
Abstract
Exosomes, nanoscale vesicles with a diameter of 30 to 150 nm, are composed of a lipid bilayer, protein, and genetic material. Exosomes are secreted by virtually all types of cells in the human body. They have key functions in cell-to-cell communication, immune regulation, inflammatory response, and neovascularization. Mounting evidence indicates that exosomes play an important role in various diseases, such as cancer, cardiovascular diseases, and brain diseases; however, the role that exosomes play in eye diseases has not yet been rigorously studied. This review covers current exosome research as it relates to ocular diseases including diabetic retinopathy, age-related macular degeneration, autoimmune uveitis, glaucoma, traumatic optic neuropathies, corneal diseases, retinopathy of prematurity, and uveal melanoma. In addition, we discuss recent advances in the biological functions of exosomes, focusing on the toxicity of exosomes and the use of exosomes as biomarkers and drug delivery vesicles. Finally, we summarize the primary considerations and challenges to be taken into account for the effective applications of exosomes.
Introduction
Extracellular vesicles (EVs) are an umbrella term for all lipid bilayer-encased extracellular structures, which are typically divided into three subtypes: exosomes (~30–150 nm in diameter), microvesicles (~100–1000 nm in diameter), and apoptotic bodies (~100–5000 nm in diameter), based on their formation mode, particle size, and functional properties.Citation1,Citation2 Exosomes are the smallest subtype of EVs.Citation2 The term “exosome” was first used in the early 1980s to describe small vesicles (~50 nm) of endosomal origin released during the maturation of sheep.Citation3–Citation5 Subsequently, it was shown that these nanovesicles could be secreted by numerous cells such as B cells, T cells, cancer cells, endothelial cells, and mesenchymal stem cells.Citation6 Exosomes, which are nanosized membrane-enclosed vesicles of 30 to 150 nm that are shed by various cell types, exist extensively in a variety of bodily fluids including tears, aqueous humor, vitreous humor, blood, urine, semen, vaginal fluid, breast milk, cerebrospinal fluid, ascites, lymph, bile, amniotic fluid, and saliva.Citation7–Citation10 They are formed as intraluminal vesicles (ILVs) after fusion of a multivesicular body (MVB) with the plasma membrane.Citation11,Citation12 A MVB is an intermediate endosomal compartment that is filled with ILVs.Citation13 Diverse bioactive molecules including lipids, proteins, and nucleic acids are the main functional components of exosomes, which are encapsulated in a lipid bilayer membrane.Citation14,Citation15 Exosomes are a promising field of cytological and biomedical research, attracting intense interest because of the biological functions and broad contribution of exosomes to a range of diseases. First, exosomes that carry surface molecules partake in intercellular communication by activating cellular signaling through receptor-ligand interactions with target cells.16 Second, it has been suggested that exosomes are involved in the progression of multiple diseases through a mechanism by which they change the physiological state of target cells via fusing with the target cell’s membrane and transferring their cargo (protein, lipids, mRNA, microRNA, and DNA) to that cell.Citation16–Citation19 Third, exosomes have great potential as biomarkers for diagnosing diseases.Citation20–Citation22 Finally, exosomes, in the form of drug delivery vehicles, have been explored as potential therapeutic tools for a variety of currently incurable diseases.Citation9,Citation23–Citation26
Vision impairment, including blindness, is an important public health concern.Citation27,Citation28 The number of people affected by vision impairment or blindness is estimated to be 2.2 billion worldwide with middle-aged and elderly individuals being particularly affected.Citation28–Citation30 In 2019, the World Health Organization reported that the leading causes of moderate or severe vision impairment are uncorrected refractive error, cataract, diabetic retinopathy (DR), age-related macular degeneration (AMD), glaucoma, corneal opacity, and trachoma.Citation29 The cornea is the transparent front surface of the eye. Characterized by its powerful refractive ability, the cornea is a key component of the optical system.Citation31,Citation32 Thus, other diseases of the cornea are contributing factors to vision impairment. In addition to these eye diseases, uveitis, which is uncommon but affects all age, is another cause of vision impairment.Citation33 There are many drawbacks and limitations to current therapies such as surgery, intraocular injections, and eye drops that primarily rely on impeding the development of these diseases. Research into more effective alternative treatments is needed, including research into regenerative cell-based therapies.Citation34,Citation35
Exosomes are known to play an important role in many diseases, such as cancer, cardiovascular disease, and nervous system diseases. In recent years, the role of various cell-derived exosomes in diverse ophthalmic diseases has been increasingly studied. However, knowledge regarding the function of exosomes in ocular diseases remains limited. Recently, some reviews of the role of exosomes in ocular diseases have emerged. Klingeborn et alCitation10 summarized various studies on the functions of exosomes in the normal and diseased eye, including AMD, diabetic retinopathy, and uveal melanoma. In addition, how exosomes are acquired and their potential as therapeutic carriers and for disease diagnosis are discussed in their review. Mead et alCitation36 reviewed the therapeutic potential of EVs in retinal diseases, including optic nerve crush, glaucoma, retinal ischemia, retinal laser injury, autoimmune uveitis, and diabetic retinopathy. Moreover, they discussed several future research directions. More recently, one review discussed the involvement of exosomes in various pathological processes and their role in several major eye diseases, including autoimmune diseases of the eye, large area corneal damage, retinopathy of prematurity, age-related macular degeneration, diabetic retinopathy, and glaucoma.Citation37 Compared with the above reviews, we report here the latest progress on more eye diseases that involve the exosomes. We discuss the role of exosomes in diabetic retinopathy, AMD, autoimmune uveitis, glaucoma, traumatic optic neuropathies, corneal diseases, retinopathy of prematurity and uveal melanoma, and we separately discuss the relationship between exosomes and oxidative stress-induced eye diseases. In addition, we discuss the biogenesis and characteristics of exosomes in detail, as well as extraction methods for studying exosomes, and summarize this in the form of a schematic for clear understanding. Furthermore, we also review the latest research on advancements elucidating the potential applications of exosomes as biomarkers, drug delivery systems, targeting modalities, and as toxic agents.
