Abstract
Platinum nanoparticle (Pt-np) species are superoxide dismutase/catalase mimetics and also have an activity similar to that of mitochondrial electron transport complex I. To examine if this complex I-like activity functions in vivo, we studied the effects of Pt-nps on the lifespan of a mitochondrial complex I-deficient Caenorhabditis elegans mutant, nuo-1 (LB25) compared with wild-type N2. We synthesized a fusion protein of a cell-penetrating peptide, human immunodeficiency virus-1 TAT (48–60), C-terminally linked to a peptide with a high affinity to platinum (GRKKRRQRRRPPQ-DRTSTWR). Pt-nps were functionalized by conjugation with this fusion protein at a 1:1 ratio of TAT-PtBP to Pt atoms. Adult worms were treated with conjugated Pt-nps for 10 days. The mean lifespan of untreated N2 and LB25 was 19.6 ± 0.4 and 11.8 ± 0.3 days, respectively. Using 5 μM of conjugated Pt-nps, the lifespan of N2 and LB25 was maximally extended. This maximal lifespan extension of LB25 was 31.9 ± 2.6%, which was significantly greater than that of N2 (21.1 ± 1.7%, P < 0.05 by Student’s t-test). Internalization of Pt into the whole body and mitochondria was similar between these two strains. Excessive accumulation of reactive oxygen species was not observed in the cytosol or mitochondria of untreated LB25. Treatment for five days with 5 μM conjugated Pt-nps decreased cytosolic and mitochondrial reactive oxygen species in N2 and LB25 to a similar extent. The ratio of [NAD+]/[NADH] was very low in the whole body and mitochondria of control LB25. After five days of treatment with 5 μM conjugated Pt-nps, the ratio of [NAD+]/[NADH] was increased in N2 and LB25. However, the degree of the increase was much higher in LB25 than in N2. Pt-nps function as NADH oxidase and recover the [NAD+]/[NADH] ratio in LB25, leading to effective extension of the lifespan of LB25.
Introduction
Platinum nanoparticles (Pt-nps) are an efficient catalyst for electro-oxidation of hydrogen peroxide (H2O2).Citation1 Owing to this catalase activity, Pt-nps are utilized as a fine element of biosensors.Citation2,Citation3 However, Pt-nps have not been developed sufficient for biologic or medical use compared with gold nanoparticles (Au-nps).Citation4–Citation6 Recently, some papers have shown biologic and medical application of Pt-nps, including usage as a platform for drug delivery.Citation7–Citation9 We regard Pt-nps as a bioactive material, ie, a kind of antioxidant, and have undertaken research on these entities. In vitro Pt-nps scavenge reactive oxygen species (ROS) including superoxide anion (O2−) as well as H2O2, and free radicals.Citation1,Citation10 Pt-nps seem to have catalytic antioxidant activity. Because the O2−-scavenging activity of Au-nps is less than that of Pt-nps, and Au-nps cannot scavenge H2O2, Pt-nps are a more potent antioxidant than Au-nps.Citation1 In vivo we tested the effects of Pt-nps on the lifespan of Caenorhabditis elegans.Citation11 Pt-nps extended the lifespan of wild-type N2 and the mitochondrial electron transport complex II mutant, mev-1(kn1), whereas a well known superoxide dismutase/catalase mimetic, EUK-8, did not do this. Furthermore, Pt-nps exhibited more effective protection of nematodes against acute oxidative stress induced by paraquat than EUK-8. These data suggest that Pt-nps scavenge excessive oxidative stress to elongate the lifespan and survival of nematodes.
Mitochondrial electron transport complex I consists of many subunits and is located in the mitochondrial inner membrane.Citation12 This complex is NADH:ubiquinone oxidoreductase that transfers electrons from NADH in the matrix to ubiquinone in the inner membrane, with proton transport from the matrix to the mitochondrial intramembrane space. In some diseases the complex I is deficient or its enzymatic activity is suppressed, leading to generation of an excessive amount of O2−. Accumulated ROS seem to contribute partially to the pathogenesis of those diseases.Citation13–Citation16 Recently, we have shown in vitro that Pt-nps have an activity similar to the complex I that drives oxidation of NADH to NAD+, as well as reduction of ubiquinone to ubiqunol.Citation17 Although it is still not known if Pt-nps transport protons, we expect that Pt-np species will be an enzymatic mimetic of mitochondrial complex I. Pt-nps are able to mediate oxidation of NADH and reduction of ubiquinone separately. In other words, Pt-nps do not require electrons from NADH to reduce ubiquinone. We suspect that Pt-nps may accept and release electrons by interaction with water molecules.
