
Abstract
Purpose
The multidrug resistance Enterobacteriaceae cause many serious infections resulting in prolonged hospitalization, increased treatment charges and mortality rate. In this study, we characterized blaNDM-5-positive multidrug resistance commensal Escherichia coli (CE) isolated from diarrheal patients in Kolkata, India.
Methods
Three CE strains were isolated from diarrheal stools, which were negative for different pathogroups of diarrheagenic E. coli (DEC). The presence of carbapenemases encoding genes and other antimicrobial resistance genes (ARGs) was detected using PCR. The genetic arrangement adjoining blaNDM-5 was investigated by plasmid genome sequencing. The genetic relatedness of the strains was determined by pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST) methods.
Results
In addition to colistin, the blaNDM-5-positive CE strains showed resistance to most of the antibiotics. Higher MICs were detected for ciprofloxacin (>32 mg/L) and imipenem (8 mg/L). Molecular typing revealed that three CE strains belonged to two different STs (ST 101 and ST 648) but they were 95% similar in the PFGE analysis. Screening for ARGs revealed that CE strains harbored Int-1, blaTEM, blaCTX-M3, blaOXA-1, blaOXA-7, blaOXA-9, tetA, strA, aadA1, aadB, sul2, floR, mph(A), and aac(6´)-Ib-cr. In conjugation experiment, transfer frequencies ranged from 2.5×10−3 to 8.4x10−5. The blaNDM-5 gene was located on a 94-kb pNDM-TC-CE-89 type plasmid, which is highly similar to the IncFII plasmid harboring an IS26-IS30-blaNDM-5-bleMBL-trpF-dsbd-IS91-dhps structure.
Conclusion
To the best of our knowledge, this is the first report on carbapenem resistance involving the blaNDM-5 gene in CE from diarrheal patients. The circulation of blaNDM-5 gene in CE is worrisome, since it has the potential to transfer blaNDM-5 gene to other enteric pathogens.
Introduction
The emergence and rapid spread of carbapenemases-producing Enterobacteriaceae is a serious public-health concern because carbapenems are the last resort antibiotics to treat extensively multidrug-resistant (MDR) bacterial infections.Citation1 Amongst the newly emerging carbapenemases-producers, NDM is very important due to its increased MDR phenotype and rapid global dissemination with frequent allelic variations.Citation2 Since its first discovery in 2008, more than 20 variants of NDM have been identified in different bacterial species spread across different countries.Citation3–5 In 2011, the NDM-5 was first identified from an MDR E. coli isolated in the United Kingdom from a patient who had been previously treated in India.Citation6 The NDM-5 varied from NDM-1 by only two amino acid replacements at positions 88 (Val → Leu) and 54 (Met → Leu).Citation6 NDM-5-producing Enterobacteriaceae has been reported worldwide, including in Algeria,Citation7 Australia,Citation8 China,Citation9 Egypt,Citation10 India,Citation11 Italy,Citation12 Mali,Citation13 South Korea,Citation14 Spain,Citation15 and United States.Citation16
The mechanism for dissemination and transmission of blaNDM-5 among Enterobacteriaceae is primarily associated with the transfer of plasmids or the transposon-related mobile genetic elements.Citation17 The blaNDM-5 has been reported worldwide with numerous incompatibility-type plasmid groups and the most prevalent being IncFIA and IncFK in India,Citation18 IncFII in the United States,Citation16 IncF and IncX3 in China,Citation9 IncFIA/B and IncX3 in Korea,Citation14 and IncFIA and IncFIB in Egypt.Citation10
E. coli is the most common pathogen associated with community acquired infections. MDR in E. coli has become an increasing problem in human health management.Citation19 Acquisition of the gene encoding NDM by E. coli isolates is an important health issue due to its fast spread across the globe and also in other Gram-negative bacteria.Citation20 This emerging trend is directly associated with an increased use of carbapenems during the treatment of several infectious diseases.Citation21 In this study, we report on the incidence of the NDM-5-producing commensal E. coli isolated from diarrheal patients in Kolkata, India. To the best of our knowledge, this is the first report on CE strains producing NDM-5 carbapenemases. The present study characterized blaNDM-5 identified in CE isolated from diarrheal patients and its dissemination through conjugative plasmid into other enteric bacteria.
