Abstract
We previously found that downregulation of protein kinase Cα (PKCα) can inhibit retinal pigment epithelium (RPE) cell proliferation involved in the development of proliferative vitreoretinopathy (PVR). In this study, we tested whether PKCα could be downregulated via small interfering RNA (siRNA)-PKCα released from foldable capsular vitreous body (FCVB) in cultured human RPE cells. SiRNA-PKCα content, determined by ultraviolet (UV) spectrophotometer, was released from FCVB containing 200, 300, 400, 500, and 600 nm siRNA-PKCα in a time-dependent manner from 1 to 96 hours and a dose-dependent manner at five concentrations. The content (y) had a good linear relationship with time (x), especially in the 600 nm siRNA-PKCα group (y = 16.214x, R2 = 0.9809). After treatment with siRNA-PKCα released from FCVBs, the PKCα was significantly decreased by RT-PCR, Western blot, and immunofluorescence analysis in RPE cells. These results indicate that PKCα was significantly downregulated by siRNA-PKCα released from FCVB in human RPE cells and provide us with a new avenue to prevent PVR.
Introduction
Proliferative vitreoretinopathy (PVR) is a severe complication of retinal detachment and the most common reason for the failure of retinal detachment surgery.Citation1,Citation2 The risk of PVR varies from 10% to 40% depending on the original diseases.Citation3,Citation4 The pathology of PVR involves a complex network of relationships of cellular and humoral factors, in which proliferating retinal pigment epithelium (RPE) cells are thought to be an initiating factor.Citation5–Citation8 Protein kinase C (PKC) is an intracellular enzyme family, which transduces extracellular signals into intracellular responses. It has been well documented that the PKC family is involved in a series of life phenomena such as cell communication, secretion, ion channel regulation, cell proliferation and differentiation, and cancer, implying it is an ideal target for blocking signal transduction.Citation9–Citation12 Our previous study found that, of the ten kinds of PKC isoforms expressed in RPE cells, only PKCα is downstream of the confluence of the complex PVR network, which can promote RPE cell proliferation by downregulation of p27 Kip1.Citation13,Citation14 All of these studies strongly suggest that PKCα is an ideal and special blocking target for PVR.
Drug delivery to the posterior segments of the eye presents considerable challenges, because drugs administered by the conventional routes of topical or systemic dosing are usually ineffective due to the existence of critical barriers (such as corneal, blood–retinal, and blood–aqueous barriers) and short drug half-lives.Citation15,Citation16 Considering these challenges, development of a drug delivery system (DDS) is becoming increasingly important in the treatment of vitreoretinal diseases not only to facilitate drug efficacy but also to attenuate adverse effects.Citation17
In our previous studies, we discovered that foldable capsular vitreous body (FCVB) is a public release vehicle DDS and can sustainably and mechanically release dexamethasone sodium phosphate (DexP) via the capsule apertures.Citation18 In the clinical trial (ClinicalTrials.gov number, NCT00910702) evaluating the flexibility, efficacy, and safety of an FCVB in the treatment of severe retinal detachment, the FCVB was shown to be flexible, effective, and safe as a vitreous substitute over a 3-month implantation time. The FCVB used in the clinical trial consists of a thin vitreous-shaped capsule with a tube-valve system made using computer and industrial technology.Citation19–Citation21
RNA interference (RNAi) is an ancient and common biological phenomenon discovered in vivo in recent years.Citation22 It is commonly used as a powerful tool in biological and biomedical research and has been used experimentally to prevent ocular neovascularization and inflammation.Citation23–Citation25 In our previous research, small interfering RNA (siRNA)-PKCα), a specific inhibitor of PKCα, was shown to be able to inhibit RPE cell proliferation.Citation14 Therefore, in this study, we investigate whether PKCα could be downregulated via siRNA-PKCα released from FCVB in cultured human RPE cells, providing us with a new avenue to prevent PVR.