A mitochondrial complex I-deficient C. elegans mutant, LB25, is a transgenic strain expressing the nuo-1 (NADH:ubiquinone oxidoreductase) gene carrying an A352V mutation.Citation18 The nuo-1 gene is the C. elegans homolog of the human NDUFV1 (NADH dehydrogenase ubiquinone flavoprotein 1) gene encoding the active site subunit of complex I. The A352V mutation corresponds to the residue A341 of the human NDUFV1 gene which causes severe neurologic disorders and muscle atrophy in early childhood.Citation13 In this study, we describe the significance of Pt-nps as a mitochondrial complex I mimetic, as well as an antioxidant, in partial recovery of the short lifespan of LB25, indicating that Pt-np species may be a new potent material for diseases with dysfunctional mitochondrial complex I.
Material and methods
C. elegans and growth conditions
Wild-type N2 and a mitochondrial complex I-deficient mutant C. elegans, nuo-1 (LB25), were obtained from the Caenorhabditis Genetic Center (University of Minnesota, St Paul, MN). These strains were maintained at 20°C by a popular procedure established by Brenner.Citation19 Age-synchronous populations were prepared as previously described.Citation11 Briefly, collected eggs were allowed to hatch overnight at 20°C in 1 mL of S-basal buffer (100 mM NaCl, 0.01 mM cholesterol, and 50 mM potassium phosphate, pH 6.0), on nematode growth medium agar plates.Citation20 Hatched worms (L1 larval stage) were transferred to fresh nematode growth medium agar plates with Escherichia coli OP50 as a food source, and cultured at 20°C until the L4 larval stage.
Preparation of Pt-nps
Pt-nps were prepared by an ethanol reduction method of hydrogen hexachloroplatinate (H2PtCl6) using poly(N-vinyl-2-pyrrolidone, PVP) as a protecting reagent.Citation11 PVP, H2PtCl6·6H2O, and ethanol were purchased from Wako Pure Chemical Industries (Osaka, Japan). Water was freshly prepared with a Millipore Milli-Q Academic Water Purification System (Millipore, Billerica, MA). We estimated the concentration of Pt in Pt-nps from the concentration of PtCl62− in a starting reaction mixture, assuming 100% reduction of Pt4+ to Pt. Therefore, the Pt-nps concentrations reported herein actually refer to the estimated concentration of Pt atoms present in Pt-nps.
The human immunodeficiency virus (HIV)-1 protein, TAT, is a transcriptional activator of HIV. TAT consists of 86 amino acids, but its translocation activity is associated with the peptide sequence (48–60; GRKKRRQRRRPPQ).Citation21 A short peptide (DRTSTWR) is known to be one with high affinity to platinum and is referred to here as a platinum-binding peptide (PtBP).Citation22 A fusion peptide was synthesized by C-terminally linking of HIV-1 TAT (48–60) to PtBP (GRKKRRQRRRPPQ-DRTSTWR). Pt-nps protected with PVP (PVP-Pt) were conjugated with this fusion protein (TAT-PtBP) at a 1:1 ratio of TAT-PtBP to Pt atom to increase the internalization of Pt-nps into worms.Citation23 The consequent conjugate was designated as TAT-PtBP-Pt. Therefore, unconjugated Pt-nps indicates PVP-Pt.
Lifespan assay
For the lifespan assay, synchronous L4 larvae were transferred to S-medium (S-basal medium supplemented with 3 mM CaCl2, 3 mM MgSO4, 50 μM ethylenediamine tetraacetic acid [EDTA], 25 μM FeSO4, 10 μM MnCl2, 10 μM ZnSO4, 1 μM CuSO4, and 10 mM KH2PO4, at pH 6.0) with E. coli OP50.Citation24 We chose a liquid medium because it was difficult to make an agar plate in which TAT-PtBP-Pt were evenly distributed and, furthermore, because worms were allowed to make contact freely with TAT-PtBP-Pt. This transfer day was recorded as Day 0. Nematodes were treated with TAT-PtBP-Pt for 10 days from Day 0. The worms were transferred to fresh culture medium every second day, and the surviving worms were counted at the same time. Worms that died unnaturally were excluded. The mortality data were subjected to Kaplan–Meier survival analysis to prepare survival curves. Statistical comparisons of mean lifespan values between untreated (control) and treated worms were analyzed by Peto’s log-rank test.
