Abstract
Background
DOTAP/cholesterol-based lipoplexes are successfully used for delivery of plasmid DNA in vivo especially to the lungs, although low systemic stability and circulation have been reported. To achieve the aim of discovering the best method for systemic delivery of DNA to disseminated tumors we evaluated the potential of formulating DOTAP/cholesterol lipoplexes with a polyethylene glycol (PEG)-modified lipid, giving the benefit of the shielding and stabilizing properties of PEG in the bloodstream.
Method
A direct comparison of properties in vitro and in vivo of 4 different DOTAP/cholesterol-based lipoplexes containing 0%, 2%, 4%, and 10% PEG was performed using reporter gene activity and radioactive tracer lipid markers to monitor biodistribution.
Results
We found that 10% PEGylation of lipoplexes caused reduced retention in lung and heart tissues of nude mice compared to nonPEGylated lipoplexes, however no significant delivery to xenograft flank tumors was observed. Although PEGylated and nonPEGylated lipoplexes were delivered to cells the ability to mediate successful transfection is hampered upon PEGylation, presumably due to a changed uptake mechanism and intracellular processing.
Conclusion
The eminent in vivo transfection potency of DOTAP/cholesterol-based lipoplexes is well established for expression in lung tumors, but it is unsuitable for expression in non first pass organs such as xenograft flank tumors in mice even after addition of a PEG-lipid in the formulation.
Efficient gene delivery in vivo based on nonviral methods remains a major challenge in the field of gene therapy, where an overwhelming variety of polymeric and liposomal compounds have been tested.Citation1 A major obstacle has been the fact that extremely efficient methods involving cationic liposomes for gene delivery to cells in vitro perform very poorly when tested in animals.Citation2 Although a regime of transfection-potent lipoplexes has been established in vitro,Citation3,Citation4 presumably in vivo applications require different physical and chemical properties and limited information about the performance for in vivo delivery has emerged.
A notable exception is 1,2-dioleoyl-3-trimethylammonium-propane/cholesterol (DOTAP/chol)-based lipoplexes, that are potent in cellular transfection and importantly facilitate systemic delivery of DNA to cells in vivo.Citation5–Citation8 DOTAP/chol complexed to plasmid DNA encoding the tumor suppressor gene FUS1 has been used successfully for the treatment of xenograft tumor model of non-small cell lung carcinoma (NSCLC)Citation9 and is currently being exploited in a clinical setting for the treatment of NSCLC patients. When these lipoplexes are administered to end-stage NSCLC patients a positive response has been reported.Citation10
DOTAP/chol-based lipoplexes show attractive properties: i) Easy formulation in that preformed liposomes are simply mixed with plasmid DNA and spontaneously organize into multilaminary vesiclesCitation11 and ii) the preparation of these particles for clinical use has been refined and is FDA approved.Citation12 However suboptimal properties relating to systemic stability have been reported when tested in comparison to other formulations.Citation2,Citation13
To develop a strategy involving transcriptionally targeted suicide gene therapy of small cell lung carcinoma (SCLC),Citation14–Citation16 which at the time of diagnosis often appear disseminated to various extrathoracic organs,Citation17 a systemic distribution of the therapeutic agent is demanded. It remains to be determined if DOTAP/chol is an optimal choice for this malignancy or whether improvements must be made for systemic stability and circulation.
Liposome modification by a shielding poly-ethylene-glycol (PEG) component in the formulation has been reported to enhance the systemic stability greatly.Citation18–Citation22 Furthermore the biophysical relationship between liposome surface concentration and structure of PEG has been characterized in detail.Citation23,Citation24 Recent reports about the delivery of plasmid and siRNA in vivo using cationic lipoplexes that also contain a PEG-lipid for improved efficiencyCitation19,Citation25,Citation26 prompted us to assess the usability of different degrees of PEGylation of DOTAP/chol/plasmid DNA lipoplexes in relation to improved systemic circulation and stability while maintaining the favorable transfection properties.
Hence, in the present study we aimed to evaluate the effect of increasing PEGylation of DOTAP/chol lipoplex-mediated gene delivery in the form of plasmid DNA and characterize the biophysical properties in relation to systemic stability, biodistribution, and uptake mechanism. Using enhanced green fluorescence protein (EGFP) and firefly luciferase (LUC) reporter genes, endocytosis inhibitors, and labeled lipids we investigate the potential use for systemic tumor delivery and assess the barriers operating at the level of intracellular uptake.
Material and methods
Materials
All chemicals eg, synthetic cholesterol were purchased from Sigma-Aldrich Inc. (Brøndby, Denmark) unless otherwise stated. DOTAP (N-[1-(2,3-dioleyl)propyl]-N,N,N-trimethylammonium chloride), DSPE-PEG2000 (1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N-[methoxy(polyethylene glycol)-2000] and 25-NBD-cholesterol (25-[N-[(7-nitro-2-1,3-benzoxadiazol-4-yl)methyl] amino]-27-norcholesterol) were purchased from Avanti Polar Lipids Inc. (Alabaster, AL, USA). [1,2-3H]-Cholesteryl Hexadecyl Ether (3H-CHE) was purchased from Perkin Elmer (Skovlunde, Denmark). High quality plasmids: pCMV-LUC (sequence available upon request) and pEGFP-N1 (Clontech, Mountain View, CA, USA) preparations were made with the endo-free Giga kit from Qiagen GmbH (Hilden, Germany) according to the manufacturer’s instructions. Glassware used for lipid work was washed and rinsed in milliQ water, baked at 180°C for 6 hours and dust was removed using pressurized air. Lipid solutions in chloroform were handled with gastight glass syringes (Hamilton, VWR, Herlev, Denmark) reserved for this purpose. Syringes were rinsed with chloroform and 20% ethanol in water only.
The H1299 and NCI-H69 cell lines (obtained from ATCC, Boras, Sweden) were cultivated in RPMI medium supplemented with pen-strep and 10% fetal calf serum (Invitrogen Inc., Taastrup, Denmark). Six week old male NMRI mice were bought from Taconic Europe (Lille Skensved, Denmark) and housed at Department of Experimental Medicine, University of Copenhagen. All animal experiments were performed according to ethical guidelines and under valid license from the Danish Animal Experimentation Board.
Method
Preparation of DOTAP/cholesterol/DSPE-PEG2000 liposomes
The protocol described by RameshCitation7 for preparation of DOTAP/chol liposomes at the 300 μmoL scale was followed with minor modifications. Briefly, equimolar amounts (150 μmoL each) of the lipids were dissolved and mixed in chloroform and placed in a rota-vaporator (BÜCHI Labortechnik AG, Flawil, Switzerland) in a 1 L round glass bottle. The solvent was evaporated under a nitrogen gas stream and the lipid film was dried by high-vacuum for several hours. Hydration was done by adding 7 mL glucose (5%) solution resulting in a 40 mM total lipid concentration and placed at 50°C for 30 minutes with repeated rotary movement to ensure complete hydration of the lipids and left overnight at room temperature. The next day the liposome preparation was placed in a metal basket and sonicated for 5 minutes at 50°C using a Bransonic water bath (MT-1510, 42 kHz, 80 W, setting “sonics”, Branson Ultrasonics, Danbury, CT, USA) and then downsized using 11 passes in a small-scale extruder (Avanti Polar) with polycarbonate nanopore filters (400 nm, 200 nm, and 100 nm, Whatman, Frisenette, Knebel, Denmark) at 50°C. This reagent was designated A-0.