Materials and methods
Characteristics of siRNA-PKC#α
SiRNA-PKCα was purchased from Ribuo Biotech (Guangzhou, China). The sequence of siRNA-PKCα was: positive-sense strand 5′GGCGUCCUGUUGUAUGAAAdAdT3′, antisense strand 3′dTdACCGCAGGACAACAUACUUU5′. Chemically synthesized siRNA-PKCα was mixed the suspension of two deoxyribose nucleotides to increase the stability in its 3′ end. Lyophilized powder of siRNA-PKCα was dissolved in TE buffer (110 mM Tris·HCl, 1 mM EDTA, pH = 8.00) into the stock concentration of 20 μM, and stored at −20°C.
Basic material and fabrication of the FCVB
FCVB is made of tailor-made modified liquid silicone rubber, which consists of a vitreous-like capsule and a tube-valve system (shown in ). The basic material, Dow Corning Class VI elastomer, was purchased from Dow Corning Company (Midland, MI). The details of FCVB were described in our previous study.Citation19
Figure 1 Small interfering RNA–protein kinase Cα (SiRNA-PKCα) sustained-release studies in vitro. (A) Foldable capsular vitreous body (FCVB) consists of a thin (60 μm) vitreous-like capsule mimicked by computer with a tube-valve system. (B) SiRNA-PKCα at concentrations of 200, 300, 400, 500, and 600 nM in TE buffer was injected into the capsules of FCVB, which were immersed in cups of modified Frans diffusion cells. Two hundred (200) μL liquid was aspirated at time intervals of 1, 2, 3, 6, 9, 12, 24, 48, 72, and 96 hours.

In vitro drug sustained-release studies
In a typical experiment, siRNA-PKCα at concentrations of 200, 300, 400, 500, and 600 nm in TE buffer was injected into the capsules of the FCVBs, and the FCVBs were separately immersed in five cups of modified Franz diffusion cells (shown in ) and 4 mL of TE buffer as release medium was transferred into each cup. These cups were kept in an orbital shaker maintained at 37°C with constant agitation. Two hundred microliters (200 μL) of the release medium was aspirated for measurement at 1, 2, 3, 6, 9, 12, 24, 48, 72, and 96 hours, and then the same amount of fresh medium was added into the cups. The collected samples were stored in a refrigerator. The siRNA-PKCα content in the liquid was then detected by UV spectrophotometer (DU800; Beckman Coulter, Brea, CA).
SiRNA-PKCα sample assay in vitro
First, a standard curve was made to demonstrate the effectiveness of the assay method. In brief, the 20 μM siRNA-PKCα stock solution was serially diluted with TE buffer to the desired concentrations: 20, 40, 60, 80, 100, 200, 300, 400, 500, and 600 nm. Samples of 100 μL were transferred into the cuvette chamber. Samples were assayed at the absorbance of 260 nm, and 100 μL of TE buffer was added to blank the background. Calibrations were made using Excel software (2003; Microsoft, Redmond, WA).
Micromorphology of the FCVB
The capsule of the FCVB was cut into a 1-cm diameter disc, and then the sample was cleaned, coated with gold, and fixed on a specimen stub. The image of the specimen surface was captured with a scanning electron microscope (XL 30 ESEM FEG; FEI-Philips, Eindhoven, the Netherlands). Examinations were carried out at 10 kV and in a humid atmosphere of approximately 1.8–2.0 Torr. Individual parameters are visible in the scale bar of each image.
Human RPE cell culture
Human RPE cells were isolated within 24 hours of death from donor eyes, which were obtained from the Eye Bank of the Zhongshan Ophthalmic Center as previously described.Citation26 The project was approved by the Ethics Committee of the Zhongshan Ophthalmic Center and followed the tenets of the Declaration of Helsinki. RPE cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen-Gibco, Karlsruhe, Germany) containing 10% fetal bovine serum, penicillin G (100 μg/mL), streptomycin sulfate (100 mg/mL), and L-glutamate in Corning culture plates. The cells were maintained at 37° C in 5% CO2 in a humidified atmosphere, and those between the third and sixth passages were used for all experiments.