Isolation of mitochondria
Isolation of mitochondria was performed as previously described.Citation18 Worms were collected from liquid culture and rinsed three times with M9 buffer (3 g/L KH2PO4, 6 g/L Na2HPO4, and 5 g/L NaCl supplemented with 1 mM MgSO4 and 0.02% gelatin after autoclaving). Rinsed worms were suspended in 0.2 M mannitol, 70 mM sucrose, 0.1 M EDTA, and 1 mM phenylmethylsulfonyl fluoride at pH 7.4. Worms were homogenized 10 times by a BRANSON Sonifier 150 with a 3 mm diameter probe (Branson Ultrasonic Co., Danbury, CT). Homogenized samples were centrifuged three times at 1000 × g for 10 minutes to eliminate unbroken cells and cuticles. The supernatant was then centrifuged at 19,200 × g for 10 minutes to precipitate mitochondria. The pellet was suspended in 0.2 M mannitol, 70 mM sucrose, and 0.1 M EDTA, pH 7.4 (MSE buffer) and centrifuged again. The resultant sediment containing mitochondria was resuspended in 0.5 mL MSE buffer. Protein concentration was estimated by the Bradford method with a commercially available kit (Bio-Rad, Hercules, CA).
Measurement of platinum content
On Day 0, treatments of N2 and LB25 with 5 μM TAT-PtBP-Pt were started and continued for five days. Nematodes treated with unconjugated PVP-Pt served as controls. Treated nematodes were rinsed three times with M9 buffer and transferred to crucibles with 1 mL of 1% nitric acid, 60% perchloric acid, and 35% chloric acid.Citation23 The samples ware heated and volatilized at 200°C using an electric hot plate, followed by heating with 1 mL of 1% nitric acid three times. Finally, the samples were resuspended in 5 mL of 1% nitric acid. Determination of amount of Pt in the resuspended samples was performed. Measurements of Pt internalized in mitochondria were performed using similar procedures. The statistical significance of differences between the control and treated groups was determined by Student’s t-test.
Measurement of reactive oxygen species
N2 and LB25 nematodes were treated with 5 μM TAT-PtBP-Pt for five days from Day 0. On day 5, the treated nematodes were washed three times with M9 buffer. The worms were then incubated for 30 minutes at 20°C in 2 mL of Hank’s solution (0.44 mM KH2PO4, 5.37 mM KCl, 0.34 mM Na2HPO4, 136.89 mM NaCl, and 5.55 mM D-glucose) containing 10 μM 5-(and 6)-chloromethyl-2’,7’-dichlorodihydrofluorescein diacetate acetyl ester (CM-H2DCFDA) (Sigma-Aldrich Co., St. Louis, MO) and/or MitoSOX™ Red (MitoSox, obtained from Invitrogen, Carlsbad, CA).Citation11 CM-H2DCF released by esterase and Mito-Sox were fluorescent dyes to assay cytosolic ROS mainly consisting of H2O2 and mitochondrial O2−, respectively. Worms were fixed with 4% formaldehyde and mounted on 2% agarose pads. To determine the fluorescence of CM-H2DCF and MitoSOX, fixed nematode samples were subjected to Leica TCS SP2 laser scanning confocal microscopy (CM-H2DCF, excitation at 488 nm and emission at 510 nm; MitoSOX, excitation at 510 nm and emission at 580 nm). The relative fluorescence of the whole body was determined densitometrically using the LCS Lite software. The statistical significance of differences between the control and treated groups was determined by Student’s t-test.
NAD+ and NADH assays
Nematodes were treated with 5 μM TAT-PtBP-Pt for five days from Day 0. On Day 5, treated animals were rinsed three times with M9 buffer. Rinsed worms were suspended in 100 mM NaCl, 1% NP-40, 1 mM DTT, 10% glycerol, and 100 mM Tris, at pH 8. Worms were homogenized 10 times by BRANSON Sonifier 150 with a 3 mm diameter probe. Protein concentration was estimated by the Bradford method. We used an NAD+/NADH assay kit (Gentaur, Kampenhout, Belgium). Homogenized samples were mixed with 100 μL of NAD+ extraction buffer for NAD+ estimation or NADH extraction buffer for NADH estimation and heated at 60°C for five minutes. Twenty microliters of assay buffer and 100 μL of NADH (or NAD+) extraction buffer were added. Samples were then vortexed and spun at 10,000 × rpm for five minutes. The resultant supernatant was used for NAD+ and NADH assays. Similar procedures were performed to determine NAD+ and NADH in isolated mitochondria. The statistical significance of differences between the control and treated groups was determined by Student’s t-test.