Materials and Methods
Clinical Specimens
In this study, stool specimens collected from acute diarrheal patients hospitalized at the Infectious Diseases Hospital, Kolkata, India, were tested for the identification of enteric pathogens. Patients who excreted >3 loose or liquid stools per day with no, some or severe dehydration were considered to have diarrhea.Citation22 Fecal specimens were collected in McCartney bottles using sterile catheters or as rectal swab in Cary Blair transportation medium and processed in the laboratory within 2 hrs for common enteric pathogens.Citation23
Identification and Isolation of Commensal E. coli
For identification and isolation of E. coli, fecal specimens were inoculated on MacConkey agar (Difco, USA) and incubated at 37°C for 16–18 hrs. Three typical colonies per sample from MacConkey agar plate (including both lactose fermenting and non-lactose fermenting colonies) were picked and sub-cultured on Luria Bertani agar (LBA, Difco, USA) plates. A small portion of cultures was taken from LBA and tested for indole-production by adding the Kovacs reagent. Colour changes to pink were recorded positive and additionally tested in triple sugar iron agar for typical E. coli. For further confirmation, all the isolates were individually tested on a Vitek-2 compact system (bioMérieux). Indole, biochemical and Vitek-2 compact-positive bacterial cultures from each of the three colonies plated on LBA plates were taken and mixed with 500 µL of sterile water or phosphate-buffer saline in 1.5 mL microfuge tubes. The bacterial suspension was boiled in a water bath for 10 min and then kept on ice for snap chill for 5 min. Bacterial suspensions were centrifuged at 8000 rpm for 10 min and the supernatants were used as DNA templates in the multiplex PCR for the detection of virulence marker genes, such as CVD432 and aaiC (enteroaggregative E. coli), eae and bfpA (enteropathogenic E. coli), elt and est (enterotoxigenic E. coli).Citation24 Simplex PCR was also performed for the detection of Shiga toxin–producing E. coli, and/or enterohemorrhagic E. coli(stx1, stx2), enteroinvasive E. coli (EIEC) (ipaH) and cytolethal distending toxin (cdt) gene for E. albertii.Citation25 We defined commensal E. coli when the tested colonies were negative for all the above virulence marker genes.
Antimicrobial Susceptibility Testing (AST)
AST was performed in accordance with the Clinical and Laboratory Standards InstituteCitation26 by disc diffusion method using commercially available discs (Becton Dickinson Company, USA) namely, ampicillin (AMP), ceftriaxone (CRO), cefotaxime (CTX), ceftazidime (CAZ), chloramphenicol (CHL), nalidixic acid (NA), ciprofloxacin (CIP), ofloxacin (OFX), norfloxacin (NOR), meropenem (MEM), streptomycin (STR), erythromycin (E), azithromycin (AZM), gentamicin (GM), tetracycline (TET), doxycycline (D), trimethoprim/sulfamethoxazole (SXT). In addition, MICs against ampicillin, azithromycin, ceftriaxone, cefotaxime, erythromycin, gentamicin, meropenem, and sulfamethoxazole/trimethoprim were determined using the E-test (AB Biodisk, Sweden), according to the manufacturer’s instructions. Detection of the extended spectrum β-lactamase (ESBL) activity was made using the combination disc test (cefotaxime, 30 μg; ceftazidime, 30 μg; with and without clavulanic acid, 10 μg). Strains were considered as ESBL positive, if they exhibit >5 mm increase in the size of the zone of inhibition of the β-lactamase inhibitor combinations in comparison to a third-generation cephalosporin without the β-lactamase inhibitor. E. coli ATCC25922 was served as control in antimicrobial susceptibility testing.
“O” Serogrouping of E. coli Isolates
The “O” serogrouping of E. coli was made using an E. coli “O” serotyping kit by slide agglutination test in accordance with the manufacturer’s instructions (Denka-Seiken Co. Ltd., Japan). This kit consists of eight “O” polyvalent and 43 monospecific antisera.
Molecular Typing
Pulsed-field gel electrophoresis (PFGE) was performed using a CHEF-Mapper (Bio-Rad, Hercules, USA) based on Pulse-Net standardized protocol.Citation27 PFGE was made by XbaI-digested genomic DNA of E. coli species. PFGE images were saved by using a Gel Doc XR system (Bio-Rad). The PFGE gel images were analysed using the BioNumerics software version 5.0 (Applied Maths, Sint-Martens-Latem, Belgium) by normalizing and aligning the peaks of the Salmonella enterica serovar Braenderup H9812 size standard. The banding pattern similarity and the clustering correlation coefficients were calculated by comparison of the dice-coefficient and UPGMA, respectively. Multilocus sequence typing (MLST) was also carried out for molecular typing. Bacterial genomic DNA was extracted from the CE strains by DNA mini kit (Qiagen, Germany). Seven housekeeping genes (adk, icd, fumC, gyrB, mdh, purA and recA) were amplified by PCR,Citation28 and the amplicons were submitted for DNA sequencing. ST analysis was done according to the protocols specified at the E. coli MLST website (http://mlst.warwick.ac.uk/mlst/dbs/Ecoli).