Characteristics and Biogenesis of Exosomes
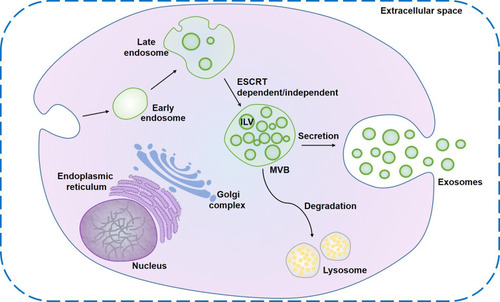
EVs are continually released by most cells into the extracellular space as vehicles to participate in diverse biological processes. EVs have two different pathways of biogenesis: 1) They are produced directly through budding from the plasma membrane; 2) The fusion between a MVB and the plasma membrane forms EVs. The latter is the production pathway of exosomes.Citation38,Citation39 As shown in , the biosynthesis of exosomes is an endosomal-dependent, progressive cytological process. ILVs accumulate in the lumen when the early endosomes formed by invagination of cell membrane mature into late endosomes. Specific-sorted ILVs, loaded with lipids, proteins, and cytosol, are obtained from inward budding of the early endosomal membrane. Late endosomes filled with ILVs are commonly known as MVBs or multivesicular endosomes (MVEs).Citation8,Citation40 There are two fates for the MVB in cells. These are the fusion of an MVB with lysosomes that degrade their cargo, and the fusion between a MVB and the plasma membrane that releases ILVs into extracellular environment as exosomes.Citation41 During the formation of exosomes, several mechanisms exist that contain the endosomal sorting complex required for transport (ESCRT)-dependent mechanisms and ESCRT-independent mechanisms (eg, tetraspanins and lipid-dependent mechanisms) to achieve specific sorting of exosomal contents.Citation38 The most well-known mechanism for forming MVB and ILVs is mediated by ESCRT, which is composed of at least twenty proteins that assemble into four complexes (involving ESCRT-0, ESCRT-I, ESCRT-II, and ESCRT-III) with several associated proteins (including VPS4, VTA1, and ALIX) that are highly conserved from yeast to mammals.Citation42,Citation43 In addition to ESCRT, some studies have reported that the biogenesis of exosomes is also regulated by other bioactive substances, such as syndecan-syntenin-ALIX,Citation44 c-Src (a membrane-associated tyrosine kinase),Citation45 small GTPase Ral,Citation46 Atg12-Atg3 (complex between two core autophagy regulators),Citation47 and mixed lineage kinase domain-like (MLKL).Citation48
Figure 1 Biogenesis of exosomes. Exosome formation originates from the early endosomes formed by plasma membrane invagination. The membranes of mature late endosomes buds inward to form ILVs and transform into MVBs. After the MVBs fuse with the plasma membrane, ILVs are released into the extracellular space to form exosomes.
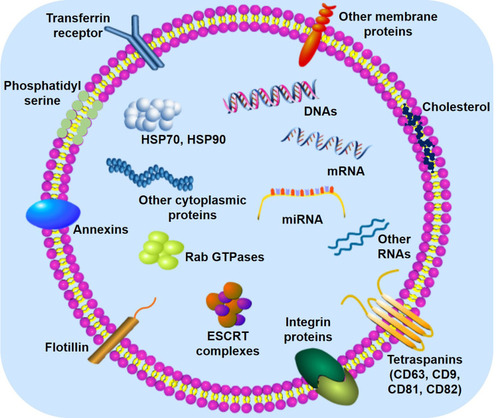
Exosomes with heterogeneous size distribution are observed to be cup-shaped and consist of numerous intracellular components including proteins, nucleic acids (eg, DNAs, mRNAs, and microRNAs), and lipids. These molecules perform different biological functions ().Citation13,Citation49 The outer membrane of exosomes are basically composed of lipids, interspersed with a variety of membrane proteins. Specific lipids that are abundant in exosomes (eg, cholesterol, phospholipids, and phosphatidylserine) have been investigated.Citation50 Undoubtedly, exosomal membrane proteins play an important role in the biological functions of exosomes, such as transmembrane proteins, as well as lipid-anchored or peripherally associated membrane proteins.Citation51 In addition to various membrane proteins, soluble proteins can exist in the exosome lumen.Citation51,Citation52 Interestingly, exosomes of different cellular origin share common protein components. These protein components include tetraspanins that contain CD63, CD9, CD81, and CD82; integrin proteins that take part in cell targeting and adhesion; membrane fusion proteins that are closely related to Rab GTPases, annexins, and flotillin; and several molecular chaperones that mediate the generation of MVBs such as heat shock proteins (HSP) 70 and 90.Citation53 Recently, the role of exosomal nucleic acids, RNAs in particular, in cell-to-cell communication and genetic exchange between cells have received widespread attention. Research has shown that mRNA and microRNA (miRNA) present in exosomes mediate their effects by transferring between cells.Citation49,Citation54 Furthermore, noncoding RNAs (ncRNAs) such as miRNA, long-ncRNA, and circular RNA are stable in exosomes and play a crucial role in the pathological process of many diseases.Citation55
Figure 2 The main components of exosomes. The lipid bilayer of exosomes contains specific lipids such as cholesterol and phosphatidylserine, which can protect the contents from degradation. In addition, various membrane proteins (eg, Tetraspanins, Annexins, Flotillin, and integrin proteins) and intracapsular proteins (eg, HSP70, HSP90, and Rab GTPases) are present in exosomes, each of which assumes different roles. ESCRT (endosomal sorting complex required for transport) is mainly involved in the generation of exosomes. DNA, mRNA, and miRNA are the key genetic materials in exosomes.
