
Abstract
Objective
Klebsiella pneumoniae is a common multidrug-resistant pathogen that jeopardizes the health of hospitalized patients. We aimed to study the phenotypic and genotypic characteristics of carbapenem-resistant K. pneumoniae (CRKP) isolates from a hospital in Beijing.
Methods
Twenty-four CRKP clinical isolates were collected within a half-year to investigate antimicrobial resistance and genomic characteristics. Illumina and Nanopore sequencing were performed to assemble and annotate genomes.
Results
All strains were multi-drug resistant. Twenty-two strains carried the blaKPC-2 gene and two harbored blaNDM-5. Multilocus sequence type(MLST) analysis identified five sequence types; most isolates belonged to ST11. Three strains were isolated from the same patient; each carried a different plasmid replicon, either IncFII (pHN7A8), IncX, or IncFIB (K).
Conclusion
This study furthers the understanding of CRKP antimicrobial resistance genotypes, and may facilitate the control of nosocomial infections caused by antimicrobial-resistant pathogens.
Introduction
Carbapenem-resistant Klebsiella pneumoniae (CRKP) comprise a set of Gram-negative bacteria that cause serious nosocomial infections, including pneumonia, meningitis, liver abscess, wound and urinary tract infections, especially in immunodeficient patients.Citation1,Citation2 The rising prevalence and increasing multidrug resistance of CRKP are of critical concern.Citation3 A World Health Organization report on antimicrobial resistance disclosed that CRKP has a worldwide geographic distribution, and that the prevalence of CRKP among K. pneumoniae isolates exceeds 50% in some patient groups.Citation4 For example, carbapenem resistance among K. pneumoniae isolates in Greece increased from 0% in 2003 to 38.3% in 2010.Citation5 Surveillance reports from CHINET show that the prevalence of meropenem resistance among K. pneumoniae isolates in China has increased annually, from 2.9% in 2005 to 26.3% in 2018.Citation6
Carbapenems are used to treat severe infections caused by multidrug-resistant Enterobacteriaceae (including K. pneumoniae). The emergence of carbapenemases and extended-spectrum beta-lactamases (ESBL) in K. pneumoniae has reduced bacterial sensitivity to almost all beta-lactams,Citation7 which confounds treatment and jeopardizes clinical outcomes.
blaKPC-2 is the major carbapenemase gene in Enterobacteriaceae isolated from Chinese adults and children.Citation6,Citation8 blaNDM-positiveK. pneumoniae isolates often produce metallo-β-lactamaseCitation9 and demonstrate high-level resistance to all carbapenems.Citation10 Understanding the types and distribution of K. pneumoniae antibiotic resistance genes (ARGs) facilitates hospital infection control and deserves attention.Citation11
We employed Illumina and Nanopore sequencing to analyze the genomic epidemiology of CPKP clinical isolates. Our findings characterized the genomic and resistance features of K. pneumoniae and revealed an evolving genotype in isolates from a single patient, and may guide prevention and treatment.
Methods
Bacterial Classification and Antimicrobial Susceptibility Testing
Samples were collected in a tertiary hospital in Beijing, China, from June to December 2021. Twenty-four CRKP isolates were obtained from sputum (11/24), urine (7/24), blood (3/24), catheters (1/24), secretions (1/24) and throat swabs (1/24). The samples were collected from 15 (62.5%) men and 9(37.5%) women between 50 and 94 years old. A fully automated analysis system (PHOENIX-100 [BD, USA]) was used for species identification and antimicrobial susceptibility testing. Susceptibilities to 18 commonly used antibacterial agents were determined; these included ampicillin, amoxicillin/clavulanate, ampicillin/sulbactam, piperacillin/tazobactam, cefazolin, cefepime, cefotaxime, ceftazidime, meropenem, imipenem, gentamicin, amikacin, ciprofloxacin, moxifloxacin, levofloxacin, trimethoprim/sulfamethoxazole, tetracycline, and aztreonam. Results were interpreted according to the breakpoints and reporting method of the Clinical Laboratory Standards Institute (2021 version, https://clsi.org).
