
Abstract
This study provides a review of the therapeutic potential of graphene dressing scaffolds and mesenchymal stem cells (MSCs) and their synergistic effects with respect to cutaneous wound healing. This study also considers their putative action mechanism based on the antibacterial, immunomodulating, angiogenic, matrix remodeling effects of materials belonging to the graphene family and MSCs during the wound healing process. In addition, this study discusses the cytocompatibility of graphene, its uses as a platform for skin substitutes, the properties it possesses with respect to providing protection against microbial invasion as well as strategies aimed at minimizing the chance of the occurrence of sepsis. MSCs are capable of secreting several factors that exert a therapeutic impact on reparative processes and tissue regeneration. In light of experiments conducted to date, graphene combined with MSCs appears to have the potential to enhance both the wound healing process and infection control at the injury site.
Introduction
Graphene, in combination with mesenchymal stem cells (MSCs), provides a potential clinical application for wound healing purposes. A number of strategies have been advanced to date aimed at enhancing and accelerating the closure of injured tissue in cutaneous wounds, one of which consists of the use of dressing materials containing graphene and derivatives thereof. Moreover, in recent years the attention of a large number of research teams has been devoted to therapy employing MSCs. This study suggests that the synergic effect of a combination of these two approaches may potentially assist in the healing of acute and chronic wounds, which presents a major clinical problem in the fields of both veterinary and human medicine, and with concern to which, due to increasing bacterial resistance, local treatment plays an especially important role. Alternative treatments for hard-to-heal wounds include the application of platelet-rich plasma and cell growth factor preparations, vacuum dressings and other dressings that exhibit antibacterial properties. Deepachitra et alCitation1 demonstrated both in vitro (fibroblasts) and in vivo (rats) that graphene oxide (GO) combined with a collagen-fibrin biofilm can be successfully employed as a dressing material. The treatment of local wounds with MSC applications has gained popularity in recent years as a promising approach for the enhancement of tissue regeneration. It is thought that the therapeutic benefit of MSCs lies principally in the various factors that they secrete such as vascular endothelial growth factor (VEGF), EGF, fibroblast growth factor (FGF), keratinocyte growth factor (KGF), insulin-like growth factor (IGF), platelet-derived growth factor (PDGF), TGF-β, prostaglandin E2 (PGE2), hepatocyte growth factor (HGF), tumor necrosis factor-α (TNF-α), interferon λ (IFNλ) and ILs such as IL-4, IL-6 and IL-10.Citation2–Citation9 All these growth factors and cytokines play an important role in the formation of new blood vessels, cell recruitment, immunomodulation and wound closure. Moreover, MSCs promote direct cell differentiation, proliferation and extracellular matrix (ECM) remodeling.Citation10,Citation11
Cell cultures are employed for both the basic research of many biological processes and for medical applications such as tissue engineering. In the first case, they provide a relatively simple experimental model in contrast to tissues that contain numerous differing cell types. Cell cultures allow for the culturing of specific types of cells and facilitate the study of processes such as cell division, the formation of organelles, protein secretion and differentiation into tissue with specific and determined phenotypes. Intracellular mechanisms, which can be investigated in detail, allow for the closer examination of metabolic processes than would otherwise be possible. With respect to the field of tissue engineering, they enable complicated manipulation leading to the creation of new tissue, which can be maintained and cultivated outside the organism of the donor/recipient. Due to rapid cell growth, in vitro experiments provide information on the process under investigation more quickly than do other methods, thus expediting the obtaining of the final product. Thus, this study suggests that graphene, in combination with MSCs, has significant potential with respect to wound healing and infection control applications.
Cutaneous wound healing
Skin makes up the most extensive organ in the body and has numerous functions including protection against microorganisms. Once this natural barrier is damaged due to injury, burns or systemic dysfunction, the risk of infection increases significantly potentially leading to the occurrence of severe general complications including sepsis.
The cutaneous wound healing process is divided into four distinct phases, ie, hemostasis, inflammation, proliferation and tissue remodeling (). It involves cells such as platelets, inflammatory cells, epithelial cells, keratinocytes, fibroblasts, a multitude of cytokines, growth factors and other bioactive molecules as well as interactions with ECM components mediated by integrin receptors and adhesive molecules. Chronic wounds are considered to be those that do not heal within 12 weeks of injury, which usually leads to prolonged pathological inflammation;Citation12–Citation15
Figure 1 Wound healing stages and the bioactive molecules involved in the healing process.
Abbreviations: FGF, fibroblast growth factor; KGF, keratinocyte growth factor; MMP, matrix metalloproteinase; PDGF, platelet-derived growth factor; TIMP, tissue inhibitor of metalloproteinase; TNF-α, tumor necrosis factor-α; VEGF, vascular endothelial growth factor.
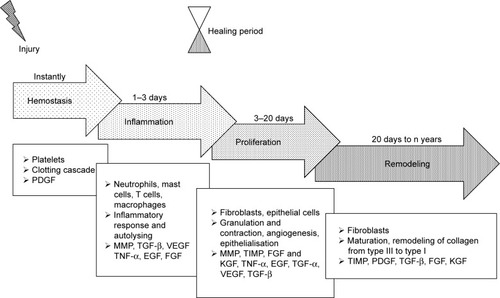
thus, the development of methods that accelerate the healing of acute and chronic wounds should make up the primary aim of the wound management process.
Hemostasis starts immediately following injury due to the constriction of the damaged vessels, which limits the extent of blood loss. This leads to tissue hypoxia and acidosis, which decreases the effect of vasoconstriction and increases the level of vascular permeability for inflammatory cells. Platelets play the most important role at this stage through the formation of a blood clot (coagulation cascade) and the production of multiple signaling molecules such as PDGF, EGF, fibronectin, fibrinogen, histamine, serotonin and the von Willebrand factor.Citation16,Citation17 PDGFs are released from the alpha granules of platelets thus promoting mitogenicity and the chemotaxis of the neutrophils, macrophages, fibroblasts and smooth muscle cells at the wound site.Citation18
The increased infiltration of neutrophils, mast cells, monocytes and T lymphocytes into the wound site occurs during the inflammation stage,Citation19 and TGF-β, TNF-α, EGF, PDGF, VEGF, FGF, IL-1, IL-6, IL-8 and IL-12 are all present in the wound environment at this phase of healing.Citation12,Citation16,Citation19–Citation22 These mediators both control the inflammatory process and modulate epithelialization, collagen accumulation and angiogenesis.Citation20 TGF-β is a potent chemoattractant for immune cells and, as with PDGF, is involved in all the phases of the wound healing process.Citation23,Citation24
First, guided by chemokines, TGF-β and peptides produced by the bacteria present,Citation25 neutrophils begin to migrate into the wound so as to prevent infection.Citation10 However, once the influx of monocytes (the second type of wound-attracted cells) commences, the infiltration of neutrophils begins to decrease. Monocytes are attracted to the wound site by factors such as PDGF and TGF-β as well as by broken-down elastin and collagen products.Citation10 Monocytes undergo phenotypic transformation within the wound so as to form macrophages which are responsible for the further reduction of inflammation and the initiation of the proliferative phase of the healing process.Citation14 The phagocytic role of macrophages, which is based on the removal of destroyed cells and debris from the wound site, is crucial with respect to wound healing. Matrix metalloproteinases (MMPs) released by macrophages, monocytes and lymphocytes are responsible for cleaning the wound of dead and damaged tissue and are secreted under the control of TNF-α, IL-1 and IL-6. Lymphocytes, which appear in the late inflammatory phase, influence both fibroblast proliferation and collagen biosynthesis.Citation12 Oxygen presence is necessary for both the actively proliferating cells and the neutrophil respiratory burst in the wound bed,Citation26 thus requiring the initiation of neovascularization.
The principal cells involved in the proliferation phase consist of fibroblasts which, following stimulation by che-motactic PDGF, EGF and TGF-β gradients, migrate to the location of tissue damage.Citation12,Citation20 Fibroblasts constitute key cells responsible for the initiation of angiogenesis, epithelialization and collagen production. Angiogenesis is essential with concern to maintaining the granulation tissue and is associated with the activity of a large number of molecules (eg, VEGF, FGF, TGF-β and TNF-α).Citation16 Moreover, MMPs promote angiogenesis via the liberation of VEGF,Citation12,Citation27 which stimulates endothelial cell functions such as proliferation, migration, differentiation and survival.Citation18,Citation28 The formation of granulation tissue, the deposition of collagen and angiogenesis occur simultaneously with epithelialization and wound contraction. Fibroblasts secrete collagen type III and fibronectin so as to create mechanically strong tissue.
The fourth and final phase of the wound healing process consists of remodeling, ie, a balance between synthesis and degradation so as to attain well organized tissue. The granulation tissue matures to form a scar. Fibroblasts change to become myofibroblasts and, together with collagen and fibronectin, participate in the wound contraction process. The collagen type I content increases in favor of collagen type III and the fibers become cross-linked and aggregated into the form of fibrillar bundles which affect both the stiffness and tensile strength of the healing tissue.Citation26 FGF plays a very important role in this phase and is produced by keratinocytes, fibroblasts, endothelial cells, smooth muscle cells, chondrocytes and mast cells, some of which are involved in the formation of granulation tissue, epithelialization and tissue remodeling.Citation18 Epithelialization occurs on the surface of the wound; epithelial cells, stimulated by EGF, KGF and TGF-α, migrate and proliferate so as to cover the new tissue. MMPs, with the inhibitors thereof (tissue inhibitor of metalloproteinases [TIMPs]), play a pivotal role in terms of regulating cell migration (keratinocytes, fibroblasts, epithelial and inflammatory cells) in the wound by modifying the wound matrix.Citation22,Citation27 The healing process is complex and long-lasting, and the maximal tensile strength of human wounds (ie, 70% of normal skin) is attained after around 1 year.Citation16
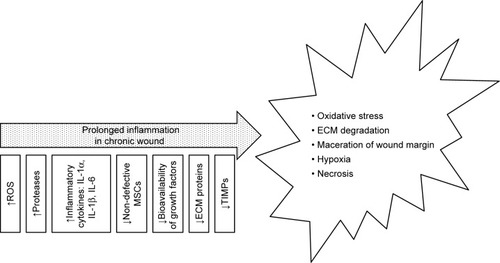
Non-healing (chronic) wounds present a serious problem both for patients themselves and the health care system, and a therapy is urgently required that accelerates the wound healing process, prevents secondary infection and which provides relief to patients. The risk factors of chronic wounds include diabetes, peripheral vascular disease, immunosuppression, acquired immunodeficiency and injury to previously wounded local tissue such as that caused by radiation therapy or certain chemicals.Citation10,Citation26 Impaired wound healing is associated with prolonged inflammation and an imbalance between the production and breakdown of the most important molecules in the wound which may be caused by excessive neutrophil infiltration, an abundance of pro-inflammatory cytokines, ROS, premature cell senescence, defective MSCs or the enhanced activity of wound proteases which degrade PDGF and TGF-β;Citation13,Citation14,Citation21,Citation26 moreover, the oxygen and moisture balance is disrupted. The inflammation phase with respect to normal wound healing lasts up to 7 days, while chronic wounds often stall in this phase and require longer healing periodsCitation13 () due to the increased quantity of ECM degradation products which promote inflammation, thus creating a self-perpetuating cycle.Citation21
Figure 2 Causes and effects of chronic wounds.