Treatment of human RPE cells with siRNA-PKCα released from FCVB
Human RPE cells grown in transwells (Corning Inc, Corning, NY) were incubated for 12 hours. The culture media was then replaced with DMEM without serum or antibiotics, and filter inserts (8 μm pore size) carrying FCVBs containing 400, 500, and 600 nm siRNA-PKCα were transferred to transwells. Lipofectamine 2000 (5 μL) (Invitrogen, Karlsruhe, Germany) was added to each well at time points of 48, 24, and 12 hours for each group, and the medium in each well was replaced with DMEM with 10% fetal bovine serum 6 hours later. Eighteen hours later, cells were harvested for Western blot and reverse transcription-polymerase chain reaction (RT-PCR). For immunofluorescence analysis, cells which were grown on cover lips in transwells were subjected to the same treatments as Western blot and RT-PCR. Cells treated by DMEM with 10% fetal bovine serum and transfected with 100 nm siRNA for 24 hours were used for blank control and positive control, respectively.
RT-PCR
Total RNA was extracted using Trizol reagent (Life Technologies, Gaithersburg, MD). The integrity of the RNA was checked by 2% agarose gel electrophoresis. Approximately 2 μg RNA was reverse-transcribed following the protocol of the Super Script™ first-strand synthesis system. CDNAs encoding PKCα and GAPDH genes were amplified by PCR as follows: denaturation at 94°C for 30 seconds, annealing at 63°C for 30 seconds and elongation at 72°C for 45 seconds. Primer sequence was designed using Primer 3. For PKCα, the forward primer was 5′-ATCCGCAGTGGAATGAGTC CTTTACAT-3′, and the reverse primer was 5′-TTGGAAG GTTGTTTCCTGTCTTCAGAG-3′. A housekeeping gene, glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the internal control. The forward primer was 5′-ACCCAGAAGACTGTGGATGG-3′, and the reverse primer was 5′-TGCTGTAGCCAAATTCGTTG-3′. PCR products were analyzed by agarose (2%) gel electrophoresis. Quantifications of signal intensity were confirmed using a specific computer program (Image J 1.43U software, Wayne Rasband, National Institutes of Health, USA).
Preparation of cell extracts
The medium was removed and the plates were washed twice with ice-cold phosphate buffered saline (PBS). The human RPE cells were lysed with sample buffer that contained 60 mM Tris, pH 6.8, 2% (w/v) sodium dodecyl sulfate (SDS), 100 mM 2-mercaptoethanol, and 0.01% (w/v) bromophenol blue.Citation27 The lysate was then incubated on ice for 30 minutes. The lysate was scraped using a cell scraper and harvested using a pipettor, then centrifuged at 4°C for 30 minutes. The supernatant was collected and boiled for 5 minutes and stored at −20°C.
Western blot analysis
Cellular extracts from human RPE cells were processed for Western blot analysis.Citation28 Briefly, 60 μg of protein per well was loaded on a 10% SDS-polyacrylamide gel electrophoresis (PAGE) gel. The protein was electro-transferred to polyvinylidene difluoride membranes (Millipore, Billerica, MA) for 1 hour at 350 mA, then blocked with Tris-buffered saline (TBS) containing 5% nonfat milk and 0.1% Tween-20 (TBST) for 1 hour and incubated with mouse anti-PKCα (BD Systems, Torrance, CA) overnight. After three washes with TBST, the membranes were incubated with horseradish peroxidase conjugated secondary antibody for 1 hour at room temperature and washed with TBST. Localization of the antibody was detected by chemiluminescence using the ECL kit (Cell Signaling, Danvers, MA) following the manufacturer’s instructions. β-actin (Boster Biologic Technology Ltd, Wuhan, China) was used as an internal control.
Immunofluorescence analysis
After treatment, cells were fixed for 15 minutes in 4% paraformaldehyde, and then rinsed three times in PBS. All reagent incubations were performed in a humidified chamber. The mouse anti-PKCα was incubated for 16 hours in PBS at room temperature; horse serum was used as the negative control instead of the primary antibody. After washing four times for 10 minutes in PBS, fluorescein isothiocyanate-labeled secondary antibodies were incubated for 40 minutes at 37°C. Then Hoechst 33342 was incubated for 10 minutes at room temperature. After three rinses in PBS, cover slips were mounted onto glass slides and analyzed on a Zeiss laser scanning confocal microscope (LSCM 510 META; Oberkochen, Germany).