Antimicrobial Resistance Gene (ARGs) Screening
Different ARGs were screened by PCR, and the positive amplicons were confirmed by DNA sequencing. The PCR assays targeted AMR genes for carbapenemases (blaNDM, blaIMP, blaKPC, blaOXA-48, blaSIM, and blaVIM), gentamicin (aadB), streptomycin (aadA1 and strA), chloramphenicol (catA1, floR), extended-spectrum β-lactamase (blaTEM, blaSHV, blaPSE4, blaCTXM-3, blaOXA-1, blaOXA-7 and blaOXA-9), ciprofloxacin [aac(6´)-Ib-cr], and macrolides {mph(A), mph(B), erm(A), erm(B)}. Simplex PCR was carried out as described previously.Citation29
Mutations in the Quinolone Resistance-Determining Regions (QRDRs)
The chromosomal genes gyrA and parC were amplified by PCR for the QRDRs and sequenced to analyse the potential mutations. We used published primers for the amplification of gyrA and parC genes.Citation30
Conjugation Assay
To test the transfer of the blaNDM-5, conjugation assay was made using NDM-5 positive commensal E. coli (CE-89) as donor with two different recipients, namely E. coli J53 (sodium azide resistant) and Salmonella enterica serovar Bareilly. In brief, overnight seed cultures of the bacteria were inoculated in LB broth for late-exponential phase culture. Cell density of bacterial culture was adjusted to 1.5×108 cells/mL. Both the cultures of donor and recipient were mixed (at the ratio of 1:2) in 5 mL of LB broth and kept at 37°C for overnight. Several selective media with antibiotics were used to recover blaNDM−5 positive transconjugants (TC). MacConkey agar containing both sodium azide (100 μg/mL) and meropenem (5 μg/mL) was used to select TC of E. coli J53 (TC-E. coli J53). Similarly, transconjugants of Salmonella (TC-Salmonella) were obtained by selecting on xylose lysine deoxycholate (XLD, Difco) containing meropenem (5 μg/mL). Putative transconjugants harboring blaNDM−5 were detected by PCR analysis followed by amplicon sequencing. The conjugation frequency was calculated as the number of transconjugants per recipient cell.
Plasmid Profiling
Plasmid DNA was extracted from the CE strains and transconjugants using a modified Kado and Liu method.Citation31 The plasmid DNA of E. coli V517 and J53 pMG252 were used as molecular weight markers for determining the molecular weight of plasmid DNAs of CE strains and the transconjugants. Plasmid incompatibility of major groups was determined by using PCR-based replicon typing.Citation32
Plasmid Sequence Analysis
Plasmid carrying blaNDM-5 was extracted from the TC-CE-89 using a QIAGEN plasmid midi kit according to the manufacturer’s instructions (QIAGEN). The complete plasmid sequence was obtained using an Illumina HiSeq platform. The DNA library was prepared for the pair end sequencing. The derived reads were subsequently trimmed and assembled de novo using the CLC genomics work bench. The plasmid sequences were annotated by Prokaryotic Dynamic Programming Gene finding Algorithm (Prodigal tool v.2.6.3). The open reading frames were compared using BLAST against the non-redundant protein database. Different ARGs in the plasmid were identified by comparing the ORFs with the genes of the Comprehensive Antibiotic Resistance Database (CARD). For the ARGs, we selected the thresholds of identity >70% and subject coverage >90%. The circular map of the pNDM-TC-CE-89 plasmid was generated using the Snap Gene server.
Nucleotide Sequence
The complete nucleotide sequences of plasmid pNDM-TC-CE-89 were submitted to GenBank accession number MZ892875.
Results
Isolation, Identification and Serotyping of E. coli from Stool Specimens
Three carbapenem-resistant CE strains were isolated from three different hospitalized acute diarrheal patients in 2015. All three strains were confirmed as E. coli using biochemical and Vitek-2 compact system. As confirmed in the multiplex PCR assay, these CE strains did not harbour any virulence encoding genes specific to DEC. Serological results showed that the “O” antigen of the CE strains is different. One CE strain was untypable (ONT) and the other strains belonged to O167 and O86a ().
Table 1 Characterization of AMR and Their Encoding Genes in NDM-5-Producing Commensal E. coli
Antimicrobial Susceptibility Testing and MIC
All the three CE strains shared the similar AMR profile. As shown in , CE strains are resistant to meropenem, ciprofloxacin, nalidixic acid, norfloxacin, ampicillin, ofloxacin, tetracycline, ceftriaxone, sulfamethoxazole/trimethoprim, streptomycin, ceftazidime, erythromycin, chloramphenicol, gentamicin, and cefotaxime but remained susceptible to doxycycline and colistin. CE displayed a higher MIC value for most of the antibiotics (). The modified Hodge test and imipenem-EDTA double-disc synergy test confirmed the production of metallo-B-lactamases (MBLs) by the CE strains. Carbapenemase gene screening revealed that all the CE strains harbored the blaNDM-5 gene that was further confirmed by amplicon sequencing. The sequence of the blaNDM−5gene exhibited 100% homology with previously reported sequences.Citation6
Table 2 Minimum Inhibitory Concentration (MIC) of Donor, Recipients, and Transconjugants
PCR for Antimicrobial Resistance Genes and QRDRs
The results of ARGs detected in the CE are shown in . Class-1 integron was identified in the blaNDM−5 positive CE strains. In addition to blaNDM-5, CE were positive for β- lactamase–encoding genes blaTEM, blaCTX-M3, blaOXA-1, blaOXA-7 and blaOXA-9; tetracycline resistance marker gene tet(A), streptomycin-encoding gene strA and aadA1; gentamicin encoding gene aadB; sulfonamides encoding gene sul2; chloramphenicol encoding gene floR, macrolide resistance phosphotransferase encoding gene mph(A); and ciprofloxacin modifying enzyme-encoding gene aac(6´)-Ib-cr (amino glycoside actetyltransferase). Plasmid-mediated quinolone resistance (PMQR) qnrB gene was detected only in CE-89. Additionally, three strains had amino acid substitutions in the QRDRs, ie, in GyrA (Ser 83 Leu and Asp 87 Asn) and ParC (Ser 80 Ile) ().
