
Abstract
Omega-3 polyunsaturated fatty acids (ω-3 PUFAs) are dietary factors involved in the prevention of cardiovascular, inflammatory, and neoplastic diseases. A multidisciplinary approach – based on recent findings in nutritional science, lipid biochemistry, biotechnology, and biology of inflammation and cancer – has been recently employed to develop ω-3 PUFA-containing nanoformulations with an aim to protect these fatty acids from degradation, increase their bioavailability and delivery to target tissues, and, thus, enhance their bioactivity. In some cases, these nanoformulations were designed to administer ω-3 PUFAs in combination with other nutraceuticals or conventional/innovative drugs. The aim of this strategy was to increase the activities of the compounds contained in the nanoformulation and to reduce the adverse effects often induced by drugs. We herein analyze the results of papers evaluating the potential use of ω-3 PUFA-containing nanomaterials in fighting cardiovascular diseases and cancer. Future directions in this field of research are also provided.
Introduction
There is a large body of evidence supporting a beneficial role of omega-3 polyunsaturated fatty acids (ω-3 PUFAs) against several pathologies, including cardiovascular diseases (CVDs) and cancer.Citation1–Citation3 The results have been obtained by using either the essential fatty acid α-linolenic acid (ALA, 18:3 ω-3), mostly found in vegetables and nuts or, and particularly, its metabolic products, the long-chain (LC)-ω-3 eicosapentaenoic acid (EPA, 20:5 ω-3) and docosahexaenoic acid (DHA, 22:6 ω-3). However, in mammal cells these two highly bioactive compounds are produced endogenously from ALA at very low levels; therefore, it is necessary to increase their main dietary sources (fish and seafood) to reach sufficient amounts in tissues. However, this requires a frequent intake of fish and seafood that appears unsustainable, particularly in the future.Citation4 Moreover, wild fish is often contaminated with heavy metals or pesticides,Citation5 whereas farmed fish contains much lower levels of LC-ω-3 PUFAs and high levels of antibiotics.Citation6 In order to overcome these problems, alternative LC-ω-3 PUFA sources are now being explored, such as microalgae grown in controlled environments,Citation7 or genetically modified plants and marine protists.Citation8–Citation11 An alternative approach may be the new nanotechnology-based strategies that are being developed to effectively deliver purified ω-3 PUFAs to the target tissues. These strategies are aimed to overcome the scarce solubility of these fatty acids, protect them from degradation, make them specifically active to target the site of injury, and/or distribute them in strict combination with other bioactive compounds/drugs. The ultimate goal is to enhance their bioavailability, thus reducing the level of intake of these fatty acids or of other co-transported drugs.Citation12–Citation14 For the first time, we have comprehensively and critically analyzed in the present review all the reports concerning the nanotechnological ω-3 PUFA-containing formulations hitherto developed, limiting our investigation to the in vitro and in vivo preclinical studies concerning the use of these nanoformulations in cellular and animal models of CVDs and cancers.
Literature search
A systematic literature search of the PubMed database was conducted from July 2017 to July 2018 to identify published peer-reviewed original articles regarding in vitro studies, in vivo animal studies, and human studies on the delivery of ω-3 PUFAs, alone, or in combination with other bioactive compounds, through nanoformulations. The key words used for the search of titles and abstracts were: “omega-3” or “n-3 PUFA” or “docosahexaenoic acid” or “eicosapen-taenoic acid” or “α-linolenic acid” or “fish oil;” and “animal studies,” or “in vitro studies,” or “in vivo studies” or “human studies;” and “nanoparticles” or “nanoformulations” and “encapsulation” and “delivery” and “nanomedicine” and “cancer” or “tumor” and “cardiovascular diseases” or “heart” and “inflammation.” We identified full-text articles written in English. The papers were chosen without restriction of time. We analyzed only the studies evaluating the biological effects of ω-3 PUFA nanoformulations and, in particular, in the cardiovascular and cancer conditions.
ω-3 PUFA-containing nanoformulations for the prevention of CVD and therapy
The prevention of CVDs is considered the main setting for ω-3 PUFA clinical application, and the major processes involved in the pathogenesis of most CVDs, including inflammation, oxidative stress, and abnormal cell proliferation,Citation15,Citation16 also represent the main targets of these fatty acids. However, the research investigating potential innovative nanomedicine strategies in CVDs is still very scarce, and currently restricted only to occlusive vasculopathies and atherosclerosisCitation17,Citation18 (; ). In the first report on this topic, Deshpande et alCitation17 investigated nanotechnology-based approaches to deliver ω-3 PUFAs in combination with other nutraceuticals/drugs to vascular walls in order to prevent occlusive vasculopathies following vascular injuries. The study was based on the recent observations demonstrating that ω-3 PUFAs and some of their bioactive metabolic derivatives (ie, the specialized proresolving mediators, including resolvins, protectins, and maresins) are crucial endogenous signals to maintain vascular homeostasis, either by exerting anti-inflammatory properties,Citation19 or by modulating the resolving phase of vascular injury and accelerating repair.Citation20
Table 1 Application of ω-3 PUFA nanoformulations in cardiovascular disease settings
Figure 1 Potential use of ALA-containing nanoemulsions against the development of restenosis and atheroma.
Note: The biological activity of nanoemulsions were evaluated in vitro or in vivo.
Abbreviations: ALA, α-linolenic acid; 17-βE, 17β-estradiol; CER, C6-ceramide; CREKA, cysteine–arginine–glutamic acid–lysine–alanine.
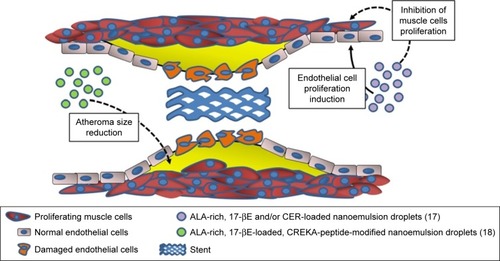
In particular, ω-3 PUFAs and their derivatives have been considered potential anti-restenosis agents, and Deshpande et alCitation17 included an ALA-rich oil (flaxseed oil) in an oil-in-water nanoemulsion system they engineered for 17β-estradiol (17-βE) and C6-ceramide (CER) (; ). This combination appeared remarkably encouraging, since ALA has been largely reported to improve CV health, and both 17-βE and CER were previously demonstrated to be cardio-protective and vasoactive.Citation21,Citation22 However, it has been reported that their effect in vivo may be markedly reduced by a scarce release due to their high hydrophobicity and capability to bind with circulating proteins.Citation23,Citation24 In addition to the observation that the two cell systems related to restenosis (human aortic endothelial cells [ECs] and vascular smooth muscle cells [VSMCs]) showed a high in vitro uptake of 17-βE and CER, the most remarkable finding was a greater biological activity of the nanoemulsion compared to each single component administered alone (ie, increased or decreased platelet-derived growth factor-induced proliferation in EC and VSMC, respectively).Citation17 The authors explained the results on the basis of the more efficient delivery of 17-βE and CER to vascular cells when they were encapsulated in ALA-based nanodroplets. For the muscular cells, they associated the findings with the increased inhibitory effect exerted by encapsulated 17-βE on the p38 MAP-kinase activity, known to be involved in VSMC proliferation. In a more recent work by the group,Citation18 the same flaxseed oil-based nanoemulsion encapsulating 17-βE was used with the clot-binding peptide cysteine–arginine–glutamic acid–lysine–alanine, for its ability to selectively bind to the atherosclerotic plaque (; ). This strategy was aimed to specifically target atherosclerotic lesions and, thus, better counteract their development and progression. The nanoemulsion was administered either to EC cultured in vitro or, intravenously, to a mouse model of atherosclerosis (apoE−/− mice fed a high-fat diet). The nanoemulsion components 17-βE and ALA were considered responsible for the increased production of nitric oxide by the EC cultured in vitro. They also minimally reduced the levels of plasma cholesterol and decreased the atherosclerotic plaque size, as well as the expression of atherosclerosis-related markers (intracellular adhesion molecule 1 (ICAM-1), vascular cell adhesion molecule 1 (VCAM-1), IL-6, tumor necrosis factor [TNF], etc.) within the aorta of the nanoemulsion-treated apoE−/− mice. The apoE−/− mice showed significantly increased levels of both triglycerides and cholesterol after a 10-week high-fat diet administration. Interestingly, the treatment with 17-βE and ALA containing nanoemulsion was able to slightly decrease, yet significantly, the level of plasma cholesterol which, conversely, remained unchanged when 17-βE was administered alone as a solution. The authors suggested that the marginal change in cholesterol induced by 17-βE and ALA nanoemulsion could partially contribute to the protective anti-atherosclerotic effects of the nanoparticles. However, they were induced to hypothesize that it was not responsible for the lesion-size reduction triggered by these 17-βE/ALA nanoformulations, since previous observations in the same mouse model of atherosclerosis had shown that the levels of circulating cholesterol were not related to the size and area of the atherosclerotic lesion.Citation25 Probably, the endothelial modifications induced by the nanoemulsion could be responsible for the size reductions of the atherosclerotic lesions.
It is worth pointing out that a recent meta-analysis did not detect any result of increased dietary intake of ALA in the blood concentrations of the same inflammation markers (soluble ICAM and VCAM, IL-6, and TNF).Citation26 This suggests that a more efficient delivery of ALA – such as the one demonstrated in the apoE−/− mouse model – could allow this fatty acid to reach vascular walls at adequate amounts to induce CV health effects, particularly if the delivery occurs in combination with other cardioprotective compounds (such as 17-βE).