Results and discussion
TAT-PtBP-Pt extends lifespan of wild-type N2 and nuo-1 mutant
The survival of control wild-type N2 and mutant LB25 delineates regular reverse-sigmoid curves (). The mutants died earlier than the wild type. Treatment with 5 μM TAT-PtBP-Pt for 10 days from Day 0 shifts the survival curves of N2 and LB25 to the right, indicating extension of their lifespan. The mean lifespan was obtained by calculating the data of the survival curves. The mean lifespans of control N2 and LB25 were 19.6 ± 0.4 and 11.8 ± 0.3 days, respectively, indicating that the mean lifespan of LB25 is approximately 40% less than that of N2 (see ). This result is in good accordance with a previous report.Citation18 The concentration of TAT-PtBP-Pt was varied from 1–25 μM. While the lifespan of N2 was significantly extended by TAT-PtBP-Pt only at 5 μM (***P < 0.001), the mutant lifespan was extended by more than 5 μM and was maximal at 5 μM (**P < 0.01 (10 and 25 μM), ***P < 0.001 (5 μM), as shown in and ). The maximal lifespan of LB25 was 15.6 ± 0.4 days by 31.9 ± 2.6% increase of their control lifespan (see ). Treatment of N2 with 5 μM TAT-PtBP-Pt extended the mean lifespan to 23.3 ± 0.4 days only by 21.1 ± 1.7%. TAT-PtBP at 25 μM alone did not affect either N2 or LB25 lifespan ( and ). These data suggest that Pt in TAT-PtBP-Pt is the active source of the nematode lifespan extension, and the lifespan extension by Pt-nps is significantly more effective in LB25 than in N2 (P < 0.05 by Student’s t-test).
Figure 1 Effects of TAT-PtBP-Pt on the lifespan of N2 and LB25. Adult nematodes were treated with 5 μM TAT-PtBP-Pt for days 0–10. Shown are survival curves of untreated control (•) and treated (○) worms A). The number of worms was 90–95 in each experiment, and three independent experiments were repeated. The mean lifespan B) was calculated from survival curves. The concentration of Pt in TAT-PtBP-Pt was varied up to 25 μM. Effect of the fusion protein (TAT-PtBP) at 25 μM alone on the lifespan of nematodes was also studied. Error bars represent the standard error of the mean.
Notes: **P < 0.01; ***P < 0.001, as compared with control worms by the log-rank test. The lifespan data are summarized in the .
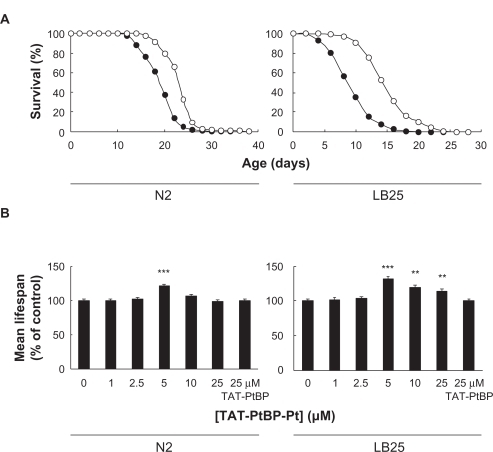
Table 1 Effects of TAT-PtBP-Pt on the lifespan of C. elegans
Recently, we have performed a similar experiment to monitor the lifespan extension of wild-type nematode N2 by conjugation of Pt-nps with TAT-PTBP.Citation23 The lifespan of control N2 was 18.0 ± 0.3 days. Treatment with TAT-PtBP-Pt significantly elongated the lifespan in the concentration range 1–50 μM (P < 0.001). The maximal extension of lifespan was observed with 5 μM TAT-PtBP-Pt. The maximal mean lifespan was 22.4 ± 0.4 days, resulting in an increase of 24%. The experimental difference between our previous and current studies is the treatment period. Nematodes were treated until death in the previous study, but only for 10 days in the current study.
The optimal increase of life span by TAT-PtBP-Pt was observed at 5 μM. Greater concentrations of TAT-PtBP-Pt showed less positive effects. Currently, we do not have a clear explanation for this phenomenon. A possible speculation is that the lifespan extension by Pt-nps may gradually decrease with the concentration increase due to toxicity. Pt-nps may extend lifespan by activating cellular signaling via a small change of ROS, but not by a big change.Citation25 Further studies are required.