MLST and PFGE
In the MLST sequence analysis, CE-88 and CE-89 were identified as ST101 Cplx (adk allele 43; fumC allele 41; gyrB allele 15; icd allele18; mdh allele 11; purA allele 7 and recA allele 6), whereas CE-93 was identified as ST648 Cplx (adk allele 92; fumC allele 4; gyrB allele 87; icd allele 96; mdh allele 70; purA allele 58 and recA allele 2, ). PFGE revealed that isolates had ~90–95% similarity, suggesting that the three CE strains are genetically related clones ().
Figure 1 Xba1 restriction patterns of genomic DNA of CE strains, Kolkata, India. Dendrogram was generated by using the unweighted pair group with the arithmetic mean method.

Plasmid Characterisation and Conjugation
The blaNDM-5 harboring CE carried multiple plasmids ranging from 5 kb to 95 kb. In the transconjugants, only a single plasmid of size ~94 kb was detected. Plasmid harboring the blaNDM-5 was transferable from CE-89 to a laboratory strain E. coli J53 and a clinical isolate of Salmonella enterica serovar Bareilly. While using E. coli J53 as recipients, the conjugation frequency was higher (∼105 transconjugants per donor cell) than S. Bareilly (∼103 transconjugants per donor cell) (). TC-E. coli J53 and TC-S. Bareilly acquired resistance to ampicillin, meropenem, ceftriaxone, ceftazidime, erythromycin, sulfamethoxazole/trimethoprim, cefotaxime, gentamicin and azithromycin, indicating the possibility that the blaNDM-5 plasmid carried the corresponding resistance to these antimicrobials. However, the CT- E. coli J53 and TC-S. Bareilly were susceptible to quinolone, fluoroquinolones and tetracycline. The MIC values of antimicrobials of donor and recipient are shown in . For meropenem, the MIC value of transconjugants was 4 mg/L, but higher MIC values were observed for other antibiotics.
Plasmid Sequence Analysis
The plasmid typing has identified that the CE harbours IncFII replicon. Considering that the blaNDM-5 harbored transferable plasmids of the same size and incompatibility group, we have chosen pNDM-TC-CE-89 as a representative for whole-genome sequencing to identify the genetic background and the flanking regions of blaNDM-5. This plasmid was identified to comprise several types of genes, including ARGs, mobile elements, and putative genes, genes encoding proteins that control plasmid replication, stability and transfer. Moreover, the sequence alignments based on BLAST revealed that pNDM-TC-CE-89 has 99% nucleotide identity with previously reported blaNDM-5 harboring plasmid of E. coli (99.9% sequence similarity, 100% query coverage; AP018144.1)Citation33 including the complete array of genes for replication, partitioning, type IV conjugative transfer system, maintenance and stabilization.
The nucleotide sequence length of pNDM-TC-CE-89 was 94,224-bp with an average GC content of 53%, which is slightly higher than that of E. coli (~50%). In addition, we observed that the plasmid has harbored different classes of ARGs, which include dihydrofolate reductase (dhfR), aminoglycoside 3”-nucleotidyltransferase (aadA), Class A β- lactamase (blaTEM), 23S rRNA dimethyl transferase (ermB) and macrolide 2’-phosphotransferase (mphA) family. The genes responsible for other antimicrobial resistance were not detected in this plasmid (). The ARGs BLAST sequence alignments and the most similar nucleotide sequences to those of the previously reported plasmid have been shown in .
Table 3 Characteristics of Plasmid Encoded AMR and Other Genes in Different Bacterial Species
Figure 2 Schematic representation and comparison of the sequences of different antimicrobial resistance encoding genes and their association with mobile genetic elements of E. coli (accession no. AP018144.1), (K) pneumoniae (accession no. CP024840.1) and commensal E. coli (CE-89) strains harboring blaNDM−5. Arrow lengths are proportionate to the lengths of the genes or open reading frames. Genetic structure of CE NDM isolates was identified the link between resistance genes and mobile genetic element. trpF: phosphoribosylanthranilate isomerase gene; IS26: IS26 transposase; IntI1: class 1 integron integrase IntI1; dfra12: dihydrofolate reductase; aadA2: aminoglycoside resistance protein; SMR: quaternary ammonium compound efflux SMR transporter QacE delta 1; Dhps: dihydropteroate synthase. IS91: IS91 family transposase; Dsbd: Thiol: disulfide interchange protein DsbD; PRAI: phosphoribosylanthranilate isomerase; bleMBL: bleomycin resistance protein; blaNDM-5: subclass B1 metallo-β-lactamase NDM-5; IS30: Transposase-like protein, IS30 family; blaTEM: β-lactamase; tnpR: Transposon Tn3 resolvase, mphA: macrolide 2’- phosphotransferase.