ω-3 PUFA-containing nanoformulations for cancer prevention and therapy
Developed ω-3 PUFA-containing nanoformulations for liver cancer prevention and therapy
Most of the studies on new nanotechnology-based strategies for a more efficient delivery of ω-3 PUFAs to cancer cells in vitro and in vivo have been conducted on liver cells. A relevant approach was used by the Corbin group.Citation12,Citation27–Citation29 They created low-density lipoprotein (LDL)-based nanoparticles with DHA as bioactive cargo (; ), and tested them on a normal mouse liver cell line, as well as on mice,Citation12,Citation28 rats,Citation29 or human liver cancer cell lines.Citation27 Interestingly, this DHA nanoformulation was successfully employed by the same group of authors also for the delivery of DHA to the brain.Citation30 In this case, they used the systemic administration of DHA-LDL nanoparticles combined with pulsed focused ultrasound exposures. In the LDL nanoparticles, DHA was placed inside the hydrophobic core, where its stability and biological activity can be preserved. The strategy of using LDLs as ω-3 PUFA transporters to liver cells has several advantages, since LDLs have a great capacity of carrying active substances, and they are naturally present in circulation, so that they are not recognized and internalized by mononuclear circulating phagocytes.Citation31 Moreover, LDLs carry and supply cells with lipids also in physiological conditions via LDL receptor-mediated endocytosis and seem particularly suitable to deliver bioactive lipid factors to cancer cells, which show higher avidity for lipids than normal cells, due to their sustained membrane turnover.Citation32–Citation34 Indeed, one particularly interesting feature of these engineered nanoparticles was their ability to be selectively cytotoxic to hepatocarcinoma cells, over the nonmalignant counterparts.Citation12,Citation28 This specificity for cancer cells is highly advantageous for antineoplastic agents, and, presumably, it was related to the fact that high concentrations of the cell growth inhibitor DHA were achieved only inside the cancer cells. Particularly, the selective growth-inhibitory effect of the LDL-DHA nanoparticles in liver cancer cells was explained on the basis of the specific and powerful anti-inflammatory and pro-resolving properties of the ω-3 PUFAs and their bioactive derivatives. Moreover, it was also related to increased levels of reactive oxygen species (ROS) and iron found in cancer cells, as well as to the reported reduced glutathione (GSH) cellular depletion.Citation35,Citation36 This pro-oxidative condition of cancer cells may easily induce peroxidation of a highly unsaturated compound, such as DHA.Citation37 In turn, the peroxidation of intracellular membranes is triggered, and that may lead to lysosomal, mitochondrial, and nuclear disruption, and, lastly, cell death.Citation28 Of great significance, a single intra-arterial injection of LDL-DHA in rats bearing transplanted orthotopic hepatocarcinoma markedly reduced tumor growth by altering redox balance and inducing tumor cell death.Citation29 The most recent application of LDL-DHA particles by these authors convinced them to formulate the stimulating hypothesis that DHA enclosed in these particles could induce a recently described regulated type of necrotic cell death called ferroptosis in liver cancer cells.Citation27 This non-apoptotic iron-dependent form of cell deathCitation38 is characterized by the simultaneous intracellular occurrence of increased lipid peroxidation, high levels of intracellular ROS originating from iron metabolism, and glutathione depletion. Moreover, it is critically and negatively regulated by the lipid antioxidant enzyme glutathione peroxidase-4 (GPX-4).Citation27 In line with this, LDL-DHA also substantially reduced the expression and activity of GPX-4 in liver cancer cells, along with causing all the pro-oxidative changes typical of ferroptosis.Citation27 Since it is usually reported that DHA – given as a free fatty acid – mainly causes apoptotic death in cancer cells,Citation39 the finding that LDL-DHA, on the other hand, triggers ferroptosis would imply that, depending on the delivery system used, ω-3 PUFAs may induce different death pathway inside the cells (ie, apoptosis or ferroptosis). The authors associated this effect to the fact that – while free DHA enters the cells by diffusion or facilitated transportCitation40 – LDL-DHA nanoparticles are endocytosed in large amounts. This different entrance pathway into the cells could enable DHA to easily reach more internal subcellular structures and, thus, exert considerable effects on them, thus contributing to the ferroptotic pathway of cytotoxicity.Citation27
Table 2 Application of ω-3 PUFA nanoformulations in liver cancer
Figure 2 Omega-3 PUFA-containing nanomaterials developed and evaluated in vitro or/and in vivo for their potential use against liver (A), breast (B) and lung (C) cancer.
Abbreviations: DHA, docosahexaenoic acid; LDL, low-density lipoprotein; LNA, linolenic acid; PEG, polyethylene glycol; PL, phospholipids; PεCL, poly-ε-caprolactone; PUFA, polyunsaturated fatty acid; SLN, solid lipid nanoparticle.
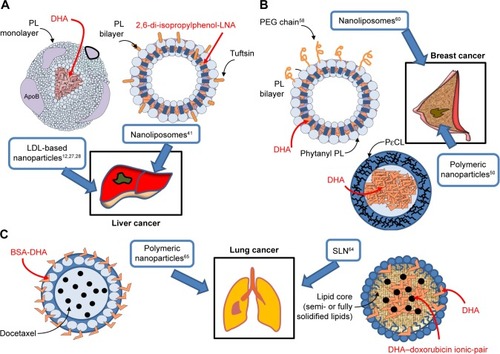
Another interesting approach for the nanodelivery of ω-3 PUFAs was recently used to deliver ALA to diethylnitrosamine-induced hepatocarcinomas in mice.Citation41 In this case, ALA was conjugated with 2,6-di-isopropylphenol, an anesthetic agent being structurally similar to vitamin E. The chemical structure of this conjugate was reported in Khan et al.Citation42 This compound, conjugated with DHA, was previously shown to possess antioxidant properties and the ability to induce apoptosis and decrease the metastatic potential of human cancer cells.Citation43 For an efficient delivery, the 2,6-di-isopropylphenol-linolenic acid conjugate was enclosed in nanoliposomes tagged with the tetrapeptide tuftsin (Thr-Lys-Pro-Arg),Citation44 a fraction of the immunoglobulin G molecule (; ). The use of tuftsin-tagged liposomes was previously and successfully used to deliver antibiotics in the area of infections,Citation45 due to the immunomodulatory properties of tuftsin.Citation44 It is worth highlighting that tuftsin was also previously administered in combination with antineoplastic drugs and shown to enhance their antitumor activity.Citation46,Citation47 The treatment with tuftsin-tagged liposomes containing the ω-3 PUFA conjugateCitation41 increased the survival of the hepatocarcinoma-bearing mice, and this effect was partly related to the markedly reduced expression of COX-2 and Bcl-2, as well as the upregulation of Bax observed in hepatocellular carcinomas. In our opinion, the co-presence of tuftsin and ω-3 PUFAs inside the same nanoliposomes is significant, since ω-3 PUFAs also show both immunomodulatory and antineoplastic activities at the same time, and their possible applications in the field of cancer immunotherapy have been recently suggested.Citation48,Citation49 Moreover, due to the safe and non-toxic nature of both ω-3 PUFAs and tuftsin and their common immunomodulating properties, the application of this liposomal delivery could be potentially useful, not only for the treatment of hepatocar-cinoma, but also for its prevention in populations at high risk (such as hepatitis C virus (HCV)-positive patients, alcoholics, or patients with inborn errors of metabolism, including type I glycogenosis or type I tyrosinemia).
Developed ω-3 PUFA-containing nanoformulations for breast cancer prevention and therapy
Roy et alCitation50 developed polymeric nanoparticles (150–200 nm) encapsulating DHA (; ) with the aim of using them in breast cancer. The polymeric wall of the nano-capsules (NCs) consisted of poly-ε-caprolactone, a polyester able to protect the small liquid droplets of DHA included in the internal part (core) from chemical and enzymatic degradation, protecting DHA against the adverse conditions present in the gastrointestinal tract. This nanoencapsulation approach was previously used for the delivery of different drugs achieving positive results, such as enhanced drug efficacy and decreased drug toxicity.Citation51 Roy et alCitation50 observed a clear cytotoxic effect (measured by the MTT assay) when they administered 50–100 µM DHA in the form of free fatty acid for 4 days to MDA-MB-231 cultured in vitro, whereas lower concentrations showed no effect. This finding was in line with the results of several papersCitation52–Citation56 analyzed by us in a recent review,Citation57 showing that treatments of the triple-negative MDA-MB-231 or MDA-MB-453 breast cancer cells with EPA or/and DHA in the 30–100 µM range for 1–5 days were necessary to inhibit cell growth and proliferation, and to enhance apoptosis or breast cancer cell sensitivity to docetaxel (DTX). Unexpectedly, however, Roy et alCitation50 found that, when lower concentrations of DHA (1–10 µM) were enclosed in NCs, they induced the proliferation of MDA-MB-231 cancer cells. On the other hand, higher concentrations (30–50 µM) of encapsulated DHA reduced breast cancer cell proliferation, similarly to what was obtained with free DHA. The simultaneous presence of vitamin E inside the NCs eliminated the growth inhibiting effect of DHA. Moreover, encapsulated DHA became more toxic than free DHA if H2O2 was added inside the NCs. Based on these results, the authors hypothesized that DHA should be oxidized and, thus, release secondary toxic products of lipid peroxidation which may be responsible for its antiproliferative effects. However, the authors suggested that H2O2 could have also acted by disrupting the nanoparticle wall, releasing DHA outside the nanoparticles and, therefore, increasing its biological activity. Otherwise, due to its hydrophobic nature, this fatty acid tends to remain bound to the oily core of these nanoparticles and lose its bioactivity. Overall, however, the delivery of DHA to scarcely differentiated breast cancer cells through this kind of polymeric nanoparticles does not appear promising for breast cancer therapy, as the antineoplastic activity of this fatty acid appears to decrease when it is encapsulated at high concentrations. Moreover, when it is encapsulated at lower concentrations, it seems to even promote breast cancer cell growth.Citation50
Table 3 Application of ω-3 PUFA nanoformulations in breast cancer
On the other hand, Alaarg et alCitation58 enclosed DHA into liposomes by using 1,2-dipalmitoyl-sn-glycero-3-phosphocholine, cholesterol, and distearoyl phospha-tidylethanolamine (PE); and polyethylene glycol (PEG) and these liposomes were used for the delivery of DHA to murine breast cancer cells in vitro (; ). PEGylated liposomes were chosen for their acknowledged ability to stay in the circulation for a longer period (about 1 day half-life) compared to non-PEGylated particles that are cleared from the circulation in ≤1 hour.Citation59 This implies that a bioactive compound (in this case, DHA) enclosed in these nanoparticles can have more chances to be taken up by cells and tissues. DHA-loaded liposomes were able to inhibit the proliferation of murine 4T1 breast cancer cells, and other kinds of epithelial cells. However, it is worth underlining that, in this study, the effects induced by the DHA-loaded liposomes were compared only to those induced by the unloaded liposomes. For a clearer understanding, however, it would have been essential also to evaluate the effects of DHA administered in its free fatty acid form.