Uptake of Exosomes
It is well known that cells can communicate with each other by means of exosomes. Exosomes, released from donor cells into the extracellular environment, undergo a free-floating period in which they circulate in body fluids.Citation19 Exosomes are ingested by recipient cells in different ways to achieve intercellular communication. Cellular uptake of exosomes can be roughly divided into the following three types: (1) exosomes can fuse directly with the plasma membrane to deliver their cargo to a recipient cell. Although this process is considered to be connected with tetraspanin complexes, a detailed mechanism remains unclear.Citation19,Citation53 (2) Exosomes and target cells transmit signals through receptor-ligand interactions.Citation53,Citation56 There are soluble signals for the proteolytic cleavage of ligands on the surface of exosomes and juxtacrine signaling for the juxtaposition of ligands and receptors on the exosomes and receptor cells.Citation19 (3) Target cells uptake exosomes by diverse forms of endocytosis. These processes of endocytosis include clathrin- and caveolin-mediated endocytosis, receptor- and raft-mediated endocytosis, macropinocytosis, and phagocytosis.Citation18,Citation19,Citation57
Whether exosome selection and uptake by cells is specific is of importance for developing exosome therapies, and several studies, including those on ocular diseases, have reported the response of exosomes from different sources to different cells. For instance, a particular study showed that different receptor cells could non-selectively uptake exosomes from the same cell source, but their uptake capacity and uptake mechanisms were different.Citation58 Additionally, Ren et alCitation59 proposed a new drug delivery method for the treatment of neuronal diseases as they found that exosomes isolated from the conditioned medium of bone marrow mesenchymal stem cells (BMSCs) might use retrograde transport to peripheral nerve endings where they were ingested. Notably, some studies have demonstrated the uptake of exosomes from specific ocular cells, which provides a theoretical basis for the clinical application of exosomes in ocular diseases. Interestingly, in vitro, exosomes secreted by human corneal epithelial cells (HCECs) could be taken up by human corneal fibroblasts while HCECs can uptake exosomes released from human mesenchymal stromal cells.Citation60,Citation61 Furthermore, in vivo, BMSC-exosomes labeled with green fluorescent protein (GFP) were injected into the vitreous humor and then tracked on retinal ganglion cells (RGCs) together with the retinal nerve fiber layer (RNFL) and resident cells morphologically identified as astrocytes, indicating that these cells could incorporate exosomes.Citation62
Methods for Exosome Isolation and Purification
To date, no technique has been used to isolate exosomes to absolute purity.Citation63 Most separation protocols obtain small EVs samples only by removing large EVs without verifying their intracellular origin.Citation16,Citation64 With the popularity of the term exosome, many scientists have used exosomes directly to refer to EVs in their publications.Citation65 In their isolation protocols, therefore, isolated samples that may contain small EVs are generally referred to directly as exosomes.Citation64 Even so, a number of techniques based on the specific traits of exosomes have been developed for the isolation of exosomes from various samples.Citation66 These methods, each with its own unique advantages and disadvantages, include differential ultracentrifugation, density-gradient separation, size exclusion chromatography, immunological isolation, and precipitation. In many studies, exosomes have been better isolated by combining different separation methods.Citation63 The following are some commonly used separation approaches. Here, we chose to use the term “exosome” to denote samples obtained by various isolation methods.
Differential Ultracentrifugation
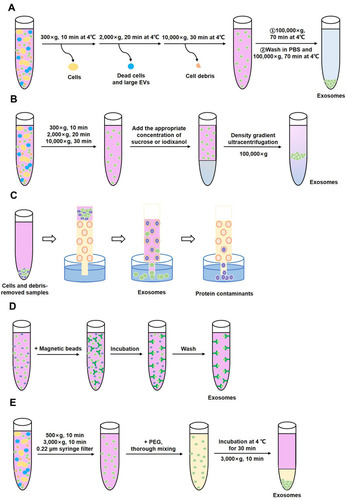
Differential ultracentrifugation is the most common method for isolating exosomes, although it is time-consuming and carries a significant risk of contamination.Citation67 It requires several steps with varying durations and rotation speeds at 4°C to obtain exosomes ().Citation68,Citation69 Under increasing centrifugal force and rotation speed, larger particles precipitate out in turn, and smaller particles are left in the supernatant. Foreign particles such as dead cells, cell debris, and apoptotic bodies are gradually removed, resulting in the separation of exosomes.Citation70
Figure 3 Major methods of exosome isolation. (A) Differential ultracentrifugation (B) Density-gradient separation (C) Size exclusion chromatography (D) Immunological isolation (E) Precipitation.
Density-Gradient Separation
Density-gradient separation can be regarded as the continuation and optimization of differential ultracentrifugation. This technique results in increased purity of exosomes by further separating particles by density and effectively removing other non-exosomes.Citation71 At present, sucroseCitation72,Citation73 and iodixanolCitation74,Citation75 are the two types of density gradient media commonly used in this method. The particular steps to be followed during density-gradient separation are dependent on the type of media chosen (). The physical integrity and bioactivity of exosomes can be better maintained by the cushioning effect of iodixanol and sucrose than by pure ultracentrifugation.
Size Exclusion Chromatography
Size exclusion chromatography (SEC) is an ideal method to separate exosomes from blood plasma. The technique is low yield but results in no obvious albumin contamination. As such, it has been applied to clinical samples.Citation76,Citation77 Unlike the protocol of differential ultracentrifugation, SEC does not rely on ultracentrifugation.Citation78 Rather, a solution containing exosomes is run through a stationary phase composed of a porous polymer. Particles larger than the pore size of the polymer are eluted first. Those smaller than the pore size are eluted later, achieving material separation ().Citation79
Immunological Isolation
Recently, highly specific immunoaffinity capture (IAC) technology has been applied to the separation of exosomes. This technique utilizes the immune affinity interactions between proteins present on the membrane of exosomes and their antibodies.Citation66 Magnetic bead-based isolation is a general method for specific capture of exosomes.Citation80 Based on the specific surface markers of exosomes, exosomes can be adsorbed and isolated after they are incubated with magnetic beads coated with corresponding antibodies ().