For investigations about the effect of PEGylation, liposomes were prepared at a smaller scale (20 μmoL). Chloroform solutions of DOTAP, cholesterol, and eventually DSPE-PEG2000 and tracer-lipids were mixed in a glass tube (12 × 75 mm) according to the scheme in . While vortexing the solvent was evaporated under a thin nitrogen gas stream. High-vacuum drying of the lipid film was followed by hydration in 0.47 mL glucose (5%) resulting in a 40 mM total lipid solution. The tubes were sealed and placed at 50°C for 30 minutes with repeated rotary movement to ensure complete hydration of the lipids and left overnight at room temperature. The next day the liposome preparations were sonicated for 2 minutes at 50°C and then downsized using a small-scale extruder (Avanti Polar) as above. Sonication for 5 minutes with this preparation caused clearing of the solution indicative of disrupted liposomes, hence the time was reduced to 2 minutes. MALDI-TOF mass spectrometry analysis of the liposome preparation made at 20 and 300 μmoL scales respectively did not reveal any differences in the composition (TL Andresen, personal communication).
Table 1 Liposome components
DNA/lipoplex formation, in vitro, and in vivo transfection
Adherent H1299 were plated the day prior to the experiment in 6-well plates, with 300,000 cells per well. NCI-H69 cells growing in suspension were single-cell resuspended on the day of the experiment and counted in a hemocytometer using Trypan Blue (0.4%) staining to discriminate from dead cells before placing 2 × 106 cells in 6 well plates. If the effect of endocytosis inhibitors was found in the experiment, the cells were incubated with inhibitors for one hour prior to lipoplex addition. Initially, the cytotoxicity of endocytosis inhibitors: chloroquine (inhibiting acidification of endosomes and lysosome fusion),Citation27 chlorpromazine (inhibiting clathrin-mediated endocytosis),Citation28 filipin (inhibiting caveaolae-mediated endocytosis),Citation29,Citation30 and cytochalasin B (inhibiting macropinocytosis)Citation31 was determined in NCI-H69 and H1299 cells in the range reported previously by others. Concentrations causing approximately 30% decrease in MTT cell viabilityCitation32 were selected (data not shown, see legend of ). Twenty microlitres (0.8 μmoL) of liposome preparations with varying amounts of PEG-lipid were diluted and mixed by rapid pipetting up and down with DNA solutions containing pEGFP-N1 and pCMV-LUC (22.5 μg each) yielding a total volume of 100 μL DNA/lipoplex solution.Citation7 For in vitro studies, after 30 minutes at RT the DNA/lipoplex (2.25 μg/40 nmoL) was added to cells in full growth medium and incubated for 2 days at 37°C before analysis of reporter activity. For in vivo applications the mixing was done with maximally 300 μL of total volume, and batches were pooled if larger amounts were required. Intravenous injection of 100 μL was administered within 2 hours of mixing.
Gene expression analysis
Adherent cells were trypsinized and suspension cells were single-cell resuspended and transferred to Eppendorf tubes in two aliquots and washed with phosphate-buffered saline (PBS). One aliquot was analyzed by flow cytometry using a FACS Canto II instrument (BD Biosciences, Franklin Lakes, NJ, USA) gating EGFP-positive cells using the blue laser (488 nm) and detecting fluorescence light in each cell at 538 ± 10 nm. A nontransfected control cell sample was used to adjust for autofluorescence. The second cell aliquot of each sample was pelleted and lysed in 100 μL passive lysis buffer (Promega Inc., Madison, WI, USA) for 10 minutes. After centrifugation for 1 minute, the supernatant was analyzed for luciferase activity (20 μL; Luciferase kit, Promega) using a luminometer (Lumat LB9507; Berthold, Bad Wildbad, Germany) and total protein concentration (20 μL, 10 times diluted, BCA kit, Pierce/Thermo, Rockford, IL, USA) using an OpsysMR microplate reader (Dynex Technologies GmbH, Berlin, Germany). Using a purified, recombinant firefly luciferase (Promega) for standardization, luciferase activity was expressed as picogram luciferase enzyme per milligram of total protein (pg luc/mg protein). When fluorescing lipid was incorporated in the lipoplex, the cell lysates of transfected cell samples were analyzed in a microplate fluorometer (Ex. 485 ± 10 nm, Em. 530 ± 10 nm, Synergy 2, Biotek Instruments Inc., Winooski, VT, USA).
Particle characterization
DNA/lipoplexes were analyzed by agarose gel (1%) electrophoresis followed by ethidium bromide staining to visualize bound and free plasmid DNA. A Zetasizer Nano ZS (Malvern Instruments Inc., Malvern, UK) was used for characterizing the particle size by dynamic light scattering and zeta potential. Preparations of liposomes and DNA/lipoplexes were diluted to approximately 1 mM total lipid and placed in a clear disposable zeta cell (Malvern). First, size was determined using four cycles of 3 minutes at standard settings for vesicles and with general purpose parameter settings. The quality of size measurements given as the volume-weighted mean diameter were analyzed by evaluating polydispersity index (PDI), scattering correlation, and cumulants fit. Subsequently samples were analyzed for zeta potential of particles using standard settings with three repeated measurements of >20 zeta runs and assessing the quality of measurements by evaluation of the phase plot.
SCLC tumor model establishment
The SCLC xenograft model was established by the injection of 5 × 106 NCI-H69 cells per flank subcutaneously into 6–8-week-old male nude NMRI mice.Citation16 Tumors from injected mice (termed passage 0) were used for serial transplantation of mice that entered experimental protocols (passage 1) or used for serial transplantation of new animals (passage 2). Xenograft tumors used in the experiments were passaged up to four times. Untreated tumors from each passage were subjected to pathological analysis to evaluate the existence of clinically validated SCLC markers.
Luciferase activity, pharmacokinetics, biodistribution in vivo
Male NMRI nude mice were injected in the lateral tail vein with 100 μL lipoplex prepared as described above. Unlabeled lipoplex without PEG-lipid (A-0*) was administered once daily for three consecutive days and on the fourth day animals were euthanized by cervical dislocation and organ samples (tumor, heart, lung, liver, kidney, spleen, and tail (1 cm upward of injection site); 50–150 mg) were isolated and snap frozen. Organ samples were mixed with 1 mL passive lysis buffer (Promega Inc.) supplemented with Protease Inhibitor Cocktail Set III (Merck Chemicals, Glostrup, Denmark) grinded in a ball mill (Qiagen) using one steel ball (5 mm) and shaking for 6 minutes. After centrifugation for 10 minutes at 4°C the supernatant was isolated and luciferase activity and protein concentration was measured as described previously.