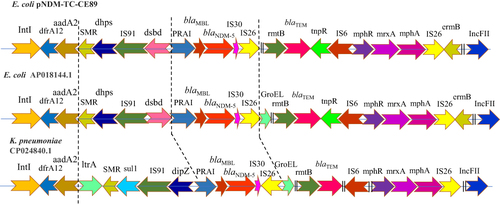
The blaNDM-5 within pNDM-TC-CE-89 was situated in a complex integron, connected by two IS26 sequences having an IS91 element and a class 1 integron with the intI1 gene truncated by one of the IS26 copies and the aadA2-dfrA12 resistance gene cassettes. The flanking genetic structure of the blaNDM-5 of pNDM-TC-CE-89 plasmid composed of an IS30 and IS26 located upstream, and the genes bleMBL (bleomycin resistance gene), trpF (phosphor ribosyl anthranilate isomerase), dsbd (Dsbd superfamily protein), IS91 and dhps (dihydropteroate synthase) in downstream (IS26-IS30-blaNDM-5-bleMBL-trpF-dsbd-IS91-dhps) ().
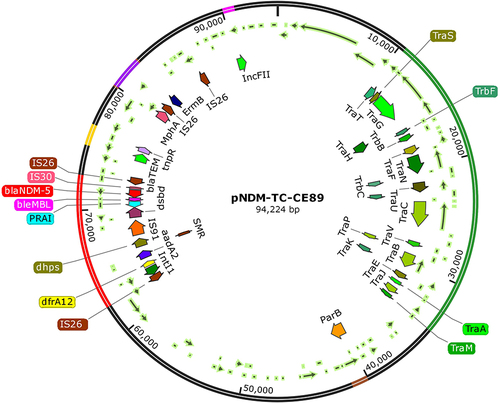
Figure 3 Salient features of blaNDM−5 encoding plasmid pNDM-TC-CE-89 plasmid with other resistance genes, transposons and type IV conjugative transfer system. The circle in red colour indicates the flanking structure of blaNDM−5. Transfer machinery are showed in green colour. Cloned par loci genes are indicated in yellow colour and the transposons IS26 elements are marked in brown.
Discussion
The emergence of NDM-producing Enterobacteriaceae is an increasing threat to global health. The blaNDM harboring strains confer resistance to most of the β-lactams and also express resistance to cephalosporins, quinolones, and aminoglycosides with different ARGs, which allows pathogens to become MDR.Citation34 AMR is not restricted only to pathogenic bacteria, but the commensal gut bacteria also play a role in harboring and transmission of ARGs. E. coli is commonly found as a commensal gut microbiota and considered as a reservoir of acquired AMR determinants.Citation35 Since blaNDM-5 was first identified in the UK, many bacterial species harboring this gene have emerged in several countries.Citation36 Although NDM-5-producing strains appear to be a common NDM variant, they remain less compared to NDM-1-variant.Citation37 In India, different sequence types (STs) of NDM-5 producing E. coli and other bacterial strains have been detected from clinical and other sources. The E. coli isolates harbour the blaNDM-5 gene in either plasmids or the transposon-related mobile genetic elements. Different incompatibility groups of plasmids belonging to the IncF, IncFII IncX3, K and FrepB are reported to carry the blaNDM-5 gene.Citation11,Citation38,Citation39 Furthermore, E. coli harbouring the blaNDM-5 found on IncX3 plasmids isolated from patients who travelled from India to Australia and Denmark, suggesting the origin of NDM-5 with STs 648 and 1284, respectively.Citation8,Citation40 In addition, the NDM-5-producing E. coli has been identified from fresh fish and mastitic milk samples in India.Citation41,Citation42 In this study, the three NDM-5-producing CE displayed close genetic similarity as evidenced from PFGE and plasmid typing and AMR profile.
CE strains harboring blaNDM-5 were typed as ST101 and ST648. ST101 of E. coli producing NDM-5 in China and Myanmar were documented as an epidemic clone of substantial public health concern carrying plasmids associated with ARGs.Citation43 E. coli ST648 was first reported in India and UKCitation6 and has also been sporadically detected worldwide.Citation8–12 The ST648 isolate detected in this study was highly resistant to several different classes of antimicrobial agents. A similar observation was made in earlier studies.Citation8,Citation43
Fluoroquinolone resistance in Enterobacteriaceae is primarily due to mutations in the QRDRs of DNA gyrase and topoisomerase IV. The two enzymes are encoded by the gyrA for DNA gyrase (topoisomerase II) and parC for topoisomerase IV. In this study, all three isolates displayed mutations in the QRDR region of gyrA and parC, which has also been reported in many investigations.Citation30
The IncFII plasmids are highly mobile and carry several blaNDM variants in Enterobacteriaceae.Citation44 A recent study has shown the ability of blaNDM bearing IncFII plasmid transfer to different members of Enterobacteriaceae along with the other ARGs.Citation45 In this study, the transfer of pNDM-CE89 carrying blaNDM−5 to other bacteria has been demonstrated along with the expression of MDR. Therefore, commensal E. coli having IncFII plasmids with blaNDM-5 are epidemiologically important, as there is a high possibility of its transfer to other pathogens, which will be a major concern in the clinical management of infections.