Skibinski et alCitation60 evaluated the improvement in the bioactivity of DHA included in a different kind of lipo-somes administered to MCF-7 and MDA-MB-231 breast cancer cells in vitro. Their DHA-loaded liposomes contained 1,2-dihexadecyl-sn-glycero-3-phosphocholine and 1,2-diphytanyl-sn-glycero-3-phosphoethanolamine (; ), phytanyl lipids similar to those present in Archaea prokaryotes and known to be highly resistant to pH and temperature changes. The authors observed that DHA loaded into the liposomes reduced more the viability of breast cancer cells and induced apoptosis more efficiently in both the cell lines. Inside the liposomes, only DHA was responsible for these effects, since the empty phytanyl lipid liposome did not significantly alter cell viability and apoptosis. Moreover, the authors found that DHA enclosed in the liposomes could more effectively decrease the expression of the activated form of Akt (p-AkT). They hypothesized that this effect could explain the increased antineoplastic activity showed by DHA loaded in the liposomes. This hypothesis is particularly interesting, since p-Akt represents a biomarker associated with poor prognosis in breast cancer, and used to evaluate the activity of the tyrosine kinase receptor signaling, often deregulated in breast cancer carcinogenesis.Citation57,Citation61
A possible future ω-3 PUFA-containing nano-based approach for breast cancer may be suggested on the basis of the above analyzed remarkable results, obtained by both Alaarg et alCitation58 and Skibinski et alCitation60 with DHA-loaded nanoliposomes. Since ω-3 PUFAs are known to potentially act as an adjuvant therapy to strengthen the activity of conventional chemotherapeutics or targeted drugs in the therapy of breast cancer,Citation57,Citation62 it would be remarkable to develop nanoliposomal structures that are able to deliver simultaneously both ω-3 PUFAs and drugs with previously acknowledged activity in breast cancer. This liposomal combined approach could also be experimented in the clinical setting, since some liposomes are known to be safe and have already been clinically approved. Moreover, liposomes are able to encapsulate both hydrophobic and hydrophilic compounds and show a high capacity, making them particularly suitable to transport combinations of antineoplastic agents with different activities (conventional, targeted, or immunoactive agents),Citation63 including also safe nutritional factors with multiple anticancer functions, such as ω-3 PUFAs.
Developed ω-3 PUFA-containing nanoformulations for lung cancer prevention and therapy
To date, only two studiesCitation64,Citation65 have evaluated the anticancer properties of ω-3 PUFA-containing nanoformulations on lung cancer cells (; ), and their main aim was to improve the delivery and antitumor efficacy of doxorubicin (DOX) and DTX, two chemotherapeutic agents widely used in the therapy of solid tumors. A variety of nanotechnological approaches have been investigated so far to improve the limited uptake and distribution of DOX in solid tumors.Citation66,Citation67 Indeed, DOX is a weakly basic compound that becomes protonated when exposed to the slightly acidic pH of the microenvironment surrounding cancer cells, making its cellular uptake difficult.Citation66 Liposomes, solid lipid nanoparticles (SLNs), and polymeric micelles are among the different types of DOX nanocarriers designed to overcome this problem. A PEGylated liposomal formulation containing DOX (Doxil) has also been approved for the treatment of some cancers (ovarian cancer, AIDS-related Kaposi sarcoma, and multiple myeloma).Citation68 Mussi et alCitation64 designed DOX-loaded SLN for lung cancer, where the oily phase included DHA and surfactants, such as Compritol, Tween 80, and triethanolamine (; ). The SLN formulation allows a site-specific release of DOX in a slightly acidic environment, such as tumor tissues.Citation66,Citation69 Moreover, the SLN solid matrix allows a more controlled release, and the lipophilicity of these nanoparticles improves the interaction with the cellular membranes. Furthermore, the SLNs obtained by encapsulating 0.4% DHA (w/v) were of smaller size (94±1 nm), which is advantageous, being the nanoparticles in the range of 100–200 nm taken up and retained at a higher concentration inside cancer cells compared to particles with a higher size.Citation70–Citation72 The simultaneous encapsulation of DOX with a lipophilic anion, such as DHA in the SLN, was aimed to overcome the low level of incorporation in lipidic nanostructures that DOX usually exhibits, due to its cationic amphiphilic nature. Moreover, the SLN exhibited negative charges due to the presence of DHA, and, inside the SLN, DOX formed an ion-pairing with DHA that could explain the high DOX encapsulation effi-ciency (almost 100%), as well as the enhanced cytotoxicity shown by the DHA-DOX-loaded SLN against A549 lung adenocarcinoma cells, as compared to free DOX and DHA administered in combination.
Table 4 Application of ω-3 PUFA nanoformulations in lung and prostate cancer
The second study was performed by Zu et al,Citation65 who evaluated the anticancer activity of ω-3 PUFA-containing polymeric nanoparticles on lung cancer cells. The nanoparticle surface contained DHA covalently bound to BSA, a natural biodegradable polymer often used to prepare polymeric nanoformulationsCitation73 (; ). Also in this case, the size of the newly developed nanoparticle (141.7±7.9 nm) was within the range of sizes allowing a more efficient uptake by cancer cells.Citation70–Citation72 The DHA-BSA nanoparticles were designed to enclose DTX, an approved semisynthetic antineoplastic drug belonging to the taxane family and, at present, largely used for the treatment of a variety of solid cancers, including lung and breast cancer.Citation74,Citation75 Its inclusion in the nanoparticle was aimed to overcome the scarce aqueous solubility of the drug. Numerous formulations of taxanes have been developed over the last decades in order to increase the aqueous solubility of the hydrophobic taxanes, such as cyclodextrin, liposomes, and albumin-bound nanoparticles (Abraxane) formulation.Citation73,Citation76 In the nanoformulation analyzed,Citation65 the addition of DHA to the albumin on the surface of the DTX-BSA nanoparticle was aimed to specifically target cancer cells, since it was previously observed that DHA preferentially accumulates in tumor cells and is considered a target ligand for cancer cells.Citation77–Citation79 This tumor targeting ability of DHA has been typically associated with the increased fluidity shown by cancer cell membranes which contain high levels of this fatty acid.Citation77–Citation79 In turn, the enhanced fluidity was related to their higher content in phospholipids, and, particularly, in PE, which was found to have a specific affinity for DHA.Citation77 Above all, targeting lung cancer cells with these nanoparticles seems mainly appropriate since PE has been found to be overexpressed not only in the cancer cells used in this study (murine Lewis Lung Carcinoma [LLC] cells)Citation77 but also, more in general, in the human non-small-cell lung cancers.Citation80 Furthermore, the addition of an antineoplastic natural agent, such as DHA, to polymeric DTX-BSA nanoparticleCitation65 was also related to the results of several human studies demonstrating that the therapeutic efficacy of this taxane can be increased after its conjugation to DHA, which is also able to decrease the side effects usually induced by taxane therapy, and improve the taxane delivery to the tumor tissues.Citation81–Citation86 Only the choice of BSA to prepare these polymeric nanoformulations could be controversial, since BSA was reported to antagonize the effect of DHA.Citation13 The new DTX-DHA-BSA nanoparticles were administered to lung cancer cells cultured in vitro and to mice injected with syngeneic LLC cells in the tibia of the right limb, which mimicked lung cancer metastasis in the bone.Citation65 The finding demonstrated that the simultaneous presence of DHA in the nanoparticles enhanced the anticancer activity of taxanes both in vitro and in vivo, and also increased the mean survival time of the mouse.
Developed ω-3 PUFA-containing nanoformulations for prostate cancer prevention and therapy
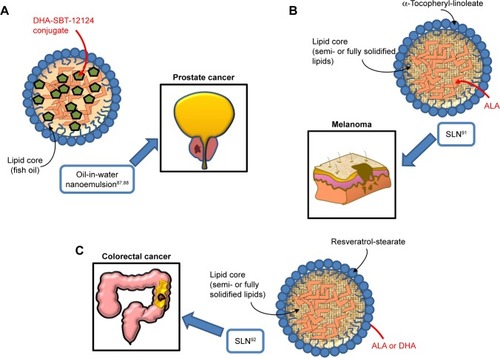
Similarly to the results of the work by Zu et al,Citation65 where nanoformulations containing DHA bound to a taxane were tested in lung cancer, two studies performed on prostate cancer used an oil-in-water nanoemulsion formulation,Citation87,Citation88 where the starting oil phase consisted of fish oil with a novel DHA-taxoid conjugate (DHA-SBT-1214) (; ). The oil phase was mixed with the water phase formed by egg phosphatidylcholine (Lipoid E80®), polysorbate 80 (Tween80®), and 1,2-distearoyl-sn-glycero-3-phosphoeth-anolamine-N-(amino[polyethylene glycol]-2000) (DSPE-PEG2000). Both the empty nanoemulsion (not containing drugs) and the nanoemulsion containing DHA-SBT-1214 showed spherical shape and a very similar particle size (225±7 and 228±7 nm, respectively). The surface charge of the oil droplets in the two formulations did not change even using the maximum encapsulation of DHA-SBT-1214 within the oil droplets. The two studies are remarkable, since they benefit from a new-generation taxoid (SBT-1214), bound to DHA, and avoid the use of albumin, which can reduce the anticancer effect of DHA.Citation13 In particular, the use of SBT-1214 was aimed to overcome the mechanism through which placlitaxelCitation89 or DTXCitation90 was observed to induce drug resistance in prostate cancer (by mechanisms related to the induced expression of either P-glycoprotein or transforming growth factor-β superfamily components, respectively). On the other hand, the rationale of SBT-1214 binding to DHA was related to the above-mentioned ω-3 PUFA ability to exert cancer-specific toxicity, to the synergistic antineoplastic effects that these fatty acids exert when bound to anticancer drugs, and to their ability to protect non-transformed prostate cells from cytotoxicity and reduce the secondary systemic harmful effects of conventional anticancer drugs, including taxanes. Moreover, the use of the non-ionic emulsifying agent Tween 80 in the preparation of this nanoemulsion is relevant, as it allows to obtain optimal oil droplet size and charge by inhibiting aggregation through competitive absorption to interfaces. This surfactant was also used for the preparation of other nanoemulsions containing ω-3 PUFAs and herein analyzed.Citation64,Citation91,Citation92 The newly designed nanoemulsion of DHA-SBT-1214 (NE-DHA-SBT-1214) was tested for its antineoplastic activity against human prostatic cancer PPT2 cells cultured in vitro, or transplanted subcutaneously in NOD/SCID mice.Citation87,Citation88 These prostate cancer cells have the features of stem-like cells and are considered a good model for testing new drugs for prostate cancer. This is a highly recurrent tumor, which has been related to the presence of high levels of cancer stem cells with the potential of self-renewing the tumor.Citation93–Citation95 The NE-DHA-SBT-1214 exerted higher cytotoxicity toward the PPT2 cells cultured in vitro, compared with DHA-SBT-1214 in solution. In addition, intravenous administrations of this nanoemulsion carrier system containing DHA-SBT-1214 in fish oil droplets to the transplanted mice markedly suppressed prostate cancer cell growth in vivo compared to a placebo nanoemulsion formulation previously approved for the treatment of metastastic human breast cancer (Abraxane™, ie, paclitaxel albumin-bound particles). Moreover, the prostate cancer cells that still remained viable after the in vivo treatment with NE-DHA-SBT-1214 had reduced neoplastic potential, which was shown by their decreased ability to give rise to colonies and spheres when analyzed ex vivo for clonogenic and sphere-forming capacities. These results suggest that the use of NE-DHA-SBT-1214 for the treatment of prostate cancer could be a promising strategy.