Abbreviations: ECM, extracellular matrix; TIMP, tissue inhibitor of metalloproteinase.
Graphene – structure, production methods, characterization techniques and biology-related properties
Graphene is an allotrope of carbon, whose structure consists of a planar sheet of single carbon atoms upon which each atom is bonded to three others densely packed within a honeycomb crystal lattice. In order to create a hexagonal lattice, the carbon must be subjected to spCitation2 hybridization. Subsequently, overlapped spCitation2 orbitals create three orbitals which are responsible for the formation of a σ covalent bond (three bonds between the nearest carbon atoms). The fourth bond consists of a π-bond, which is perpendicularly oriented (z-direction, out of the plane).
The potential applications of graphene are highly dependent on the production method employed and the form in which it is obtained (). However, it has proved difficult to date to obtain an ideal graphene which is both flat and homogeneous,Citation29,Citation30 ie, the creation of an ideal single layer graphene remains a major challenge. In order to fulfill the expectations of engineers, who use graphene for various applications, and scientists who use it in a wide range of research fields, a number of graphene production techniques have been (and continue to be) developed. It is already known that the best quality graphene in terms of structural integrity and electrical properties is obtained by means of the mechanical cleavage of highly oriented pyrolytic graphite.Citation31 Therefore, it will be necessary to evaluate the results of any new graphene production methods via a comparison of the materials produced with the properties of mechanically exfoliated pristine graphene.
Table 1 LPE and CVD advantages
Several strategies are presently being explored aimed at attaining reproducible and scalable graphene on various substrates ().
Currently, techniques such as scanning electron microscopy (SEM) and Raman spectroscopy are usually employed for the identification of the structure of graphene.Citation54,Citation55 The advantages of these techniques are shown in . shows a set of SEM images of samples collected simultaneously using two different detectors. The typical features of graphene grown on Cu foil are marked in the images.
Table 2 Selected techniques for graphene morphology characterization
Figure 3 SEM images recorded using (A) in-lens and (B) ESB detectors with the graphene features marked: the scale bars are 2 µm. Reprinted from Pasternak I. Synthesis and properties of graphene obtained on metallic and germanium substrates by CVD method [unpublished PhD thesis]; 2016; Copyright © 2016 Pasternak. (C) Raman spectra of the graphene.
Notes: The three most prominent peaks in the Raman spectrum of graphene consist of the G band at ~1,585 cm−1, the 2D band at ~2,700 cm−1 and the disorder-induced D band at ~1,350 cm−1 (for laser excitation energy of 2.33 eV). The G band, which is related to C−C bond stretching, is caused by the in-plane vibration of spCitation2 carbon atoms and corresponds to the first-order Raman-allowed E2g phonon in the center of the Brillouin zone (BZ). The D band, known as the disorder or defect band representing the ring breathing mode (A1g symmetry) of spCitation2 carbon rings, is induced by defects in the graphene lattice. The 2D band consists of the second order of the D band, sometimes referred to as an overtone of the D band, and is the result of the two phonon process involving two D phonons from the vicinity of the K point of the BZ. Unlike the D band, it does not need to be activated by proximity to a defect.Citation60 The intensity ratio of the G and D bands can be used to determine the number of defects in a graphene sample.Citation61 The number of layers of graphene can be defined on the basis of the line shape of the 2D peak as well as its intensity relative to the G peak. Single-layer graphene is characterized by a sharp, symmetrical, Lorentzian-shaped 2D peak with an intensity greater than that of the G peak. As the number of layers increases, the 2D peak becomes broader and less symmetrical accompanied by a decrease in its intensity.Citation62
Abbreviation: SEM, scanning electron microscopy.
![Figure 3 SEM images recorded using (A) in-lens and (B) ESB detectors with the graphene features marked: the scale bars are 2 µm. Reprinted from Pasternak I. Synthesis and properties of graphene obtained on metallic and germanium substrates by CVD method [unpublished PhD thesis]; 2016; Copyright © 2016 Pasternak. (C) Raman spectra of the graphene.Notes: The three most prominent peaks in the Raman spectrum of graphene consist of the G band at ~1,585 cm−1, the 2D band at ~2,700 cm−1 and the disorder-induced D band at ~1,350 cm−1 (for laser excitation energy of 2.33 eV). The G band, which is related to C−C bond stretching, is caused by the in-plane vibration of spCitation2 carbon atoms and corresponds to the first-order Raman-allowed E2g phonon in the center of the Brillouin zone (BZ). The D band, known as the disorder or defect band representing the ring breathing mode (A1g symmetry) of spCitation2 carbon rings, is induced by defects in the graphene lattice. The 2D band consists of the second order of the D band, sometimes referred to as an overtone of the D band, and is the result of the two phonon process involving two D phonons from the vicinity of the K point of the BZ. Unlike the D band, it does not need to be activated by proximity to a defect.Citation60 The intensity ratio of the G and D bands can be used to determine the number of defects in a graphene sample.Citation61 The number of layers of graphene can be defined on the basis of the line shape of the 2D peak as well as its intensity relative to the G peak. Single-layer graphene is characterized by a sharp, symmetrical, Lorentzian-shaped 2D peak with an intensity greater than that of the G peak. As the number of layers increases, the 2D peak becomes broader and less symmetrical accompanied by a decrease in its intensity.Citation62Abbreviation: SEM, scanning electron microscopy.](/cms/asset/be828683-866c-4d5c-a5e4-74457d9f9356/dijn_a_12190716_f0003_c.jpg)
Graphene properties with respect to biology (wettability, nanotopography and defects)
Recently published scientific reports and reviews have discussed both the cytotoxic effects of materials belonging to the graphene group and the biosafety of graphene nanotechnologies.Citation64–Citation68 This review, however, focuses on the stimulating properties of graphene and its putative action mechanism, particularly with respect to wound healing and the reduction of infection. First, we distinguished a selection of graphene samples in terms of their physical form, surface chemistry, topography and surface energy; the graphene production method determines its physical form. The chemical vapor deposition (CVD) method depends on the deposition of carbon atoms from hydrocarbon gas on a substrate (eg, copper foil). Graphene films can be transferred onto a wide range of substratesCitation53,Citation55 and are used in both physicsCitation69 and biology (eg, surfaces for cell growth).Citation70 While graphene solutions, in which graphene flakes and sheets are formed, are employed in the majority of toxicological studiesCitation65,Citation68,Citation71–Citation73 involving the dispersion of graphene in biological fluids, much less attention has been devoted to the biocompatibility of graphene in the form of a monolayerCitation63,Citation70,Citation74–Citation76 which can be used as a scaffold for the transplanting of cells into damaged tissue, especially with respect to acute and chronic wound therapy.
Wettability
Water molecule and protein absorption occurs once the substrate comes into contact with its biological surroundings. The behavior of the substrate in contact with water depends on its hydrophobic and hydrophilic surface properties. GO consists of a highly defective graphene sheet functionalized with oxygen groups (hydroxyl, carboxyl, and epoxy) which evinces high levels of hydro-philicity and protein absorption. Reduced graphene oxide (rGO) is produced via the reduction of GO using high temperatures or chemicals and is considered to have the same favorable level of solubility as GO despite the hydrophobic nature of the pristine form of graphene.Citation77 Huang et alCitation78 indicated that a graphene film produced by means of the dispersing method (in a solvent of tetrahydrofuran, distilled water and dimethylformamide) exhibits reversible hydrophobic and hydrophilic transition in response to UV illumination and dark storage, respectively. Moreover, graphene can be further modulated so as to obtain hydrophobic and hydrophilic surfaces.Citation79
Wettability affects the ability of cell adhesion-mediated proteins to attach to the substrateCitation80 and, consequently, determines cell adhesion.Citation81 Cells attach to the underlying substrate (protein layer) by means of focal contacts, ie, adhesive connections containing a large number of proteins (eg, integrins). The extracellular parts of integrins bind to the ECM, their integral parts anchor integrin into the cell membrane and their intracellular parts bind to the focal adhesion proteins thus forming a physical link between the ECM and the actin cytoskeletal network.Citation63,Citation82,Citation83 Focal complexes that connect cells with the external environment are crucial for the functioning of cellular processes and mechanisms such as mechano-sensoring, spreading, cell migration and proliferation.Citation84 The focal adhesion assembly responds to matrix stiffness,Citation85 a phenomenon that is used by researchers for the qualitative and quantitative analysis of the effect on cellular behavior of the nanotopography of the various substrates used in regenerative medicine.Citation63,Citation79,Citation86–Citation88 James and TourCitation88 indicated that there are numerous permutations of graphene differentiated in terms both of their physical form and the number of layers. Dai et alCitation30 measured water contact angle dependence on the number of graphene layers and summarized that the wettability of graphene depends on the number of layers, the graphene preparation substrate and its surface chemical composition. They determined that in the case of the presence of more than six layers, the water contact angle value of graphene equaled that of graphite.
Nanotopography
Nanotopography makes up a fundamental factor with respect to the design of biomaterials intended for tissue engineering applications.Citation89 The surface properties of graphene can be described according to its morphology, ie, the presence of wrinkles, fluctuations and N2 adsorption, which alters its mechanical and chemical properties. Wrinkles may arise as the result of thermal stress occurring during the production process or due to the transfer technique employed, ie, it may be influenced by the metal substrate.Citation90,Citation91 Corrugation appearing on transferred surface-grown graphene has been determined at ~2–15 nm in height and ~20–100 nm in widthCitation90 or even smaller (1–2 nm).Citation79 Graphene deformations also include ripple formations with a height of up to 1 nm in suspended graphene membranes and the formation of crumples which may be produced via the rapid evaporation of aerosol droplets.Citation92 Roughness caused by the nanostructure of the graphene is capable of changing its hydrophobicityCitation93 and, consequently, its interaction with molecules and cells (eg, focal adhesion, cytoskeleton contraction).