Similar internalization of Pt-nps by N2 and nuo-1
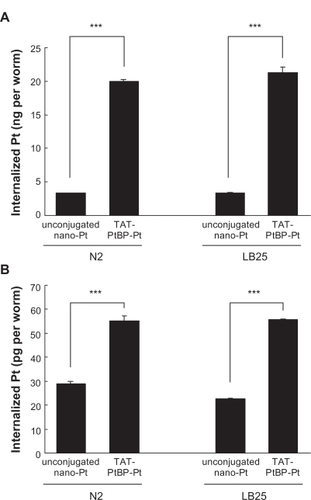
The amount of Pt internalized in N2 and LB25 whole bodies by treatment with 5 μM unconjugated PVP-Pt for five days from Day 0 was 3.30 ± 0.05 and 3.33 ± 0.12 ng/worm, respectively (). Conjugation of Pt-nps with TAT-PtBP enhances Pt internalization approximately six times in both nematodes (***P < 0.001; amount of internalized Pt in N2 and LB25 whole bodies, 19.9 ± 0.4 and 21.3 ± 0.9 ng/worm, respectively). The mitochondrial Pt accumulation from PVP-Pt was approximately 1% of the whole body Pt accumulation (29.1 ± 0.8 and 22.7 ± 0.3 pg/worm in N2 and LB25 mitochondria, respectively, ). Although TAT was used as a cell-penetrating peptide (CPP), it was significantly effective even in mitochondrial translocation (***P < 0.001; N2 and LB25, 55.3 ± 2.0 and 55.7 ± 0.3 ng/worm, respectively). There was not a significant difference in whole body or mitochondrial accumulation of Pt from TAT-PtBP-Pt between these two strains. Therefore, the more effective lifespan extension of LB25 than N2 by treatment with 5 μM TAT-PtBP-Pt is not due to the higher permeability of Pt in LB25 than N2.
Figure 2 Amount of Pt internalized in whole bodies and mitochondria of N2 and LB25. Animals were treated with 5 μM TAT-PtBP-Pt for five days from Day 0. Pt accumulation in worm whole bodies A) and mitochondria B) was determined by an inductively-coupled plasma mass spectrometer ICP-MS. For control, unconjugated Pt-nps (PVP-Pt) was used. The number of worms used was 100 and 500–508 in each measurement for whole bodies and mitochondria, respectively, and three independent experiments were repeated. Error bars represent the standard error of the mean.
Note: Statistical significance was determined by Student’s t-test (***P < 0.001).
CPPs are referred to as protein transduction domains and utilized to facilitate cellular uptake of hydrophilic bioactive large molecules.Citation26 TAT is one of the popular CPPs and successful in delivery of proteins, peptides, and nucleotides across the cell membrane.Citation27 TAT has been used for protein delivery, even in C. elegans.Citation28 CPPs are usually linked to cargoes by covalent binding.Citation29 For example, in the case of Au-nps, tiopronin, a kind of spacer, was first covalently bound to gold atoms via a sulfhydryl group during reduction of AuCl4− by NaBH4.Citation30 TAT was then covalently bound to tiopronin to functionalize the Au-nps. We used PtBP to link TAT to Pt-nps.Citation23 This is a unique approach to transcellular delivery of metal-nps. Although precise mechanisms for internalization of Pt-nps into nematodes are unknown, we assume that intestinal permeability of Pt-nps is enhanced by conjugation with TAT-PtBP. Previous studies have shown that TAT conjugation leads Au-nps to nuclear translocation in cells.Citation30,Citation31 Furthermore, CPPs interrupt mitochondrial targetting.Citation32 Our results indicate that TAT-PtBP conjugation facilitates mitochondrial translocation of Pt-nps to a small but significant extent. We need more sophisticated research on intracellular localization of TAT conjugated Pt-nps.
Reactive oxygen species in cytosol and mitochondria of N2 and nuo-1
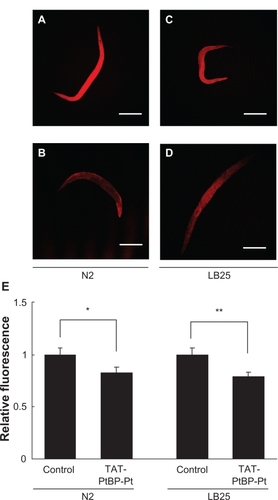
The effects of TAT-PtBP-Pt treatment on ROS were investigated in N2 and LB25. Typical fluorescence from cytosolic CM-H2DCF in the whole body is shown in (A, control N2; B, treated N2; C, control LB25; D, treated LB25). The fluorescent intensity obtained using LCS Lite software was similar between control N2 and LB25 (46.4 ± 2.8 and 42.5 ± 2.8, respectively). The relative fluorescence is shown in . Treatment with 5 μM TAT-PtBP-Pt for five days from Day 0 significantly decreased the fluorescence, and the extent of this reduction was also similar between these two strains (−39.2 ± 2.1 and −33.7 ± 1.8%, respectively, *P < 0.05). Mitochondrial O2− was also measured with MitoSox in ; A, control N2; B, treated N2; C, control LB25; D, treated LB25). The fluorescent intensity from mitochondrial MitoSox was 48.0 ± 3.0 and 45.7 ± 2.8 in control N2 and LB25, respectively ( depicted with relative fluorescence). A similar treatment with 5 μM TAT-PtBP-Pt significantly decreased the MitoSox fluorescence (39.6 ± 2.7; *P < 0.05 and 35.9 ± 2.0; **P < 0.01 as intensity in N2 and LB25, respectively). These results show that ROS accumulation in control LB25 is not excessive, and TAT-PtBP-Pt treatment decreases ROS in N2 and LB25 to a similar extent. A main cause of the short life of LB25 may not be ROS accumulation.