Using the lipid marker 3H-CHE,Citation13,Citation33 tritium-labeled DNA/lipoplexes with varying degrees of PEGylation were injected in a single dose (100 μL) containing approximately 1 μCi tritium label. In these experiments half of the homogenate (500 μL) was isolated for scintillation counting before centrifugation. If more than 90% of the counts were found in the tail sample the injection was considered as failed and the mouse was excluded from the experiment. Blood samples (100 μL) were drawn from the eye by periobital plexus puncture after 15 minutes, 2 hours, 5 hours, and 24 hours, and immediately mixed with 10 mL scintillation liquid (UltimaGold, Perkin Elmer, Skovlunde, Denmark) and counted in a Beckman LS 6500 liquid scintillation counter (Beckman Instruments, Fullerton, CA, USA). After the last blood sampling the animals were euthanized and organs sampled and analyzed as above. Due to the low systemic distribution of radiolabel no blood correction factor was applied to organ values.Citation34
Results
Gene expression analysis in vivo
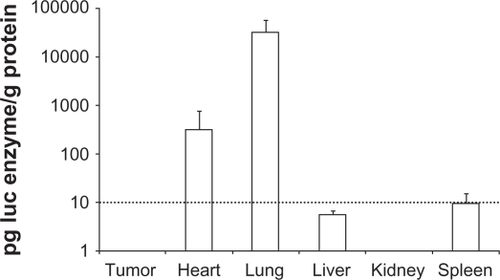
The DOTAP/chol/DNA lipoplex has previously been shown to facilitate gene expression in primarily lung and heart tissue when administered intravenously by tail vein injection to NSCLC models in nude mice.Citation6,Citation9,Citation35,Citation36 We inoculated NCI-H69 cells subcutaneously (s.c.) on the flanks of NMRI nude mice for testing of the ability of the DOTAP/chol lipoplexes to facilitate gene expression in SCLC s.c. xenografts. The tumors reached approximately 200–500 mm3 in size when the experiment was started. Lipoplexes were prepared with DOTAP/chol (formulation A-0*) and pCMV-LUC plasmid and three daily tail vein injections of xenografted nude mice were performed. The animals were sacrificed on the fourth day and selected organs were sampled and luciferase activity was assayed in protein extracts (). We detected luciferase expression in lung and heart tissue, whereas the activity was three to four orders of magnitude lower, below assay background (10 pg luc per g protein) in tumor, liver, kidney, and spleen tissue samples. We interpreted this finding as showing that DOTAP/chol lipoplexes have insufficient systemic stability to reach beyond first-pass organs and speculated that the required improvement in biodistribution might be obtained by adding a PEGylated lipid to the lipoplex formulation.
Figure 1 Luciferase reporter gene expression in vivo. After three consecutive daily tail vein injections of DOTAP/chol/DNA lipoplex (A-0*) in nude mice organs were sampled on the fourth day and assayed for luciferase activity, n = 4. Assay background was at 10 pg luc/g protein (indicated with dotted line).
Liposome particle and lipoplex characterization
In order to investigate the usability of modifying the lipoplex with PEG-polymer, four formulations of 100 nm-extruded DOTAP/chol liposomes designated A-0, B-2, C-4, and D-10, containing 0%, 2%, 4%, or 10% DSPE-PEG2000 lipid respectively, were evaluated. shows the composition of each formulation. Immediately after mixing the liposomes with plasmid DNA the solution became cloudy white indicating lipoplex formation, as observed by others.Citation5,Citation7
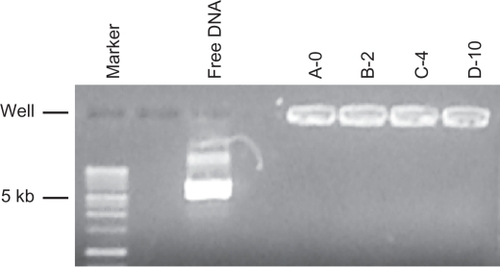
A small aliquot of lipoplex (0.2% of each preparation) was analyzed by agarose gel electrophoresis.Citation37 In agreement with predicted molar excess of lipid, all DNA was complexed to lipoplex, since no DNA was observed migrating as free plasmid DNA ().
Figure 2 Agarose gel electrophoresis/ethidium bromide staining of DNA/lipoplexes with different content of PEG-lipid compared to the migration of free plasmid DNA (free DNA) and a DNA size marker.
The ratio of positively charged lipids to negatively charged DNA was calculated to be 2.6.Citation9 Before and after mixing with DNA the size, polydispersity index (PDI), and charge were assessed using dynamic light scattering and zeta potential measurements (a typical experiment is described in ).
Figure 3 Physical characterization of lipoparticles using a Zetasizer. Each of the four liposome formulations was analyzed before (white columns) and after (grey columns) mixing with plasmid DNA. A) The hydrodynamic size. B) The polydispersity index. C) The zeta potential of the particles. The estimate and the standard error are given for each measurement. Three independent experiments yielded equivalent results.
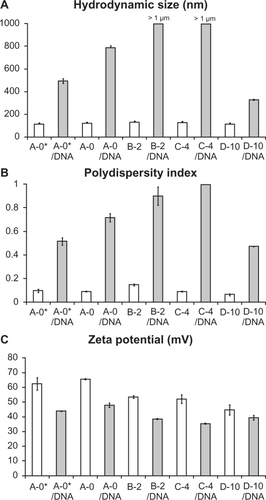
All four liposome preparations have particle sizes in the range 113 ± 0.5–130 ± 0.4 nm and low polydispersity indices (0.066 ± 0.012–0.149 ± 0.01). The zeta potential is gradually decreasing from +65.6 ± 0.74 mV in A-0 to +44.7 ± 3.47 mV in D-10 concurrent with increasing content of the negatively charged DSPE-PEG2000 lipid. The model of PEG-surface structure being mushroom-like at concentrations below 4%, whereas brush-like structures exist at higher PEG-lipid mol-percentages (eg, 10%).Citation23,Citation24 Interestingly, we found that upon mixing with DNA the size of lipoplexes increase and lipoplexes containing 2% or 4% PEG-lipid form micrometer scale aggregates, whereas the A-0 and D-10 lipoplex have sizes 784 ± 49 and 327 ± 5.3 nm, respectively. This finding is in accordance with PDI values that are close to 1 for A-0, B-2, and C-4 lipoplexes indicating high turbidity, whereas D-10 lipoplex has a medium PDI of 0.472. Upon mixing with negatively charged DNA, the zeta potential of the particles is reduced accordingly, but the lipoplex particles/aggregates maintain a high positive charge (ranging from 47.8 ± 1.56 to 35.4 ± 1.58). The properties of lipoplexes described above were consistently found when lipids were prepared at the scale of 20 μmoL in at least three independent experiments, however when preparing the DOTAP/chol particles according to the standard protocolCitation7 at the scale of 300 μmoL (designated A-0* in ) the lipoplexes were measured to be below half a micrometer (492 ± 21 nm) in accordance with previously published results of lipoplexes used for translation investigations.Citation9 Zeta-potential measurements were similar for lipoplexes arranged at both preparation scales (A-0 and A-0*).
Gene expression analysis in vitro
Equal amounts of pCMV-LUC- and pEGFP-N1-expression plasmids encoding LUC and EGFP genes were mixed with A-0, B-2, C-4, or D-10 liposomes and used in transfection of two lung cancer cell lines H1299 or NCI-H69. Forty-eight hours later the cells were analyzed for reporter gene expression, either by measuring the percentage of EGFP-positive cells by flow cytometry () or by measuring the overall amount of luciferase activity expressed in relation to total protein in cell lysates ().
Figure 4 Reporter gene expression by transfection of NCI-H69 (SCLC) and H1299 (NSCLC) cells in vitro. A) EGFP fluorescence in single cells was measured by FACS flow cytometry and expressed as the percentage of EGFP positive cells. *** indicates a significant difference from A-0 (P < 0.001). B) Luciferase activity was measured in cell lysates and expressed as picogram luciferase per milligram of protein. Data from four independent experiments with two different lipid preparations were collected and the average and standard error of the mean are given. *indicates a significant difference from A-0 (P < 0.05).