Figure 3 Omega-3 PUFA-containing nanomaterials developed and evaluated in vitro or/and in vivo for their potential use against prostate (A), melanoma (B), and colorectal cancer (C).
Abbreviations: PUFA, polyunsaturated fatty acid; DHA, docosahexaenoic acid; SLN, solid lipid nanoparticle; ALA, α-linolenic acid.
Developed ω-3 PUFA-containing nanoformulations for melanoma and colorectal cancer (CRC) prevention and therapy
We have studied the antineoplastic potential of ω-3 PUFAs in CRC and melanoma for many yearsCitation96–Citation101 and investigated the possible mechanisms underlying the beneficial action of these fatty acids. We have recently developed new SLNs for the delivery of ω-3 PUFAs to C32 melanoma cellsCitation91 and HT-29 and HCT116 CRC cells,Citation92 to reduce possible ω-3 PUFA oxidative degradation of these compounds, and, thus, prolong their shelf-life, enhance their bioavailability, and improve their antineoplastic activity (; ). In particular, the matrix of the SLN tested in melanoma cells contained ALA esterified to α-tocopherol and encapsulated ALA itself. ALA carried in these nanoformulations was always more cytotoxic than ALA or α-tocopherol administered alone to melanoma cells.Citation91 On the other hand, the lipid matrix of the SLN formulation tested in CRC cells contained stearic acid esterified to the natural dietary antioxidant resveratrol (RV). Furthermore, these RV-based SLN (RV-SLN) carried either ALA or DHA inside.Citation92 Compared with the other SLN tested on melanoma cells, the substitution of ALA in the matrix with stearic acid was aimed to further increase the hydrophobicity of the encapsulated ω-3 PUFAs, thus potentially improving the efficiency of their delivery. Even though stearic acid is a saturated fatty acid, it has been shown that it may lower serum LDL cholesterol levels,Citation102 and also protect from the development of prostate cancer.Citation103,Citation104 In particular, this protective action appears remarkable, since it has been recently hypothesized that the incidence of prostate cancer may be associated with a high intake of ω-3 PUFAs. Thus, the combination stearic acid/ω-3 PUFAs inside the SLN could neutralize possible prostate-specific adverse effects of these fatty acids.Citation105 Moreover, the encapsulation of both ALA and DHA makes them more efficient in inhibiting CRC cell growth, by enhancing their inhibitory effect on cell proliferation rather than further increasing their already high ability to induce apoptosis.Citation92 These increased antineoplastic effects were strictly related to the enhanced cellular incorporation observed when DHA and ALA were enclosed in the RV-SLN to CRC cells. It was remarkable to find that, only after the encapsulation, ALA induced an evident cellular increase in both EPA and DHA and that encapsulated DHA was more efficiently retroconverted to EPA. This implies that the encapsulation of either DHA or ALA into RV-based SLN could also markedly increase the CRC content of EPA, which, according to several recent reports, seems to protect against CRC risk better than all the other main ω-3 PUFAs.Citation106–Citation108 The more efficient conversion of SLN-encapsulated ALA is particularly interesting when considering that ALA is present at high levels in a variety of seeds and vegetables, and its increased intake could be achieved more easily and in a more sustainable way compared to an increased consumption of EPA itself.
Table 5 Application of ω-3 PUFA nanoformulations in melanoma and colon cancer
In conclusion, it is important to underline that the possibility to further enhance the protective activity of these dietary compounds against CRC is appealing, since the possible prevention and treatment of CRC with PUFA has involved many researchers in the past two decades.Citation96,Citation109–Citation112 In particular, possible successful future investigations in the field of CRC nanomedicine-based therapy could be related to the powerful modulator effect recently demonstrated for ω-3 PUFAs on the intestinal microbiota.Citation107,Citation113 Indeed, since the microbiome may deeply affect the individual response to cancer immunotherapy, it would be worth investigating the approach of combined nanodelivery of immunomodulating innovative drugs, already used for CRCCitation114,Citation115 with ω-3 PUFAs against CRC in vivo. Moreover, co-delivering powerful anti-inflammatory agents, such as ω-3 PUFAs, and these drugs in nanoparticles could also have the potential to overcome the severe colon immune-related adverse inflammatory events induced by these innovative treatments in CRC patients.Citation116
Mixed and contrasting outcomes of recent human studies and the well-known benefits of ω-3 PUFAs in CVD and cancer
We cannot avoid focusing hereinafter on the topic outlined by the following question: are the contrasting outcomes of recent large human trials weakening the accepted dogma of ω-3 PUFA healthy effects? In our opinion, this topic deserves particular attention, particularly by the researchers who have been studying for decades the beneficial effects of ω-3 PUFAs in the prevention of CVD and cancer.Citation1,Citation39,Citation57,Citation62,Citation96,Citation98,Citation117 Even though this issue cannot be tackled exhaustively in just one paragraph, it is essential to outline some considerations.
As far as the ω-3 PUFA cardioprotective role is concerned, the beneficial outcomes of early human studies (epidemiological studies, prospective cohort studies, and randomized controlled trials) appear in contrast with the mixed or less positive results of some recent prospective trials, and meta-analysis of clinical trials.Citation117 Indeed, they provided scarce or no evidence that ω-3 PUFA may reduce CV events or mortality (for a review see Bowen et alCitation118). It has been hypothesizedCitation117 that these discrepancies may be attributed to the difficulty of ω-3 PUFA to induce healthy effects in a completely changed scenario. In fact, in recent years, the enrolled patients have often been simultaneously subject to the administration of statins and/or other pharmaco-logic agents widely used to prevent CVD. Moreover, higher levels of ω-3 PUFA containing foods have been generally added to the diet following the numerous recommendations formulated over the last two decades. The results of the recently published VITAL studyCitation119 also obtained mixed and less promising results. It had the novelty of being a large, 5-year randomized, double-blind, placebo-controlled trial of a daily fish oil supplementation (1 g, Omacor® fish oil) for primary prevention in healthy subjects at baseline. On the contrary, the earlier trials mostly evaluated whether a supplementation with fish oil or purified ω-3 PUFAs could prevent CV adverse events in patients with a history of CVD or at high risk for CVD. Interestingly, the primary outcomes of the VITAL study indicated only a small but non-significant decrease in the rate of a major adverse CV event (ie, myocardial infarction [MI] or stroke) in the ω-3 PUFA group with respect to the placebo group. However, a secondary analysis in those patients that were specifically affected by stroke or MI revealed that the intervention was able to significantly reduce the risk of MI (by 28%) and the death rate of MI (by 50%). Interestingly, a much higher reduction in MI risk (by 77%) was observed in the African subpopulation, thus enforcing the notion that the ω-3 PUFA beneficial effects may be modified depending on the individual genetic background.Citation120 Remarkably, the need for angioplasty was also significantly reduced (by 22%) by the fish oil supplementation. The accuracy of this trial was also demonstrated by the evaluation of the basal plasma ω-3 index (EPA + DHA as a percentage of total fatty acids) of the subjects, which was evaluated also after 1-year treatment to show its increase (by about 50%). The outcomes of another recently published randomized double-blind, placebo-controlled trial (REDUCE-IT) reported more positive results. However, it evaluated the effects of simultaneous treatment with a very high dose (4 g/day) of purified EPA and statin therapy for almost 5 years in patients with CVD or at high risk for CVD and high fasting triglyceride levels.Citation121 The study showed that the EPA treatment reduced the levels of serum triglycerides, CV events, and CV death. In particular, the rates of emergency or urgent revascularizations or MI were reduced by the EPA supplementation, even though the high dose used slightly increased the rate of bleeding events (see next paragraph), as well as the onset of atrial arrhythmias. It can be underlined that many differences do exist between these two most recent and largest trials, which can however help us to better understand the actual potential of ω-3 PUFAs in preventing CVD. For instance, the use of fish oil at a quite low dose may lack in showing the induction of health CV effects. On the contrary, EPA administered alone at a very high dose, even though in the presece of a statin therapy, may elicit beneficial effects, increasing, however, also the risk of some detrimental properties. Overall, the debate on the ω-3 PUFA-induced CV benefits is ongoing, also considering the manifold data from in vitro studies and in vivo animal studies supporting a variety of different biological and molecular mechanisms whereby ω-3 PUFA may confer coronary protection and reduce the risk of CVD.Citation122 Therefore, further large interventional trials comparing different doses and types of fatty acid (EPA and DHA) are needed for a better knowledge of this topic. Moreover, it would be more advantageous that the subjects/patients would be stratified according to the ω-3 index expressed as the content of EPA and DHA in red cell membranes, which is considered a more reliable and stable indicator of the real ω-3 PUFA intake than their plasma level.Citation123,Citation124 Moreover, it would be important to evaluate the ω-3 index several times along the long-term studies.
With regard to cancer, plenty of evidence obtained from in vitro and in vivo preclinical studies has clearly indicated the anticancer activity of omega-3 PUFA.Citation120 On the contrary, as we have widely discussed in a recent critical review of the literature,Citation120 the results of the human studies are more controversial. In particular, most of the recent results confirmed the beneficial effects of the preclinical studies. However, there are studies where no effect was observed, or the antineoplastic effects were confined only to specific subpopulations (obese subjects, with specific dietary habits or with a particular fatty acid pattern, or having a specific T-cell infiltration in the tumor microenvironment). Only a few reports found positive associations between the development/progression of cancer (mainly for prostate cancer), and the high level of ω-3 PUFA introduced with the diet, present in blood, or accumulated in tissues. In our opinion, to clarify the issue, it would be essential to know the actual levels of ω-3 PUFA intake and the efficiency of their incorporation in the tissues of each subject along the study period. To this end, we recommended repeated evaluations of biomarkers reflecting the intake of ω-3 PUFAs, and in particular, the ω-3 index expressed as the content of EPA and DHA in red cell membranes.Citation120 Moreover, we suggested, whenever possible, not to overlook some characteristics of the subjects being studied, such as the race/genetic background, and the metabolic/nutritional status.