Statistical analysis
All data were expressed as mean ± standard deviation (SD). All experiments were repeated at least 4 times, and similar results were obtained. One-way analysis of variance (ANOVA), the test of homogeneity of variances, and a post hoc test (Bonferroni test) were used to determine significant differences between the treated groups and the control groups. P < 0.05 was considered statistically significant.
Results
Selectivity and acceptability of the standard curve
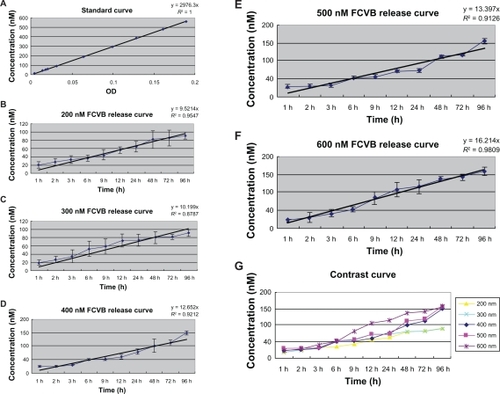
A representative result of the standard curve is shown in . The standard curve showed a very good linear relationship (y = 2976.3x, R2 = 1), demonstrating that the method selected had good selectivity and acceptability.
Figure 2 Standard curve and in vitro release profile in the cumulative amount of five different concentrations. (A) The standard curve had a very good linear relationship. With the absorbance (x) increase, the small interfering RNA–protein kinase Cα (SiRNA-PKCα) concentration (y) rose accordingly (y = 2976.3x, R2 = 1). B–G) Time and dose-dependent release. The siRNA-PKCα released from the foldable capsular vitreous body (FCVB) increased in a linear manner with time. Of the five different concentrations, the 600 nM group had a best linear relationship (y = 16.214x, R2 = 0.9809). When siRNA-PKCα concentration rose inside the FCVBs, the content released from the capsules rose accordingly. The tendency began from 1 hour, and lasted until 96 hours.
Notes: Results shown were representative of four repeated experiments. The concentrations of different time points are statistically different (P < 0.05). The data are expressed as the mean ± standard deviation (SD).
In vitro release of siRNA-PKC#α
The cumulative release of siRNA-PKCα from FCVB is shown in –. SiRNA-PKCα released from the capsules of FCVB increased with time in a 96-hour experimental time period. The content (y) had a good linear relationship with time (x), especially in the 600 nm group, indicating that the FCVB released siRNA-PKCα stably with time (y = 16.214x, R2 = 0.9809) (P < 0.05).
The contents released from FCVB increased accordingly, with siRNA-PKCα concentration rising in the FCVB. The released content was much greater in the 600 nm group at the same release time than in the other groups. All data indicated that siRNA-PKCα was released from the capsules of FCVB in a dose-dependent manner in vitro in 5 concentrations during a 96-hour experiment time period.
Scanning electron microscopy image of the FCVB
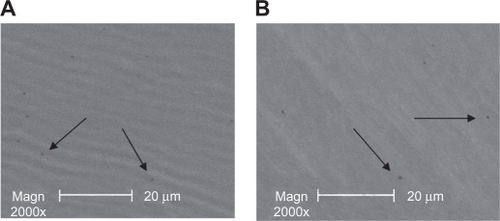
Before the experiment, scattered mini apertures of 300 nm could be seen in the capsule of FCVB by scanning electron microscopy. shows 2 of the typical apertures. After a 96-hour experimental time period, 300-nm mini apertures still existed in the capsules, as shown in . No great changes were observed in the capsule apertures by scanning electron microscopy.
Figure 3 Scanning electron microscope images of the capsule of the foldable capsular vitreous body. Mini apertures of 300 nm in the capsules were observed before the release (A) and at the end of the experiment time (B) (arrows).