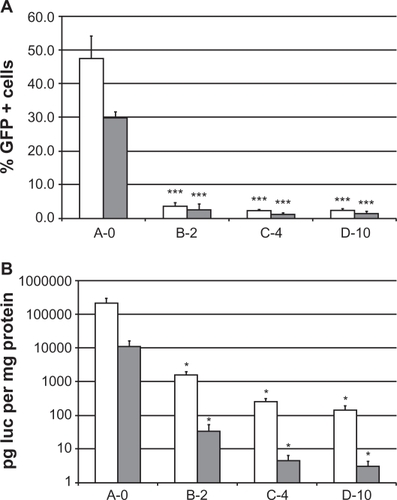
With the presence of PEG-lipid in the lipoplex a dramatic decrease in the number of EGFP-positive cells from 47% ± 6.9% with A-0 to 3.6% ± 1.8% with B-2 in H1299 cells and from 30% ± 1.9% with A-0 to 2.6% ± 1.8% with B-2 in NCI-H69 cells. Transfections with lipoplexes C-4 or D-10 yielded even lower transfection efficiencies around 1% to 2% of the cells being successfully transfected (). In accordance with this, the luciferase activity is gradually decreasing roughly 10-fold in each step from 213 ± 89 ng luc/mg protein with A-0 to 145 ± 47 pg luc/mg protein with D-10 in H1299 cells and 11 ± 5.2 ng luc/mg protein with A-0 to 3.0 ± 1.2 pg luc/mg protein with D-10 in NCI-H69 when analyzing the cell samples transfected with lipoplexes A-0, B-2, C-4, and D-10, respectively ().
Kinetics of cellular uptake
In order to investigate the kinetics of cellular uptake of the lipoplexes the four formulations with increasing PEG-lipid were prepared with 1.5% 25-NBD-cholesterol, a green fluorescing lipid reporter that has previously been used to study liposome transport.Citation38,Citation39 Since the EGFP expression would overlap spectrally with the labeled cholesterol fluorescence, only LUC-expression plasmid was used for lipoplex formation. Essentially the same reporter activities were obtained as with the unlabeled lipoplexes (data not shown). Furthermore, microscopic and FACS flow cytometry analyses showed that for all four mol-percentages of PEG-lipid, virtually all cells (99%) contained the fluorescing cholesterol in similar subcellular compartments two days after transfection () and this prompted us to measure the uptake kinetics on a shorter time scale.
Figure 5 Cellular uptake of fluorescently labeled lipoplexes. A) Bright-field and fluorescence microscope pictures taken two days after H1299 cells were transfected with NBD-labeled A-0 (upper panel) and D-10 lipoplexes (lower panel). Scale bar marked in red indicates 15 μm. B) NBD fluorescence in cell lysates was measured at different time points after addition to cell cultures. Non-PEGylated A-0 lipoplex is rapidly taken up by cells to a saturating level within one hour and remain constant hereafter (fitted line [estimate ± standard error]: Fluorescence index = (1001 ± 102) + (2.8 ± 3.6) × time) Slope confidence interval includes 0. In contrast, PEGylated lipoplexes are gradually taken up over two days. Fitted lines: B-2: Fluorescence index = (626 ± 115) + (15 ± 4.3) x time; C-4: Fluorescence index = (487 ± 71) + (15 ± 2.9) × time; D-10: Fluorescence index = (678 ± 92) + (44 ± 3.7) × time. Slope confidence interval exclude 0. Data are normalized and averaged from triplicates of three independent experiments (average ± SD).
![Figure 5 Cellular uptake of fluorescently labeled lipoplexes. A) Bright-field and fluorescence microscope pictures taken two days after H1299 cells were transfected with NBD-labeled A-0 (upper panel) and D-10 lipoplexes (lower panel). Scale bar marked in red indicates 15 μm. B) NBD fluorescence in cell lysates was measured at different time points after addition to cell cultures. Non-PEGylated A-0 lipoplex is rapidly taken up by cells to a saturating level within one hour and remain constant hereafter (fitted line [estimate ± standard error]: Fluorescence index = (1001 ± 102) + (2.8 ± 3.6) × time) Slope confidence interval includes 0. In contrast, PEGylated lipoplexes are gradually taken up over two days. Fitted lines: B-2: Fluorescence index = (626 ± 115) + (15 ± 4.3) x time; C-4: Fluorescence index = (487 ± 71) + (15 ± 2.9) × time; D-10: Fluorescence index = (678 ± 92) + (44 ± 3.7) × time. Slope confidence interval exclude 0. Data are normalized and averaged from triplicates of three independent experiments (average ± SD).](/cms/asset/b7fb4259-bb43-40d5-819b-b17a6d878ed0/dijn_a_12184590_f0005_c.jpg)
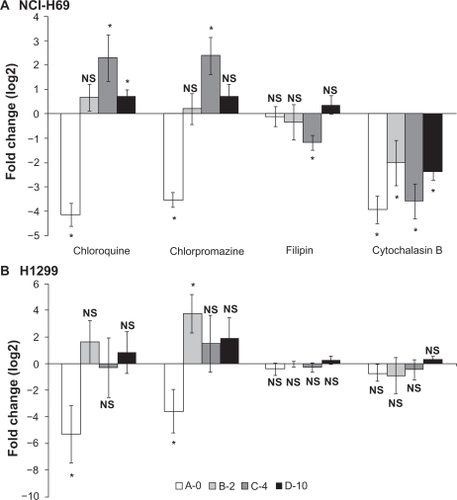
Figure 6 Effects of endocytosis inhibitors on luciferase reporter activity measured in (A) NCI-H69 and (B) 1299 cells. Cells were incubated one hour prior to transfection and then exposed to inhibitors for the entire incubation (48 hours). Chloroquine (20 μM (NCI-H69), 30 μM (H1299)), chlorpromazine 10 μM (NCI-H69), 20 μM (H1299), filipin (4 μM), and cytochalasin B (25 μM). Data from 3–5 independent experiments were pooled and log2 transformed to obtain fold change in expression. Error bars indicate standard error of the mean, *means that 0 is not included in confidence interval (P < 0.05), no change: N.S.
Hence shows the cell lysate fluorescence of NBD-cholesterol at 1, 4, 24, and 48 hours after addition of labeled lipoplexes to H1299 cells. A remarkable difference in uptake kinetics is observed. The fluorescence in cells transfected with lipoplex without PEG-lipid (A-0) is unchanged from one hour after transfection to two days (ANOVA: P = 0.45: no change), whereas fluorescence in cells transfected with lipoplexes with 2%, 4%, or 10% PEG-lipid (B-2, C-4, or D-10) accumulates over time (ANOVA: P values < 0.002 in all three cases: significant change), reaching a maximum of fluorescence after 24 to 48 hours. Regression analysis confirms this difference (linear equation fits are shown in legend).
Mechanism of cellular uptake, effect of endocytosis inhibitors
In order to investigate whether the changed transfectability of PEGylated lipoplexes could be due to an altered uptake mechanism cell culture transfections were done in the presence of four different drugs known to interfere with endocytosis, namely chloroquine that inhibits acidification of endosomes and lysosome fusion,Citation27 chlorpromazine that inhibit clathrin-mediated endocytosis,Citation28 filipin, a sterol binding compound that inhibit caveaolae-mediated endocytosisCitation29 and cytochalasin B that inhibit actin polymerization and macropinocytosis,Citation31 all of which have been described as being useful in relation to nonviral gene transfer.Citation40 Luciferase reporter activity was evaluated in H1299 and NCI-H69 cells and most profound effects were observed in NCI-H69 cells ().