Interestingly, the outcomes of the more recent interventional VITAL studyCitation119 indicated that the daily supplementation with 0.8 g/day ω-3 EPA + DHA was unable to lower the incidence of different kinds of invasive cancer of any type, as well as that of colorectal, breast, and prostate cancer, or the incidence of death from cancer. These findings were consistent with the results of two recent meta-analyses of ω-3 PUFA trials of CVD that had shown either no association between the supplementations with ω-3 PUFA and the risk of cancer in generalCitation125 or a slightly increased but not significant risk.Citation126 Again, however, attention should be focused on the doses supplemented and the consequent ω-3 PUFA tissue enrichment, that is, in our experience, the most related parameter to the antineoplastic activity of these fatty acids.Citation124 It is possible that the daily dose of about 1.0 g (generally recommended for the secondary prevention of CVD) may not be sufficient to induce any anticancer effect. In a recently published review,Citation124 we critically analyzed the doses of ω-3 PUFA that had been able to induce anticancer effects both in the animal studies and in the few available interventional human studies. We found that supplementation to humans with safe doses of EPA + DHA of ~2.0 g/day produced a plasma and tissue enrichment in these fatty acids of about 2–5 folds, which was associated with significant anticancer effects.Citation127–Citation129 Higher doses were not able to further increase the amount of EPA and DHA incorporated in tissues. In mice or rats the dietary doses of EPA + DHA that have clear antineoplastic effects were variable in relation to their body mass and surface. However, these doses always produced an increase of ~2–5 folds of these fatty acids in the plasma, as well as in the normal and neoplastic tissues. Altogether, these observations prompted us to suggest that a daily dose of about 2 g could be the most appropriate for future human studies evaluating the anticancer effects of ω-3 PUFAs.
Possible adverse effects of ω-3 PUFAs: are they real obstacles to their use?
Increased bleeding risk
It is not possible to completely overlook here the controversies regarding the possible adverse effects of high doses of ω-3 PUFAs. In particular, high intakes of ω-3 PUFAs have been suggested to exert adverse effects on bleeding risk, since, in an earlier study, the diet with high levels of ω-3 PUFAs of the Greenland Inuit population was associated with an increased bleeding time.Citation130 The finding that these fatty acids were able to displace the ω-6 PUFA arachidonic acid from cell membranes and decrease its metabolic conversion to the powerful pro-thrombotic platelet-aggregating factor thromboxane A2, further substantiated this hypothesis.Citation96 Moreover, increased intakes of ω-3 PUFAs were also reported to be associated with reduced blood levels of several clotting factors such as factor V, prothrombin, and von Willebrand factor.Citation131 However, a recent review of the literatureCitation132 examining this issue supported the safe consumption of EPA and DHA even in highly vulnerable and sensitive populations, such as patients having gastrointestinal cancer or hospitalized in Intensive Care Units and consuming fish oil–enriched medical nutrition. No increased risk of bleeding-related event was identified in the analyzed reports, even if the patients were simultaneously treated with platelet aggregation inhibitors. On the contrary, the results concurred to support the safety of either very high daily doses of EPA + DHA given for a short period (up to 10 g/day) or much lower doses consumed for 1 year. Results obtained more recently by the large VITAL study having a 5-year median follow-up confirmed these findings.Citation119 In this study, healthy subjects supplemented with 1 g/day fish oil (providing 460 mg EPA + 380 mg DHA) did not show increased major bleeding episodes as compared to the subjects of the placebo group. The other recent large REDUCE-IT studyCitation121 was performed on patients already receiving statin therapy and showing high levels of serum triglycerides, patients with a previously diagnosed CVD or at high risk of developing CVD. They were supplemented for almost 5 years with a high daily dose (4 g) of purified EPA. This study reported a modestly but significantly higher overall rate of adverse events, but not fatal bleeding events in the EPA group (2.7% vs 2.1% in the placebo group). However, it was observed that there were no significant differences between the placebo end EPA groups in severe bleeding events, such as hemorrhagic stroke, serious central nervous system bleeding and gastrointestinal bleeding. Moreover, CV patients are often subject to chronic antithrombotic therapies, and this factor was not considered in the study. In any case, the inclusion of ω-3 PUFA in nanoformulation and the consequent increased efficiency of their delivery and absorbance at specific sites may prevent the induction of adverse bleeding effects at the circulatory level.
Immunosuppression
Concern has also been expressed/raised over the possibility that ω-3 PUFAs could have the potential to suppress any possible basal immunoreactivity against cancer, and decrease the impact of cancer immunotherapy.Citation133 This alarming possibility is related to the widely acknowledged anti-inflammatory and immunosuppressive activities of ω-3 PUFAs, and it is clear that their inclusion in nanoformulations to increase their bioactivity could also intensify these effects. However, in the last decades, a great deal of information has been gathered supporting the anti-cancer activities of ω-3 PUFAs, which oppose the preconceived alarm of a possible ω-3 PUFA cancer-promoting activity.Citation120 Moreover, it should be underlined that, whereas acute inflammation is a response to antagonize infections, repair, and resolve tissue damage, chronic inflammation has been involved in the promotion of cancer, by inducing immunosuppression in tumor microenvironment.Citation134,Citation135 Per se, it is considered a hallmark of cancer. To this respect, ω-3 PUFAs reveal their multifaceted nature, on one hand, by inhibiting persistent inflammation, thus preventing the promotion of cancer, and by also exerting pro-inflammatory actions to inhibit the immunosuppression existing in the tumor environment.Citation136 In fact, recent findings have shown that one of the mechanisms through which ω-3 PUFAs may inhibit tumor development in vivo is the induction of a local antitumor immune response.Citation137,Citation138 In particular, Liang et alCitation137 demonstrated a decreased expression of markers of tumor-associated macrophages in the tumor microenvironment of immunocompetent mice bearing prostate cancer and fed a fish oil–enriched diet. They also observed a decreased expression of a chemokine (CCL-2) that specifically recruits monocytes and macrophages to inflammatory sites.Citation137 It was also hypothesized that more than the dietary intake of ω-3 PUFAs by themselves, an increased dietary ω-3 PUFA/ω-6 PUFA ratio could maximally and beneficially influence the immune characteristics of the tumor microenvironment and reduce the risk of developing some forms of cancer.Citation139 This was demonstrated by Gevariya et alCitation138 that used mice transplanted with syngeneic prostate transgenic cell line and supplemented with a high ω-3 PUFA/ω-6 PUFA ratio diet (3.3). They found increased levels of cytokines associated with Th1 and Th2 immune response in the tumors growing in these mice compared to those of mice fed a diet with a much lower ω-3 PUFA/ω-6 PUFA ratio (0.002). These findings are in line with recent in vitro studies showing that ω-3 PUFAs are able to stimulate the innate and the adaptive immune response, both in the humoral and in the cellular branch.Citation140–Citation142
Finally, considering the immunosuppressive activity of ω-3 PUFAs, it could be argued that they may have the potential to inhibit the immune response to infections. Again, this detrimental effect could be augmented by their inclusion in nanoformulations aimed to increase their efficacy. On the contrary, however, according to recent relevant results comprehensively analyzed by Costantini et al,Citation143 dietary ω-3 PUFA can positively modulate the immune response to microbe infections. In particular, evidence has been provided that the strict relationship between ω-3 PUFAs, gut microbiota, and immune system may influence the activity of the host immune cells.Citation143 In particular, plenty of animal studies and some evidence in humans have demonstrated that dietary supplementations with ω-3 PUFAs are able to normalize the gut microbial imbalance.Citation143 This appears interesting, since the intestinal microbiota has been reported to exert a crucial role in the development of the systemic and gut immune response.Citation144,Citation145
Conclusion
To date, only a limited number of recent studies performed in vitro or in animals have directly investigated the biological effects of newly developed ω-3 PUFA-containing nanomaterials in CVDs and cancer. To date, none of these nanoformulations have been tested in the clinical setting. The studies have almost univocally demonstrated that a more efficient delivery of ω-3 PUFAs through these newly developed nanosystems may markedly enhance the therapeutic potential of nutraceuticals and other co-delivered drugs toward inflammatory lesions and cancer. Different strategies have been used, and these fatty acids have generally been included in the newly developed systems because of their ability to induce healthy effects. However, in some cases, they were also used just as a component of the lipidic phase to obtain nanoparticles carrying different bioactive products. Moreover, their position in the external part of some nanoparticles was also thought to specifically target cancer cells, which are avid of lipids necessary for their high-level metabolism and high proliferative turnover rate. Thus, through this expedient, such ω-3 PUFA-coated nanoparticles could be preferentially taken up by cancer cells.
Notably, the inclusion of these fatty acids – showing both anti-inflammatory and anti-neoplastic activities – in nanoformulations could be significant in making them more bioavailable and to better deliver them to cancer tissues. Therefore, ω-3 PUFAs could better inhibit cancer cell growth, but simultaneously decrease the adverse inflammatory effects induced by the currently used conventional and innovative antineoplastic agents and, also, reduce the cachexia associated with the advanced cancer stages.
It is worth observing that ALA becomes more bioactive when enclosed in some of the nanoformulations analyzed in this review. This happened since ALA in this form was more efficiently delivered to the target cells and converted into its more bioactive products (EPA and DHA). This implies that these ALA-containing nanoformulations could allow to easily enrich our tissues with EPA and DHA, which, unlike ALA, largely found in vegetables and nuts, are mainly obtained from fish that indeed are not considered a sustainable food source for the entire world population.
Altogether, the reports analyzed univocally suggest that multiple potential advantages could be obtained with the administration of nanoformulations containing ω-3 PUFAs for the prevention and cure of both CVDs and cancer. In particular, the inclusion in nanoparticles has been often designed to offer protection to these quite unstable compounds from oxidative insults and to increase and enhance their delivery to the sites of the pathological lesions, resulting in an increased efficacy. Moreover, the safety of ω-3 PUFAs at the doses generally used in human studies,Citation124 their inexpensiveness, and the results so far obtained made them optimal candidates to be enclosed in nanoparticles in combination with other pharmacological agents already used in the clinical setting. The potential of these therapeutic combinations is to deliver all the agents enclosed successfully, increase their activity, and make the therapy more effective.
Acknowledgments
This work was funded in part by grants Linea D1 2016, Linea D.3.2 2013, and D.3.2 2015 to GC from Università Cattolica del S Cuore, Rome, Italy, within its program of promotion and diffusion of scientific research.
Disclosure
The authors report no conflicts of interest in this work.