BLAST-based sequence alignments revealed that the plasmid pNDM-CE-89 had the most similar nucleotide sequences in other plasmids such as pYJ6-NDM5 (accession number AP023236.1) from an E. coli strain, pM941-NDM5 (accession number AP023454.1) from an K. pneumoniae strain, and pM2_FII (GenBank accession AP018144.1) from an E. coli strain. The plasmid pNDM-CE-89 possesses complex structures as it contains the gene encoding MBLs blaNDM-5, a large group of genetic elements (integrons, transposons, and ISCRs), IntI1 (class 1 integron integrase) and mphA, a family of macrolide phosphotransferase.
The location of blaNDM−5 in pNDM-CE-89 was identified between the two IS26 elements. The similar flanking genetic structure has previously been observed in other blaNDM−5 bearing plasmids.Citation33 The IS26 element has been reported in many members of Enterobacteriaceae and exists together with the β-lactamases region, which is a part of transposon-like structure in many plasmids.Citation46 In pNDM-CE-89, the association of blaNDM−5 with IS26 makes a condition favourable for its mobilization and function. The flanking region of NDM producers carried a highly conserved gene segment (IS26-IS30-blaNDM5-bleMBL-trpF) suggesting that these four genes are important for the transmission and expression of AMR.
In conclusion, the global dissemination of NDM-producing Gram-negative bacteria is of great concern. Our data suggest that commensal E. coli in the gut acts as a reservoir for NDM carrying plasmid and the other ARGs. More epidemiological and clinical studies are needed to elucidate the mechanisms of emergence, evolution and dissemination of NDM-5 in commensal E. coli and other gut microbes.
Ethical Approval
This study was approved by the “Institutional Ethics Committee (IEC) of the National Institute of Cholera and Enteric Diseases in Kolkata, India (registration number: A‐1/2015‐IEC)”. Written consent was obtained from each adult patient or the parent/guardian of the child patients enrolled in this study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in relation to this work.
Acknowledgment
Dr. Goutam Chowdhury acknowledges the support from Okayama University, Okayama, Japan. TR is a Senior Scientist, Indian National Science Academy, New Delhi, India.
Additional information
Funding
References
- van Duin D, Doi Y. The global epidemiology of carbapenemase-producing Enterobacteriaceae. Virulence. 2017;8(4):460–469. doi:10.1080/21505594.2016.1222343
- Suay-García B, Pérez-Gracia MT. Present and future of carbapenem resistant Enterobacteriaceae (CRE) infections. Antibiotics. 2019;8:122. doi:10.3390/antibiotics8030122
- Yong D, Toleman MA, Giske CG, et al. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother. 2009;53:5046–5054. doi:10.1128/AAC.00774-09
- Wu W, Feng Y, Tang G, Qiao F, McNally A, Zong Z. NDM Metallo-β-Lactamases and their bacterial producers in health care settings. Clin Microbiol Rev. 2019;32:e00115–e00118. doi:10.1128/CMR.00115-18
- Basu S. Variants of the New Delhi metallo-β-lactamase: new kids on the block. Future Microbiol. 2020;15:465–467. doi:10.2217/fmb-2020-0035
- Hornsey M, Phee L, Wareham DW. A novel variant, NDM-5, of the New Delhi metallo- β-lactamase in a multidrug-resistant Escherichia coli ST648 isolate recovered from a patient in the United Kingdom. Antimicrob Agents Chemother. 2011;55:5952–5954. doi:10.1128/AAC.05108-11
- Sassi A, Loucif L, Gupta SK, Dekhil M, Chettibi H, Rolain JM. NDM-5 carbapenemase encoding gene in multidrug-resistant clinical isolates of Escherichia coli from Algeria. Antimicrob Agents Chemother. 2014;58:5606–5608. doi:10.1128/AAC.02818-13
- Wailan AM, Paterson DL, Caffery M, Sowden D, Sidjabat HE. Draft Genome Sequence of NDM-5-producing Escherichia coli sequence type 648 and genetic context of blaNDM5 in Australia. Genome Announc. 2015;3:e00194–e001915. doi:10.1128/genomeA.00194-15