Graphene exhibits an extremely high specific surface area (theoretically 2,630 m2·g−1)Citation30 depending on the preparation method employed and the number of layers, whereas the specific surface areas of the various graphene derivatives range from 600 to 1,600 m2·g−1.Citation94 In conclusion, the various physical forms and chemical structures of graphene derivatives exert differing effects on cells.Citation66
Defects
Each graphene production method leads to the production of differing properties and quality levels and influences the number of defects in the material,Citation55 all of which are capable of affecting the material’s degree of impact on cells and, consequently, its therapeutic effect. Defect-free graphene does not exist; defects in the structure of graphene may arise spontaneously during the production process or may be introduced through changing the properties of the materialCitation29,Citation55 which can be identified via the application of Raman spectroscopy ().Citation54 While certain defects exert a favorable effect, such as increasing the reactivity of the graphene, others must be eliminated prior to medical application, including hexagons that transform into pentagons (the Stone–Wales defect, SW), single (the absence of one lattice atom) or multiple vacancies, dimensional defects (eg, dislocations – line defects) and defects along the edges.Citation29,Citation54
The influence of graphene on cells and the putative mechanism of this process
The surface properties of graphene allow for its use as a platform for cell adhesion and even induced cell proliferation.Citation75 It has been shown previously that graphene monolayers are non-toxic, stimulate the mitochondrial activity of mouse fibroblasts (L929)Citation63 and potentiate the adhesion and proliferation of osteoblasts and MSCs.Citation70,Citation79,Citation95 In addition, two-dimensional multilayer pristine graphene film has been found to demonstrate good biocompatibility with human stromal fibroblasts.Citation96 Graphene–polycaprolactone composites have also been found to exhibit good biocompatibility employing L929 fibroblasts.Citation97 As mentioned above, the presence of fibroblasts is crucial in the proliferation phase at which time they are recruited into the wound. Confirmation of the biocompatibility of graphene and fibroblasts is essential prior to considering further research on its use in the wound healing process. The use of graphene as a scaffold material exerts an influence on cells by means of its nanotopography; cells are influenced by mechanical forces in the local environment to which they respond via nano-transduction either by adaptation or death. The expected role of graphene in the wound healing process is to mimic the architecture of the native ECM in such a way that proliferation, migration and spatial organization lead to enhanced wound closure, an increase in the strength of the new tissue formed and a reduction in scar formation.
Immunofluorescence staining has revealed that L929 cells created more focal adhesions, and the migration of the cells appeared to be more regular, than on a glass control of similar roughness to graphene.Citation63 Cell migration depends on the roughness of the substrate,Citation82,Citation89 according to which cells that migrate on graphene require a greater amount of energy for the dissolution of old adhesions, tail retraction and the balancing of internal and external forces. Increasing the effective surface energy (associated with moderately rough substrates) enhances the total amount of work per unit area required for full detachment,Citation89 which may result in increased mitochondrial activity and the activation of pathways involved in all the processes concerned with cell migration and other processes essential for cell growth and proliferation. Rho family GPTase are involved in the translation of the signals that regulate the various cellular processes such as cell adhesion, actin cytoskeleton re-organization, polarity, cell growth, proliferation and chemotaxis ().Citation98–Citation100 Rougher and stiffer substrates appear to provide better cell scaffolds resulting in enhanced intracellular tension and an increase in the number of focal adhesions and cell proliferation.Citation63,Citation89,Citation100 Focal adhesions play an important role in the transduction of mechanical signals, and a complex network of signaling pathways is involved in the cellular response (). Integrin activation leads to the activation of focal adhesion kinase (FAK) and Src kinase. The activation of FAK may lead to enhanced cell proliferation as mediated by extracellular signal-regulated kinases (ERKs) via various signaling pathways.Citation98,Citation100 Moreover, FAK is able to regulate cell migration by means of binding to and promoting the Src-mediated phosphorylation of p130Cas and via the regulation of the RhoA–ROCK pathway.Citation98,Citation100,Citation101 The absence of FAK negatively affects the production of lamellipodia by the cells on the edge of the wound.Citation101
Figure 4 Mechanotransduction of fibroblasts in response to contact with graphene substrate.
Notes: External cues occur in terms of intracellular regulation through a number of signaling cascades including the Rho family GPTase (Rho, Rac and Cdc42) and the activators thereof. These proteins induce the creation of stress fibers and enhanced focal adhesions and lead to the formation of filopodia and lamellipodia.
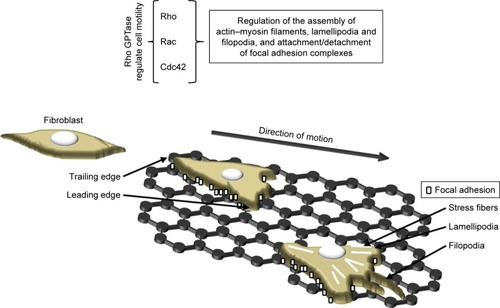
The actin cytoskeleton is composed of actin filaments and, through combining microfilaments with multiple actin binding proteins, it creates various cellular forms such as cortical actin networks, stress fibers within the cytoplasm, shrinkage rings formed during cytokinesis and surface tabs (lamellipodia, filopodia) in the cells. A strong actin network is required in order to stabilize the cells on the substrate (). Marked stress fibers observed in cells cultivated on a graphene scaffoldCitation63 indicate a struggle with the substrate architecture and stiffness forces. Kim et alCitation102 and Zhang et alCitation103 suggested that GO micropatterns might provide a suitable cell-guiding substrate for the purposes of tissue engineering and regenerative medicine. Moreover, triangular GO micro-patterns fabricated using meniscus-dragging deposition and photolithography techniques have been determined to enhance the speed, distance and directionality of L929 fibroblasts.Citation102 Due to its ability to guide cells in a specific direction (essential with respect to wound closure and scar formation), the use of a micro-pattern graphene substrate for wound treatment is particularly advantageous. However, Keshavan et alCitation104 showed that the response to identical surface cues (topographical and chemical) is a cell-type dependent mechanism. They noted the higher primary adhesion of Chinese hamster ovary cells on poly-d-lysine coated micro-patterned single-layer graphene (SLG) compared to that on adjacent SLG ablated stripes. However, during the incubation period, the cells were observed to migrate onto the adjacent SLG ablated stripes, which proved a more favorable environment for their subsequent proliferation. Interestingly, these same surfaces resulted in differing primary neuron cell arrangements.Citation105 Osteoblasts have been found to adhere to GO patterns exhibiting highly aligned, oriented and elongated actin filaments which have adapted to the pattern width.Citation103 Moreover, the authors also point out that polarized cells with high cytoskeleton tension and traction forces are capable of overcoming the strong adhesion between cells and GO, thus leading to higher cell contractility and mobility. Following the discovery of the surface patterning of graphene, new tissue engineering applications emerged employing these unique properties. Finally, it has also been shown that materials with unique nano-topographical characteristics – including graphene – offer properties which are similar to those of growth factors.Citation11 To sum up therefore, cells are affected by the mechanical properties of the scaffold, its nano-architecture and chemical signals (eg, growth factors, cytokines and ECM modifying enzymes bound to the graphene). These stimuli lead to tensile, compressive and shear stress which lead to changes in the cell structure and the initiation of signal transduction (eg, FA-Rho GTPase crosstalk) ().
One of the additional benefits of graphene consists of its antibacterial activity, which may offer an alternative to the use of antibiotics in the wound healing context. A range of bacteria limitation/death mechanisms initiated by both graphene and its derivatives have been described in the literatureCitation14,Citation106–Citation109 depending on the diversity of the various forms of graphene and their chemical properties and the structure of the bacteria wall. The antibacterial activity of graphene materials is associated with membrane stress, which may be accompanied by ROS-dependentCitation110 or -independent oxidative stress.Citation111 Some authors report that the sharp edges of graphene cut through the cell membranes of bacteria thus causing lethal damage to cellular integrity.Citation112,Citation113 A further mechanism consists of the isolation of bacteria through wrapping/trapping them in a sheet-form blanket of graphene thus limiting bacterial access to nutrients.Citation108 Shuai et al proposed GO–Ag nanosystem (polymer scaffold containing 1 wt% GO–1 wt% Ag) with synergistic effect on antibacterial action via combining the capturing effects of GO nanosheets and the killing effects of Ag and showed bacterial inhibition rate >95%.Citation114 Escherichia coli cells were found to lose their cellular integrity accompanied by severe membrane damage following 2.5 hours of incubation with 100 µg mL−1 GO nanosheets;Citation115 moreover, the authors indicated that a large amount of phospholipids were freed from the bacteria cell membranes as a result of interactions between the graphene and lipid molecules. Kurantowicz et alCitation116 determined that 250 µg mL−1 of pristine graphene, GO and rGO consistently inhibited the growth of Salmonella enterica and Listeria monocytogenes by 100%. They further demonstrated that bacterial cells interacted with the sp3-hybrized oxidative group of the GO and distributed themselves over the surface thereof, while the bacterial cells were arranged at the edges of the pristine graphene and rGO. Moreover, they also showed that pristine graphene and rGO exhibit lower levels of antibacterial activity than does GO. On the other hand, Barbolina et alCitation117 pointed out that graphene contaminants are responsible for the reported antibacterial properties rather than graphene alone and concluded that GO purification is crucial in order to ensure the true biological effect of the material. The authors, using highly purified and thoroughly washed GO, failed to discover either bactericidal or bacteriostatic properties over a broad concentration range with concern to planktonic cultures of either E. coli or Staphylococcus aureus.
In addition, the antiviral action of graphene has been demonstrated by Ye et alCitation118 who suggested that this property can be attributed to the unique single-layer structure and negative charge. A non-cytotoxic concentration (6 µg mL−1) of GO was added to PK-15 cells infected with pseudorabies virus and Vero cells infected with porcine epidemic diarrhea virus and was found to suppress both infections. The authors noticed that the GO in the cell culture did not block viral replication and the subsequent spread to neighboring cells, rather the pre-incubation of the viruses with GO induced the significant inhibition of infection. Thus, they suggested that GO inhibits virus infection by inactivating virus particles prior to entering cells. They concluded that the antiviral action mechanism is based on the electrostatic interaction of negatively charged sharp-edged GO with positively charged virus particles, resulting in viral morphology damage (both the envelope and the spikes were destroyed) and subsequent inactivation. Moreover, the authors indicated that both GO and rGO exhibit similar antiviral activity and that the oxygen-containing group is not essential for the initiation of such activity. Song et alCitation119 demonstrated that negatively charged GO efficiently captured the enteric EV71 and H9N2 viruses and that GO surfaces are capable of destabilizing enveloped viruses.