Precipitation
Precipitation is a simple method of isolation. Biological samples are mixed with a polymer. Exosome precipitation is then achieved at lower speeds of centrifugation ().Citation81 Polyethylene glycol (PEG) is a polymer-based precipitation reagent that is commonly used to extract exosomes from cell culture supernatant. PEG has the ability to form exosome aggregates by encapsulating a large quantity of exosomes.Citation82
Biological Functions of Exosomes
Signal Transduction
Signal transduction is the most basic and important biological function of exosomes. Early studies reported that exosomes may be cellular garbage carriers to expel metabolic waste in cells.Citation83 However, subsequent studies have demonstrated that exosomes can achieve intercellular signaling, thereby regulating various physiological and pathological processes, without the classical pathways of autocrine, paracrine, and direct intercellular contact. Exosomes released from their parent cells can act as vectors of intercellular communication, transporting their contents to adjacent cells or distant cells. They can also transmit information to multiple cells and locations.Citation84 Exosomes can serve as a signaling complex by directly binding to receptors on recipient cells and activating those cells. Furthermore, protein fragments, cleaved from the surface of exosomes by proteases, can serve as soluble ligands interacting with surface receptors on target cells to activate associated signaling cascades.Citation85 Exosomes can also transfer their cargoes to recipient cells through endocytosis, thus changing the cell status and producing functional effects.Citation86 The uptake capacity of exosomes is determined by the type of recipient cell rather than the type of donor cell, which may be closely related to organ-specific metastasis.Citation58 Many studies have shown that oxidative stress signals closely related to the processes of various major eye diseases are transferred through exosomes.Citation87,Citation88 However, further research is needed to understand the specific cellular and molecular basis for exosomal signal transduction.
Immune Regulation
The role of exosomes in immunology has been studied extensively. Early studies have found that B lymphocyte-derived exosomes execute antigen presentation through major histocompatibility complex (MHC) proteins (MHC-I and MHC-II).Citation89 Cancer cells, like immune cells, can generate immunologically active exosomes, affecting the immune regulatory mechanism. Exosomes that carry tumor antigens have anti-tumor effects. These exosomes can use CD8+ and CD4+ T cells to eliminate tumors. They can also directly inhibit the growth and development of tumors.Citation90 Kim et alCitation91 demonstrated that exosomes secreted from immature dendritic cells (DCs) take part in diminishing inflammatory and autoimmune responses via the MHC-II dependent pathway. A recent study indicated that mesenteric lymph (ML) exosomes released by gut epithelial cells after trauma/hemorrhagic shock (T/HS) may act as key mediators in the induction of post-trauma immune dysfunction, which can markedly increase the expression of MHC-II and Fas ligand. However, ML exosomes may reduce the antigen-presenting capability of DCs that can cause lymphocytes proliferation.Citation92
Although the research on the immunomodulatory effect of exosomes is mainly focused on cancer diseases, the immunomodulatory potential of exosomes in ocular diseases has also been investigated in recent years.Citation15,Citation93,Citation94 For example, retinal pigment epithelium (RPE) cells can release small EVs under different conditions to inhibit the immune response. Knickelbein et alCitation95 confirmed that EVs from non-stimulated ARPE-19 only suppress the proliferation of T cells and the immunity of monocytes, but have no effect on cell survival. However, when ARPE-19 cells are stimulated by inflammation-related cytokines, RPE-derived small EVs kill monocytes in addition to inhibiting T cell proliferation, possibly to mitigate harmful immune responses. More studies further examining the role of exosomes in immune-mediated eye diseases are needed.
Repair Regeneration
Exosomes contain specific components that play a role in repair and regeneration in a variety of disease models. For example, miRNAs may be essential mediators in the induction of tissue regeneration by the activation or inhibition of numerous downstream pathways.Citation96,Citation97 Stem cell-derived exosomes are able to induce cardiac angiogenesis and promote postinjury repair by the transfer of specific components (mRNAs, miRNAs, and proteins) into target cells or the site of injury.Citation98 In addition, exosomes from mesenchymal stem cells (MSCs) participate in the repair and regeneration of cartilage damage by promoting migration and proliferation, enhancing matrix synthesis, decreasing cell apoptosis, and regulating the immune response.Citation99 Existing studies have shown that exosomes also play a key role in liver repair and regeneration.Citation100,Citation101 Nojima et alCitation102 observed that hepatocyte exosome number increases significantly in the hepatic ischemia-reperfusion model. Hepatocyte exosomes induce hepatocyte proliferation in target cells by transporting enzymes that foster the generation sphingosine-1-phosphate (S1P). Similarly, the ability of exosomes to promote regeneration has been verified in eye diseases models. For example, exosomes within RPE cells subjected to oxidative stress have been shown to exhibit increased levels of mRNAs and proteins specific for the vascular endothelial growth factor receptor (VEGFR). The release of these exosomes results in increased expression of VEGFR-1 and VEGFR-2 and subsequent angiogenic capacity in endothelial cells.Citation103
Exosomes as Biomarkers
Exosomes have a promising potential to serve as biomarkers for a wide range of diseases. The number of exosomes and the specific bioactive substances present in exosomes have been found to vary distinctly between healthy individuals and patients with a variety of diseases.Citation104 Li et alCitation105 reported that colon cancer can be identified by the presence of a large number of circRNAs in serum exosomes. Recently, Xiao et alCitation106 demonstrated for the first time that cytokeratin 19 (CK19), carbohydrate antigen 125 (CA125), and tumor-associated glycoprotein 72 (TAG72) can be used as biomarkers for colorectal cancer (CRC) due to their different expression levels in exosomes derived from different cells. Moreover, glypican-1 (GPC1) is a proteoglycan exclusively enriched in the surface of cancer exosomes. Melo et alCitation107 reported that healthy people and patients with pancreatic cancer at different stages can be distinguished by GPC1+ circulating exosomes measured in serum. In addition to exosomal cargo differences, increased levels of exosomes were found in the supernatant of cancer cell cultures and in the blood of cancer patients.Citation108,Citation109 An innovative study using small populations found that different types of lipids in exosomes in urine may be specific markers for diagnosing prostate cancer.Citation110
