant of ST307 (4-1-1-52-1-1-7 instead of 4-1-2-52-1-1-7); thus, further analyses are needed to confirm the novel ST. Interestingly, MLST revealed that ST512 was the most frequently detected in 2014–2015, while ST101 prevailed during 2016–2017, suggesting a changed circulation in the latest years in our hospital. Remarkably, PFGE cluster VIII grouped 77% of col-R cultures, four of which isolated from patients within the ICU as indistinguishable pulsotypes, all carried transposons in mgrB and were isolated during Christmas season holidays (December 2014–January 2015). In particular, the cluster VIII included the cultures KP8 (isolation data 17/11/14), KP12 and KP14 (isolation data 12/1/14), and KP18 (isolation data 5/1/15) (), belonging to a group of strains isolated during an outbreak in the ICU ward, which was likely related to a low level of compliance to standard hygiene procedures because of reduced personnel availability, and underlined the likelihood of bacterial persistence in the hospital environment.Citation10
The discriminatory abilities of PFGE and MLST were compared by the number of unique STs and number of clusters identified. PFGE showed good discriminatory power, and it is still considered a reference method for the epidemiological investigations of infectious diseases, including nosocomial outbreaks.Citation55 PFGE, generating genome-wide DNA fingerprints with rare-cutter restriction enzymes, is also a cost-effective method; nevertheless, it is labor-intensive and may lack comparability between laboratories due to operator errors in identifying bands particularly when shifted or weak on PFGE gel image analysis. In our study, MLST was less discriminating than PFGE, as found elsewhere.Citation24 Anyway, MLST is considered the most suitable genotyping method for strains comparison, further providing data within laboratories, and appropriate for global and long term or evolutionary studies rather than local epidemiology.Citation55
Conclusions
To our knowledge, this is the first study concerning colistin and carbapenems resistance characteristics in clinical K. pneumoniae isolates from the Molise Region, central Italy. Although focusing on topic investigated elsewhere, our findings can be useful to better understand the most significant concerns on hospital infections by K. pneumoniae at a local level, and can support the molecular epidemiology data related to CR-Kp both nationally and internationally, hence contributing to complete the framework of the epidemiology of this microorganism.
This study confirms that CR-Kp infections are most commonly detected in ICU patients due to their critical conditions and invasive procedures like catheterization or tracheostomy. In our setting, the KPC enzyme remains the predominant carbapenemase in K. pneumoniae, followed by VIM. The highest prevalence of KPC was linked with ST512 prevalence, although a switching towards ST101 circulation was detected. A high level of colistin resistance was found, more than the rate reported in other studies, likely due to an overuse of colistin in our hospital setting, and it is associated with acquisition of insertion elements or accumulation of deleterious mutations in the mgrB gene. Further investigations are warranted to clarify the entire role of Pmr signaling system in col-R strains.
In conclusion, infections with carbapenem-resistant organisms, particularly when KPC-producing, are widely distributed, and antimicrobial treatments selected should be critically evaluated, since an optimal therapy is not yet defined.Citation56 In light of the lack of novel antimicrobial agents for the treatment of difficult healthcare infections, the implementation of proper prevention strategies and adequate staffing is essential to control the spread of MDR K. pneumoniae.Citation26 Moreover, the routine application of molecular analyses for rapid and accurate detection of determinants and mutations conferring resistance is crucial to reduce and control the burden of MDR bacterial infectionsCitation40 and to guide best-choice therapy for better patient outcomes, as well as to elucidate epidemiology and dynamics of dissemination in the hospital environment.
Certainly, while, on one hand, not all the infections are associated with modifiable factors, evidences suggest that the spectrum of situations in which currently it is possible to intervene is broader than in the past. Furthermore, triggering and modifiable factors are mostly due to inadequate healthcare practices, particularly to the failure in applying standard and specific precautions for infectious diseases to avoid unnecessary procedures, the inappropriate use of antibiotics, and the lack of human and technological resources to be committed in the care and prevention.