PKCα downregulation after treatment by siRNA-PKCα released from FCVB
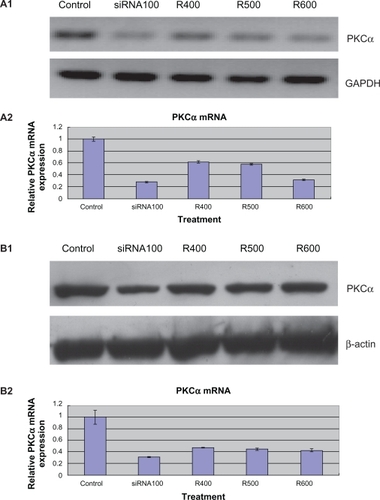
As shown in , RT-PCR results showed that PKCα messenger RNA (mRNA) was significantly downregulated following 24-hour release siRNA-PKCα treatments in the 400 nm, 500 nm, and 600 nm release groups. Consistent with a change at the mRNA level, the PKCα protein after treatment decreased 45.1% ± 6.73%, 50.8% ± 4.96%, and 54.3% ± 3.62% in the 400 nm, 500 nm, and 600 nm release groups, respectively, and 64.0% ± 4.5% in the positive control of 100 nm siRNA-PKCα treated group compared with that in the untreated control group (). In the untreated RPE cells, both the mRNA and protein level remained constant. These data indicate that siRNA-PKCα can actually be released from the FCVBs and further decrease expression of PKCα in cultured human RPE cells.
Figure 4 Expression of protein kinase Cα (PKCα) messenger RNA (mRNA) and protein downregulation by released small interfering RNA (siRNA)-PKCα in retinal pigment epithelium (RPE) cells. A1, A2) RT-PCR showed that released siRNA-PKCα reduced the mRNA levels of PKCα significantly compared with control groups. *P < 0.05 vs the corresponding value for control groups. Data are the mean ± standard deviation of quadruplicates from an experiment that was repeated with similar results (siRNA100 means RPE cells treated with 100 nM siRNA-PKCα; R400, R500, and R600 mean RPE cells treated with siRNA-PKCα released from foldable capsular vitreous bodies containing 400, 500, and 600 nM siRNA-PKCα). B1, B2) Western blot showed that released siRNA-PKCα reduced the protein levels of PKCα significantly compared with control groups.
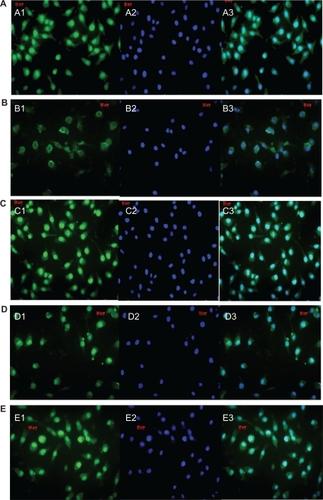
We further examined the expression changes of PKCα using immunocytochemistry. The cultured human RPE cells formed a monolayer with typical polygonal cellular arrays, as shown in . Compatible with findings from RT-PCR and Western blot analysis, staining of PKCα decreased significantly after exposure to both 100 nm siRNA-PKCα and siRNA-PKCα released from FCVBs. Compared with the positive control of 100 nm siRNA-PKCα treated group, the stain did not change greatly in the experimental groups.
Figure 5 Confocal images of protein kinase Cα (PKCα) expression in retinal pigment epithelium (RPE) cells by different treatments. PKCα staining was much lighter in the small interfering RNA (siRNA)-PKCα treatment than in the control, with no significant difference in the released siRNA-PKCα treatment groups. (A) blank control. (B) RPE cells treated with 100 nM siRNA-PKCα. (C–E) RPE cells treated with siRNA-PKCα released from foldable capsular vitreous bodies containing 400, 500, and 600 nM siRNA-PKCα, respectively. (1) PKCα staining in the cytoplasm and nuclei of RPE cells. (2) Staining of the nuclei with Hoechst 33342. (3) An overlay of images 1 and 2. Scale bars, 20 μm. Magnification × 1000.