References
- CalvielloGSuHMWeylandtKHFasanoESeriniSCittadiniAExperimental evidence of ω-3 polyunsaturated fatty acid modulation of inflammatory cytokines and bioactive lipid mediators: their potential role in inflammatory, neurodegenerative, and neoplastic diseasesBiomed Res Int2013201374317110.1155/2013/74317123691510
- MarventanoSKolaczPCastellanoSA review of recent evidence in human studies of n-3 and n-6 PUFA intake on cardiovascular disease, cancer, and depressive disorders: does the ratio really matter?Int J Food Sci Nutr201566661162210.3109/09637486.2015.107779026307560
- DyallSCLong-chain omega-3 fatty acids and the brain: a review of the independent and shared effects of EPA, DPA and DHAFront Aging Neurosci201575210.3389/fnagi.2015.0005225954194
- KitessaSMAbeywardenaMWijesunderaCNicholsPDDHA-containing oilseed: a timely solution for the sustainability issues surrounding fish oil sources of the health-benefitting long-chain omega-3 oilsNutrients2014652035205810.3390/nu605203524858407
- Bushkin-BedientSCarpenterDOBenefits versus risks associated with consumption of fish and other seafoodRev Environ Health201025316119121038755
- NicholsPDMcManusAKrailKSinclairAJMillerMRecent advances in omega-3: health benefits, sources, products and bioavailabilityNutrients2014693727373310.3390/nu609372725255830
- WellsMLPotinPCraigieJSAlgae as nutritional and functional food sources: revisiting our understandingJ Appl Phycol201729294998210.1007/s10811-016-0974-528458464
- MillerMRNicholsPDCarterCGn-3 Oil sources for use in aquaculture–alternatives to the unsustainable harvest of wild fishNutr Res Rev2008212859610.1017/S095442240810241419087364
- Lenihan-GeelsGBishopKSFergusonLRAlternative sources of omega-3 fats: can we find a sustainable substitute for fish?Nutrients2013541301131510.3390/nu504130123598439
- MartinsDACustódioLBarreiraLAlternative sources of n-3 long-chain polyunsaturated fatty acids in marine microalgaeMar Drugs20131172259228110.3390/md1107225923807546
- SpragueMBetancorMBTocherDRMicrobial and genetically engineered oils as replacements for fish oil in aquaculture feedsBiotechnol Lett201739111599160910.1007/s10529-017-2402-628721583
- ReynoldsLMulikRSWenXDilipACorbinIRLow-density lipoprotein-mediated delivery of docosahexaenoic acid selectively kills murine liver cancer cellsNanomedicine (Lond)20149142123214110.2217/nnm.13.18724397600
- KannoSKurauchiKTomizawaAYomogidaSIshikawaMAlbumin modulates docosahexaenoic acid-induced cytotoxicity in human hepa-tocellular carcinoma cell linesToxicol Lett2011200315416110.1016/j.toxlet.2010.11.00921108996
- NamaniTIshikawaTMorigakiKWaldePVesicles from docosahexaenoic acidColloids Surf B Biointerfaces200754111812310.1016/j.colsurfb.2006.05.02216829059
- CalderPCThe role of marine omega-3 (n-3) fatty acids in inflammatory processes, atherosclerosis and plaque stabilityMol Nutr Food Res20125671073108010.1002/mnfr.20110071022760980
- InnesJKCalderPCThe differential effects of eicosapentaenoic acid and docosahexaenoic acid on cardiometabolic risk factors: a systematic reviewInt J Mol Sci2018192E53210.3390/ijms1902053229425187
- DeshpandeDJaneroDRAmijiMEngineering of an ω-3 polyunsaturated fatty acid-containing nanoemulsion system for combination C6-ceramide and 17β-estradiol delivery and bioactivity in human vascular endothelial and smooth muscle cellsNanomedicine20139788589410.1016/j.nano.2013.02.00723473744
- DeshpandeDKethireddySJaneroDRAmijiMMTherapeutic effi-cacy of an ω-3-fatty acid-containing 17-β estradiol nano-delivery system against experimental atherosclerosisPLoS One2016112e014733710.1371/journal.pone.014733726840601
- SerhanCNPro-resolving lipid mediators are leads for resolution physiologyNature201451075039210110.1038/nature1347924899309
- SansburyBESpiteMResolution of acute inflammation and the role of resolvins in immunity, thrombosis, and vascular biologyCirc Res2016119111313010.1161/CIRCRESAHA.116.30730827340271
- CongBXuYShengHCardioprotection of 17β-estradiol against hypoxia/reoxygenation in cardiomyocytes is partly through up-regulation of CRH receptor type 2Mol Cell Endocrinol20143821172510.1016/j.mce.2013.09.00224035863
- BorodziczSCzarzastaKKuchMCudnoch-JedrzejewskaASphingolipids in cardiovascular diseases and metabolic disordersLipids Health Dis2015145510.1186/s12944-015-0053-y26076974
- BoweyKTanguayJFTabrizianM2-Dioleoyl-sn-glycero-3-phosphocholine-based nanoliposomes as an effective delivery platform for 17β-estradiolEur J Pharm Biopharm201486336937510.1016/j.ejpb.2013.10.00624161656
- StoverTCSharmaARobertsonGPKesterMSystemic delivery of liposomal short-chain ceramide limits solid tumor growth in murine models of breast adenocarcinomaClin Cancer Res20051193465347410.1158/1078-0432.CCR-04-177015867249
- BourassaPAMilosPMGaynorBJBreslowJLAielloRJEstrogen reduces atherosclerotic lesion development in apolipoprotein E-deficient miceProc Natl Acad Sci U S A19969310022100278816744
- SuHLiuRChangMHuangJJinQWangXEffect of dietary alpha-linolenic acid on blood inflammatory markers: a systematic review and meta-analysis of randomized controlled trialsEur J Nutr201857387789110.1007/s00394-017-1386-228275869
- OuWMulikRSAnwarAMcDonaldJGHeXCorbinIRLow-density lipoprotein docosahexaenoic acid nanoparticles induce ferroptotic cell death in hepatocellular carcinomaFree Radic Biol Med201711259760710.1016/j.freeradbiomed.2017.09.00228893626
- MossLRMulikRSVan TreurenTKimSYCorbinIRInvestigation into the distinct subcellular effects of docosahexaenoic acid loaded low-density lipoprotein nanoparticles in normal and malignant murine liver cellsBiochim Biophys Acta2016186011 Pt A2363237610.1016/j.bbagen.2016.07.00427418237
- WenXReynoldsLMulikRSHepatic arterial infusion of low-density lipoprotein docosahexaenoic acid nanoparticles selectively disrupts redox balance in hepatoma cells and reduces growth of ortho-topic liver tumors in ratsGastroenterology2016150248849810.1053/j.gastro.2015.10.00826484708
- MulikRSBingCLadouceur-WodzakMMunaweeraIChopraRCorbinIRLocalized delivery of low-density lipoprotein docosahexaenoic acid nanoparticles to the rat brain using focused ultrasoundBiomaterials20168325726810.1016/j.biomaterials.2016.01.02126790145
- GottoAMJrPownallHJHavelRJIntroduction to the plasma lipoproteinsMethods Enzymol19861283413523141
- FavreGTargeting of tumor cells by low density lipoproteins: principle and use of ellipticin derivativesC R Seances Soc Biol Fil19921861–273871450992
- GalDOhashiMMacDonaldPCBuchsbaumHJSimpsonERLow-density lipoprotein as a potential vehicle for chemotherapeutic agents and radionucleotides in the management of gynecologic neoplasmsAm J Obstet Gynecol198113988778857223790
- HoYKSmithRGBrownMSGoldsteinJLLow-density lipoprotein (LDL) receptor activity in human acute myelogenous leukemia cellsBlood197852610991114214187
- CasarilMCorsoFBassiADecreased activity of scavenger enzymes in human hepatocellular carcinoma, but not in liver metastasesInt J Clin Lab Res199424294977919435
- YangLYChenWLLinJWDifferential expression of antioxidant enzymes in various hepatocellular carcinoma cell linesJ Cell Biochem20059662263110.1002/jcb.2054116052474
- MerendinoNLoppiBD’AquinoMDocosahexaenoic acid induces apoptosis in the human PaCa-44 pancreatic cancer cell line by active reduced glutathione extrusion and lipid peroxidationNutr Cancer20055222523310.1207/s15327914nc5202_1216201853
- BertrandRLIron accumulation, glutathione depletion, and lipid peroxidation must occur simultaneously during ferroptosis and are mutually amplifying eventsMed Hypotheses2017101697410.1016/j.mehy.2017.02.01728351498
- SeriniSPiccioniEMerendinoNCalvielloGDietary polyunsaturated fatty acids as inducers of apoptosis: implications for cancerApoptosis200914213515210.1007/s10495-008-0298-219130233
- SchwenkRWHollowayGPLuikenJJFPBonenAGlatzJFCFatty acid transport across the cell membrane: regulation by fatty acid transportersProstaglandins Leukot Essent Fatty Acids2010824–614915410.1016/j.plefa.2010.02.02920206486
- KhanAAAlanaziAMJabeenMHassanIBhatMATargeted nano-delivery of novel omega-3 conjugate against hepatocellular carcinoma: regulating COX-2/bcl-2 expression in an animal modelBiomed Pharmacother20168139440110.1016/j.biopha.2016.04.03327261618
- KhanAAAlamMTufailSMustafaJOwaisMSynthesis and characterization of novel PUFA esters exhibiting potential anticancer activities: an in vitro studyEur J Med Chem201146104878488610.1016/j.ejmech.2011.07.04421868138
- AltenburgJDHarveyKAMcCraySXuZSiddiquiRAA novel 2,6-diisopropylphenyl-docosahexaenoamide conjugate induces apoptosis in T cell acute lymphoblastic leukemia cell linesBiochem Biophys Res Commun2011411242743210.1016/j.bbrc.2011.06.17221749860
- SiemionIZKluczykATuftsin: on the 30-year anniversary of Victor Najjar’s discoveryPeptides199920564567410465518
- KhanMAAljarbouAKhanAOwaisMImmune stimulating and therapeutic potential of tuftsin-incorporated nystatin liposomes against Cryptococcus neoformans in leukopenic BALB/C miceFEMS Immunol Med Microbiol2012661889710.1111/j.1574-695X.2012.00992.x22612788
- KhanAKhanAADwivediVAhmadMGHakeemSOwaisMTuftsin augments antitumor efficacy of liposomized etoposide against fibrosarcoma in Swiss albino miceMol Med2007135–626627610.2119/2007-00018.Khan17622310
- BaiKBLángOOrbánEDesign, synthesis, and in vitro activity of novel drug delivery systems containing tuftsin derivatives and methotrexateBioconjug Chem200819112260226910.1021/bc800115w18959436