Ethics
A formal institutional review board process for the ethical approval of this research was not required; thus, it is not available since no experimental, clinical or diagnostic procedures other than ones required for clinical management of the patients were performed. Furthermore, patients' anonymous information were provided from the microbiology hospital laboratory, which isolated the strains. The study completely followed the principles outlined in the Declaration of Helsinki.
Author Contributions
All authors substantially contributed to conception and design, acquisition, analysis and interpretation of data, drafted and critically revised the article for important intellectual content, approved the final version to be published, and agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Acknowledgment
The authors are grateful to Dr. Jim McLauchlin, Public Health England, London, UK, for the useful comments and revisions provided to this work.
Data Availability
All data generated or analysed during this study are included in the manuscript .
Disclosure
The authors report no conflicts of interest in this work.
References
- Pitout JDD, Nordmann P, Poirel L. Carbapenemase-producing Klebsiella pneumoniae, a key pathogen set for global nosocomial dominance. Antimicrob Agents Chemother. 2015;59(10):5873–5884. doi:10.1128/AAC.01019-1526169401
- European Centre for Disease Prevention and Control. Point Prevalence Survey of Healthcare associated Infections and Antimicrobial Use in European Acute Care Hospitals. Stockholm: ECDC; 2013 Available from: https://ecdc.europa.eu/sites/portal/files/media/en/publications/Publications/healthcare-associated-infections-antimicrobial-use-PPS.pdf. Accessed 64, 2019.
- Navon-Venezia S, Kondratyeva K, Carattoli A. Klebsiella pneumoniae: a major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol Rev. 2017;41(3):252–275. doi:10.1093/femsre/fux01328521338
- Rello J, Kalwaje EV, Lagunes L, et al. A global priority list of the TOp TEn resistant Microorganisms (TOTEM) study at intensive care: a prioritization exercise based on multi-criteria decision analysis. Eur J Clin Microbiol Infect Dis. 2019;38(2):319–323. doi:10.1007/s10096-018-3428-y30426331
- Sabbatucci M, Iacchini S, Iannazzo S, et al. Sorveglianza nazionale delle batteriemie da enterobatteri produttori di carbapenemasi. Rapporti ISTISAN 17/18 ISSN:1123-3117.
- European Centre for Disease Prevention and Control. Surveillance of Antimicrobial Resistance in Europe 2017. Stockholm: ECDC; 2018 Available from: https://ecdc.europa.eu/sites/portal/files/documents/EARS-Net-report-2017-update-jan-2019.pdf. Accessed 64, 2019.
- Girmenia C, Serrao A, Canichella M. Epidemiology of carbapenem resistant Klebsiella pneumoniae infections in mediterranean countries. Mediterr J Hematol Infect Dis. 2016;8(1):2016032. doi:10.4084/mjhid.2016.032
- Munoz-Price LS, Poirel L, Bonomo RA, et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect Dis. 2013;13(9):785–796. doi:10.1016/S1473-3099(13)70190-723969216
- Giani T, Arena F, Vaggelli G, et al. Large nosocomial outbreak of colistin-resistant, carbapenemase-producing Klebsiella pneumoniae traced to clonal expansion of an mgrB deletion mutant. J Clin Microbiol. 2015;53(10):3341–3344. doi:10.1128/JCM.01017-1526202124
- Ripabelli G, Tamburro M, Guerrizio G, et al. Tracking multidrug-resistant Klebsiella pneumoniae from an Italian hospital: molecular epidemiology and surveillance by PFGE, RAPD and PCR-based resistance genes prevalence. Curr Microbiol. 2018;75(8):977–987. doi:10.1007/s00284-018-1475-329523910
- Ghasemi Y, Archin T, Kargar M, Mohkam M. A simple multiplex PCR for assessing prevalence of extended-spectrum β-lactamases producing Klebsiella pneumoniae in Intensive Care Units of a referral hospital in Shiraz, Iran. Asian Pac J Trop Med. 2013;6(9):703–708. doi:10.1016/S1995-7645(13)60122-423827147