As mentioned above, exosomes also exist in bodily fluids closely related to the eye such as aqueous humor (AH)Citation111,Citation112 and vitreous humor (VH).Citation113 Polarity-specific functions of RPE monolayers can be revealed by directional proteomes of exosomes.Citation114 Exosomes isolated from the cultured supernatant of ARPE-19 cells as well as the AH of patients with age-related macular degeneration (AMD) were identified and compared.Citation115 Results indicated that the specific proteins were elevated in the AH of individuals with AMD subjects. This result provides evidence that exosomal proteins in AH may be used as a biomarker for the diagnosis of AMD. In addition, the basal side of RPE cells, the main lesion site of AMD can release exosomes. These exosomes that enter the blood from the choroid are a potential biomarker for the diagnosis of retinal diseases.Citation116 Similarly, the potential of exosomes and their RNA payloads as specific markers in the diagnosis of eye diseases has attracted much attention as of late. Dismuke et al first demonstrated that exosomes are the major EV type in AH and contain characteristic exosomal RNA, which may provide important basic information for the development of exosomes for the diagnosis of diseases.Citation111
Exosomes as Drug Delivery Vesicles
Exosomes are potential natural drug carriers due their structural composition, diverse biological functions, and unique biological origin. Research has suggested that exosomes could be used to passively and actively deliver drugs with different properties and components.Citation117 The efficacy and stability of a drug can be improved by anchoring it to tetraspanins embedded in the exosomal membrane. For example, the attachment of myostatin propeptide to CD63 on the exosomal membrane not only solves the problem of its low stability in serum, but also enhances its inhibitory effect on mature myostatin without toxicity.Citation118 CP05, a peptide of phage display, can increase its efficiency in capturing and loading exosomes and increases the expression of corresponding proteins in muscles by attaching to exosomal CD63.Citation119 Furthermore, exosomes facilitate the function of certain bioactive substances by encapsulating them and delivering them to receptor cells. For example, exogenous proteins can be loaded by exosomes and transferred across the blood-brain barrier to target cells, which may be a promising treatment for brain diseases.Citation120 Chen et alCitation121 found that endogenous exosomes can independently carry gRNA or Cas9 proteins of the clustered regularly interspaced short palindromic repeats/associated nuclease (CRISPR/Cas) system, thereby transferring the gene editing activity of this system between donor and recipient cells. In addition to the direct effect, autologous exosomes can indirectly promote the delivery of exogenous nanodrugs in vivo and increase the efficacy of drugs.Citation122
Because of the limitations of traditional methods for the treatment of eye diseases, increasing attention is being given to exosomes as a medium for drug delivery. Comparison of intravitreal injection of traditional AAV2 with the novel exosome-associated AAV2 (exo-AAV2) revealed that exo-AAV2 carrying the GFP-coding gene adeno-associated virus (AAV) was more strongly expressed and had greater penetrability in the retina.Citation123 Additionally, when exosomes from MSCs overexpress miR-126, intravitreal injection can effectively inhibit the signaling pathways associated with inflammatory response in DR.Citation124
Toxicity of Exosomes
Despite increasing research focused on the therapeutic effects of exosomes, little is known about exosomal toxicity. The potential widespread applications of exosomes as therapeutic agents raise concerns about their potential adverse impacts. Exosomes secreted by genetically engineered T cells whose surfaces bind to chimeric antigen receptor (CAR) have been found to have low toxicity, although they effectively inhibit tumor development.Citation125 Recent studies have shown that exosomes contribute to the development of AIDS. HIV-1-infected T cells can not only release exosomes containing the transactivation response (TAR) element RNA to activate the associated cascade and thus aggravate the cancer, but also uninfected cells can produce exosomes to activate latent HIV-1 in infected cells to promote cancer development.Citation126,Citation127 In addition, some liver-derived exosomes can cause toxic reactions when applied to target cells. Exogenous exosomes released after acetaminophen liver injury in mice can be absorbed by mouse primary hepatocytes, causing toxicity of receptor cells and induce cell death.Citation128 Gao et alCitation129 demonstrated that circulating exosomes loaded with metallothionein 1D pseudogene (MT1DP), released by hepatocytes, target kidney cells to enhance cadmium-induced nephrotoxicity. Toxic amyloid-beta oligomers can be carried by exosomes from the brain of people with Alzheimer’s disease, allowing these oligomers to move between neurons and worsen the disease.Citation130 Zhang et alCitation14 have shown that exosomes in serum produced after hepatic ischemia-reperfusion may cause oxidative stress and damage hippocampal and cortical neurons. The eye is an extremely sensitive organ, and with the emergence of exosomes in the study of eye diseases, the toxicity of exosomes on the eyes is a matter of concern that needs to be addressed. However, only a few studies have focused on the adverse effects of exosomes in the eye. Huang et alCitation131 reported that when exosomes are loaded with IgG, they can activate the classical complement pathway, promoting microvascular damage and aggravating DR. However, exosomes lacking Ig-G have the opposite effect. Recently, Ke et alCitation132 extracted exosomes from normal RPE cells and rotenone-induced RPE cells. Compared with the control group, exosomes released by RPE cells under oxidative stress induced inflammation and apoptosis in normal RPE cells through the Apaf1/Caspase-9 signaling pathway. This led to retinal functional damage. Further research into exosomal toxicity in the eye is warranted.