- Liu Z, Xiao X, Li Y, Liu Y, Li R, Wang Z. Emergence of incx3 plasmid- harboring blaNDM-5 dominated by Escherichia coli ST48 in a goose farm in Jiangsu, China. Front Microbiol. 2019;10:2002. doi:10.3389/fmicb.2019.02002
- Gamal D, Fernández-Martínez M, El-Defrawy I, Ocampo-Sosa AA, Martínez-Martínez L. First identification of NDM-5 associated with OXA-181 in Escherichia coli from Egypt. Emerg Microbes Infect. 2016;5(1):e30. doi:10.1038/emi.2016.24
- Rahman M, Shukla SK, Prasad KN, et al. Prevalence and molecular characterisation of New Delhi metallo-β-lactamases NDM-1, NDM-5, NDM-6 and NDM-7 in multidrug-resistant Enterobacteriaceae from India. Int J Antimicrob Agents. 2014;44(1):30–37. doi:10.1016/j.ijantimicag.2014.03.003
- Giufrè M, Errico G, Accogli M, et al. Emergence of NDM-5-producing Escherichia coli sequence type 167 clone in Italy. Int J Antimicrob Agents. 2018;52(1):76–81. doi:10.1016/j.ijantimicag.2018.02.020
- Muggeo A, Maiga A, Maiga I, et al. First description of IncX3 NDM-5-producing plasmid within Escherichia coli ST448 in Mali. J Med Microbiol. 2020;69(5):685–688. doi:10.1099/jmm.0.001182
- Baek JY, Cho SY, Kim SH, et al. Plasmid analysis of Escherichia coli isolates from South Korea co-producing NDM-5 and OXA-181 carbapenemases. Plasmid. 2019;104:102417. doi:10.1016/j.plasmid.2019.102417
- Pitart C, Solé M, Roca I, et al. Molecular Characterization of bla NDM-5 carried on an IncFII Plasmid in an Escherichia coli Isolate from a Nontraveler Patient in Spain. Antimicrob Agents Chemother. 2015;59(1):659–662. doi:10.1128/AAC.04040-14
- Rojas LJ, Hujer AM, Rudin SD, et al. NDM- 5 and OXA-181 Beta-Lactamases, a significant threat continues to spread in the Americas. Antimicrob Agents Chemother. 2017;61(7):e00454–e00417. doi:10.1128/AAC.00454-17
- Zou H, Jia X, Liu H, Li S, Wu X, Huang S. Emergence of NDM-5- producing Escherichia coli in a Teaching Hospital in Chongqing, China: incF-type Plasmids may contribute to the prevalence of blaNDM-5. Front Microbiol. 2020;11:334. doi:10.3389/fmicb.2020.00334
- Ahmad N, Khalid S, Ali SM, Khan AU. Occurrence of blaNDM variants among Enterobacteriaceae from a neonatal intensive care unit in a Northern India Hospital. Front Microbiol. 2018;9:407. doi:10.3389/fmicb.2018.00407
- Paitan Y. Current trends in antimicrobial resistance of Escherichia coli. Curr Top Microbiol Immunol. 2018;416:181–211. doi:10.1007/82_2018_110
- Ceccarelli D, Hesp A, van der GJ, et al. Antimicrobial resistance prevalence in commensal Escherichia coli from broilers, fattening turkeys, fattening pigs and veal calves in European countries and association with antimicrobial usage at country level. J Med Microbiol. 2020;69:537–547. doi:10.1099/jmm.0.001176
- Kumarasamy KK, Toleman MA, Walsh TR, et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: a molecular, biological, and epidemiological study. Lancet Infect Dis. 2010;10(9):597–602. doi:10.1016/S1473-3099(10)70143-2
- Gidudu J, Sack DA, Pina M, et al. Diarrhea: case definition and guidelines for collection, analysis, and presentation of immunization safety data. Vaccine. 2011;29(5):1053–1071. doi:10.1016/j.vaccine.2010.11.065
- Nair GB, Ramamurthy T, Bhattacharya MK, et al. Emerging trends in the etiology of enteric pathogens as evidenced from an active surveillance of hospitalized diarrhoeal patients in Kolkata, India. Gut Pathog. 2010;2(1):4. doi:10.1186/1757-4749-2-4
- Panchalingam S, Antonio M, Hossain A, et al. Diagnostic microbiologic methods in the GEMS-1 case/control study. Clin Infect Dis. 2012;55(suppl_4):S294–S302. doi:10.1093/cid/cis754
- Hinenoya A, Ichimura H, Yasuda N, et al. Development of a specific cytolethal distending toxin (cdt) gene (Eacdt)-based PCR assay for the detection of Escherichia albertii. Diagn Microbiol Infect Dis. 2019;95:119–124. doi:10.1016/j.diagmicrobio.2019.04.018
- Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing, 29th ed. CLSI supplement M100S. Wayne, PA: Clinical and Laboratory Standards Institute; 2019.