- HalderRCAlmasiASagongBLeungJJewettAFialaMCurcuminoids and ω-3 fatty acids with anti-oxidants potentiate cytotoxicity of natural killer cells against pancreatic ductal adenocarcinoma cells and inhibit interferon γ productionFront Physiol2015612910.3389/fphys.2015.0009826052286
- SongMNishiharaRCaoYMarine ω-3 polyunsaturated fatty acid intake and risk of colorectal cancer characterized by tumor-infiltrating T cellsJAMA Oncol2016291197120610.1001/jamaoncol.2016.060527148825
- RoyJOliveiraLTOgerCPolymeric nanocapsules prevent oxidation of core-loaded molecules: evidence based on the effects of docosahexaenoic acid and neuroprostane on breast cancer cells proliferationJ Exp Clin Cancer Res20153415510.1186/s13046-015-0273-z26689718
- LeiteEAGrabe-GuimarãesAGuimarãesHNMachado-CoelhoGLLBarrattGMosqueiraVCFCardiotoxicity reduction induced by halofantrine entrapped in nanocapsule devicesLife Sci200780141327133410.1016/j.lfs.2006.12.01917303179
- SchleyPDJijonHBRobinsonLEFieldCJMechanisms of omega-3 fatty acid-induced growth inhibition in MDA-MB-231 human breast cancer cellsBreast Cancer Res Treat200592218719510.1007/s10549-005-2415-z15986129
- RogersKRKikawaKDMouradianMDocosahexaenoic acid alters epidermal growth factor receptor-related signaling by disrupting its lipid raft associationCarcinogenesis20103191523153010.1093/carcin/bgq11120525880
- ChauvinLGoupilleCBlancCLong chain n-3 polyunsaturated fatty acids increase the efficacy of docetaxel in mammary cancer cells by downregulating Akt and PKCε/δ-induced ERK pathwaysBiochim Biophys Acta20161861438039010.1016/j.bbalip.2016.01.01226821209
- SatoSBParkJKawamotoJSatoSKuriharaTInhibition of constitutive Akt (PKB) phosphorylation by docosahexaenoic acid in the human breast cancer cell line MDA-MB-453Biochim Biophys Acta20131831230631310.1016/j.bbalip.2012.10.00423085420
- LeeEJYunUJKooKHDown-regulation of lipid raft-associated oncoproteins via cholesterol-dependent lipid raft internalization in docosahexaenoic acid-induced apoptosisBiochim Biophys Acta20141841119020310.1016/j.bbalip.2013.10.00624120917
- SeriniSCalvielloGModulation of Ras/ERK and phosphoinositide signaling by Long-Chain n-3 PUFA in breast cancer and their potential complementary role in combination with targeted drugsNutrients201793E18510.3390/nu903018528241486
- AlaargAJordanNYVerhoefJJMetselaarJMStormGKokRJDocosahexaenoic acid liposomes for targeting chronic inflammatory diseases and cancer: an in vitro assessmentInt J Nanomedicine2016115027504010.2147/IJN.S11599527785012
- GabizonAShmeedaHBarenholzYPharmacokinetics of PEGylated liposomal doxorubicin: review of animal and human studiesClin Pharmacokinet200342541943610.2165/00003088-200342050-0000212739982
- SkibinskiCGDasAChenKMA novel biologically active acid stable liposomal formulation of docosahexaenoic acid in human breast cancer cell linesChem Biol Interact20162521810.1016/j.cbi.2016.03.03527041074
- ClarkASWestKStreicherSDennisPAConstitutive and inducible Akt activity promotes resistance to chemotherapy, trastuzumab, or tamoxifen in breast cancer cellsMol Cancer Ther20021970771712479367
- SeriniSOttes VasconcelosRFasanoECalvielloGHow plausible is the use of dietary n-3 PUFA in the adjuvant therapy of cancer?Nutr Res Rev201629110212510.1017/S095442241600004427172872
- VaidyaTStraubingerRMAit-OudhiaSDevelopment and evaluation of tri-functional immunoliposomes for the treatment of HER2 positive breast cancerPharm Res20183559510.1007/s11095-018-2365-x29536232
- MussiSVSilvaRCOliveiraMCLucciCMAzevedoRBFerreiraLANew approach to improve encapsulation and antitumor activity of doxorubicin loaded in solid lipid nanoparticlesEur J Pharm Sci2013481–228229010.1016/j.ejps.2012.10.02523178339
- ZuYHuYYuXJiangSDocetaxel-loaded bovine serum albumin nanoparticles conjugated docosahexaenoic acid for inhibiting lung cancer metastasis to boneAnticancer Agents Med Chem201717454255110.2174/187152061666616081714365627539313
- TrédanOGalmariniCMPatelKTannockIFDrug resistance and the solid tumor microenvironmentJ Natl Cancer Inst200799191441145410.1093/jnci/djm13517895480
- PrimeauAJRendonAHedleyDLilgeLTannockIFThe distribution of the anticancer drug Doxorubicin in relation to blood vessels in solid tumorsClin Cancer Res20051124 Pt 18782878810.1158/1078-0432.CCR-05-166416361566
- Johnson-ArborKDubeyRStatPearls [Internet]Treasure Island (FL)StatPearls Publishing2017 2018
- SubediRKKangKWChoiHKPreparation and characterization of solid lipid nanoparticles loaded with doxorubicinEur J Pharm Sci2009373–450851310.1016/j.ejps.2009.04.00819406231
- LiYWangJWientjesMGAuJLDelivery of nanomedicines to extracellular and intracellular compartments of a solid tumorAdv Drug Deliv Rev2012641293910.1016/j.addr.2011.04.00621569804
- MaedaHBharateGYDaruwallaJPolymeric drugs for efficient tumor-targeted drug delivery based on EPR-effectEur J Pharm Biopharm200971340941910.1016/j.ejpb.2008.11.01019070661
- TorchilinVTumor delivery of macromolecular drugs based on the EPR effectAdv Drug Deliv Rev201163313113510.1016/j.addr.2010.03.01120304019
- ZhangYSunTJiangCBiomacromolecules as carriers in drug delivery and tissue engineeringActa Pharm Sin B201881345010.1016/j.apsb.2017.11.00529872621
- AlkenSKellyCMBenefit risk assessment and update on the use of docetaxel in the management of breast cancerCancer Manag Res2013535736510.2147/CMAR.S4932124143122
- HeXWangJLiYEfficacy and safety of docetaxel for advanced non-small-cell lung cancer: a meta-analysis of Phase III randomized controlled trialsOnco Targets Ther201582023203110.2147/OTT.S8564826346649
- HennenfentKLGovindanRNovel formulations of taxanes: a review. Old wine in a new bottle?Ann Oncol200617573574910.1093/annonc/mdj10016364960
- LiSQinJTianCThe targeting mechanism of DHA ligand and its conjugate with Gemcitabine for the enhanced tumor therapyOncotarget20145113622363510.18632/oncotarget.196925004114
- ZhangJYeEWangJVariations of cellular membrane phospholipids with genesis of gastric cancerMod Oncol20101808550857
- StubbsCDSmithADThe modification of mammalian membrane polyunsaturated fatty acid composition in relation to membrane fluidity and functionBiochim Biophys Acta19847791891376229284
- MarienEMeisterMMuleyTNon-small cell lung cancer is characterized by dramatic changes in phospholipid profilesInt J Cancer201513771539154810.1002/ijc.2951725784292
- WolffACDonehowerRCCarducciMKPhase I study of docosahexaenoic acid-paclitaxel: a taxane-fatty acid conjugate with a unique pharmacology and toxicity profileClin Cancer Res2003910 Pt 13589359714506145
- FracassoPMPicusJWildiJDPhase 1 and pharmacokinetic study of weekly docosahexaenoic acid paclitaxel, Taxoprexin, in resistant solid tumor malignanciesCancer Chemother Pharmacol200963345145810.1007/s00280-008-0756-018414864
- PayneMEllisPDunlopDDHA-paclitaxel (Taxoprexin) as first-line treatment in patients with stage IIIB or IV non-small cell lung cancer: report of a phase II openlabel multicenter trialJ Thorac Oncol20061998499017409983
- HomsiJBedikianAYKimKBPhase 2 open-label study of weekly docosahexaenoic acid-paclitaxel in cutaneous and mucosal metastatic melanoma patientsMelanoma Res200919423824210.1097/CMR.0b013e32832a1e2f19521262
- BedikianAYDe ContiRCConryRPhase 3 study of docosahexaenoic acid-paclitaxel versus dacarbazine in patients with metastatic malignant melanomaAnn Oncol201122478779310.1093/annonc/mdq43820855467
- HomsiJBedikianAYPapadopoulosNEPhase 2 open-label study of weekly docosahexaenoic acid paclitaxel in patients with metastatic uveal melanomaMelanoma Res201020650751010.1097/CMR.0b013e3283403ce920881508
- AhmadGEl SaddaRBotchkinaGOjimaIEganJAmijiMNanoemulsion formulation of a novel taxoid DHA-SBT-1214 inhibits prostate cancer stem cell-induced tumor growthCancer Lett2017406718010.1016/j.canlet.2017.08.00428803993
- AhmadGGattaceccaFEl SaddaRBiodistribution and phar-macokinetic evaluations of a novel taxoid DHA-SBT-1214 in an oil-in-water nanoemulsion formulation in naïve and tumor-bearing micePharm Res20183549110.1007/s11095-018-2349-x29520477
- VredenburgMROjimaIVeithJEffects of orally active taxanes on P-glycoprotein modulation and colon and breast carcinoma drug resistanceJ Natl Cancer Inst200193161234124511504769
- Marín-AguileraMCodony-ServatJKalkoSGIdentification of docetaxel resistance genes in castration-resistant prostate cancerMol Cancer Ther201211232933910.1158/1535-7163.MCT-11-028922027694
- CassanoRMellaceSMarrelliMConfortiFTrombinoSα-Tocopheryl linolenate solid lipid nanoparticles for the encapsulation, protection, and release of the omega-3 polyunsaturated fatty acid: in vitro anti-melanoma activity evaluationColloids Surf B Biointerfaces201715112813310.1016/j.colsurfb.2016.11.04327988473
- SeriniSCassanoRCorsettoPARizzoAMCalvielloGTrombinoSOmega-3 PUFA loaded in resveratrol-based solid lipid nanoparticles: physicochemical properties and antineoplastic activities in human colorectal cancer cells in vitroInt J Mol Sci2018192E58610.3390/ijms1902058629462928
- ReyaTMorrisonSJClarkeMFWeissmanILStem cells, cancer, and cancer stem cellsNature2001414685910511110.1038/3510216711689955
- WichaMSLiuSDontuGCancer stem cells: an old idea – a paradigm shiftCancer Res20066641883189010.1158/0008-5472.CAN-05-315316488983
- ClarkeMFFullerMStem cells and cancer: two faces of eveCell200612461111111510.1016/j.cell.2006.03.01116564000