Exosomes and Oxidative Stress
The concept of oxidative stress was formulated in redox biology and medicine in the 1980s, and it has been studied in a wide range of fields, including chemistry, biology, and pathology.Citation133 Oxidative stress is closely linked with a variety of diseases. Subsequently, increased research has focused on the relationship between exosomes and oxidative stress. Plasma exosomes from HIV patients treated with antiretroviral therapies have been reported to be associated with changes in the levels of certain oxidative stress markers, indicating that these exosomes are loaded with oxidative stress-related proteins.Citation134 In addition, exosomes and their cargoes may play a role in various diseases by mediating different molecular mechanisms associated with oxidative stress. For example, oxidative stress not only induced more exosomes from cardiac progenitor cells, but it also significantly upregulated miR-21 in exosomes under this condition. Importantly, miR-21 can downregulate its target gene programmed cell death 4 to inhibit oxidative stress-related apoptosis in myocardial cells.Citation135 Saeed-Zidane et alCitation136 found that exosomes, secreted by bovine granulosa cells under oxidative stress, contained abundant Nrf2 mRNA and possible antioxidants, which may mediate the molecular defense mechanism related to oxidative stress in granulosa cells. Similarly, oxidative stress plays an important role in many ocular diseases, such as diabetic retinopathy and age-related macular degeneration.Citation137–Citation139 There is no doubt that the role of exosomes released by eye cells exposed to oxidative stress is worthy of considerable attention.
Exosomes in Diseased Eyes
Exosomes contain proteins, lipids, and nucleic acids that are associated with their parental cells and thus have a wide range of biological functions. The following is an introduction to their functions in eye diseases ().
Table 1 A Brief Description of the Biological Role of Exosomes in Related Eye Diseases
Diabetic Retinopathy
Diabetic retinopathy (DR), one of the serious complications of diabetes, is the most important manifestation of diabetic microangiopathy, resulting in a fundus lesion with specific changes. The main pathological stages of DR include non-proliferative DR, pre-proliferative DR, and proliferative DR. Diabetic macular oedema, characterized by vascular leakage and macula, is a special subtype of non-proliferative of DR.Citation140,Citation141 The pre-proliferative stage refers to capillary closures and the existence of a non-perfusion area. The development of neovascularization is a marker of proliferative DR, and hypoxia plays an important role in this stage.Citation141 Obviously, retinal ischemia-reperfusion injury (IRI) to a large extent mediates the pathological process of DR.Citation142 Retinal ischemia directly leads to temporary hypoxia and the loss of important nutrients. Reperfusion then promotes the production of excessive reactive oxygen species (ROS) inducing oxidative stress and further inflammatory responses.Citation143 Finally, IRI causes necrosis, apoptosis, and autophagy resulting in a decrease in retinal neurons, especially RGCs.Citation144
The role of exosomes secreted by MSCs in the eye is coming into focus as studies confirm that DR can be improved after intravitreal injection of MSC derived exosomes.Citation124 Moisseiev et alCitation145 demonstrated the effect of exosomes on retinal ischemia by intravitreal injection of hypoxia-cultured exosomes containing angiogenic active ingredients from human-MSCs (hMSCs) and control saline in oxygen-induced retinopathy (OIR) mouse models. Compared with the control group, exosome treatment inhibited retinal thinning and alleviated retinal ischemia. Studies have also been conducted on the role of exosomes in diabetic microangiopathy. EVs in plasma of diabetic patients with retinopathy were found to increase the expression of cytokines and angiogenic factors, suggesting that these EVs may be involved in the transmission of carrier-related factors.Citation146 Recently, Maisto et alCitation147 used high concentrations of glucose to stimulate retinal photoreceptors to simulate DR. They found that high glucose levels increased vascular endothelial growth factor (VEGF) and diminished anti-angiogenic miRNA levels in photoreceptors and exosomes. A study on diabetic microvascular complications found that exosomes can carry circular RNAs-cPWWP2A, an endogenous miR-579 sponge, and indirectly mediate the function of retinal vessels in patients with diabetes through different expression levels of cPWWP2A and miR-579.Citation148 However, it has been found that the level of platelet-rich plasma exosomes is significantly increased in the plasma of diabetic rats, which can up-regulate the TLR4 signaling pathway that regulates inflammatory response and can cause retinal endothelial injury.Citation149 Kamalden et alCitation150 confirmed that miR-15a, delivered by circulating exosomes after pancreatic production, can cause oxidative stress to enhance type-2-diabetes (T2D)-induced DR. PPARγ is an active ingredient in exosomes. Katome et alCitation151 showed that PPARγ was significantly increased in AH and VH of patients with proliferative DR, revealing that exosomes play an important role in DR.