Graphene has also been investigated with respect to hemocompatibility and angiogenic action.Citation65,Citation120–Citation122 GO was shown to exhibit prothrombotic properties which are able to activate Src kinases and induce the release of calcium from intracellular stores; the prothrombotic character was shown to be dependent on the surface charge distribution.Citation123 Jaworski et al,Citation65 based on the results of experiments on chicken embryo red blood cells, demonstrated that different forms of graphene exhibit differing hemocompatibility depending on the production method employed and the surface modification. In addition, Mukherjee et alCitation120 demonstrated the pro-angiogenic activity of graphene and proposed a mechanism based on the intracellular formation of ROS and reactive nitrogen species and the activation of phospho-eNOS and phospho-Akt. Shine et alCitation122 reported that with higher concentrations of graphene (from 0.25% to 1% in the composite), the expression level of angiogenic proteins was enhanced in human mesenchymal stem cells (hMSCs) cultured on calcium silicate/graphene composites. Park et alCitation121 indicated that the incorporation of rGO flakes into MSC spheroids and monolayer cultures promoted the expression of proangiogenic growth factors (VEGF, FGF-2, and HGF) and that the highest expression concerned hybrid spheroids with 5 µg mL−1 rGO flakes. The authors also demonstrated that enhanced cell–ECM interaction through the incorporation of rGO flakes into MSC spheroids leads to an increased amount of VEGF via mediated FN-integrin binding, which leads to the enhanced expression of phosphorylated FAK, phosphorylated ERK and thus VEGF.
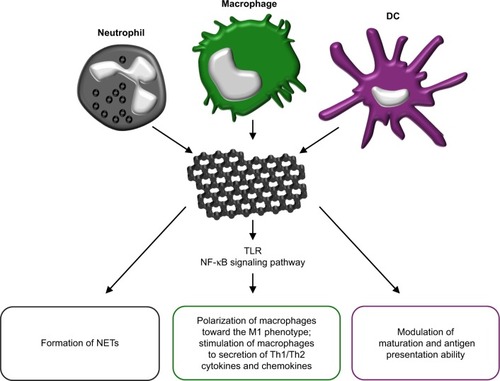
Graphene and its derivatives have also been shown to possess immunomodulatory properties depending on their physicochemical features and functionalization.Citation124 These nanocompounds are able to modulate the functions of phagocytic immune cells that participate in supporting the normal wound healing process, including neutrophils,Citation125 macrophagesCitation19 and dendritic cells (DCs).Citation126 Neutrophils constitute the first inflammatory cells recruited to the wound tissue from the blood and both act to sterilize the wound via the production of antimicrobial peptides and proteases and to regulate the inflammatory response via the secretion of multiple cytokines and growth factors.Citation125 Recently, it has been determined that primary human neutrophils exposed to small (50–300 nm) and large (10–40 µm) sheets of GO produce neutrophil extracellular traps (NETs), ie, fibrillar networks that contribute toward defense against pathogens ().Citation127 The production of NETs was seen to be dependent on the size of the GO sheets and was associated with both ROS production and the influx of Ca2+. Despite the fact that the GO-induced formation of NETs eventually led to neutrophil-cell death (NETosis),Citation127 it represents an important mechanism in terms of immobilizing and killing medically relevant bacteria.Citation128 Macrophages fulfill a large number of beneficial functions with respect to promoting the wound healing process, including the regulation of the inflammatory response, the removal of neutrophils/apoptotic cells, the promotion of angiogenesis, fibroblast proliferation and ECM reorganization.Citation19 In general, the various nanomaterials (gra-phene family nanomaterials [GFNs]) of the graphene family are cytotoxic in a dose-dependent manner and induce differing types of cell death (apoptosis, autophagy and necrosis) in macrophages.Citation124 However, sub-cytotoxic concentrations of GFNs and the appropriate variations in their physicochemical properties are capable of modulating the immune functions of these cells. It has been shown that a sub-cytotoxic dose of pristine graphene stimulates primary murine macrophages and immortalized macrophages into secreting Th1/Th2 cytokines (IL-1α, IL-6, IL-10, TNF-α and GM-CSF) and chemokines (MCP-1, MIP-1α, MIP-1β and RANTES), most probably due to the toll-like receptor (TLR)-dependent activation of the nuclear factor-κB (NF-κB) signaling pathway.Citation129 (). Moreover, GO induces an inflammatory response (together with autophagy) in murine RAW 264.7 macrophages by activating TLRs (TLR4 and TLR9) and their downstream MyD88-, TRAF6- and NF-κB-dependent signaling pathways.Citation130 In addition, GO sheets polarize macrophages toward the M1 phenotype and enhance their pro-inflammatory response in a size-dependent manner. Larger GO sheets exhibit a stronger interaction with the TLR4 plasma membrane, resulting in NF-κB activation and M1 polarization both in vitro and in vivo.Citation131 The M1 immune polarization effect has also been observed with respect to monocytes treated with GO functionalized with amino groups.Citation132 DCs infiltrate wounds quickly following injury and accelerate early wound closure, most likely via the secretion of factors that increase cellular proliferation, granulation tissue formation and angiogenesis.Citation126 GO and other carbonaceous nanoparticles, ie, C60 fullerenes and C60-TRIS fullerenes, have been shown to be taken up by conventional DCs and differentially modulate the antigen presentation ability of these cells (). GO only (ie, not fullerenes – a further form of carbon along with graphite and diamond) was found to downregulate intracellular levels of immunoproteasome subunit low molecular mass polypeptide 7 and thus decrease the level of antigen processing in DCs.Citation133 Another study demonstrated that pure GO induces the higher maturation and stronger production of TNF-α and IL-1β in human DCs than does polyvinylpyrrolidone-functionalized GO.Citation134 Moreover, the treatment of DCs with a mixture of GO and a glioma peptide antigen enhances their anti-tumor immune response.Citation135 Taken together, the immune regulatory properties of GFNs are determined largely by the dosage and the variation in their physicochemical properties.
Figure 5 The immunomodulatory properties of grapheme.
Notes: Graphene and its derivatives may act on neutrophiles, inducing neutrophil extracellular traps (NETs) formation. Moreover, graphene induces TLR-dependent activation of NF-κB signaling pathway in macrophages, resulting in polarization of macrophages toward the M1 phenotype and stimulation of secretion of Th1/Th2 cytokines and chemokines. Graphene derivatives also modulate maturation of dendritic cells and their antigen processing and presentation capacity.
Abbreviations: DC, dendritic cell; TLR, toll-like receptor.
Graphene has also been used in in vivo studies usually incorporated into composites. A study by Deepachitra et alCitation1 indicated that GO incorporated into a collagen-fibrin biofilm resulted in no adverse effects and enhanced the wound healing process in Wistar rats. In addition, they noticed an increase in the mechanical strength of the composite films with GO and rat healing tissue, thus indicating its potential use as a structural reinforcement material. Zhong et alCitation136 proposed the use of GO as a delivery system for recombinant TIMP-1 (a metalloproteinase inhibitor) and demonstrated the continuous release of TIMP-1 from the GO for up to 40 days. The subcutaneous administration of TIMP-1-GO to rats with experimental wounds has been shown to promote both vascularization and collagen regeneration. Mahmoudi et alCitation15 prepared nanofibers containing GO nanosheets via the electrospinning of chitosan–PVP and demonstrated that GO promoted adhesion and the viability of human skin fibroblast cells, enhanced bactericidal capacity and accelerated the wound healing process in Sprague Dawley rats.
MSCs and their putative mechanism of action in wound repair
The use of MSCs in the treatment of wounds raises great hope for regenerative medicine. MSCs have the capacity for multi-lineage differentiation.Citation83,Citation137,Citation138 They adhere to the surface of the culture vessel, exhibit fibroblast-like morphology and develop into symmetrical colonies. They express such antigens as CD73, CD90 and Cd105 and should not express CD14, D19, CD34, CD45, CD11b, CD79a and HLA-DR surface molecules.Citation3,Citation6,Citation9,Citation139–Citation142 Bone marrow, the umbilical cord, adipose tissue, placenta and cord blood all provide sources of MSCs () containing the therapeutic potential for the treatment of wound healing disorders.Citation143 With respect to normal cutaneous wound healing, MSCs are mobilized from their host sources to the injury site where they support skin repair despite hypoxia and a lack of nutrients.Citation22,Citation144 MSC therapy is dependent on both the sufficient extent of MSC engraftment at the injury site and cell survival within the wound. While autologous MSC transplantation provides a number of reasons for optimism, allogenic MSC transplantation is also feasible since these cells, as with group O red blood cells, are immunologically silent.Citation145,Citation146 A conditioned medium of MSCs (MSC-CM), which includes bioactive molecules secreted by the MSCs in the culture, has also demonstrated regenerative effects with concern to wound healing tissue.Citation2,Citation140,Citation145,Citation147
Figure 6 The effects of MSCs and graphene in the wound healing process.
Abbreviations: VEGF, vascular endothelial growth factor; PDGF, platelet-derived growth factor; EGF, epidermal growth factor; FGF, fibroblast growth factor; HGF, hepatocyte growth factor; IFNλ, interferon λ; IGF, insulin-like growth factor; KGF, keratinocyte growth factor; MMP-9, matrix metalloproteinase-9; MSC, mesenchymal stem cells; PGE2, prostaglandin E2; RNS, reactive nitrogen species; SDF-1, stromal cell-derived factor-1.