- CalvielloGSeriniSPiccioniEn-3 polyunsaturated fatty acids and the prevention of colorectal cancer: molecular mechanisms involvedCurr Med Chem200714293059306918220742
- FasanoESeriniSPiccioniEDHA induces apoptosis by altering the expression and cellular location of GRP78 in colon cancer cell linesBiochim Biophys Acta20121822111762177210.1016/j.bbadis.2012.08.00322898250
- SeriniSOttes VasconcelosRFasanoECalvielloGEpigenetic regulation of gene expression and M2 macrophage polarization as new potential omega-3 polyunsaturated fatty acid targets in colon inflammation and cancerExpert Opin Ther Targets201620784385810.1517/14728222.2016.113908526781478
- SeriniSFasanoEPiccioniEDHA induces apoptosis and differentiation in human melanoma cells in vitro: involvement of HuR-mediated COX-2 mRNA stabilization and β-catenin nuclear translocationCarcinogenesis201233116417310.1093/carcin/bgr24022045024
- SeriniSFasanoECellenoLCittadiniACalvielloGPotential of long-chain n-3 polyunsaturated fatty acids in melanoma preventionNutr Rev201472425526610.1111/nure.1209324665956
- SeriniSZinziAOttes VasconcelosRRole of β-catenin signaling in the anti-invasive effect of the omega-3 fatty acid DHA in human melanoma cellsJ Dermatol Sci201684214915910.1016/j.jdermsci.2016.06.01027600927
- MensinkRPEffects of stearic acid on plasma lipid and lipoproteins in humansLipids200540121201120516477803
- CroweFLAllenNEApplebyPNFatty acid composition of plasma phospholipids and risk of prostate cancer in a case-control analysis nested within the European prospective investigation into cancer and nutritionAm J Clin Nutr20088851353136310.3945/ajcn.2008.2636918996872
- LiuYZWuKHuangJThe PTEN/PI3K/Akt and Wnt/β-catenin signaling pathways are involved in the inhibitory effect of resveratrol on human colon cancer cell proliferationInt J Oncol201445110411210.3892/ijo.2014.239224756222
- BraskyTMDarkeAKSongXPlasma phospholipid fatty acids and prostate cancer risk in the SELECT trialJ Natl Cancer Inst2013105151132114110.1093/jnci/djt17423843441
- HullMASandellACMontgomeryAAA randomized controlled trial of eicosapentaenoic acid and/or aspirin for colorectal adenoma prevention during colonoscopic surveillance in the NHS Bowel Cancer Screening Programme (The seAFOod Polyp Prevention Trial): study protocol for a randomized controlled trialTrials20131423710.1186/1745-6215-14-5923895505
- PiazziGD’ArgenioGProssomaritiAEicosapentaenoic acid free fatty acid prevents and suppresses colonic neoplasia in colitis-associated colorectal cancer acting on Notch signaling and gut microbiotaInt J Cancer201413592004201310.1002/ijc.2885324676631
- NakanishiMHanleyMPZhaRA novel bioactive derivative of eicosapentaenoic acid (EPA) suppresses intestinal tumor development in ApcΔ14/+ miceCarcinogenesis201839342943810.1093/carcin/bgx13629206907
- RoseDPConnollyJMOmega-3 fatty acids as cancer chemopreventive agentsPharmacol Ther199983321724410576293
- RoynetteCECalderPCDupertuisYMPichardCn-3 polyunsaturated fatty acids and colon cancer preventionClin Nutr200423213915110.1016/j.clnu.2003.07.00515030953
- DupertuisYMMeguidMMPichardCColon cancer therapy: new perspectives of nutritional manipulations using polyunsaturated fatty acidsCurr Opin Clin Nutr Metab Care200710442743210.1097/MCO.0b013e3281e2c9d417563460
- LeeJYSimTBLeeJENaHKChemopreventive and chemotherapeutic effects of fish oil derived omega-3 polyunsaturated fatty acids on colon carcinogenesisClin Nutr Res20176314716010.7762/cnr.2017.6.3.14728770178
- NoriegaBSSanchez-GonzalezMASalyakinaDCoffmanJUnderstanding the impact of omega-3 rich diet on the gut microbiotaCase Rep Med20162016308930310.1155/2016/389830727065349
- NappiABerrettaMRomanoCMetastatic colorectal cancer: role of target therapies and future perspectivesCurr Cancer Drug Targets201818542142910.2174/15680096176661702009514328183254
- NikpoorARTavakkol-AfshariJSadriKJalaliSAJaafariMRImproved tumor accumulation and therapeutic efficacy of CTLA-4-blocking antibody using liposome-encapsulated antibody: in vitro and in vivo studiesNanomedicine20171382671268210.1016/j.nano.2017.08.01028847682
- CoutzacCAdamJSoularueEColon immune-related adverse events: anti-CTLA-4 and anti-PD-1 blockade induce distinct immunopathological entitiesJ Crohns Colitis201711101238124610.1093/ecco-jcc/jjx08128967957
- ZárateREl Jaber-VazdekisNTejeraNPérezJARodríguezCSignificance of long chain polyunsaturated fatty acids in human healthClin Transl Med2017612510.1186/s40169-017-0153-628752333
- BowenKJHarrisWSKris-EthertonPMOmega-3 fatty acids and cardiovascular disease: are there benefits?Curr Treat Options Cardiovasc Med201618116910.1007/s11936-016-0487-127747477
- MansonJECookNRLeeIMMarine n-3 fatty acids and prevention of cardiovascular disease and cancerN Engl J Med20193801233210.1056/NEJMoa181140330415637
- SeriniSCalvielloGLong-chain omega-3 fatty acids and cancer: any cause for concern?Curr Opin Clin Nutr Metab Care2018212838910.1097/MCO.000000000000043929232261
- BhattDLStegPGMillerMCardiovascular risk reduction with icosapent ethyl for hypertriglyceridemiaN Engl J Med20193801112210.1056/NEJMoa181279230415628
- MozaffarianDWuJHOmega-3 fatty acids and cardiovascular disease: effects on risk factors, molecular pathways, and clinical eventsJ Am Coll Cardiol2011582047206710.1016/j.jacc.2011.06.06322051327
- HarrisWSvon SchackyCThe omega-3 index: a new risk factor for death from coronary heart disease?Prev Med200439121222010.1016/j.ypmed.2004.02.03015208005
- FasanoESeriniSCittadiniACalvielloGLong-chain n-3 PUFA against breast and prostate cancer: which are the appropriate doses for intervention studies in animals and humans?Crit Rev Food Sci Nutr201757112245226210.1080/10408398.2013.85006025897862
- AungTHalseyJKromhoutDAssociations of omega-3 fatty acid supplement use with cardiovascular disease risks: meta-analysis of 10 trials involving 77 917 individualsJAMA Cardiol2018322523410.1001/jamacardio.2017.520529387889
- ZhangYFGaoHFHouAJZhouYHEffect of omega-3 fatty acid supplementation on cancer incidence, non-vascular death, and total mortality: a meta-analysis of randomized controlled trialsBMC Public Health20141420410.1186/1471-2458-14-20424568238
- AntiMArmelaoFMarraGEffects of different doses of fish oil on rectal cell proliferation in patients with sporadic colonic adenomasGastroenterology19941076170917187958682
- BougnouxPHajjajiNFerrassonMNGiraudeauBCouetCLe FlochOImproving outcome of chemotherapy of metastatic breast cancer by docosahexaenoic acid: a phase II trialBr J Cancer2009101121978198510.1038/sj.bjc.660544119920822
- WestNJClarkSKPhillipsRKEicosapentaenoic acid reduces rectal polyp number and size in familial adenomatous polyposisGut201059791892510.1136/gut.2009.20064220348368
- DyerbergJBangHOHaemostatic function and platelet polyunsaturated fatty acids in EskimosLancet1979243343589498
- BragaMLjungqvistOSoetersPFearonKWeimannABozzettiFESPEN Guidelines on Parenteral Nutrition: SurgeryClin Nutr20092837838619464088
- JeansenSWitkampRFGarthoffJAvan HelvoortACalderPCFish oil LC-PUFAs do not affect blood coagulation parameters and bleeding manifestations: analysis of 8 clinical studies with selected patient groups on omega-3-enriched medical nutritionClin Nutr20183794895710.1016/j.clnu.2017.03.02728427779
- D’EliseoDVelottiFOmega-3 fatty acids and cancer cell cytotoxicity: implications for multi-targeted cancer therapyJ Clin Med201652E1510.3390/jcm502001526821053
- DiakosCICharlesKAMcMillanDCClarkeSJCancer-related inflammation and treatment effectivenessLancet Oncol20141511e493e50310.1016/S1470-2045(14)70263-325281468
- PawelecGGoldeckDDerhovanessianEInflammation, ageing and chronic diseaseCurr Opin Immunol201429232810.1016/j.coi.2014.03.00724762450
- IlagLLAre long-chain polyunsaturated fatty acids the link between the immune system and the microbiome towards modulating cancer?Medicines (Basel)201853E10210.3390/medicines503010230201858
- LiangPHenningSMSchokrpurSEffect of dietary omega-3 fatty acids on tumor-associated macrophages and Prostate Cancer ProgressionProstate201676141293130210.1002/pros.2321827338236
- GevariyaNBesançonMRobitailleKOmega-3 fatty acids decrease prostate cancer progression associated with an anti-tumor immune response in eugonadal and castrated miceProstate201979192010.1002/pros.2370630073695
- KhadgeSSharpJGMcGuireTRImmune regulation and anti-cancer activity by lipid inflammatory mediatorsInt Immunopharmacol20186558059210.1016/j.intimp.2018.10.02630447537
- GuesdonWKosarajuRBrophyPEffects of fish oils on ex vivo B-cell responses of obese subjects upon BCR/TLR stimulation: A pilot studyJ Nutr Biochem201853728010.1016/j.jnutbio.2017.10.00929195133
- HouTYMcMurrayDNChapkinRSOmega-3 fatty acids, lipid rafts, and T cell signalingEur J Pharmacol20167852910.1016/j.ejphar.2015.03.09126001374
- WachtelNRohwerNPietznerALoewAWeylandtKHOmega-3 fatty acid supplementation-a possible dietary adjunct to enhance immune therapy in cancer?J Cell Biotech201814[1–2]8388
- CostantiniLMolinariRFarinonBMerendinoNImpact of omega-3 fatty acids on the gut microbiotaInt J Mol Sci20171812E264510.3390/ijms1812264529215589
- HondaKLittmanDRThe microbiota in adaptive immune homeostasis and diseaseNature2016535758410.1038/nature1884827383982
- SanzYDe PalmaGGut microbiota and probiotics in modulation of epithelium and gut-associated lymphoid tissue functionInt Rev Immunol20092839741310.3109/0883018090321561319954356