Whole-Genome Sequencing Using Illumina and Nanopore
Genomic DNA was extracted from cultured bacteria. The Illumina sequencing library was prepared using NEBNext Ultra II FS DNA Library Prep Kit for Illumina (New England BioLabs, CA, USA) according to the manufacturer’s instructions. Sequencing was performed on the Illumina NovaSeq 6000 platform at Novogene company with a strategy of pair-end 150bp. Nanopore sequencing libraries were prepared using the Ligation Sequencing Kit SQK-LSK109 (Oxford Nanopore Technologies, Cambridge, UK). The prepared libraries were sequenced on a MinION MK1B sequencer (Oxford Nanopore Technologies, Cambridge, UK) with an R9.4 flow cell.
Dry Lab: Bioinformatics Analysis
The term dry lab denotes the bioinformatic portion of the technology.Citation12 Illumina and Nanopore sequencing data were filtered by FastpCitation13 (version 0.19.7) and NanoFilt (v2.8.0). Hybrid assembly of clean data was conducted with Unicycler (v0.4.8).Citation14 The ARGs of CRKP were identified using the ABRicate (1.0.1) and ResfinderCitation15 databases (https://cge.food.dtu.dk/services/ResFinder/). Kleborate (v2.0.1)Citation16 was used for multilocus sequence typing (MLST). Plasmids were identified using the PlasmidFinderCitation17 database. Genomes were annotated using Prokka (1.14.6).Citation18 Core genome alignments were created by Roary,Citation19 and maximum likelihood phylogenetic trees were generated using RAxML-NGCitation20 (v1.0.3), visualized using FigTree (v1.4.4) and the R ggplot2 package. Plasmid genome circle map visualization using proksee (https://proksee.ca).
Results
CRKP Isolation and Antimicrobial Susceptibility Testing
From June to December 2021, 24 CRKP isolates were obtained from sputum (11/24), urine (7/24), blood (3/24), catheters (1/24), secretions (1/24), and throat swabs (1/24). Specimens were primarily from the Intensive Care Unit (ICU) (17/24), followed by neurology (3/24), neonatology (2/24) and nephrology (2/24) wards. All strains were ESBL-positive. Three strains (BJ10-0010, BJ10-0013 and BJ10-0015) were isolated from urine, blood, and sputum of the same ICU patient in June, September, and October 2021, respectively. The samples were collected from 15 (62.5%) men and 9(37.5%) women between 50 and 94 years old. Most strains were resistant to multiple agents, including meropenem and imipenem (). All 24 strains (100%) were resistant to ampicillin, cefazolin, amoxicillin/clavulanate, ampicillin/sulbactam, meropenem, piperacillin/tazobactam, cefepime, cefotaxime and ceftazidime, but most (66.67%) were sensitive to amikacin. Interestingly, the three strains from the same patient showed slightly different levels of imipenem resistance. The MIC values of the 22 strains are provided in Supplementary Table S1.
Table 1 Antimicrobial Susceptibility of K. pneumoniae Isolates
Genomic Characterization of CRKP
The genomes of 24 CRKP strains were obtained by hybrid assembly using Illumina and nanopore sequencing, and have been deposited in NCBI under the BioProject accession number PRJNA874878. Whole genome MLST identified five sequence types. Most strains belonged to ST11 (19/24), and others belonged to ST15 (2/24), ST37 (1/24), ST403 (1/24) and ST1530 (1/24). Genomic annotation identified diverse carbapenemase- and β-lactamase-encoding ARGs. The blaLAP-2 (13/24), blaCTX-M-65 (7/24) and blaSHV-12 (7/24) genes were chromosomal, while blaKPC-2 (13/24), blaTEM-1B (12/24), blaOXA-1 (8/24), blaNDM-5 (2/24) blaSHV-12 (2/24) and blaCTX-M-3 (1/24) blaCTX-M-65 (1/24) were plasmid-encoded. The distribution of ARGs of strains of different STs is shown in .
Figure 1 Distribution of antibiotic resistance genes, plasmids and STs of all strains. Colors represent different clades. Red represents subclade 1, blue represents subclade 2, and purple represents subclade 3. The antique white squares represent corresponding ARGs or replicons.