Here it was found that all lipoplexes were sensitive to cytochalasin B decreasing luciferase activity between 4- and 16-fold (22–24-fold) indicating that the uptake mechanism involves macropinocytosis or that actin filaments are involved in intracellular processing. Only the transfection with the largest particles made with DNA/C-4 lipoplexes was decreased two fold by filipin indicative of a caveolae-dependent uptake. With regards to chloroquine and chlorpromazine a large difference in sensitivity was observed between non-PEGylated and PEGylated lipoplexes. A-0-based lipoplexes yielded roughly 16-fold less reporter activity in the presence of these inhibitors, and in contrast hereto, PEGylated lipoplexes made with C-4 yielded four fold more reporter activity, and D-10-based lipoplexes facilitated 1.6-fold more luciferase activity in the presence of chloroquine. In adherent H1299 cells (), effects of inhibitors were less significant, although the same pattern of lipoplex sensitivity could be recognized from that of NCI-H69 cells. Chloroquine and chlorpromazine reduced luciferase activity 16- and 32-fold, respectively in case of A-0 lipoplex, whereas B-2 lipoplexes yielded 16-fold more activity when the cells were exposed to chlorpromazine. Although the exact action of inhibitors may be uncertain, taken together these data strongly suggest that the cellular uptake of naked and PEGylated lipoplexes depend on different mechanisms. All experiments were averaged from 3–5 independent experiments and at least two different preparations of liposomes.
Biodistribution and stability
Discrepancies between transfection in vitro and in vivo has been reported previouslyCitation41 and the finding that PEGylated DOTAP/chol lipoplexes are taken up efficiently over time by cancer cells in vitro, albeit gene expression is hampered, prompted us to investigate the potential of the lipoplexes formulated with different moL-percentage of PEG-lipid in vivo. The intracellular processing of lipoplexes is different in various organs and cell types,Citation40 and the putative stabilizing effect of PEG-lipid in the formulation could influence the effectiveness in cellular transfection between the four formulations in vivo. Using the nonexchangeable and nonmetabolizeable lipid marker 3H-CHE,Citation33 lipoplexes were prepared and injected in the tail vein of tumor-bearing NMRI nude mice and blood was drawn by periobital plexus sampling 15 minutes, 2 hours, 5 hours, and 24 hours after injection of lipoplex.
The radioactive dose was 0.5 μCi corresponding to approximately 1.2 ×107 cpm (background 9 cpm). Interestingly, when 100 μL blood samples from animals were counted by scintillation, only small amounts of tritium were found, indicating a poor systemic spreading of the radioactive dose to the vein blood drawn from the eye. shows the tritium counts of isolated blood samples over time after background subtraction. Blood sampled 15 minutes after tail vein injection of lipoplexes with 0%, 2%, or 4% PEG-lipid contained tritium only 6 to 9 times over background, whereas samples drawn from animals injected with D-10 lipoplex contained approximately 18 times over background, however in all cases, only a very small fraction of the injected dose. Five hours after injection all blood samples contained the same low amount of radioactivity.
Figure 7 A) Blood availability of tritium-labeled lipoplexes with PEGylation. Following tail vein injection of lipoplexes blood samples were drawn by periobital plexus puncture (eye vein blood) after 15 minutes, 2 hours, 5 hours, and 24 hours, and the amount of radioactivity was quantified by scintillation counting. B) Biodistribution of luciferase activity. Mice injected with tritium-labeled DNA/lipoplex with different degrees of PEGylation were sacrificed after 24 hours and samples from organs were analyzed for luciferase activity. Background level in assay is 10 pg/g protein (indicated with dotted line). C) Biodistribution of radioactivity. Tritium counts were expressed as CPM per gram organ sample weight (CPM/g) and the relative distribution in tumor, heart, lung, liver, kidney, and spleen was calculated. Error bars indicate standard deviations. In each group, n = 3–5. A two sided t-test of accumulation in heart/lung samples between A-0 and D-10 results in P = 1.9E-05 (***).
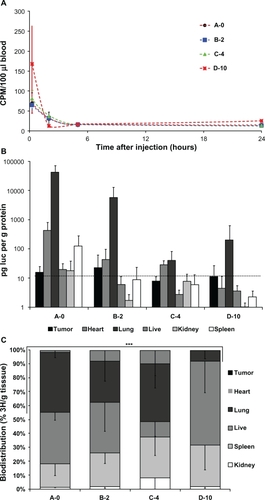
After 24 hours the animals were sacrificed and selected organs were sampled and assayed for luciferase activity and accumulation of tritium-tracer. shows the luciferase activity in tumor, heart, lung, liver, kidney, and spleen samples from animals injected with a single dose of lipoplex formulated with 0%, 2%, 4%, or 10% PEG-lipid. In general, only heart and lung tissue of animals receiving plasmid DNA in lipoplexes contain luciferase activity considerably above assay background, and this was gradually decreasing with increasing PEGylation. D-10 lipoplexes yielded more luciferase activity in the lung than C-4, however no activity in heart tissue could be measured. Additionally, the spleen sample from mice injected with A-0 lipoplex showed luciferase activity above background. Importantly, no luciferase activity was measured in tumor tissue from mice injected with lipoplex.
The relative distribution of radioactive lipid tracer in tumor, heart, lung, liver, kidney, and spleen tissues is shown in and there is a gradual change with high accumulation in the heart and lungs in case of lipoplex A-0, B-2, and C-4 to high liver accumulation of the lipoplex containing 10% PEG-lipid (D-10).
Discussion
Effect of PEGylation in vitro
Previous efforts in nonviral gene delivery using DOTAP/chol-based lipoplexes have proven successful in obtaining efficient cellular transfection in vitro and in vivo using xenograft subcutaneous flank tumors by intratumoral injection or orthotopic micrometastatic lung tumor model of NSCLC by intravenous injection.Citation6 We recently reported the successful use of DOTAP/chol lipoplexes for delivery of suicide gene therapy by intratumoral injection.Citation16 The rapid clearance from the system of intravenously injected DOTAP/chol-based lipoplexes have been reported,Citation13,Citation42 and since our gene therapy strategy is aimed at the systemic treatment of subcutaneous xenograft tumorsCitation14 and eventually disseminated SCLC, in this study we sought to stabilize and increase the circulation time of DOTAP/chol-based lipoplexes that have been modified with different amounts of PEG-lipid, ie, without abolishing the eminent ability to transfect cells.Citation43,Citation44 Several other works have reported the effect of PEGylation of lipoplexes made from DOTAP or other cationic lipid and with DOPE as the helper lipid,Citation25,Citation26,Citation45–Citation47 and we discuss these in relation to our findings with PEGylation of DOTAP/chol-based lipoplexes further below.
A remarkable threshold in physical properties appears to exist at 5% PEG-lipid in lipoplexes. PEG-lipid content of a few to 4% causes aggregation and micrometer-sized particles whereas 5% to 10% of PEGylated particles remain smaller. We applied the lipoplex conditions of RameshCitation7 which utilize a P/N ratio of 2.6, whereas others have used less DNA.Citation25,Citation48 Our findings are in alignment with the recently reported result of Mignet et alCitation26 who tested several percentages of DSPE-PEG2000 in lipoplexes and found that large aggregates occur in case of 1.25% and 2.5%, but not 5% PEG-lipid in the formula. Similarly, a size stabilizing effect exist when more than 4–5 moL% of a mono-alkyl- or cholesteryl-anchored PEG-lipid is incorporated into lipoplexes at neutral charge ratio.Citation47
The biophysical basis of this difference may rely on the surface density of PEG-lipid that in less than 5% allow for a mushroom-structured PEG-lipids where PEG-polymers do not interact.Citation23,Citation24 However, upon lipoplexing where negatively charged DNA is added, PEG-lipids may interdigit with PEG of other particles and hence favor aggregation.