Figure 3 Effects of TAT-PtBP-Pt on cytosolic reactive oxygen species in N2 and LB25. Animals were treated with 5 μM TAT-PtBP-Pt for five days from Day 0. Worms were then treated with 10 μM CM-H2DCFDA for 30 minutes at 20°C. Fluorescence was measured by confocal florescence microscopy (Ex 488 nm and Em 510 nm). Shown is cytosolic reactive oxygen species in N2 and LB-25 whole bodies: A) control N2; B) treated N2; C) control LB25; D) treated LB25. The panel E) shows the average of relative intensity for three independent experiments with 10 worms per each experiment. Error bars represent the standard error of the mean.
Notes: *P < 0.05; **P < 0.01, as compared with control worms by Student’s t-test.
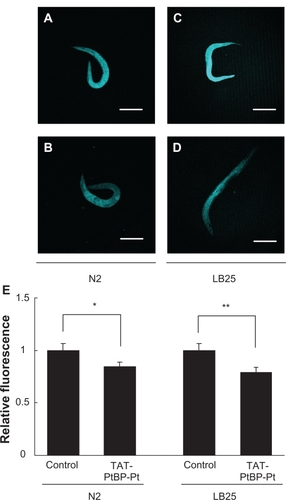
Figure 4 Effects of TAT-PtBP-Pt on O2− in N2 and LB mitochondria. Animals were cultured with TAT-PtBP-Pt for five days from Day 0. Worms were then treated with 10 μM MitoSOX for 30 minutes at 20°C. Fluorescence was measured by confocal florescence microscopy (Ex 510 nm and Em 580 nm). Shown is O2− in N2 and LB25 mitochondria: A) control N2; B) treated N2; C) control LB25; D) treated LB25. The panel E) shows the average of relative intensity for three independent experiments with 10 worms per each experiment. Error bars represent the SEM.
Notes: *P < 0.05; **P < 0.01, as compared with control worms by Student’s t-test.
We measured cytosolic and mitochondrial ROS in LB25 for the first time. Excessive accumulation of ROS was not observed in the cytosol or mitochondria of control LB25 in comparison with control N2. We think that the quantification of ROS level is insufficient due to the ROS estimation by fluorescent dyes. However, these results are still interesting. It has been reported that 1-methyl-4-phenylpyridinium and rotenone, inhibitors of complex I, lead to NADH-dependent generation of O2− using prepared bovine mitochondrial particles.Citation33 However, this increase of O2− generation by the inhibitors was measured for a short period. Thus, we have to study the perturbation of ROS level by suppression of complex I for a long time in vivo. In fact, oxygen consumption was reduced by 50% in LB25, compared with N2.Citation18 Decreased oxygen consumption is supposed to help decrease the generation of ROS. Currently, we do not know how LB25 worms maintain ROS at a relatively normal level during life. Because the oxidative state must be high in LB25, oxidized substances may be accumulated to reduce free ROS. Oxidative marker assays will be required.
Increase of [NAD+]/[NADH] ratio in nuo-1 by TAT-PtBP-Pt
We measured NAD+ and NADH levels in the whole body homogenate and mitochondrial fraction from N2 and LB25. The [NAD+]/[NADH] ratio was considerably smaller in the whole body (0.057 ± 0.004) and mitochondria (0.026 ± 0.005) of control LB25 than in those of control N2 (whole body, 0.167 ± 0.005; mitochondria, 0.146 ± 0.006), indicating excessive accumulation of NADH in LB25 (). Treatment of N2 with 5 μM TAT-PtBP-Pt significantly increased the [NAD+]/[NADH] ratio by approximately twice (***P < 0.001). A similar treatment of LB25 with 5 μM TAT-PtBP-Pt greatly increased the [NAD+]/[NADH] ratio by 4.6 and 8.8 times in whole body and mitochondria, respectively (***P < 0.001). These results indicate that Pt-nps function as NADH oxidase in C. elegans, and accumulated NADH in LB25 is efficiently oxidized to NAD+. This recovery of the [NAD+]/[NADH] ratio may take part in the effective lifespan extension of LB25.