- Calbo E, Garau J. The changing epidemiology of hospital outbreaks due to ESBL-producing Klebsiella pneumoniae: the CTX-M-15 type consolidation. Future Microbiol. 2015;10(6):1063–1075. doi:10.2217/fmb.15.2226059626
- Cannatelli A, Giani T, D’Andrea MM, et al. MgrB inactivation is a common mechanism of colistin resistance in KPC-producing Klebsiella pneumoniae of clinical origin. Antimicrob Agents Chemother. 2014;58(10):5696–5703. doi:10.1128/AAC.03110-1425022583
- Jafari Z, Harati AA, Haeili M, et al. Molecular epidemiology and drug resistance pattern of carbapenem-resistant Klebsiella pneumoniae isolates from Iran. Microb Drug Resist. 2019;25(3):336–343. doi:10.1089/mdr.2017.040430351186
- Cannatelli A, D’Andrea MM, Giani T, et al. In vivo emergence of colistin resistance in Klebsiella pneumoniae producing KPC-type carbapenemases mediated by insertional inactivation of the PhoQ/PhoP mgrB regulator. Antimicrob Agents Chemother. 2013;57(11):5521–5526. doi:10.1128/AAC.01480-13
- Poirel L, Jayol A, Bontron S, et al. The mgrB gene as a key target for acquired resistance to colistin in Klebsiella pneumoniae. J Antimicrob Chemother. 2015;70(1):75–80. doi:10.1093/jac/dku32325190723
- Barbieri NL, Nielsen DW, Wannemuehler Y, et al. mcr-1 identified in Avian Pathogenic Escherichia coli (APEC). PLoS One. 2017;12(3):e0172997. doi:10.1371/journal.pone.017299728264015
- Xavier BB, Lammens C, Ruhal R, et al. Identification of a novel plasmid-mediated colistin-resistance gene, mcr-2, in Escherichia coli, Belgium, June 2016. Euro Surveill. 2016;21(27). doi:10.2807/1560-7917.ES.2016.21.27.30280
- Yin W, Li H, Shen Y, et al. Novel plasmid-mediated colistin resistance gene mcr-3 in Escherichia coli. MBio. 2017;8(4):e01166–17.28811346
- Carattoli A, Villa L, Feudi C, et al. Novel plasmid-mediated colistin resistance mcr-4 gene in Salmonella and Escherichia coli, Italy 2013, Spain and Belgium, 2015 to 2016. Euro Surveill. 2017;22(31). doi:10.2807/1560-7917.ES.2017.22.31.30589
- Borowiak M, Fischer J, Hammerl JA, Hendriksen SR, Szabo I, Malorny B. Identification of a novel transposon-associated phosphoethanolamine transferase gene, mcr-5, conferring colistin resistance in d-tartrate fermenting Salmonella enterica subsp. enterica serovar Paratyphi B. J Antimicrob Chemother. 2017;72(12):3317–3324. doi:10.1093/jac/dkx32728962028
- Wang X, Wang Y, Zhou Y, et al. Emergence of a novel mobile colistin resistance gene, mcr-8, in NDM-producing Klebsiella pneumoniae. Emerg Microbes Infect. 2018;7(1):122. doi:10.1038/s41426-018-0124-z29970891
- Yang YQ, Li YX, Lei CW, Zhang AY, Wang HN. Novel plasmid-mediated colistin resistance gene mcr-7.1 in Klebsiella pneumoniae. J Antimicrob Chemother. 2018;73(7):1791–1795. doi:10.1093/jac/dky11129912417
- Sammarco ML, Ripabelli G, Tamburro M. Epidemiologia molecolare delle malattie infettive: metodi di analisi ed interpretazione dei risultati. Ann Ig. 2014;26:10–45. doi:10.7416/ai.2014.195624452182
- Guo C, Yang X, Wu Y, et al. MLST-based inference of genetic diversity and population structure of clinical Klebsiella pneumoniae, China. Sci Rep. 2015;5:7612. doi:10.1038/srep0761225556771
- Ripabelli G, Salzo A, Mariano A, Sammarco ML, Tamburro M; Collaborative Group for HAIs Point Prevalence Surveys in Molise Region. Healthcare-associated infections point prevalence survey and antimicrobials use in acute care hospitals (PPS 2016-2017) and long-term care facilities (HALT-3): a comprehensive report of the first experience in Molise Region, Central Italy, and targeted intervention strategies. J Infect Public Health. 2019;2(4):509–515.