Since 5% to 10% PEG on liposome surface forms brush-like structure it seems an attractive model that mushroom-structured PEG on the cationic liposome surface will lead to aggregated lipoplexes and brush like-structured PEG lead to more stable structures. It may be speculated that all PEG-lipid is not present in the lipoplex surface or even that some may form micelles, since the effect of polymer-based micelle formation has been reported when attempting to incorporate high mol% of DSPE-PEG2000 in liposome formulation.Citation49 However, in this theoretical study it was predicted that 10% PEG2000-lipid would incorporate into liposome membrane spontaneously, whereas longer PEG-chains might form micellar structures. The effect of inserting saturated lipid-PEG components in saturated membranes has been studied by spin-label electron spin resonance (ESR) spectroscopy.Citation50 To our knowledge it has not been shown to what extent saturated PEG-lipids (DSPE-PEG2000) form micelles or pack into an unsaturated DOTAP/“bulky” lipid (cholesterol) membrane, but the aforementioned studies render it likely that although some PEG-lipid may be buried into the lamellar lipoplex structure, all PEG-lipid is retained in the lipoplex. Furthermore, the fact that the zeta potential of DOTAP/chol/DSPE-PEG2000 liposomes with 10% PEG-lipid is lower than 4% PEG-lipid (, white columns), presumably due to the negative charge in the PE-head group, support that liposome composition is in accordance with the expected from the formulation.
When analyzing lipoplexes in vitro we found an inverse correlation between the amount of PEG-lipid in the lipoplex and reporter gene activity in both easy-to-transfect, adherent lung cancer H1299 cells and hard-to-transfect, suspension lung cancer NCI-H69 cells. Luciferase activity measurements showed that each stepwise increase in PEG-lipid percentage roughly reduced the luciferase activity by 10-fold, or three orders of magnitude. In accordance, the percentage of EGFP expressing cells was reduced one to two orders of magnitude to a level just above the detection limit. Using DOPE as the helper lipid several researchers have demonstrated a similar negative influence of increasing PEG-lipid in lipoplex formulation.Citation45,Citation46 Nicolazzi et al studied PEGylated lipoplexes with various charged peptide linkers in the PEG-lipid and confirmed the negative influence on transfection efficiency when analyzing lipoplexes with 5% PEG-lipid.Citation25 Intriguingly, a post insertion method for PEGylation of formed lipoplexes was used that enabled particles of much lower positive charge between 5 and 20 mV compared to 35 to 40 mV for PEGylated lipoplexes in this study, where PEG-lipid was incorporated from the beginning.
It could be speculated that lipoplex uptake mechanism and intracellular release could be altered by PEGylation, since it has been suggested that DOTAP/chol-based lipoplexes enter the cell by direct plasma membrane fusionCitation5 or endocytosis-based upon electrostatic interactions.Citation40,Citation51,Citation52 Using a NBD-cholesterol derivate we labeled the lipoplexes so that the lipid uptake could be monitored fluorometrically and it became evident that the kinetics of cellular uptake differs between PEGylated and non-PEGylated lipoplexes. Using a cell lysate microplate assay it was observed that within the first hour after addition, non-PEGylated lipoplexes enter the cells to a saturating limit. In contrast the uptake of PEGylated lipoplexes is gradually increasing over time and supposedly depends on an endocytotic mechanism that allow the accumulation of higher amounts of fluorescently labeled lipoplex – at least in the case of lipoplexes with 10% PEG-lipid where the fluorescent signal is twice that obtained from cells transfected with non-PEGylated lipoplexes.
This finding was further supported when radiolabeled lipoplexes were tested in cell culture and the label accumulating in cells after two days was measured (data not shown). Interestingly, fluorescence microscopy and flow cytometry show that with all four formulations virtually all cells contain the NBD-label after two days, hence it is indicated that the lost ability of PEGylated lipoplex to successfully transfect cells and allow the expression of reporter gene is due to defective intracellular processing and endocytic release of plasmid DNA. In support of this hypothesis we noted that lipoplexes made with plasmid and DODAC/DOPE/DSPE-PEG2000 showed that increasing mol% of PEG-lipid reduced the transfection after 24 hours.Citation46 However, when analyzing cellular uptake of lipoplexes with radioactive lipids or nucleic acids it was concluded that PEG-induced inhibition of transfection did not correlate with a reduction in endocytosis. In addition, the uptake of oligodeoxynucleotides (ODN) complexed in DODAC/DOPE/PEG2000-ceramide (5%) lipoplexes was investigated and it was found that the stronger uptake correlated with longer ceramide PEG-anchors.Citation45 Furthermore, using confocal microscopy an inverse correlation between ceramide lipid length and successful nuclear transfer of fluorescently labeled ODN was suggested, supporting the notion that DSPE-PEG2000 being C-18-anchored would inhibit endosomal escape similarly in our experiments and that unsuccessful transgene expression from PEGylated lipoplexes relate to that.
A profound change in sensitivity between non-PEGylated and PEGylated lipoplexes towards four endocytosis inhibitors thought to interfere with different uptake pathways was observed. Although the inhibitors may exert nonspecific effects,Citation53 apparently, clathrin-coated endosomes are engaged in transfection using naked lipoplexes, since chlorpromazine reduced luciferase activity 8–16-fold in the lung cancer cell lines in our study and this was also found by others.Citation54 The increase in luciferase activity from PEGylated lipoplexes when cells were exposed to chlorpromazine could be a competition effect of endocytic pathways – one being inhibited and thereby favoring another. Similarly, the sensitivity of A-0 lipoplexes towards chloroquine was also observed in both cell lines and in contrast chloroquine seems to have a stimulating effect on reporter activity from PEGylated lipoplexes. The latter is in alignment with the notion that PEGylated lipo-particles need to unload the DNA cargo before endosome fusion to lysosomes and degradation,Citation55 however the inhibiting effect of chloroquine on transfection with A-0-based lipoplexes was not easily interpretable. Several researchers have demonstrated that gene expression is increased in the presence of chloroquine when using 4% to 10% PEGylated small unilaminary liposomes as delivery vehicle involving passively targetedCitation27 or receptor-targetedCitation28 endocytosis. At least in NCI-H69 cells the sensitivity towards cytochalasin B was significant with all lipoplex formulations and hence a macropinocytosis-dependent uptake mechanism is suggested,Citation56 although it cannot be formally ruled out that the disruption of endosomal trafficking by actin de-polymerization will influence the uptake through an unknown mechanism.Citation31
Difference in inhibitor sensitivity may depend on cell size and growth properties, NCI-H69 being small, growing in suspension, and adherent H1299 that are large and flattened out may require longer preincubation time, hence the cellular uptake of cationic lipoplexesCitation57 and PEGylated lipoparticles may be cell type dependent in relation to endocytotic pathways at work.Citation58
Effect of PEGylation in vivo
The fate of the radiolabel allowed us to assess the systemic availability and retention in various organs. Surprisingly little radioactivity was detectable in blood isolated from the eye, only 0.01% to 0.1% of the injected dose was isolated in 100 μL blood 15 minutes after injection. A mouse weighing 30 g is estimated to contain approximately 3 g bloodCitation59 and hence if the entire dose was freely distributed in the blood and available we would expect 3.3%. One possibility is that due to the high positive charge (>+40 mV) and large aggregate size of the lipoplexes (all >300 nm) they do not make a full systemic passage, but are retained in the first primary organs after intravenous injection. In the heart and lungs all lipoplexes with 0% to 10% PEG-lipid are successful in transfection, but gradually decreasing corresponding to the activities measured in vitro. Comparing the reporter activity measured from lipoplexes made with A-0* in and A-0 in , there is no difference (t-test, P value = 0.35), although the first was applied in three daily injections and the latter only once. We speculate that the duration of expression may not be long or that one dose is saturating in the reachable sites with this intravenous delivery method. Furthermore it is suggested that A-0 lipoplexes being 700 nm in size are as efficient in heart/lung transfection as A-0* lipoplexes.