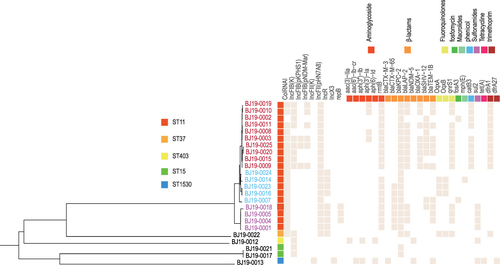
Distribution of Antibiotic Resistance Genes Among Plasmid Replicon Types
Plasmid incompatibility groups were determined according to replication sequences. The strains contained multiple plasmid replicon types, most of which distributed in ST11 strains. All ST11 strains containing plasmid replicons IncFII (pHN7A8). IncFIB (K), IncFIB (pKPHS1) or IncFIB (pNDM-Mar) were distributed exclusively in subclade1, while IncR was carried only in subclades 2 and 3. Twenty-five plasmids contained single replicons, and 14 carried multiple replicons. On average, each single-replicon plasmid carried 1.52 ARGs, and each multi-replicon plasmid encoded 2.71 ARGs. Plasmids containing multiple-replicons IncFIB(K)/IncFII(pHN7A8) and IncFIB(K)/IncFIB(pKPHS1) carried blaKPC-2, blaTEM-1B, and rmtB. The plasmids of BJ10-0013 included the IncFII (K) replicon and encoded eight ARGs, the most of all plasmids.
Comparison of BJ19-0010, BJ19-0013 and BJ19-0015 from a Single Patient
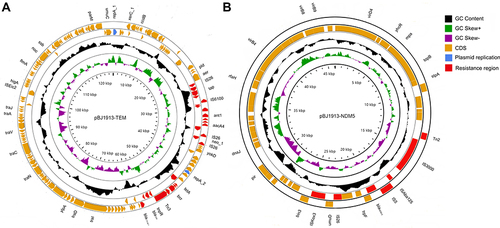
These three strains were isolated from different samples from the same patient. Interestingly, BJ19-0010 and BJ19-0015 belonged to ST11 and were resistant to ciprofloxacin and levofloxacin, while strain BJ19-0013 belonged to ST1503 and was fluoroquinolone-sensitive. In the phylogenetic tree, BJ19-0010 and BJ19-0015 fell in subclade 1, while BJ19-0013 comprised a distinct branch. Strains BJ19-0010, BJ19-0013 and BJ19-0015 had diverse plasmid replicons. Interestingly, BJ19-0010 and BJ19-0015 shared IncFII (pHN7A8), while BJ19-0010 carries IncFIB(K)/IncFIB(pNDM-Mar) and BJ19-0015 carries IncFIB (K). Both BJ19-0010 and BJ19-0015 carried blaKPC-2, blaLAP-2 and blaOXA-1. The blaKPC-2 and blaOXA-1 genes were plasmid-encoded, while the blaLAP-2 gene was located on the chromosome. Strain BJ19-0010 also contained two copies of blaTEM-1B that were located on different plasmids. However, BJ19-0013 carries IncX3 and IncFII(K), which are different from the other two strains. The hybrid assembly showed that the IncFII(K) plasmid pBJ1913-TEM had a length of 127942bp and carries the β- Lactams ARGs blaTEM-1B and blaCTX-M-3(). The other plasmid, pBJ1913-NDM5, had a length of 46068bp and carries the carbapenem-resistance gene blaNDM-5 (), which is nearly identical with plasmid pR15_NDM-5 in Escherichia coli strain R15 isolated from wastewater treatment plant in China.
Figure 2 The genome circle map of plasmids pBJ1913-TEM (A) and pBJ1913-NDM5 (B) carried by strain BJ19-0013. The blue color in the outer circle represents the plasmid replicon protein, the red color represents the drug resistance gene region, and the Orange color represents the protein coding region. The inner-circle indicates the positive and negative GC skew of the plasmid in green and purple, respectively.