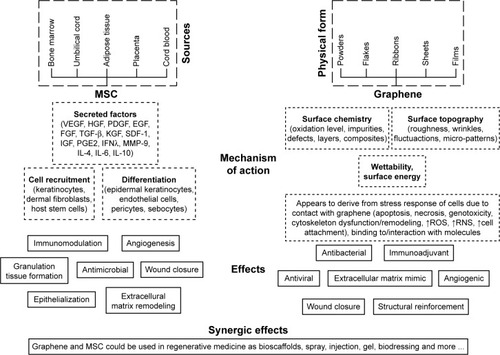
One of the most important features regarding the clinical application of MSCs consists of their ability to recruit other cells for the purpose of tissue repair, concerning which differentiation and paracrine signaling have been identified as mechanisms of their action.Citation3,Citation139 It has been shown that MSC-CM enhances wound closure via the acceleration of the in vitro migration of fibroblasts and keratinocytes.Citation147 The wound healing process requires interaction between cells, ECM proteins and biomolecules (growth factors, cytokines and chemokines), and MSCs play a key role in the coordination of individual damaged tissue regeneration processes.Citation11,Citation139 The number of connections with the ECM exerts a significant impact on the strength of the new tissue that replaces lost tissue in the wound. To date, a number of routes have been studied concerning the introduction of MSCs into the organism for wound healing purposesCitation146,Citation148,Citation149 and the most recent study conducted on this theme revealed that the subcutaneous injection of MSCs provided a much more efficient method than intravenous injection with concern to the healing of skin wounds.Citation150 The use of exogenous MSCs also provides a promising strategy with respect to the treatment of non-healing wounds as in the case of those caused by diabetes, vascular insufficiency and several other medical conditions.Citation22,Citation151 While it is believed that MSCs have the therapeutic potential for application with concern to wound-healing disorders, the action mechanism is still not fully understood.Citation143
Although the injection of MSCs into the blood stream leads to healing to a certain extent, the delivery of MSCs on scaffolds provides a significantly more potent therapeutic approach (ie, it is site-specific). Multi-functional scaffolds have the potential to guide the adhesion, growth and differentiation of MSCs so as to form skin-functional and structural tissue. When designing suitable MSC scaffolds, it is important to remember that the physicochemical properties of the biomaterials used may well determine and change the fate of MSCs.Citation138 The stiffness, elasticity, porosity and reactivity of the material may potentially affect cellular behavior through the forces applied, the activation of various molecules during the cell adhesion process and interactions with the scaffold.Citation152–Citation154 Thus, the creation of an effective physical platform will allow for the control of various processes such as the attachment, proliferation and differentiation of MSCs. Moreover, it may also assist in the development of a specific required biological effect via the direction of the behavior of the cell. A detailed knowledge of the signaling mechanism activated by scaffold–cell interactions would reveal the direction of a range of cellular activities thus making it possible to achieve a specific aim via the manipulation of the physiochemical properties of the biomaterial scaffold.
MSC as a producer of bioactive molecules
The secretion of bioactive factors is thought to constitute the principal MSC action mechanism during the wound healing process ().Citation7,Citation140,Citation143 The paracrine effect of MSCs is based on the release of growth factors, ILs and other bioactive molecules secreted or packaged into extracellular vesicles or exosomes.Citation8 Growth factors play a pivotal role in the modulation and coordination of cellular processes in all phases of the wound healing process () and the sustained release of these molecules coupled with their bioactivity both stimulate the wound environment and promote wound closure.
The immunomodulatory effects of MSCs are related to the reprogramming of macrophages from type M1 to type M2 anti-inflammatory phenotypes which suppresses the proliferation of T cells, modulates TNF-α production, reduces the NK cell function in the inflammatory phase and lowers the level of IFN-γ activity in the process.Citation8–Citation10,Citation144,Citation155–Citation157 MSCs secrete factors that upregulate the secretion of IL-10 and inhibit neutrophil infiltration into the wound.Citation10 They also secrete IL-10 by themselves.Citation139 MSCs also release TGF-β1 and HGF, which suppresses T cell proliferation, and PGE2, which prevents the differentiation of CD4+ T cells into Th17 cells.Citation157 MSCs also release anti-inflammatory cyto-kines, eg, IL-4, which is important with respect to chronic wound healing.Citation10 The secretion of IL-6 by MSCs has been demonstrated both in mice and in humans and its dual nature (pro-inflammatory and/or anti-inflammatory effects) has been emphasized.Citation4 MSCs produce IL-6 in a p38MAPK pathway-dependent manner.Citation158 Tamama and KerpedjievaCitation145 reviewed the relevant literature and summarized that both MSCs themselves and MSC-CM encourage wound repair and that multiple growth factors and cytokines (VEGF, bFGF, IL-6, IL-8) are involved in the MSC-mediated wound healing process.
MSCs promote new vessel formation through the release of VEGF.Citation144 An et alCitation150 revealed that autophagy in MSCs improves cutaneous wound healing via the paracrine secretion of VEGF and the direct phosphorylation of ERK, resulting in the further promotion of the VEGF-induced vascularization of endothelial cells. MSCs have been found to extensively express those factors involved in vessel stabili-zation, smooth muscle cell migration and matrix remodeling such as TGF-β, PDGF-β and MMP-9 as well as high levels of stromal cell-derived factor-1 (SDF-1) α chemokine, which is known to be involved in the recruitment and retention of proangiogenic macrophages and MSCs themselves.Citation83,Citation159
Wu et alCitation160 revealed that BM-MSC-treated wounds exhibited accelerated wound closure in normal BALB/c mice and diabetic mice compared with fibroblast- or vehicle control medium-treated wounds. Shin et alCitation161 demonstrated that the administration of tonsil-derived MSCs into wound beds significantly promoted the repair of surgical defects in mice. Luo et alCitation162 discovered that MSC-treated wounds exhibited a more regular fiber alignment than did the wounds of the control animals and, moreover, that the former developed both hair follicles and sweat glands.
Li et alCitation140 revealed that the proliferation and migration of dermal fibroblasts was enhanced by MSC-CM; moreover, the activity of the MMPs thereof and the expression of TGF-β3 increased following MSC-CM treatment. The authors also indicated more rapid wound healing and less scarring following the application of MSC-CM in vivo. High levels of TGF-β3 and low levels of TGF-β1 were found in an embry-otic wound microenvironment in which scar-free healing generally occurs.Citation163 Hence, the ratio of TGF-β3 to TGF-β1 appears to be an important factor with concern to scar-free wound healing. It is proposed therefore that the anti-scarring potential of TGF-β3 released by MSCs should be employed in medical applications. The benefit of employing MSCs compared to the exogenous administration of TGF-β3 lies in the greater action spectrum associated with the release from the MSCs of a range of other bioactive factors. Scars occur as the result of the excessive amount of the ECM deposited by fibroblasts in the wound bedCitation144 and feature a lack of follicles and nerve endings. In addition, the tensile strength of the new tissue is substantially weaker. MSCs promote anti-scarring properties through the secretion of PGE2, which induces the increased expression of IL-10 via T cells and macrophages.Citation156 The MSC upregulation of IL-10 decreases the expression of the IL-6 and IL-8 necessary for the prevention of the occurrence of an excessive increase in the deposition of collagen in the wound.Citation144,Citation156 The HGF secreted by MSCs acts to modulate fibroblasts via the down-regulation of the expression of TGF-β1 which drives both myofibroblast differentiation and the production of collagens types I and III.Citation144,Citation157 HGF also enhances the degradation of the ECM through the upregulation of the fibroblast expression of MMPs.Citation157 MMP-9, ie, one of the MMPs released by MSCs exhibits a high degree of activity against gelatin and degrades other ECM molecules including collagens, laminin and aggrecan.Citation27
The antimicrobial activity of MSCs is based on the secretion of antimicrobial factors such as LL-37 and immune-modulative factors which upregulate the killing of bacteria and the phagocytosis thereof by immune cells.Citation10,Citation22 LL-37 makes up one of the antimicrobial peptides and proteins group – also known as “host defense peptides” and the low production thereof is associated with skin disorders.Citation141
The environment (ie, the tension of oxygen) makes up a further crucial factor, which influences the behavior of MSCs (gene expression, the release of cytokines and other factors). Chen et alCitation2 revealed, based on real-time PCR analysis, that MSCs treated under hypoxic conditions expressed significantly greater amounts of EGF, KGF, IGF-1, VEGF-α and SDF-1 but lower amounts of TGF-β1 than did dermal fibroblasts. In addition, EGF is, for example, an important growth factor with respect to re-epithelialization and the promotion of wound closure. Thus, it is also possible to control the function of MSCs via the triggering of the cultivation conditions.
In summary, MSCs contain a wide range of growth factors and cytokines, which work in synergy to accelerate the wound healing process. The features of the abovemen-tioned bioactive molecules with respect to cell recruitment and MSC differentiation in the context of wound healing will be further described below.
MSC as a coordinator of cell recruitment and differentiation
The therapeutic action of exogenously delivered MSCs and MSC-CM lies in its selective recruitment of host cells to the injury site and the direct differentiation of MSCs. SDF-1 is thought to play an important role in terms of the recruitment of stem cells from bone marrow to the injury site via a CXCR4-dependent mechanism. The enhancement of SDF-1 signaling within injured tissue can also be used to augment cellular transplantation.Citation6,Citation164 MSCs provide support for native cells at the injury site via the secretion of a variety of pro-survival and pro-migratory cytokines and growth factors.Citation165 Chen et alCitation2 found that MSC-CM significantly enhances the migration and proliferation of keratinocytes and endothelial cells; the authors suggested that the various factors released by MSCs (VEGF-α, IGF-1, EGF, KGF, angiopoietin-1, SDF-1, macrophage inflammatory protein-1 alpha and beta and erythropoietin) recruit macrophages and endothelial cells to the wound, thus enhancing the healing process. Lee et alCitation166 demonstrated that MSC-CM harvested under hypoxia promoted fibroblast migration in vitro and dermal wound closure more rapidly than did MSC-CM collected under the normoxic culture condition. Rodriguez-Menocal et alCitation167 revealed that healthy donor MSCs were significantly better than MSCs derived from chronic wound patients in terms of inducing normal fibroblasts to migrate; the authors also indicated that bone marrow-derived MSCs induce fibroblast migration in a dose-dependent manner. Increased fibroblast migration was observed in the presence of MSCs in a low concentration (10% of the population); however, an increased MSC concentration (20% or higher) led to a decrease in the migration of fibroblasts. On the other hand, the attenuated infiltration of inflammatory cells has been observed following the transplantation of tonsil MSCs into mice.Citation161 It might be concluded therefore that the secreted factors of MSCs create a specialized cell recruitment microenvironment and offer great potential with respect to stem cell-based therapies.
Certain evidence suggests that MSCs may also induce tissue regenerative processes through in situ differentiation. When MSCs were cocultured in vitro with keratinocytes they exhibited trans-differentiation to keratinocytes.Citation157 Mishra et alCitation168 demonstrated that a keratinocyte-conditioned medium induced MSC differentiation to dermal myofibroblast-like cells and also enhanced the expression of cytokines including SDF-1, IL-8, IL-6 and CXCL5. The authors also noticed the organization of MSCs around the keratinocytes in vitro and in vivo, which they subsequently compared to the wound repair granulation phase. Sasaki et alCitation169 indicated that MSCs have the capacity to differentiate into multiple skin cell types including keratinocytes, endothelial cells and pericytes in cutaneous wounds in mice following intravenous injection. Wu et alCitation160 demonstrated that the MSC treatment of wounds enhanced the healing process in both normal and diabetic mice. Thus, MSCs play a dual role in wound healing by both producing specific factors and differentiating to specific cell types.