- Ribot EM, Fair MA, Gautom R, et al. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157: H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog Dis. 2006;3:59–67. doi:10.1089/fpd.2006.3.59
- Wirth T, Falush D, Lan R, et al. Sex and virulence in Escherichia coli: an evolutionary perspective. Mol Microbiol. 2006;60(5):1136–1151. doi:10.1111/j.1365-2958.2006.05172.x
- Chowdhury G, Ramamurthy T, Ghosh A, Dutta S, Takahashi E, Mukhopadhyay AK. Emergence of azithromycin resistance mediated by phosphotransferase-Encoding mph (A) in diarrheagenic Vibrio fluvialis. mSphere. 2019;4(3):e00215–e00219. doi:10.1128/mSphere.00215-19
- Rawat N, Jamwal R, Devi PP, et al. Detection of unprecedented level of antibiotic resistance and identification of antibiotic resistance factors, including QRDR mutations in Escherichia coli isolated from commercial chickens from North India. J Appl Microbiol. 2021;10. doi:10.1111/jam.15209
- Kado CI, Liu ST. Rapid procedure for detection and isolation of large and small plasmids. J Bacteriol. 1981;145(3):1365–1373. doi:10.1128/jb.145.3.1365-1373.1981
- Carattoli A, Bertini A, Villa L, Falbo V, Hopkins KL, Threlfall EJ. Identification of plasmids by PCR-based replicon typing. J Microbiol Methods. 2005;63(3):219–228. doi:10.1016/j.mimet.2005.03.018
- Sugawara Y, Akeda Y, Sakamoto N, et al. Genetic characterization of blaNDM-harboring plasmids in carbapenem-resistant Escherichia coli from Myanmar. PLoS One. 2017;12:e0184720. doi:10.1371/journal.pone.0184720
- Nordmann P, Poirel L. Epidemiology and diagnostics of carbapenem resistance in gram-negative bacteria. Clin Infect Dis. 2019;69:S521–S528. doi:10.1093/cid/ciz824
- Thanh DP, Thi Nguyen TN, Vu TD, et al. Commensal Escherichia coli are a reservoir for the transfer of XDR plasmids into epidemic fluoroquinolone-resistant Shigella sonnei. Nat Microbiol. 2020;5(2):256–264. doi:10.1038/s41564-019-0645-9
- Baloch Z, Lv L, Yi L, et al. Emergence of almost identical F36: a-:B32 Plasmids carrying blaNDM-5 and qepA in Escherichia coli from Both Pakistan and Canada. Infect Drug Resist. 2019;12:3981–3985. doi:10.2147/IDR.S236766
- Marchetti VM, Bitar I, Mercato A, et al. Complete nucleotide sequence of plasmids of two Escherichia coli Strains carrying and blaNDM-5 and blaOXA-181 from the same patient. Front Microbiol. 2020;10:3095. doi:10.3389/fmicb.2019.03095
- Krishnaraju M, Kamatchi C, Jha AK, et al. Complete sequencing of an IncX3 plasmid carrying blaNDM-5 allele reveals an early stage in the dissemination of the blaNDM gene. Indian J Med Microbiol. 2015;33:30–38. doi:10.4103/0255-0857.148373
- Paul D, Garg A, Bhattacharjee A. Occurrence of blaNDM-1 and blaNDM-5 in a tertiary referral hospital of North India. Microb Drug Resist. 2017;23:815–821. doi:10.1089/mdr.2016.0124
- Hammerum AM, Littauer P, Hansen F. Detection of Klebsiella pneumoniae co-producing NDM-7 and OXA-181, Escherichia coli producing NDM-5 and Acinetobacter baumannii producing OXA-23 in a single patient. Int J Antimicrob Agents. 2015;6:597–598. doi:10.1016/j.ijantimicag.2015.07.008
- Ghatak S, Singha A, Sen A, et al. Detection of New Delhi Metallo-beta-Lactamase and Extended-Spectrum beta-lactamase genes in Escherichia coli isolated from mastitic milk samples. Transbound Emerg Dis. 2013;60(5):385–389. doi:10.1111/tbed.12119
- Singh AS, Lekshmi M, Nayak BB, Kumar SH. Isolation of Escherichia coli harboring blaNDM-5 from fresh fish in India. J Microbiol Immunol Infect. 2016;49(5):822–823. doi:10.1016/j.jmii.2014.11.004
- Yang RS, Feng Y, Lv XY, et al. Emergence of NDM-5- and MCR-1-producing Escherichia coli Clones ST648 and ST156 from a single muscovy duck (Cairina moschata). Antimicrob Agents Chemother. 2016;60:6899–6902. doi:10.1128/AAC.01365-16
- Yang L, Lin Y, Lu L, et al. Coexistence of Two blaNDM-5 genes carried on IncX3 and IncFII plasmids in an Escherichia coli isolate revealed by illumina and nanopore sequencing. Front Microbiol. 2020;11:195. doi:10.3389/fmicb.2020.00195
- Takayama Y, Sekizuka T, Matsui H, et al. Characterization of the IncFII-IncFIB (pB171) plasmid carrying blaNDM-5 in Escherichia coli ST405 clinical isolate in Japan. Infect Drug Resist. 2020;13:561–566. doi:10.2147/IDR.S232943
- Sarkar A, Pazhani GP, Chowdhury G, Ghosh A, Ramamurthy T. Attributes of carbapenemase encoding conjugative plasmid pNDM-SAL from an extensively drug-resistant Salmonella enterica Serovar Senftenberg. Front Microbiol. 2015;6:969. doi:10.3389/fmicb.2015.00969