Age-Related Macular Degeneration
The retina is a soft transparent membrane that is sensitive to light stimulation and is attached to the inner surface of the choroid through the outermost layer of RPE and the Burch membrane.Citation152 The RPE is composed of monolayer pigment epithelial cells, which are vulnerable to oxidative stress, especially ROS, due to their anatomical position and function.Citation103
Age-related macular degeneration (AMD), which occurs mostly in people over 50 years of age, is the leading cause of blindness among the elderly in developed countries. According to the clinical manifestations and pathological changes, it is divided into dry AMD and wet AMD. Wet AMD, also known as neovascular AMD, is characterized by the occurrence of choroidal neovascularization (CNV) and accompanied by choroidal and retinal hemorrhage. Wet AMD produces more serious visual impairment than dry AMD. Neovascularization does not occur in dry AMD, but yellow drusen form underneath the RPE. Dry AMD is accompanied by geographical atrophy, also known as atrophic AMD.Citation153
RPE, which forms the outer blood-retina barrier (BRB), plays an important role in the pathogenesis of AMD. It has been reported that oxidative stress, an important factor leading to AMD, can cause donor cells to secrete EVs that transmit stress signals to normal RPE cells to weaken the barrier function of RPE.Citation154 Many types of retinal cells can generate and release exosomes, such as RPE cells and retinal astroglial cells (RACs), which may be involved in the pathological process of AMD. However, the roles of these different cell-derived exosomes in the retina are also markedly different. Hajrasouliha et alCitation155 demonstrated that RACs-derived exosomes contain anti-angiogenic components that can reduce retinal vascular leakage and curb CNV. These results suggest a new treatment of AMD. Exosomes from RPE cells, however, do not have this capability. Interestingly, unlike healthy RPE cells, RPE cells under stress release large amounts of exosomes containing VEGFR2 that enhance the formation of new blood vessels. In addition, exosomes from RPE cells under stress increase autophagy to promote AMD.Citation156 Moreover, Wang et alCitation157 conjectured that exosomes and autophagy might mediate and enhance the formation of drusen during AMD. Bevacizumab, an anti-VEGF antibody against AMD, has also been found to be taken up by RPE cells after intravitreal injection and then re-released by exosomes.Citation158
Numerous studies have revealed that exosomes contain cargo that may serve as biomarkers for the diagnosis of AMD, particularly microRNA and proteins. For instance, Kang et alCitation115 indicated that exosome proteins have great potential as novel biomarkers for the diagnosis of neovascular AMD as they found changes in exosomal proteins levels in the AH of subjects with AMD. A recent study found that the expression of three new circulating microRNAs, including miR-486-5p, miR-626, and miR-885-5p, significantly differed in control and individuals with AMD, but their effects were not identical. The results showed that the expression of miR-486-5p and miR-626 were up-regulated in patients, while the expression of miR-885-5p was down-regulated in patients.Citation159 In addition, Biasutto et alCitation160 used reverse phase protein arrays analysis to find that the content of some phosphoproteins in cultured ARPE-19 cells-derived exosomes subjected to oxidative stress were changed, suggesting that these exosomal proteins may be involved in cell apoptosis and cell metabolism. All of the above studies provide theoretical and experimental basis for exosomes to be used as specific markers in the diagnosis of AMD.
Autoimmune Uveitis
Uveitis is the general name for inflammation of iris, ciliary body, and choroidal. According to the location, the disease can be divided into anterior uveitis, posterior uveitis middle uveitis, and panuveitis. Autoimmune uveitis (AU) involves not only inflammation of the uveitis itself, but also inflammation of adjacent tissues such as the retina and vitreous. RPE cells, which have immunosuppressive properties, may modulate the immune response by releasing exosomes.Citation15 In recent years, the role of exosomes in autoimmune uveitis has been studied.
Studies have shown that MSCs used in the treatment of eye diseases have a significant immunomodulatory effect and can repair damaged or diseased tissues and organs. Previous studies have demonstrated that MSCs can improve experimental AU in rat models. Based on the relevant experiments of MSCs, Bai et alCitation161 verified the role of exosomes secreted by MSCs in the experimental AU rat model through periocular injection. They found that periocular injection of hMSCs-derived exosomes reduced leukocyte infiltration in the eyes and alleviated uveitis. These results suggest that exosomes from hMSCs have promising potential in the treatment of AU. Interestingly, the MSC exosomes were found to regulate the immune response by inhibiting the flow of inflammatory cells into the eye rather than by inhibiting the specific immune response of T cells. In addition, a study has shown that intravenous administration of exosomes released by MSCs immediately after immunization can protect the retinal structure of experimental AU rat model and reduce inflammatory cell infiltration. These results are similar to those found with injection of MSC exosomes around the eye.Citation162
Glaucoma and Traumatic Optic Neuropathies
Glaucoma is a group of diseases characterized by atrophy and depression of the optic nerve, visual field defects, and visual decline. Pathologically increased intraocular pressure (IOP) and insufficient blood supply to the optic nerve are the primary risk factors for glaucoma. The tolerance of the optic nerve to pressure damage is also related to the occurrence and development of glaucoma. There are two subtypes of glaucoma: open-angle glaucoma and closed-angle glaucoma. The two subtypes can be further divided into primary and secondary glaucoma.Citation163 Patients with primary glaucoma have normal or elevated IOP, but no clear cause has been identified.Citation163–Citation165 In addition, normal tension glaucoma, with no increase in IOP, is considered likely to be a singular subtype of primary open-angle glaucoma (POAG).Citation166 Unlike primary glaucoma, secondary glaucoma has identifiable causes, such as elevated IOP.Citation163,Citation164 During the development of glaucoma, the RGCs are gradually lost due to the increased IOP caused by the increased generation and reduced outflow of AH,Citation167 which hinders the transmission of information from the retina to the brain. Therefore, reducing intraocular pressure and decreasing the loss of RGCs are the main methods of glaucoma treatment. Trabecular meshwork (TM) that can drain AH to reduce IOP is the main objective of clinical treatment.Citation168
Lerner et alCitation169 performed protein analysis on exosomes secreted by primary cells of the non-pigmented ciliary epithelium (NPCE) and a NPCE cell line. They found that these exosomes contained a large number of negative modulators of Wnt signaling miR-29b. After treating TM cells with these exosome samples, the expression of COL3A1 was down-regulated, suggesting that NPCE cells-derived EVs may mediate the regulation of TM classical Wnt signaling. Lerner et alCitation170 also reported that exosomes derived from NPCE cells are involved in the regulation of Wnt signaling proteins in TM cells, suggesting that EVs are involved in signal communication between related cells in the ocular water system. A study reported by Liu et alCitation171 suggested the potential role of exosomes in POAG. They conducted a large-scale study of TM gene expression in patients with POAG and healthy subjects, and for the first time demonstrated that TM gene expression occurred in patients with a Q368X myocilin mutation. It is worth noting that flotillin-2 and tetraspanin-6, two exosome-enriched proteins, had different TM gene expression in POAG patients. Moreover, exosome-related proteins, including matrix GLA protein, SPARC, and TIMP2, are expressed by a few TM characteristic genes that were different in non-myocilin POAG individuals.Citation171,Citation172 One study used miRNA sequencing (miRNA-Seq) to detect the expression of miR-182 in various types of human eye tissues, human AH, and exosomes derived from human primary TM cells, respectively, to verify the correlation between miRNA variation and POAG.Citation112 In addition to studies on TM, the therapeutic effect of MSC-derived exosomes from different tissues on glaucoma and traumatic optic neuropathies has been extensively investigated. For example, a series of tests were carried out after weekly intravitreal injection of exosomes released by bone marrow-derived MSCs (BMSCs) in rats of the optic nerve crush (ONC) model. Exosomes from BMSCs were found to enhance the survival of RGCs and promote axonal regeneration, thus significantly protecting the optic nerve.Citation62 This neuroprotective effect of BMSC-derived exosomes has also been demonstrated in the genetic DBA/2J mouse model of glaucoma.Citation173 However, Pan et alCitation174 investigated the effect of exosomes released by umbilical cord MSCs on ONC rats and found that cell-derived exosomes increased the survival of RGCs but did not promote axonal regeneration.