The senescence of MSCs
Cultured primary cells exhibit a limited division number and the aging of MSCs appears to present a major problem with respect to clinical applications, which require a significant number of cells.Citation170–Citation172 Many authors have remarked that later passage MSCs exhibited morphological abnormalities (an increase in cytoplasm granularity and the formation of vacuoles), enlargement and slower proliferation rates.Citation170,Citation171,Citation173–Citation175 They concluded that the long-term culturing of MSCs results in an increase in cell senescence. Turinetto et alCitation171 suggested that it is difficult to predict which passage or number of cell divisions characterize the senescent state of MSCs due to variations in terms both of seeding densities and the time of harvesting. Whitfield et alCitation174 observed a human adult bone marrow stromal cell population during increased in vitro passaging and discovered that the cells increased in size over time; they concluded that the larger MSCs had originated from several different generations and that they had exited the normal cell cycle, thus no longer fulfilling the MSC criterion of exhibiting the capacity for self-renewal in vitro.
Outlook and conclusion
Graphene–MSC dressings present a potentially attractive therapy involving the alteration of the wound environment via both mechanical and chemical stimuli. It is possible to create and alter the trophic functions of MSCs via dynamic ECM–cytoskeletal interactions, cell–cell contacts and soluble and transcription factor signaling,Citation8 and they can be poten-tiated by means of graphene scaffolds. Kalbacova et alCitation70 were the first to demonstrate that graphene in the monolayer form is non-toxic for MSCs and, moreover, stimulates the growth thereof. Kazantseva et alCitation152 showed that graphene-augmented inorganic nanofiber scaffolds do not impede the normal growth of adipose-derived hMSC; moreover, they are able to both direct the preferential orientation and alter the morphology of MSCs. Shine et alCitation122 showed that hMSCs are capable of uniformly covering calcium silicate/graphene composites and that composites with a higher content of graphene (1%) enhance cell proliferation. Going forward, the greatest challenge for scientists is to produce graphene in a way that closely resembles the nanotopography of the natural ECM of human skin and to achieve a similar biological effect. The direct delivery of MSCs to wounds may induce rapid cell death;Citation143,Citation157 thus, the introduction of MSCs via graphene nanoscaffolds presents a promising alternative delivery method which is capable of minimizing unprogrammed cell death. Li et alCitation176 proposed a 3D graphene foam scaffold loaded with MSCs in connection with wound healing which both exhibited good biocompatibility and promoted the growth and proliferation of MSCs. The authors emphasized that the mechanical properties of graphene foam and MSCs strongly promote integration with the host tissue, which results in enhanced and more rapid wound closure. Li et alCitation176 concluded that a 3D graphene foam together with MSCs synergized so as to promote wound closure via the enhancement of early vascularization accompanied by a reduction in scarring in an animal model, most probably due to the specific electrical properties of 3D graphene foam. Chu et al showed that hybrid scaffold containing in wound healing in diabetic rats.Citation177 The results of experiments conducted to date allow us to conclude that graphene in combination with certain cells has the potential for use in the enhancement of the healing of complicated wounds and that MSCs introduced to wounds directly via graphene scaffolds presents a viable alternative to traditional dressing materials. We believe that the stiffness and nanotopography of cell culture scaffolds like graphene generate the mechanical signals required for the regulation of cell signaling that triggers cell response: migration, proliferation and differentiation.
Acknowledgments
This work was supported by the Warsaw University of Life Sciences (grant number 505-10-072500-P00191-99), the project of National Sustainability Program I No LO1503 provided by the Ministry of Education, Youth and Sports of the Czech Republic, and PROGRES Q26 provided by Charles University, Czech Republic.
Disclosure
The authors report no conflicts of interest in this work.
References
- DeepachitraRRamnathVSastryTPGraphene oxide incorporated collagen–fibrin biofilm as a wound dressing materialRSC Adv201441076271762727
- ChenLTredgetEEWuPYGWuYParacrine factors of mesenchy-mal stem cells recruit macrophages and endothelial lineage cells and enhance wound healingPLoS One200834e188618382669
- HockingAMGibranNSMesenchymal stem cells: paracrine signaling and differentiation during cutaneous wound repairExp Cell Res2010316142213221920471978
- KyurkchievDBochevIIvanova-TodorovaESecretion of immunoregulatory cytokines by mesenchymal stem cellsWorld J Stem Cells20146555257025426252
- IsaksonMde BlacamCWhelanDMcArdleACloverAJPMesen-chymal stem cells and cutaneous wound healing: current evidence and future potentialStem Cells Int201520157112
- Marquez-CurtisLAJanowska-WieczorekAMcGannLEElliottJAMesenchymal stromal cells derived from various tissues: biological, clinical and cryopreservation aspectsCryobiology201571218119726186998
- DuscherDBarreraJWongVWStem cells in wound healing: the future of regenerative medicine? A mini-reviewGerontology201662221622526045256
- HoferHRTuanRSSecreted trophic factors of mesenchymal stem cells support neurovascular and musculoskeletal therapiesStem Cell Res Ther20167113127612948
- LeeDEAyoubNAgrawalDKMesenchymal stem cells and cutaneous wound healing: novel methods to increase cell delivery and therapeutic efficacyStem Cell Res Ther201673726960535
- MaxsonSLopezEAYooDDanilkovitch-MiagkovaALerouxMAConcise review: role of mesenchymal stem cells in wound repairStem Cells Transl Med20121214214923197761
- SalmasiSKalaskarDMYoonWWBlunnGWSeifalianAMRole of nanotopography in the development of tissue engineered 3D organs and tissues using mesenchymal stem cellsWorld J Stem Cells20157226628025815114
- HarperDThe physiology of wound healingSurgery2014329445450
- FrykbergRGBanksJChallenges in the treatment of chronic woundsAdv Wound Care (New Rochelle)20154956058226339534
- ParaniMLokhandeGSinghAGaharwarAKEngineered nano-materials for infection control and healing acute and chronic woundsACS Appl Mater Interfaces2016816100491006927043006
- MahmoudiNEslahiNMehdipourATemporary skin grafts based on hybrid graphene oxide-natural biopolymer nanofibers as effective wound healing substitutes: pre-clinical and pathological studies in animal modelsJ Mater Sci Mater Med2017287311327866344
- SinnoHPrakashSComplements and the wound healing cascade: an updated reviewPlast Surg Int20132013617
- João de MasiECCamposACJoão de MasiFDRattiMAIkeISJoão de MasiRDThe influence of growth factors on skin wound healing in ratsBraz J Otorhinolaryngol201682551252126832633
- BarrientosSBremHStojadinovicOTomic-CanicMClinical application of growth factors and cytokines in wound healingWound Repair Regen201422556957824942811
- KohTJDiPietroLAInflammation and wound healing: the role of the macrophageExpert Rev Mol Med201113e2321740602
- OlczykPMencnerŁKomosinska-VassevKThe role of the extracellular matrix components in cutaneous wound healingBio Med Res Int201474758418
- ZhaoRLiangHClarkeEJacksonCXueMInflammation in chronic woundsInt J Mol Sci201617122085
- KanjiSDasHAdvances of stem cell therapeutics in cutaneous wound healing and regenerationMediators Inflamm20172017521796729213192
- GilbertRWDVickaryousMKViloria-PetitAMSignalling by transforming growth factor beta isoforms in wound healing and tissue regenerationJ Dev Biol20164221
- GökşenSBalabanlıBCoşkun-CevherŞApplication of platelet derived growth factor-BB and diabetic wound healing: the relationship with oxidative eventsFree Radic Res201751549850528480814
- GonzalezACCostaTFAndradeZAMedradoARWound healing – a literature reviewAn Bras Dermatol201691561462027828635
- HanGCeilleyRChronic wound healing: a review of current management and treatmentsAdv Ther201734359961028108895
- CaleyMPMartinsVLCO’TooleEAMetalloproteinases and wound healingAdv Wound Care (New Rochelle)20154422523425945285
- KeJWilgusTAVascular endothelial growth factor and angiogenesis in the regulation of cutaneous wound repairAdv Wound Care (New Rochelle)201431064766125302139
- LiuLQingMWangYChenSDefects in graphene: generation, healing and their effects on the properties of graphene: a reviewJ Mater Sci Technol2015316599606
- DaiJ-FWangG-JMaLWuC-KSurface properties of graphene: relationship to graphene-polymer compositesRev Adv Mater Sci2015406071
- NovoselovKSGeimAKMorozovSVElectric field effect in atomically thin carbon filmsScience2004306569666666915499015
- StankovichSDikinDADommettGHBGraphene-based composite materialsNature2006442710028228616855586
- HernandezYNicolosiVLotyaMHigh-yield production of graphene by liquid-phase exfoliation of graphiteNat Nanotechnol20083956356818772919
- LotyaMHernandezYKingPJLiquid phase production of graphene by exfoliation of graphite in surfactant/water solutionsJ Am Chem Soc2009131103611362019227978
- MatteviCEdaGAgnoliSEvolution of electrical, chemical, and structural properties of transparent and conducting chemically derived graphene thin filmsAdv Funct Mater2009191625772583
- deSKingPJLotyaMFlexible, transparent, conducting films of randomly stacked graphene from surfactant-stabilized, oxide-free graphene dispersionsSmall20106345846419859943
- EdaGChhowallaMChemically derived graphene oxide: towards large-area thin-film electronics and optoelectronicsAdv Mater201022222392241520432408
- KaruAEBeerMPyrolytic formation of highly crystalline graphite filmsJ Appl Phys196637521792181
- EizenbergMBlakelyJMCarbon monolayer phase condensation on Ni(111)Surf Sci1979821228236
- KimKSZhaoYJangHLarge-scale pattern growth of gra-phene films for stretchable transparent electrodesNature2009457723070619145232