Figure 5 Effects of TAT-PtBP-Pt on the [NAD+]/[NADH] ratio in N2 and LB25. The concentrations of NAD+ and NADH were measured in whole bodies A) and mitochondria B) using an assay kit. The number of worms was 100 and 500 in each measurement for whole bodies and mitochondria, respectively, and three independent experiments were repeated. Error bars represent the standard error of the mean.
Note: ***P < 0.001 as compared with control worms by Student’s t-test.
![Figure 5 Effects of TAT-PtBP-Pt on the [NAD+]/[NADH] ratio in N2 and LB25. The concentrations of NAD+ and NADH were measured in whole bodies A) and mitochondria B) using an assay kit. The number of worms was 100 and 500 in each measurement for whole bodies and mitochondria, respectively, and three independent experiments were repeated. Error bars represent the standard error of the mean.Note: ***P < 0.001 as compared with control worms by Student’s t-test.](/cms/asset/f8d9c154-8d7d-44e7-a8a8-1643c10aab97/dijn_a_12184693_f0005_b.jpg)
In nuo-1 mutants, including LB25, excessive cellular NADH caused reduction of pyruvate to lactate.Citation18 Depletion of pyruvate slows down tricarboxylic acid cycle and oxidative phosphorylation. We believe that Pt-nps oxidize NADH to NAD+ to normalize the NAD+/NADH ratio, which recovers these metabolic pathways and increases adenosine triphosphate synthesis, even though the activity of complex IV is decreased by approximately 50%.Citation18 Increase of adenosine triphosphate helps recover the short life of LB25. One may think that accelerated oxidative phosphorylation may produce O2− as a byproduct because LB25 has the low activity of complex I. Pt-nps are able to scavenge ROS including O2. A similar decrease of ROS by TAT-PtBP-Pt treatment was observed in the cytosol and mitochondria of N2 and LB25 ( and ). However, if Pt-nps efficiently scavenge more ROS in treated LB25, a similar decrease of ROS may be apparent. The lifespan recovery of LB25 by Pt-nps may be supported by the maintenance of ROS using Pt-nps. We believe that the NADH oxidase activity of Pt-nps in scavenging ROS plays a pivotal role in effective recovery of the short lifespan of LB25.
Furthermore, even in wild-type N2, the [NAD+]/[NADH] ratio was increased by treatment with Pt-nps. The NAD+-dependent SIR-2.1 pathway may be involved in the lifespan extension caused by treatment with Pt-nps.Citation34 If it is, this signal pathway seems to be more effective in LB25 to elongate the lifespan because of a marked increase of NAD+ by Pt-nps treatment.
Conclusion
Pt-nps functionalized with a TAT fusion protein are internalized in the mitochondria as well as in the cytosol and exert NADH oxidase activity. Therefore, Pt-np species may be a potent medicinal material to improve symptoms caused by mitochondrial complex I defect. Active targeting of the Pt-nps to mitochondrial matrix and/or inner membrane using signal peptides will be helpful to develop this potency further.
Acknowledgements
This work was supported in part by a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science, and Technology, Japan.
Disclosure
The authors report no conflicts of interest in this work.