Discussion
In this study, we determined the siRNA-PKCα release of a nonbiodegradable device (FCVB) and its effect on PKCα in cultured human RPE cells. Our present study indicates that FCVB can mechanically and sustainably release siRNA-PKCα in a time-dependent and dose-dependent manner during a 96-hour experimental time period. Furthermore, PKCα was decreased in RPE cells treated with siRNA-PKCα released from FCVBs. However, compared with the positive control group, expression of the target protein did not decrease remarkably with our experimental systems even in the presence of transfection reagents. This is likely attributable to the fact that our FCVB drug delivery system released siRNA-PKCα progressively and to the fact that siRNA-PKCα of the positive control group reached peak concentration quickly. The present results were consistent with a previous study that FCVB can sustainably, mechanically release DexP via the apertures of the capsule in a time-dependent and dose-dependent manner.Citation18
In this study, siRNA-PKCα was shown to be fairly permeable through the capsule apertures of FCVB and to downregulate PKCα in RPE cells. We speculated that there were two main factors involved in the mechanism. The first is the concentration gradient of siRNA-PKCα. With the concentration gradient increase, siRNA-PKCα released from the FCVBs subsequently increased, as shown in –. Second, the diffusion coefficient was related to the concentration gradient. The two factors mentioned previously have been reported by Acharya and colleagues in a homogeneous poly(lactic-co-glycolic acid) microstructures drug release system.Citation29
The current intraocular DDS confronts some major disadvantages. For example, they are opaque, may interfere with vision, and result in a nonuniform drug distribution.Citation30 In contrast to the current intraocular DDS, FCVB has good transparency and induces very little refractive shift,Citation31 permitting the siRNA-PKCα in solution to make direct contact with the retina and achieve targeted drug delivery. Therefore, the FCVB can sustainably and mechanically release drugs without changing their chemical properties, and may provide a common vehicle for releasing different drugs.
The ideal intravitreal DDS is a kind of material that is not only an intravitreal drug delivery system but also a vitreous tamponade agent.Citation30 Previous studies demonstrated that FCVB was a fine vitreous substitute that closely mimicked vitreous morphology and restored its physiological functions, such as support and cellular barriers, during a 3-month implantation period without the obvious complications commonly induced by silicone oil.Citation32 In our study, siRNA-PKCα was released from FCVB in a time-dependent and a dose-dependent manner in vitro. SiRNA-PKCα was detected until 96 hours, indicating that FCVB can be used as a DDS in addition to serving as a vitreous substitute.
Instability is one of the essential obstacles of the RNAi therapeutic applications.Citation33 SiRNA is easily degraded by nucleases in vitro and in vivo. Andrieu-Soler and his colleagues have demonstrated that there was high nuclease activity in the vitreous.Citation34 To solve the problem, siRNA-PKCα combined with FCVB shows great advantages. On the one hand, chemically synthesized siRNA-PKCα was mixed with the suspension of 2 deoxyribose nucleotides to increase the stability in its 3′ end; on the other hand, siRNA-PKCα was injected into the capsule of FCVB, protecting it from contacting nucleases in vitro and in vivo. In addition, Ambati J and colleagues reported that 21-nt or longer siRNAs suppress hemanogiogenesis and lymphangiogenesis in various mouse models of neovascularization in a sequence- and target-independent manner through activating cell surface Toll-like receptor 3.Citation35,Citation36 Therefore, it is critical to address the safety issue using in vivo systems.
The underlying factor in many vision-threatening vitreo-retinal diseases is unwanted intraocular cell proliferation.Citation37 In the pathology of PVR, proliferating RPE cells are thought to be an initiating factor.Citation5–Citation8 In a previous study, we found that PKCα affects the cell cycle progression and proliferation in RPE cells through the downregulation of p27kip1.Citation14 Since PKCα was downregulated after exposure to siRNA-PKCα released from FCVB, it may be a rational approach for targeted therapies against RPE cell proliferation and PVR disease.