- ReinaAJiaXHoJLarge area, few-layer graphene films on arbitrary substrates by chemical vapor depositionNano Lett200991303519046078
- KwonSYCiobanuCVPetrovaVGrowth of semiconducting graphene on palladiumNano Lett20099123985399019995079
- MarchiniSGüntherSWintterlinJScanning tunneling microscopy of graphene on Ru(0001)Phys Rev B2007767075429
- SutterPWFlegeJ-ISutterEAEpitaxial graphene on rutheniumNat Mater20087540641118391956
- CorauxJN’DiayeATBusseCMichelyTStructural coherency of graphene on Ir(111)Nano Lett20088256557018189442
- HamiltonJCBlakelyJMCarbon segregation to single crystal surfaces of Pt, PD and COSurf Sci1980911199217
- LandTAMichelyTBehmRJHemmingerJCComsaGSTM investigation of single layer graphite structures produced on Pt(111) by hydrocarbon decompositionSurf Sci19922643261270
- SutterPSadowskiJTSutterEGraphene growth on PtPhys Rev B200980245411
- WangYZhengYXuXElectrochemical delamination of CVD-grown graphene film: toward the recyclable use of copper catalystACS Nano20115129927993322034835
- LiXZhuYCaiWTransfer of large-area graphene films for high-performance transparent conductive electrodesNano Lett20099124359436319845330
- LiangXSperlingBACalizoIToward clean and crackless transfer of grapheneACS Nano20115119144915321999646
- LinWXuLZwingenbergerSGibonEGoodmanSBLiGMesen-chymal stem cells homing to improve bone healingJ Orthop Translat20179192729662796
- CiukTPasternakIKrajewskaAProperties of chemical vapor deposition graphene transferred by high-speed electrochemical delami-nationJ Phys Chem C2013117402083320837
- EckmannAFeltenAMishchenkoAProbing the nature of defects in graphene by Raman spectroscopyNano Lett20121283925393022764888
- SkodaMDudekIJaroszASzukiewiczDGraphene: one material, many possibilities – application difficulties in biological systemsJ Nanomater201420146111
- GuptaAChenGJoshiPTadigadapaSEklundPCRaman scattering from high-frequency phonons in supported n-graphene layer filmsNano Lett20066122667267317163685
- FerrariACMeyerJCScardaciVRaman spectrum of graphene and graphene layersPhys Rev Lett20069718189401
- GrafDMolitorFEnsslinKSpatially resolved Raman spectroscopy of single- and few-layer grapheneNano Lett20077223824217297984
- DsLRiedlCKraussBvon KlitzingKStarkeUSmetJHRaman spectra of Epitaxial graphene on sic and of epitaxial graphene transferred to SiO2Nano Lett2008943204325
- ZhNWangYYYuTShenZXRaman spectroscopy and imaging of grapheneNano Res200814273
- PimentaMADresselhausGDresselhausMSCançadoLGJorioASaitoRStudying disorder in graphite-based systems by Raman spectroscopyPhys Chem Chem Phys20079111276129017347700
- WangYNiZYuTRaman studies of monolayer graphene: the substrate effectJ Phys Chem C2008112291063710640
- LasockaISzulc-DąbrowskaLSkibniewskiMBiocompatibility of pristine graphene monolayer: scaffold for fibroblastsToxicol In Vitro20184827628529409908
- SyamaSMohananPVSafety and biocompatibility of graphene: a new generation nanomaterial for biomedical applicationInt J Biol Macromol20168654655526851208
- JaworskiSHinzmannMSawoszEInteraction of different forms of graphene with chicken embryo red blood cellsEnviron Sci Pollut Res201724272167121679
- MajeedWBourdoSPetiboneDMThe role of surface chemistry in the cytotoxicity profile of grapheneJ Appl Toxicol201737446247027593524
- PelinMFuscoLLeónVDifferential cytotoxic effects of gra-phene and graphene oxide on skin keratinocytesSci Rep201774057228079192
- DuanGZhangYLuanBGraphene-induced pore formation on cell membranesSci Rep201774276728218295
- KorenevSDishmanRYebraACharacterization of graphene stripper foils in 11-MeV cyclotronsPhys Procedia201790369373
- KalbacovaMBrozAKongJKalbacMGraphene substrates promote adherence of human osteoblasts and mesenchymal stromal cellsCarbon2010481543234329
- WangBLuoPGTackettKNIIRuizONSunY-PGraphene oxides as substrate for enhanced mammalian cell growthJ Nanomater Mol Nanotechnol2012122
- GurunathanSHanJWEppakayalaVDayemAAKwonD-NKimJ-HBiocompatibility effects of biologically synthesized graphene in primary mouse embryonic fibroblast cellsNanoscale Res Lett20138111323279756
- GurunathanSWoong HanJKimEKwonD-NParkJ-KKimJ-HEnhanced green fluorescent protein-mediated synthesis of biocompatible grapheneJ Nanobiotechnology201412111624411017
- ConroyJVermaNKSmithRJBiocompatibility of pristine graphene monolayers nanosheets and thin films2014 Available from: arxiv.org/abs/1406.2497Accessed March 4, 2019
- VerdanovaMRezekBBrozANanocarbon allotropes – graphene and nanocrystalline diamond – promote cell proliferationSmall201612182499250927000766
- KimJKimSJungWHighly enhanced compatibility of human brain vascular pericyte cells on monolayer grapheneBioengineered201781859127689961
- McCallionCBurthemJRees-UnwinKGolovanovAPluenAGraphene in therapeutics delivery: problems, solutions and future opportunitiesEur J Pharm Biopharm201610423525027113141
- HuangCXuFSunYEffects of morphology, tension and vibration on wettability of graphene: a molecular dynamics studyComput Mater Sci2017139216224
- KalbacovaMHVerdanovaMBrozAVetushkaAFejfarAKalbacMModulated surface of single-layer graphene controls cell behaviorCarbon201472207214
- XuLCSiedleckiCAEffects of surface wettability and contact time on protein adhesion to biomaterial surfacesBiomaterials200728223273328317466368
- Rosales-LealJIRodríguez-ValverdeMAMazzagliaGEffect of roughness, wettability and morphology of engineered titanium surfaces on osteoblast-like cell adhesionColloids Surf A Physicochem Eng Asp20103651–3222229
- KhaliliAAAhmadMRA review of cell adhesion studies for biomedical and biological applicationsInt J Mol Sci2015168181491818426251901
- GaoCPengSFengPShuaiCBone biomaterials and interactions with stem cellsBone Res201751705929285402
- GuptaMDossBLimCTVoituriezRLadouxBSingle cell rigidity sensing: a complex relationship between focal adhesion dynamics and large-scale actin cytoskeleton remodelingCell Adh Migr201610555456727050660
- YehYCLingJYChenWCLinHHTangMJMechanotransduction of matrix stiffness in regulation of focal adhesion size and number: reciprocal regulation of caveolin-1 and β1 integrinSci Rep2017711500829118431
- JesionISkibniewskiMSkibniewskaEGraphene and carbon nanocompounds: biofunctionalization and applications in tissue engineeringBiotechnol Biotechnol Equip2015293415422
- FellerLJadwatYKhammissaRAGMeyerovRSchechterILemmerJCellular responses evoked by different surface characteristics of intraosseous titanium implantsBiomed Res Int2015201517194525767803
- JamesDKTourJMGraphene: powder, flakes, ribbons, and sheetsAcc Chem Res201346102307231823276286
- GentileFTirinatoLBattistaECells preferentially grow on rough substratesBiomaterials201031287205721220637503
- LiuNPanZFuLZhangCDaiBLiuZThe origin of wrinkles on transferred grapheneNano Res20114109961004
- WangWYangSWangAObservation of the unexpected morphology of graphene wrinkle on copper substrateSci Rep201771824428811526
- DengSBerryVWrinkled, rippled and crumpled graphene: an overview of formation mechanism, electronic properties, and applicationsMater Today2015194198213
- HuangYChenXZhangMQReversible surface wettability conversion of graphene films: optically controlled mechanismJ Mater Sci201449830253033
- Ramakrishna MatteHSSSubrahmanyamKSRaoCNRSynthetic aspects and selected properties of grapheneNanomater Nanotechnol20111513
- KalbacovaMBrozAKalbacMInfluence of the fetal bovine serum proteins on the growth of human osteoblast cells on grapheneJ Biomed Mater Res A2012100A1130013007
- TanXWThompsonBKonstantopoulosAApplication of graphene as candidate biomaterial for synthetic keratoprosthesis skirtInvest Ophthalmol Vis Sci201556116605661126465888
- SayyarSMurrayEThompsonBCGambhirSOfficerDLWallaceGGCovalently linked biocompatible graphene/polycaprolactone composites for tissue engineeringCarbon201352296304
- ProvenzanoPPKeelyPJMechanical signaling through the cytoskel-eton regulates cell proliferation by coordinated focal adhesion and Rho GTPase signalingJ Cell Sci201112481195120521444750
- HannaSEl-SibaiMSignaling networks of Rho GTPases in cell motilityCell Signal201325101955196123669310
- HoonJTanMKohC-GThe regulation of cellular responses to mechanical cues by Rho GTPasesCells201652E1727058559
- SchallerMDCellular functions of FAK kinases: insight into molecular mechanisms and novel functionsJ Cell Sci2010123Pt 71007101320332118
- KimSKimMShinYCell migration according to shape of graphene oxide micropatternsMicromachines (Basel)2016710E18630404359
- ZhangHHouRXiaoPSingle cell migration dynamics mediated by geometric confinementColloids Surf B Biointerfaces2016145727827137805
- KeshavanSOropesa-NuñezRDiasproACanaleCDanteSAdhesion and migration of CHO cells on micropatterned single layer graphene2D Materials201742025022
- LorenzoniMBrandiFDanteSSimple and effective graphene laser processing for neuron patterning applicationSci Rep20133195423739674
- GuoXMeiNAssessment of the toxic potential of graphene family nanomaterialsJ Food Drug Anal201422110511524673908
- HegabHMElmekawyAZouLMulcahyDSaintCPGinic-MarkovicMThe controversial antibacterial activity of graphene-based materialsCarbon2016105362376
- ZouXZhangLWangZLuoYMechanisms of the antimicrobial activities of graphene materialsJ Am Chem Soc201613872064207726824139
- WangLHuCShaoLThe antimicrobial activity of nanoparticles: present situation and prospects for the futureInt J Nanomedicine2017121227124928243086
- GurunathanSHanJWDayemAAEppakayalaVKimJHOxidative stress-mediated antibacterial activity of graphene oxide and reduced graphene oxide in Pseudomonas aeruginosaInt J Nanomedicine201275901591423226696
- SongCYangC-MSunX-FInfluences of graphene oxide on biofilm formation of Gram-negative and Gram-positive bacteriaEnviron Sci Pollut Res201825328532860
- LiuSZengTHHofmannMAntibacterial activity of graphite, graphite oxide, graphene oxide, and reduced graphene oxide: membrane and oxidative stressACS Nano2011596971698021851105
- NandaSSYiDKKimKStudy of antibacterial mechanism of gra-phene oxide using Raman spectroscopySci Rep20166111228442746