References
- KajitaMHikosakaKIitsukaMKanayamaAToshimaNMiyamotoYPlatinum nanoparticle is a useful scavenger of superoxide anion and hydrogen peroxideFree Rad Res200741615626
- CaoZZouYXiangCSunLXXuFAmperometric glucose biosensor based on ultrafine platinum nanoparticlesAnal Lett20074021162127
- PolskyRGillRKaganovskyLWillnerINucleic acid-functionalized Pt nanoparticles: Catalytic labels for the amplified electrochemical detection of biomoleculesAnal Chem2006782268227116579607
- BhattacharyaRMukherjeePBiological properties of “naked” metal nanoparticlesAdv Drug Deliv Rev2008601289130618501989
- BoisselierEAstrucDGold nanoparticles in nanomedicine: Preparation, imaging, diagnostics, therapies and toxicityChem Soc Rev2009381759178219587967
- GhoshPHanGDeMKimCKRotelloVMGold nanoparticles in delivery applicationsAdv Drug Deliv Rev2008601307131518555555
- BaroneMSciortinoMTZaccariaDMazzagliaASortinoSNitric oxide photocaging platinum nanoparticles with anticancer potentialJ Mater Chem20081855315536
- ZangLLaugLMünchgesangWReducing stress on cells with apoferritin-encapsulated platinum nanoparticlesNano Lett201019219223
- PelkaJGehrkeHEsselenMCellular upake of platinum nanoparticles in human colon carcinoma cells and their impact on cellular redox systems and DNA integrityChem Res Toxicol20092264965919290672
- WatanabeAKajitaMKimJIn vitro free radical scavenging activity of platinum nanoparticlesNanotechnology20092045510519834242
- KimJTakahashiMShimizuTEffects of a potent antioxidant, platinum nanoparticle, on the lifespan ofCaenorhabditis elegans Mech Ageing Dev2008129322331
- HirstJTowards the molecular mechanism of respiratory complex IBiochem J201042532733920025615
- SchuelkeMSmeitinkJMarimanEMutant NDUFV1 subunit of mitochondrial complex I causes leukodyatrophy and myoclonic epilepsyNat Genet19992126026110080174
- BénitPChretienDKadhomNLarge-scale deletion and point mutations of nuclear NDUFV1 and NOUFS1 genes in mitochondrial complex I deficiencyAm J Hum Genet2001681344135211349233
- WinklhoferKHaassCMitochondrial dysfunction in Parkinson’s diseaseBiochim Biophys Acta20101802294419733240
- KorenagaMOkudaMOtaniKWangTLiYWeinmanSAMitochondrial dysfunction in hepatitis CJ Clin Gasteroenterol200539s162s166
- HikosakaKKimJKajitaMKanayamaAMiyamotoYPlatinum nanoparticles have an activity similar to mitochondrial NADH: Ubiquinone oxidoreductaseColloids Surf Biointerfaces200866195200
- GradLILemireBDMitochondrial complex I mutations in Caenorhabditis elegans produce cytochrome c oxidase deficiency, oxidative stress and vitamin-responsive lactic acidosisHum Mol Genet20041330331414662656
- BrennerSThe genetics of Caenorhabditis elegansGenetics19747771944366476
- SulstonJEBrennerSThe DNA of Caenorhabditis elegansGenetics197477951044858229
- JärverPLangelÜCell-penetrating peptides – A brief introductionBiochim Biophys Acta2006175826026316574060
- SarikayaMTamerlerCJenAKYSchultenKBaneyxFMolecular biomimetics: Nanotechnology through biologyNat Mater2003257758512951599
- KimJShirasawaTMiyamotoYThe effect of TAT conjugated platinum nanoparticles on lifespan in a nematode Caenorhabditis elegans modelBiomaterials2010315849585420434216
- JohnsonTEWoodWBGenetic analysis of the life-span of Caenorhabditis elegansProc Natl Acad Sci U S A198279660366076959141
- HeidlerTHarwingKDanielHWenzelUCaenorhabditis elegans lifespan extension caused by treatment with an orally activity ROS-generator in dependent on DAF-16 and SIR-2.1Biogerontology20101118319519597959
- El-AndaloussiSHolmTLangelÜCell-penetrating peptides: Mechanisms and ApplicationsCurr Pharm Des2005113597361116305497
- VivèsELebleuBThe tat-derived cell-penetrating peptideLangelÜCell-Penetrating Peptides: Processes and ApplicationsBoca Raton, FLCRC Press LLC2002
- DelomFFessartDCarusoMEChevetETat-mediated protein delivery in living Caenorhabditis elegansBiochem Biophys Res Commun200735258759117141180
- WagstaffKMJansDAProtein transduction: Cell penetrating peptides and their therapeutic applicationsCurr Med Chem2006131371138716719783
- de la FuenteJMBerryCCTat peptide as an efficient molecule to translocate gold nanoparticles into the cell nucleusBioconjug Chem2005161176118016173795
- BerryCCde la FuenteJMMullinMChuSWICurtisASGNuclear localization of HIV-1 tat functionalized gold nanoparticlesIEEE Trans Nanobiosci20076262269
- RossMFMurphyMPCell-penetrating peptides are excluded from the mitochondrial matrixBiochem Soc Trans2004321072107415506968
- HasegawaETakeshigeKOishiTMuraiYMinakamiS1-Methyl-4-phenylpyridinium (MPP+) induces NADH-dependent superoxide formation and enhances NADH-dependent lipid peroxidation in bovine heart submitochondrial particlesBiochem Biophys Res Commun1990170104910552167668
- TissenbaumHGurenteLIncreased dosage of a sir-2 gene extends lifespan inCaenorhabditis elegans Nature2001410227230