Intravitreal sustained-release systems are exciting new modalities of drug delivery that offer effective treatment of some visually devastating diseases.Citation38 SiRNA-PKCα combined with FCVB shows great prospects for preventing PVR, and it will greatly reduce the suffering and economic burden of patients. As a public release vehicle, FCVB has the potential to release several drugs at the same time, which will meet the requirements of drug combinations for many diseases.
In conclusion, FCVB can sustainably and mechanically release siRNA-PKCα via capsule apertures in a time-dependent and dose-dependent manner in addition to serving as a vitreous substitute. The in vitro study demonstrated that PKCα was inhibited after exposure to siRNA-PKCα released from FCVBs. The present research offers a new avenue for the treatment of diseases of the posterior segment of eyes, especially proliferative eye diseases.
Conclusion
In this study, we developed a FCVB drug delivery system to release siRNA-PKCα to inhibit PKCα expression in RPE cells in vitro. We found that PKCα was significantly downregulated by siRNA-PKCα released from FCVB, providing a new avenue to prevent PVR.
Acknowledgements
This study was supported by Natural National Science Foundation of China (NSFC-30973258) and the National Basic Research Program of China (“973” program, Number 2007CB512200). The Eye Bank of Zhongshan Ophthalmic Center is thanked for providing eye cups for primary cell culture.
Disclosure
The authors report no conflicts of interest in this work.
References
- SunalpMAWiedemannPSorgenteNRyanSJEffect of adriamycin on experimental proliferative vitreoretinopathy in the rabbitExp Eye Res19854111051154029281
- CowleyMConwayBPCampochiaroPAKaiserDGaskinHClinical risk factors for proliferative vitreoretinopathyArch Ophthalmol19891078114711512757544
- KirchhofBStrategies to influence PVR developmentGraefes Arch Clin Exp Ophthalmol2004242869970315309556
- CardilloJAStoutJTLaBreeLPost-traumatic proliferative vitreoretinopathy. The epidemiologic profile, onset, risk factors, and visual outcomeOphthalmology19971047116611739224471
- MurphyTLSakamotoTHintonDRMigration of retinal pigment epithelium cells in vitro is regulated by protein kinase CExp Eye Res19956066836957641851
- HarrisMSSakamotoTKimuraHHypericin inhibits cell growth and induces apoptosis in retinal pigment epithelial cells: possible involvement of protein kinase CCurr Eye Res19961532552628654105
- SheuSJSakamotoTOsuskyRTransforming growth factor-beta regulates human retinal pigment epithelial cell phagocytosis by influencing a protein kinase C-dependent pathwayGraefes Arch Clin Exp Ophthalmol1994232116957017531168
- SakamotoTHintonDRSakamotoHCollagen gel contraction induced by retinal pigment epithelial cells and choroidal fibroblasts involves the protein kinase C pathwayCurr Eye Res19941364514597924409
- NishizukaYThe molecular heterogeneity of protein kinase C and its implications for cellular regulationNature198833461846616653045562
- MoriartyPDicksonAJErichsenJTBoultonMProtein kinase C isoenzyme expression in retinal cellsOphthalmic Res2000322–3576010754435
- EvansVGMultiple pathways to apoptosisCell Biol Int19931754614768339064
- KizakiHSuzukiKTadakumaTIshimuraYAdenosine receptor-mediated accumulation of cyclic AMP-induced T-lymphocyte death through internucleosomal DNA cleavageJ Biol Chem19902659528052841690738
- YuKMaPGeJExpression of protein kinase C isoforms in cultured human retinal pigment epithelial cellsGraefes Arch Clin Exp Ophthalmol2007245799399917124607
- GaoQTanJMaPPKC alpha affects cell cycle progression and proliferation in human RPE cells through the downregulation of p27kip1Mol Vis2009152683269520011080