- European committee on antimicrobial susceptibility testing, EUCAST, 2019 Eucast.org. Available from: www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_9.0_Breakpoint_Tables.pdf. Accessed 87, 2019.
- Dallenne C, Da Costa A, Decré D, Favier C, Arlet G. Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J Antimicrob Chemother. 2010;65(3):490–495. doi:10.1093/jac/dkp49820071363
- Nordmann P, Naas T, Poirel L. Global spread of carbapenemase-producing Enterobacteriaceae. Emerg Infect Dis. 2011;17(10):1791–1798. doi:10.3201/eid1710.11065522000347
- Choi Y, Sims GE, Murphy S, Miller JR, Chan AP. Predicting the functional effect of amino acid substitutions and indels. PLoS One. 2012;7(10):46688. doi:10.1371/journal.pone.0046688
- Han H, Zhou H, Li H, et al. Optimization of pulsed-field gel electrophoresis for subtyping of Klebsiella pneumoniae. Int J Environ Res Public Health. 2013;10(7):2720–2731. doi:10.3390/ijerph1007272023880721
- Tenover FC, Arbeit RD, Goering RV, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33(9):2233.7494007
- Diancourt L, Passet V, Verhoef J, Grimont PAD, Brisse S. Multilocus sequence typing of Klebsiella pneumoniae nosocomial isolates. J Clin Microbiol. 2005;43(8):4178–4182. doi:10.1128/JCM.43.8.4178-4182.200516081970
- Hunter PR, Gaston MA. Numerical index of the discriminatory ability of typing systems: an application of Simpson’s index of diversity. J Clin Microbiol. 1988;26(11):2465–2466.3069867
- O’Neill J. Tackling drug-resistant infections globally: final report and recommendations. 2016 Available from: https://amr-review.org/sites/default/files/160518_Final%20paper_with%20cover.pdf. Accessed 55, 2019.
- Codjoe FS, Donkor ES. Carbapenem resistance: a review. Med Sci (Basel). 2018;6(1):E1.
- Cristina ML, Sartini M, Ottria G, et al. Epidemiology and biomolecular characterization of carbapenem-resistant Klebsiella pneumoniae in an Italian hospital. J Prev Med Hyg. 2016;57(3):149–156.
- Klontz EH, Tomich AD, Günther S, et al. Structure and dynamics of fosA-mediated fosfomycin resistance in Klebsiella pneumoniae and Escherichia coli. Antimicrob Agents Chemother. 2017;61(11):e01572–17. doi:10.1128/AAC.01572-1728874374
- Granata G, Petrosillo N. Resistance to colistin in Klebsiella pneumoniae: a 4.0 strain? Infect Dis Rep. 2017;9(2):7104. doi:10.4081/idr.2017.710428626539
- Giordano C, Barnini S, Tsioutis C, et al. Expansion of KPC-producing Klebsiella pneumoniae with various mgrB mutations giving rise to colistin resistance: the role of IS L3 on plasmids. Int J Antimicrob Agents. 2018;51(2):260–265. doi:10.1016/j.ijantimicag.2017.10.01129097338
- Moradigaravand D, Martin V, Peacock SJ, Parkhill J. Evolution and epidemiology of multidrug-resistant Klebsiella pneumoniae in the United Kingdom and Ireland. MBio. 2017;8(1):e01976–16. doi:10.1128/mBio.e01976-1628223459
- Lomonaco S, Crawford MA, Lascols C, et al. Resistome of carbapenem- and colistin-resistant Klebsiella pneumoniae clinical isolates. PLoS One. 2018;13(6):0198526. doi:10.1371/journal.pone.0198526
- Corbella M, Caltagirone M, Gaiarsa S, et al. Characterization of an outbreak of extended-spectrum β-lactamase-producing Klebsiella pneumoniae in a neonatal intensive care unit in Italy. Microb Drug Resist. 2018;24(8):1128–1136. doi:10.1089/mdr.2017.027029368992
- Shakib P, Ramazanzadeh R, Taherikalani M, Nouri B. Detection of extended-spectrum beta-lactamases (ESBLs) and antibiotic susceptibility patterns in Klebsiella pneumoniae in Western, Iran. Infect Disord Drug Targets. 2018;18(2):156–163. doi:10.2174/187152651766617071310173428707597
- Day MJ, Hopkins KL, Wareham D. Typing and epidemiological surveillance show that UK bloodstream Escherichia coli with extended-spectrum β-lactamases correspond to human gut strains, but not those from dinner. Lancet Infect Dis. 2019 ISSN: 1473-3099.