- ShuaiCGuoWWuPA graphene oxide-Ag co-dispersing nanosystem: dual synergistic effects on antibacterial activities and mechanical properties of polymer scaffoldsChem Eng J2018347347322333
- TuYLvMXiuPDestructive extraction of phospholipids from Escherichia coli membranes by graphene nanosheetsNat Nanotechnol20138859460123832191
- KurantowiczNSawoszEJaworskiSInteraction of graphene family materials with Listeria monocytogenes and Salmonella entericaNanoscale Res Lett201510111225977644
- BarbolinaIWoodsCRLozanoNKostarelosKNovoselovKSRobertsISPurity of graphene oxide determines its antibacterial activity2D Materials201632025025
- YeSShaoKLiZAntiviral activity of graphene oxide: how sharp edged structure and charge matterACS Appl Mater Interfaces2015738215712157926370151
- SongZWangXZhuGVirus capture and destruction by label-free graphene oxide for detection and disinfection applicationsSmall2015119–101171117625285820
- MukherjeeSSriramPBaruiAKGraphene oxides show angiogenic propertiesAdv Healthc Mater20154111722173226033847
- ParkJKimYSRyuSGraphene potentiates the myocardial repair efficacy of mesenchymal stem cells by stimulating the expression of angiogenic growth factors and gap junction proteinAdv Funct Mater2015251725902600
- ShineM-YChiangW-HChenI-WLiuW-YChenY-WSynergistic acceleration in the osteogenic and angiogenic differentiation of human mesenchymal stem cells by calcium silicate-graphene compositesMater Sci Eng C201773726735
- SinghSKSinghMKNayakMKThrombus inducing property of atomically thin graphene oxide sheetsACS Nano2011564987499621574593
- SaleemJWangLChenCImmunological effects of graphene family nanomaterialsNanoImpact20175109118
- WilgusTARoySMcDanielJCNeutrophils and wound repair: positive actions and negative reactionsAdv Wound Care (New Rochelle)20132737938824527354
- VinishMCuiWStaffordEDendritic cells modulate burn wound healing by enhancing early proliferationWound Rep and Reg2016241613
- MukherjeeSPLazzarettoBHultenbyKGraphene oxide elicits membrane lipid changes and neutrophil extracellular trap formationChem201842334358
- KaplanMJRadicMNeutrophil extracellular traps: double-edged swords of innate immunityJ Immun201218962689269522956760
- ZhouHZhaoKLiWThe interactions between pristine gra-phene and macrophages and the production of cytokines/chemokines via TLR- and NF-κB-related signaling pathwaysBiomaterials201233296933694222796167
- ChenGYYangHJLuCHSimultaneous induction of autoph-agy and toll-like receptor signaling pathways by graphene oxideBiomaterials201233276559656922704844
- MaJLiuRWangXCrucial role of lateral size for graphene oxide in activating macrophages and stimulating pro-inflammatory responses in cells and animalsACS Nano2015910104981051526389709
- OrecchioniMBedognettiDNewmanLSingle-cell mass cytometry and transcriptome profiling reveal the impact of graphene on human immune cellsNat Commun201781110929061960
- TkachAVYanamalaNStanleySGraphene oxide, but not fullerenes, targets immunoproteasomes and suppresses antigen presentation by dendritic cellsSmall201399–101686169022887961
- ZhiXFangHBaoCThe immunotoxicity of graphene oxides and the effect of PVP-coatingBiomaterials201334215254526123566800
- WangWLiZDuanJWangCFangYYangX-DIn vitro enhancement of dendritic cell-mediated anti-glioma immune response by graphene oxideNanoscale Res Lett20149131125024678
- ZhongCShiDZhengYNelsonPJBaoQSuccessive release of tissue inhibitors of metalloproteinase-1 through graphene oxide-based delivery system can promote skin regenerationNanoscale Res Lett20171253328916996
- SinghASinghASenDMesenchymal stem cells in cardiac regeneration: a detailed progress report of the last 6 years (2010–2015)Stem Cell Res Ther2016718227259550
- KenryLeeWCLohKPLimCTWhen stem cells meet graphene: opportunities and challenges in regenerative medicineBiomaterials201815523625029195230
- Al-ShaibaniMBHWangX-NLovatPEDickinsonAMCellular therapy for wounds: applications of mesenchymal stem cells in wound healingAlexandrescuVWound Healing – New Insights into Ancient ChallengesLondonInTech2016
- LiMLuanFZhaoYMesenchymal stem cell-conditioned medium accelerates wound healing with fewer scarsInt Wound J2017141647326635066
- Alcayaga-MirandaFCuencaJKhouryMAntimicrobial activity of mesenchymal stem cells: current status and new perspectives of antimicrobial peptide-based therapiesFront Immunol201782150733928424688
- LinW-HChenT-HChangJ-KA direct and polymer-free method for transferring graphene grown by chemical vapor deposition to any substrateACS Nano2014821784179124471977
- HamdanSPastarIDrakulichSNanotechnology-driven therapeutic interventions in wound healing: potential uses and applicationsACS Cent Sci20173316317528386594
- NuschkeAActivity of mesenchymal stem cells in therapies for chronic skin wound healingOrganogenesis2014101293724322872
- TamamaKKerpedjievaSSAcceleration of wound healing by multiple growth factors and cytokines secreted from multipotential stromal cells/mesenchymal stem cellsAdv Wound Care (New Rochelle)20121417718224527301
- ZhangJHuangXWangHThe challenges and promises of allogeneic mesenchymal stem cells for use as a cell-based therapyStem Cell Res Ther20156123426620426
- WalterMNMWrightKTFullerHRMacneilSJohnsonWEBMesenchymal stem cell-conditioned medium accelerates skin wound healing: an in vitro study of fibroblast and keratinocyte scratch assaysExp Cell Res201031671271128120206158
- EmingSAMartinPTomic-CanicMWound repair and regeneration: mechanisms, signaling, and translationSci Transl Med20146265265sr625473038
- SteffensDMathorMBSantiBTLucoDPPrankePDevelopment of a biomaterial associated with mesenchymal stem cells and kerati-nocytes for use as a skin substituteRegen Med201510897598726542841
- AnYLiuWJXuePAutophagy promotes MSC-mediated vascularization in cutaneous wound healing via regulation of VEGF secretionCell Death Dis20189211429298988
- Latifi-PupovciHKuçiZWehnerSIn vitro migration and proliferation (“wound healing”) potential of mesenchymal stromal cells generated from human CD271+ bone marrow mononuclear cellsJ Transl Med201513131526407865
- KazantsevaJIvanovRGasikMNeumanTHussainovaIGraphene-augmented nanofiber scaffolds demonstrate new features in cells behaviourSci Rep2016613015027443974
- JastrzebskaETomeckaEJesionIHeart-on-a-chip based on stem cell biologyBiosens Bioelectron201675678126298640
- BrzozkaZJastrzebskaECardiac Cell Culture TechnologiesChamSpringer International Publishing2018
- EnnisWJSuiABartholomewAStem cells and healing: impact on inflammationAdv Wound Care (New Rochelle)20132736937824587974
- ChenDHaoHFuXHanWInsight into reepithelialization: how do mesenchymal stem cells perform?Stem Cells Int20162016612017326770209
- SeoBFJungS-NThe immunomodulatory effects of mesenchymal stem cells in prevention or treatment of excessive scarsStem Cells Int20162016218
- YewTLHungYTLiHYEnhancement of wound healing by human multipotent stromal cell conditioned medium: the paracrine factors and p38 MAPK activationCell Transplant201120569370621176394
- BortolottiFUkovichLRazbanVIn vivo therapeutic potential of mesenchymal stromal cells depends on the source and the isolation procedureStem Cell Reports20154333233925660405
- WuYChenLScottPGTredgetEEMesenchymal stem cells enhance wound healing through differentiation and angiogenesisStem Cells200725102648265917615264
- ShinS-CSeoYParkHYRegenerative potential of tonsil mesenchymal stem cells on surgical cutaneous defectCell Death Dis20189211229298988
- LuoGChengWHeWPromotion of cutaneous wound healing by local application of mesenchymal stem cells derived from human umbilical cord bloodWound Repair Regen201018550651320840520
- FinnsonKWMcLeanSDi GuglielmoGMPhilipADynamics of transforming growth factor beta signaling in wound healing and scarringAdv Wound Care (New Rochelle)20132519521424527343
- RennertRCSorkinMGargRKGurtnerGCStem cell recruitment after injury: lessons for regenerative medicineRegen Med20127683385023164083
- SpeesJLLeeRHGregoryCAMechanisms of mesenchymal stem/ stromal cell functionStem Cell Res Ther20167112527581859
- LeeEYXiaYKimW-SHypoxia-enhanced wound-healing function of adipose-derived stem cells: increase in stem cell proliferation and up-regulation of VEGF and bFGFWound Repair Regen200917454054719614919
- Rodriguez-MenocalLSalgadoMFordDVan BadiavasEStimulation of skin and wound fibroblast migration by mesenchymal stem cells derived from normal donors and chronic wound patientsStem Cells Transl Med20121322122923197781
- MishraPJMishraPJBanerjeeDKeratinocyte induced differentiation of mesenchymal stem cells into dermal myofibroblasts: a role in effective wound healingInt J Transl Sci20161532
- SasakiMAbeRFujitaYAndoSInokumaDShimizuHMesenchymal stem cells are recruited into wounded skin and contribute to wound repair by transdifferentiation into multiple skin cell typeJ Immunol200818042581258718250469
- EstradaJCTorresYBengurãaAHuman mesenchymal stem cell-replicative senescence and oxidative stress are closely linked to aneuploidyCell Death Dis201346e69123807220
- TurinettoVVitaleEGiachinoCSenescence in human mesenchymal stem cells: functional changes and implications in stem cell-based therapyInt J Mol Sci20161771164
- LiYWuQWangYLiLBuHBaoJSenescence of mesenchymal stem cells (review)Int J Mol Med201739477578228290609
- WagnerWHornPCastoldiMReplicative senescence of mesenchymal stem cells: a continuous and organized processPLoS One200835e221318493317
- WhitfieldMJLeeWCJVan VlietKJOnset of heterogeneity in culture-expanded bone marrow stromal cellsStem Cell Res20131131365137724103495
- KundrotasGGasperskajaESlapsyteGIdentity, proliferation capacity, genomic stability and novel senescence markers of mesen-chymal stem cells isolated from low volume of human bone marrowOncotarget2016710107881080226910916
- LiZWangHYangBSunYHuoRThree-dimensional graphene foams loaded with bone marrow derived mesenchymal stem cells promote skin wound healing with reduced scarringMater Sci Eng C Mater Biol Appl20155718118826354253
- ChuJShiPYanWPEGylated graphene oxide-mediated quercetin-modified collagen hybrid scaffold for enhancement of MSCs differentiation potential and diabetic wound healingNanoscale201810209547956029745944