The positive charge of lipoplexes led to interaction with albumin and other serum proteins when exposed to serum or injected intravenously.Citation48,Citation60 Binding of lipoplexes to serum lipoproteins or complement factor C3 showed that it may have strong influence on particle size, DNA integrity, and transfection ability.Citation60–Citation62 When lipoplex size was assessed in the presence of serum proteins it was found that 5% PEG-lipid prevented a large increase in size over several hours that was observed in the case of non-PEGylated lipoplexes. The difference to the lipoplexes of the present study is that the N/P ratio is lower (1/5, less DNA complexed) and our particle’s surface charge is much higher presumably due to preincorporation of PEG-lipid that may not shield the charge as effectively as in case of postincorporation. Hence it may be amenable to utilize a lower N/P ratio, eg, 1/4 as reported, however that cause a decreased transfection in vivo.Citation48
Opsonization and uptake by phagocytic cells of the reticuloendothelial system (RES) present in blood was therefore expected in case of all four lipoplexes and indeed approximately 20% of the label resides in the spleen, since phagocytic blood cells accumulate later in the spleen as it was reported for liposome particles previously.Citation20,Citation63 Furthermore, luciferase activity found in the spleen when using A-0 lipoplexes could indicate successful transfection of RES cells accumulating in this organ ().
Analysis of lipoplexes with 0%, 2%, 4%, and 10% PEG-lipid allowed us to demonstrate that low PEG-lipid percentage (B-2, C-4) leads to severe lipoplex aggregation causing retention in lung/heart tissue similar to naked lipoplexes (A-0) that is not highly aggregated. Interestingly, with D-10 lipoplexes we observed a distribution in abdominal organs that suggests that heart/lung retention is overcome and although not being able to transfect cells in liver, kidney, and spleen lipoplexes are accumulating here. A similar distribution in inner organs was found when non-transplanted mice were used (data not shown) indicating that the inverse relation between lung/heart accumulation and 10% PEGylation is true also in noncancerous animals. Mice injected with lipoplexes maintained a normal behavior during the experiment, although we measured a significant weight loss of up to 10% in most of the animals. This could relate to some kind of pulmonary obstruction, however this was not visible in H&E staining of tissue sections, that were evaluated by a trained pathologist.
These observations support the notion that PEG shielding of lipo-particles increase systemic circulation,Citation64 however due to the high positive surface charge only in the case of D10 lipoplexes was a small increase in blood availability measured. It has been previously reported that lipoplexes with 5% PEG-lipid could increase the blood availability marginally, however we found no increase in tumor accumulation.Citation25,Citation26 Neither of the lipoplex formulations convey delivery to tumor sites, hence our data confirm previous reports that (non-PEGylated) DOTAP/chol lipoplexesCitation2,Citation13 or cationic DOPE-based lipoplexes including PEG-lipidsCitation25,Citation45,Citation48 do not mediate delivery to xenograft subcutaneous tumors to any useful extend.
Conclusion
We explored the potential of improving systemic stability and circulation of DOTAP/chol-based lipoplexes for in vivo cell transfection by including a PEG-lipid in the formulation. Although we observed a decrease in the retention in lung and heart tissues the systemic circulation was not considerably improved and the eminent properties of providing transfection and ectopic gene expression are lost upon increasing degrees of PEGylation, hereby rendering it unsuitable as a systemic delivery method for plasmid DNA-bound genetic medicine for extrathoracic sites.
Acknowledgements
The authors wish to extend their appreciation to Birgit Guldhammer for pathological evaluation of tissue section, Hanne Mørck Nielsen for access to Zetasizer, and Pia Pedersen for excellent technical assistance. This work was supported in part by a grant from the Danish Cancer Society and the Novo Nordisk Foundation. The authors report no conflicts of interest in this work.
References
- KangHCLeeMBaeYHPolymeric gene carriersCrit Rev Eukaryot Gene Expr20051531734216472063
- KawanoTOkudaTAoyagiHNiidomeTLong circulation of intravenously administered plasmid DNA delivered with dendritic poly(L-lysine) in the blood flowJ Control Release20049932933715380641
- EwertKKAhmadABouxseinNFEvansHMSafinyaCRNon-viral gene delivery with cationic liposome-DNA complexesMethods Mol Biol200843315917518679623
- SafinyaCREwertKAhmadACationic liposome-DNA complexes: from liquid crystal science to gene delivery applicationsPhilos Transact A Math Phys Eng Sci20063642573259616973477
- TempletonNSLasicDDFrederikPMStreyHHRobertsDDPavlakisGNImproved DNA: liposome complexes for increased systemic delivery and gene expressionNat Biotechnol1997156476529219267
- RameshRSaekiTTempletonNSSuccessful treatment of primary and disseminated human lung cancers by systemic delivery of tumor suppressor genes using an improved liposome vectorMol Ther2001333735011273776
- RameshRNanoparticle-mediated gene delivery to the lungMethods Mol Biol200843330133118679632
- SakuraiFTeradaTMaruyamaMTherapeutic effect of intravenous delivery of lipoplexes containing the interferon-beta gene and poly I: poly C in a murine lung metastasis modelCancer Gene Ther20031066166812944985
- ItoIJiLTanakaFLiposomal vector mediated delivery of the 3p FUS1 gene demonstrates potent antitumor activity against human lung cancer in vivoCancer Gene Ther20041173373915486560
- LuCSepulvedaCAJiLSystemic therapy with tumor suppressor FUS1-nanoparticles for stage IV lung cancerAmerican Association for Cancer Research Annual Meeting: Proceedings2007 Apr 14–18Los Angeles, CA Philadelphia (PA)AACR2007 Abstract nr LB-348.2007.