- Habboush Y, Guzman N. Antibiotic Resistance. StatPearls Publishing; 2018. Available from: https://www.ncbi.nlm.nih.gov/books/NBK513277. Accessed July 15, 2019.
- Cantón R, Akóva M, Carmeli Y, et al. Rapid evolution and spread of carbapenemases among Enterobacteriaceae in Europe. Clin Microbiol Infect. 2012;18(5):413–431. doi:10.1111/j.1469-0691.2012.03821.x22507109
- Giacobbe DR, Del Bono V, Trecarichi EM, et al. Risk factors for bloodstream infections due to colistin-resistant KPC-producing Klebsiella pneumoniae: results from a multicenter case–control–control study. Clin Microbiol Infect. 2015;21(12):1106.e1–8. doi:10.1016/j.cmi.2015.08.001
- Wise MG, Estabrook MA, Sahm DF, Stone GG, Kazmierczak KM. Prevalence of mcr-type genes among colistin-resistant Enterobacteriaceae collected in 2014-2016 as part of the INFORM global surveillance program. PLoS One. 2018;13(4):0195281. doi:10.1371/journal.pone.0195281
- Siguier P, Gourbeyre E, Chandler M. Bacterial insertion sequences: their genomic impact and diversity. FEMS Microbiol Rev. 2014;38(5):865–891. doi:10.1111/1574-6976.1206724499397
- Huddleston JR. Horizontal gene transfer in the human gastrointestinal tract: potential spread of antibiotic resistance genes. Infect Drug Resist. 2014;7:167–176. doi:10.2147/IDR25018641
- López-Camacho E, Gómez-Gil R, Tobes R, et al. Genomic analysis of the emergence and evolution of multidrug resistance during a Klebsiella pneumoniae outbreak including carbapenem and colistin resistance. J Antimicrob Chemother. 2014;69(3):632–636. doi:10.1093/jac/dkt41924155060
- Esposito EP, Cervoni M, Bernardo M, et al. Molecular epidemiology and virulence profiles of colistin-resistant Klebsiella pneumoniae blood isolates from the hospital agency “Ospedale dei Colli”, Naples, Italy. Front Microbiol. 2018;9:1463. doi:10.3389/fmicb.2018.0146330061868
- Bakour S, Sahli F, Touati A, Rolain JM. Emergence of KPC-producing Klebsiella pneumoniae ST512 isolated from cerebrospinal fluid of a child in Algeria. New Microbes New Infect. 2014;3:34–36. doi:10.1016/j.nmni.2014.09.00125755890
- Giacometti F, Piva S, Vranckx K, et al. Application of MALDI-TOF MS for the subtyping of Arcobacter butzleri strains and comparison with their MLST and PFGE types. Int J Food Microbiol. 2018;277:50–57. doi:10.1016/j.ijfoodmicro.2018.04.02629684765
- Ripabelli G, Sammarco ML, Scutellà M, Felice V, Tamburro M. Carbapenem-resistant KPC- and TEM-producing Escherichia coli ST131 isolated from a hospitalized patient with urinary tract infection: first isolation in Molise region, Central Italy, July 2018. Microb Drug Resist. 2019. doi:10.1089/mdr.2019.0085