Discussion
Carbapenems are usually effective for the treatment of drug-resistant bacterial infections. However, CRKP can express plasmid-encoded carbapenemases, which inactivate most β-lactam antibiotics and confer carbapenem resistance.Citation21 Our study showed that 24 CRKP strains were resistant to most tested drugs. The resistance rate to fluoroquinolones was relatively high, which was consistent with the results of a previous study.Citation22 Three strains were isolated from different samples from the same patient within 4 months and carried different carbapenemase genes, which were located on separate plasmids. Genome annotation revealed that Tn3 was located upstream of the blaKPC-2 gene on chromosomes of BJ19-0010 and BJ19-0015, while Tn552 was located downstream of blaNDM-5 in BJ19-0013. The Tn3 transposon promotes the dissemination of AMR,Citation23 and Tn552 has a co-integrate resolution system homologous to Tn3 family elements,Citation24 which may have facilitated plasmid and ARG exchange among these three strains. A previous study showed that blaKPC−2-positive K. pneumoniae isolates in China belonged primarily to ST11.Citation25 In this study, the majority of CRKP strains belonged to ST11, all of which contained blaKPC-2. Most (68.42%, 19/24) ST11 strains were isolated from ICU patients; consequently, the monitoring of AMR strains in ICUs should be strengthened. In K. pneumoniae, the plasmid with an IncR replicon was identified as a multidrug resistance vector with variable ARGs.Citation26 ST11 strains in this study contained the IncR and IncFII (pHN7A8) replicons with blaCTX-M-65, blaTEM-1B, blaKPC-2, and rmtB genes.
Conclusion
Our study revealed the prevalence of CRKP strains isolated primarily from ICU patients. All 24 strains (100%) were resistant to β- Lactams and carbapenems and carried diverse plasmids with different ARGs. The rapid dissemination of ST11 strains highlights the necessity of strengthening CRKP surveillance and may provide a theoretical basis for infection control and treatment.
Data Sharing Statement
The genomes of 24 CRKP strains have been deposited in NCBI with the BioProject accession number PRJNA874878.
Ethics Approval and Informed Consent
The institutional ethics committees of Dong zhimen Hospital Beijing University of Chinese Medicine approved the study. As all data were anonymously collected and interpreted, the institutional ethics committees waived the need for written informed consent from the participants. The study was also supervised by Chinese PLA Center for Disease Control and Prevention.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
Additional information
Funding
References
- Paczosa MK, Mecsas J. Klebsiella pneumoniae: going on the offense with a strong defense. Microbiol Mol Biol Rev. 2016;80(3):629–661. doi:10.1128/MMBR.00078-15
- Ahmadi M, Ranjbar R, Behzadi P, et al. Virulence factors, antibiotic resistance patterns, and molecular types of clinical isolates of Klebsiella Pneumoniae. Expert Rev Anti Infect Ther. 2022;20(3):463–472. doi:10.1080/14787210.2022.1990040
- Lee CR, Lee JH, Park KS, et al. Global dissemination of carbapenemase-producing Klebsiella pneumoniae: epidemiology, genetic context, treatment options, and detection methods. Front Microbiol. 2016;7:895. doi:10.3389/fmicb.2016.00895
- Antimicrobial resistance, global report on surveillance; 2014. Available from: https://www.who.int/publications/i/item/9789241564748. Accessed December 14, 2022.
- Zagorianou A, Sianou E, Iosifidis E, et al. Microbiological and molecular characteristics of carbapenemase-producing Klebsiella pneumoniae endemic in a tertiary Greek hospital during 2004–2010. Euro Surveill. 2012;17(7):20088.
- China antimicrobial surveillance network, 2021.Available from: http://www.chinets.com/Data/AntibioticDrugFast. Accessed December 16, 2022.