- KoltoverISaldittTSafinyaCRPhase diagram, stability, and overcharging of lamellar cationic lipid-DNA self-assembled complexesBiophys J199977291592410423436
- TempletonNSNonviral delivery for genomic therapy of cancerWorld J Surg20093368569719023615
- TamPMonckMLeeDStabilized plasmid-lipid particles for systemic gene therapyGene Ther200071867187411110420
- ChristensenCLZandiRGjettingTCramerFPoulsenHSSpecifically targeted gene therapy for small-cell lung cancerExpert Rev Anticancer Ther2009943745219374598
- PedersenNMortensenSSorensenSBTranscriptional gene expression profiling of small cell lung cancer cellsCancer Res2003631943195312702587
- ChristensenCLGjettingTPoulsenTTCramerFRothJAPoulsenHSTargeted cytosine deaminase-uracil phosphoribosyl transferase suicide gene therapy induces small cell lung cancer specific cytotoxicity and tumor growth delayClin Cancer Res2010162308231920371678
- ChuaYJSteerCYipDRecent advances in management of small-cell lung cancerCancer Treat Rev20043052154315325033
- CharroisGJAllenTMRate of biodistribution of STEALTH liposomes to tumor and skin: influence of liposome diameter and implications for toxicity and therapeutic activityBiochim Biophys Acta2003160910210812507764
- SantelAAlekuMKeilOA novel siRNA-lipoplex technology for RNA interference in the mouse vascular endotheliumGene Ther2006131222123416625243
- WoodleMCLasicDDSterically stabilized liposomesBiochim Biophys Acta199211131711991510996
- AllenTMCullisPRDrug delivery systems: entering the mainstreamScience20043031818182215031496
- PasutGVeroneseFMPEG conjugates in clinical development or use as anticancer agents: an overviewAdv Drug Deliv Rev2009611177118819671438
- KenworthyAKHristovaKNeedhamDMcIntoshTJRange and magnitude of the steric pressure between bilayers containing phospholipids with covalently attached poly(ethylene glycol)Biophys J199568192119367612834
- TiroshOBarenholzYKatzhendlerJPrievAHydration of polyethylene glycol-grafted liposomesBiophys J199874137113799512033
- NicolazziCMignetNde laFNAnionic polyethyleneglycol lipids added to cationic lipoplexes increase their plasmatic circulation timeJ Control Release20038842944312644368
- MignetNCadetMBessodesMSchermanDIncorporation of poly(ethylene glycol) lipid into lipoplexes: On-line incorporation assessment and pharmacokinetics advantagesLiposome technologyThird ed2007273292
- HeyesJPalmerLChanKGiesbrechtCJeffsLMaclachlanILipid encapsulation enables the effective systemic delivery of polyplex plasmid DNAMol Ther20071571372017299407
- SkjorringeTGjettingTJensenTGA modified protocol for efficient DNA encapsulation into pegylated immunoliposomes (PILs)J Control Release200913914014519545595
- SchnitzerJEOhPPinneyEAllardJFilipin-sensitive caveolae-mediated transport in endothelium: reduced transcytosis, scavenger endocytosis, and capillary permeability of select macromoleculesJ Cell Biol1994127121712327525606
- RejmanJOberleVZuhornISHoekstraDSize-dependent internalization of particles via the pathways of clathrin- and caveolae-mediated endocytosisBiochem J200437715916914505488
- TheodoropoulosPAGravanisATsaparaACytochalasin B may shorten actin filaments by a mechanism independent of barbed end cappingBiochem Pharmacol199447187518818204105
- PoulsenTTPedersenNJuelHPoulsenHSA chimeric fusion of the hASH1 and EZH2 promoters mediates high and specific reporter and suicide gene expression and cytotoxicity in small cell lung cancer cellsCancer Gene Ther20081556357518421308
- CharroisGJAllenTMDrug release rate influences the pharmacokinetics, biodistribution, therapeutic activity, and toxicity of pegylated liposomal doxorubicin formulations in murine breast cancerBiochim Biophys Acta2004166316717715157619
- BallyMMMayerLDHopeMJNayaRPharmacodynamics of liposomal drug carriers: methodological considerationsGregoriadisGLiposome technologySecond edLondon, UKCRC Press19932741
- DengWGWuGUedaKXuKRothJAJiLEnhancement of antitumor activity of cisplatin in human lung cancer cells by tumor suppressor FUS1Cancer Gene Ther200815293917828283
- RameshRItoISaitoYLocal and systemic inhibition of lung tumor growth after nanoparticle-mediated mda-7/IL-24 gene deliveryDNA Cell Biol20042385085715684712
- LleresDWeibelJMHeisslerDZuberGDuportailGMelyYDependence of the cellular internalization and transfection efficiency on the structure and physicochemical properties of cationic detergent/DNA/liposomesJ Gene Med2004641542815079816
- WustnerDFluorescent sterols as tools in membrane biophysics and cell biologyChem Phys Lipids200714612517241621
- KheirolomoomAFerraraKWCholesterol transport from liposomal delivery vehiclesBiomaterials2007284311432017610949
- ElouahabiARuysschaertJMFormation and intracellular trafficking of lipoplexes and polyplexesMol Ther20051133634715727930
- LuoDSaltzmanWMSynthetic DNA delivery systemsNat Biotechnol200018333710625387
- MonckMAMoriALeeDStabilized plasmid-lipid particles: pharmacokinetics and plasmid delivery to distal tumors following intravenous injectionJ Drug Target2000743945210758914
- FelgnerPLGadekTRHolmMLipofection: a highly efficient, lipid-mediated DNA-transfection procedureProc Natl Acad Sci U S A198784741374172823261
- SmythTNCationic liposomes as in vivo delivery vehiclesCurr Med Chem2003101279128712678799
- SongLYAhkongQFRongQCharacterization of the inhibitory effect of PEG-lipid conjugates on the intracellular delivery of plasmid and antisense DNA mediated by cationic lipid liposomesBiochim Biophys Acta2002155811311750259
- HarviePWongFMBallyMBUse of poly(ethylene glycol)-lipid conjugates to regulate the surface attributes and transfection activity of lipid-DNA particlesJ Pharm Sci20008965266310756331
- MassonCGarinotMMignetNpH-sensitive PEG lipids containing orthoester linkers: new potential tools for nonviral gene deliveryJ Cont Rel200499423434
- ZhangYBradshaw-PierceELDelilleAGustafsonDLAnchordoquyTJIn vivo comparative study of lipid/DNA complexes with different in vitro serum stability: Effects on biodistribution and tumor accumulationJ Pharm Sci20089723725017721944
- Rovira-BruMThompsonDHSzleiferISize and structure of spontaneously forming liposomes in lipid/PEG-lipid mixturesBiophys J2002832419243912414678
- MontesanoGBartucciRBelsitoSMarshDSportelliLLipid membrane expansion and micelle formation by polymer-grafted lipids: Scaling with polymer length studied by spin-label electron spin resonanceBiophys J2001801372138311222298
- PiresPSimoesSNirSGasparRDuzgunesNPedroso de LimaMCInteraction of cationic liposomes and their DNA complexes with monocytic leukemia cellsBiochim Biophys Acta19991418718410209212
- ChenevierPVeyretBRouxDHenry-ToulmeNInteraction of cationic colloids at the surface of J774 cells: a kinetic analysisBiophys J2000791298130910968993
- IvanovAIPharmacological inhibition of endocytic pathways: is it specific enough to be usefulMethods Mol Biol2008440153318369934
- RejmanJBragonziAConeseMRole of clathrin- and caveolae-mediated endocytosis in gene transfer mediated by lipo- and polyplexesMol Ther20051246847415963763
- HoekstraDRejmanJWasunguLShiFZuhornIGene delivery by cationic lipids: in and out of an endosomeBiochem Soc Trans200735687117233603
- WadiaJSStanRVDowdySFTransducible TAT-HA fusogenic peptide enhances escape of TAT-fusion proteins after lipid raft macropinocytosisNat Med20041031031514770178
- CornelisSVandenbrandenMRuysschaertJMElouahabiARole of intracellular cationic liposome-DNA complex dissociation in transfection mediated by cationic lipidsDNA Cell Biol200221919711953008
- BarefordLMSwaanPWEndocytic mechanisms for targeted drug deliveryAdv Drug Deliv Rev20075974875817659804
- Joint Working Group on RefinementRemoval of blood from laboratory mammals and birds. First report of the BVA/FRAME/RSPCA/UFAW Joint Working Group on RefinementLab Anim1993271228437430
- ZhangYAnchordoquyTJThe role of lipid charge density in the serum stability of cationic lipid/DNA complexesBio Biophys Acta-Biomem20041663143157
- FanecaHSimoesSde LimaMCPAssociation of albumin or protamine to lipoplexes: enhancement of transfection and resistance to serumJ Gene Med2004668169215170739
- TandiaBMLonezCVandenbrandenMRuysschaertJMElouahabiALipid mixing between lipoplexes and plasma lipoproteins is a major barrier for intravenous transfection mediated by cationic lipidsJ Biol Chem2005280122551226115661732
- McleanJWFoxEABalukPOrgan-specific endothelial cell uptake of cationic liposome-DNA complexes in miceAm J Phys-Heart Circu Phys199742387404
- GabizonACataneRUzielyBProlonged circulation time and enhanced accumulation in malignant exudates of doxorubicin encapsulated in polyethylene-glycol coated liposomesCancer Res1994549879928313389