- HughesPMOlejnikOChang-LinJEWilsonCGTopical and systemic drug delivery to the posterior segmentsAdv Drug Deliv Rev200557142010203216289435
- Cunha-VazJGThe blood-ocular barriers: past, present, and futureDoc Ophthalmol1997931–21491579476613
- YasukawaTOguraYTabataYKimuraHWiedemannPHondaYDrug delivery systems for vitreoretinal diseasesProg Retin Eye Res200423325328115177203
- LiuYKeQChenJSustained mechanical release of dexamethasone sodium phosphate from a foldable capsular vitreous bodyInvest Ophthalmol Vis Sci20105131636164219834025
- LiuYJiangZGaoQTechnical standards of a foldable capsular vitreous body in terms of mechanical, optical, and biocompatible propertiesArtif Organs2010341083684520618225
- LinXGeJGaoQEvaluation of the flexibility, efficacy, and safety of a foldable capsular vitreous body in the treatment of severe retinal detachmentInvest Ophthalmol Vis Sci201152137438120811065
- GaoQMouSGeJA new strategy to replace the natural vitreous by a novel capsular artificial vitreous body with pressure-control valveEye (Lond)200822346146817525767
- BernsteinECaudyAAHammondSMHannonGJRole for a bidentate ribonuclease in the initiation step of RNA interferenceNature2001409681836336611201747
- NovinaCDSharpPAThe RNAi revolutionNature2004430699616116415241403
- ReichSJFosnotJKurokiASmall interfering RNA (siRNA) targeting VEGF effectively inhibits ocular neovascularization in a mouse modelMol Vis2003921021612789138
- NakamuraHSiddiquiSSShenXRNA interference targeting transforming growth factor-beta type II receptor suppresses ocular inflammation and fibrosisMol Vis20041070371115475878
- CinatlJJrMichaelisMFleckensteinCWest Nile virus infection induces interferon signaling in human retinal pigment epithelial cellsInvest Ophthalmol Vis Sci200647264565116431963
- CampaMJWangMZHowardBFitzgeraldMCPatzEFJrProtein expression profiling identifies macrophage migration inhibitory factor and cyclophilin a as potential molecular targets in non-small cell lung cancerCancer Res20036371652165612670919
- WeaverSASchaeferALDixonWTWestern blotting for detection of glucocorticoid receptors in the brain and pituitary gland from adrenal intact pigsBrain Res20008691–213013610865067
- AcharyaGShinCSVedanthamKA study of drug release from homogeneous PLGA microstructuresJ Control Release2010146220120620381555
- ColthurstMJWilliamsRLHiscottPSGriersonIBiomaterials used in the posterior segment of the eyeBiomaterials200021764966510711963
- GaoQChenXGeJRefractive shifts in four selected artificial vitreous substitutes based on Gullstrand-Emsley and Liou-Brennan schematic eyesInvest Ophthalmol Vis Sci20095073529353419264881
- ChenJGaoQLiuYClinical device-related article evaluation of morphology and functions of a foldable capsular vitreous body in the rabbit eyeJ Biomed Mater Res B Appl Biomater201197239640421442743
- FattalEBarrattGNanotechnologies and controlled release systems for the delivery of antisense oligonucleotides and small interfering RNABr J Pharmacol2009157217919419366348
- Andrieu-SolerCBejjaniRAde BizemontTNormandNBenEzraDBehar-CohenFOcular gene therapy: a review of nonviral strategiesMol Vis2006121334134717110916
- KleinmanMEYamadaKTakedaASequence- and target-independent angiogenesis suppression by siRNA via TLR3Nature2008452718759159718368052
- ChoWGAlbuquerqueRJKleinmanMESmall interfering RNA-induced TLR3 activation inhibits blood and lymphatic vessel growthProc Natl Acad Sci U S A2009106177137714219359485
- ChengLHostetlerKValiaevaNIntravitreal crystalline drug delivery for intraocular proliferation diseasesInvest Ophthalmol Vis Sci201051147448119696179
- GuidettiBAzémaJMalet-MartinoMMartinoRDelivery systems for the treatment of proliferative vitreoretinopathy: materials, devices and colloidal carriersCurr Drug Deliv20085171918220546