- Nordmann P, Cuzon G, Naas T. The real threat of Klebsiella pneumoniae carbapenemase-producing bacteria. Lancet Infect Dis. 2009;9(4):228–236. doi:10.1016/S1473-3099(09)70054-4
- Han R, Shi Q, Wu S, et al. Dissemination of carbapenemases (KPC, NDM, OXA-48, IMP, and VIM) among carbapenem-resistant Enterobacteriaceae isolated from adult and children patients in China. Front Cell Infect Microbiol. 2020;10:314. doi:10.3389/fcimb.2020.00314
- Behzadi P, García-Perdomo HA, Karpiński TM, et al. Metallo-ß-lactamases: a review. Mol Biol Rep. 2020;47(8):6281–6294. doi:10.1007/s11033-020-05651-9
- Fu B, Yin D, Sun C, et al. Clonal and horizontal transmission of bla(NDM) among Klebsiella pneumoniae in children’s intensive care units. Microbiol Spectr. 2022;10(4):e0157421. doi:10.1128/spectrum.01574-21
- Ahmadi Z, Noormohammadi Z, Ranjbar R, et al. Prevalence of tetracycline resistance genes tet (A, B, C, 39) in Klebsiella pneumoniae isolated from Tehran, Iran. Iran J Med Microbiol. 2022;16(2):141–147. doi:10.30699/ijmm.16.2.141
- Behzadi P, Ranjbar R. DNA microarray technology and bioinformatic web services. Acta Microbiol Immunol Hung. 2019;66(1):19–30. doi:10.1556/030.65.2018.028
- Chen S, Zhou Y, Chen Y, et al. Fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 2018;34(17):i884–i890. doi:10.1093/bioinformatics/bty560
- Wick RR, Judd LM, Gorrie CL, et al. Unicycler: resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput Biol. 2017;13(6):e1005595. doi:10.1371/journal.pcbi.1005595
- Zankari E, Hasman H, Cosentino S, et al. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012;67(11):2640–2644. doi:10.1093/jac/dks261
- Lam MMC, Wick RR, Watts SC, et al. A genomic surveillance framework and genotyping tool for Klebsiella pneumoniae and its related species complex. Nat Commun. 2021;12(1):4188. doi:10.1038/s41467-021-24448-3
- Carattoli A, Zankari E, García-Fernández A, et al. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob Agents Chemother. 2014;58(7):3895–3903. doi:10.1128/AAC.02412-14
- Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014;30(14):2068–2069. doi:10.1093/bioinformatics/btu153
- Page AJ, Cummins CA, Hunt M, et al. Roary: rapid large-scale prokaryote pan genome analysis. Bioinformatics. 2015;31(22):3691–3693. doi:10.1093/bioinformatics/btv421
- Kozlov AM, Darriba D, Flouri T, et al. RAxML-NG: a fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics. 2019;35(21):4453–4455. doi:10.1093/bioinformatics/btz305
- Jeon JH, Lee JH, Lee JJ, et al. Structural basis for carbapenem-hydrolyzing mechanisms of carbapenemases conferring antibiotic resistance. Int J Mol Sci. 2015;16(12):9654–9692. doi:10.3390/ijms16059654
- Ahmadi Z, Noormohammadi Z, Behzadi P, et al. Molecular detection of gyrA mutation in clinical strains of Klebsiella pneumoniae. Iran J Public Health. 2022;51(10):2334–2339. doi:10.18502/ijph.v51i10.10992
- Nicolas E, Lambin M, Dandoy D, et al. The Tn3-family of replicative transposons. Microbiol Spectr. 2015;3(4). doi:10.1128/microbiolspec.MDNA3-0060-2014
- Rowland SJ, Dyke KG. Tn552, a novel transposable element from Staphylococcus aureus. Mol Microbiol. 1990;4(6):961–975. doi:10.1111/j.1365-2958.1990.tb00669.x
- Qi Y, Wei Z, Ji S, et al. ST11, the dominant clone of KPC-producing Klebsiella pneumoniae in China. J Antimicrob Chemother. 2011;66(2):307–312. doi:10.1093/jac/dkq431
- Yu X, Zhang W, Zhao Z, et al. Molecular characterization of carbapenem-resistant Klebsiella pneumoniae isolates with focus on antimicrobial resistance. BMC Genom. 2019;20(1):822. doi:10.1186/s12864-019-6225-9