Corneal Disease
Corneal disease, one of the primary eye diseases leading to blindness, causes the clear cornea to appear pale and opaque, which can lead to blurred vision, decreased vision, and even blindness. Corneal defects in mice with mucopolysaccharidosis VII have been found to be repaired by human umbilical cord MSC transplantation.Citation175 Han et alCitation176 isolated exosomes containing matrix metalloproteinase (MMP) 14 from corneal fibroblasts. MMP14, an angiogenic enzyme, is transported by corneal fibroblast exosomes to vascular endothelial cells to promote corneal angiogenesis. In addition, MMP14 is involved in the recruitment of its target MMP2 on corneal fibroblast exosomes. Samaeekia et alCitation61 extracted exosomes from the secretome of human corneal mesenchymal stromal cells by differential ultracentrifugation and conducted in vitro and in vivo experiments. Compared with a control group, both the remaining wound area in an in vitro scratch assay and the degree of wounds in in vivo corneal epithelial debridement wounds in mice were significantly improved by these exosomes. The results suggest that human corneal mesenchymal stromal cells exosomes may be used to treat corneal epithelial trauma. More recently, one study has shown that the treatment of HCEC-derived EVs increased myofibroblast differentiation via the expression of α-smooth muscle actin and the elevation of contractility, suggesting that these EVs may conduct corneal scarring.Citation60
Retinopathy of Prematurity
The data shows that there are different causes of childhood blindness in different regions of the world, and retinopathy of prematurity (ROP) is one of the leading causes of childhood blindness in the United States.Citation177 Infants born before term (40 weeks) are prone to various lesions because their tissues and organs, especially those related to the visual system, are not fully developed. ROP, clinically characterized by retinal vasculature abnormalities, is a clinical entity involving the retina.Citation178,Citation179 ROP consists of two oxygen-dependent postpartum phases: Phase 1, characterized by the arrest of vascular growth, and Phase 2, characterized by vascular proliferation.Citation180
In phase 1, hyperoxia, which inhibits VEGF, is an important inducer of this disease. Importantly, normal air conditions can also lead to hyperoxia in addition to oxygen therapy to increase infant survival.Citation180 As a baby grows, the increased metabolism changes the retina from a hyperoxic state to a hypoxic state, and the disease enters phase 2. In contrast, this stage promotes VEGF.Citation180 One study showed that exosomes from MSCs could effectively improve hyperoxia-induced retinopathy, and that exosomes were well tolerated after intravitreal injection, providing a novel idea for safe non-cellular therapy for ROP.Citation145 Of note, Xu et alCitation181 investigated the role of exosomes derived from microglia in an animal model of ROP. Recently, the idea that microglia cells are involved in the formation and development of the central nervous system has emerged,Citation182 and a recent study has found that microglia may improve retina-related diseases.Citation183 Microglia-derived exosomes were injected into the vitreous body of a mouse model and showed a series of manifestations: 1) reduction of avascular areas and neovascular tufts; 2) decreased expression of VEGF and transforming growth factor β (TGF-β), and 3) inhibition of photoreceptor apoptosis.Citation181 These findings also provide a theoretical basis for exosomes to be used as a new treatment method for ROP. Indeed, the efficacy of exosomes in ROP remains to be further studied.
Uveal Melanoma
Although eye cancer is rare, it can be devastating when it occurs. Uveal melanoma (UM) is a common cancer of the eye.Citation184 UM, a primary intraocular malignancy, is a general term for melanoma of the choroid, ciliary body, or iris.Citation185 Notably, nearly half of patients with UM have organ metastases, especially liver metastases.Citation186 Eldh et alCitation187 extracted exosomes from liver perfusate from patients with hepatic metastatic UM who underwent hepatic perfusion. In metastatic UM, exosomes originating from melanoma are released for hepatic circulation, and these exosomes contain Melan-A and melanoma-related microRNAs, thus further demonstrating their origin. More importantly, exosomes can be isolated from local blood circulation in patients with metastatic UM, and their RNA clusters are specific.Citation187 Similarly, miRNA profiles from vitreous exosomes from patients with UM were analyzed in another study, which also suggested that exosomal RNAs may be potential specific markers for diagnosing UM.Citation188 Thus, exosomes have not been well studied in UM, but their potential and prospects in the diagnosis of disease are worth exploring.
Conclusions and Future Perspectives
Current evidence has shown that exosomes contain a variety of proteins and genetic material that contribute to their broad biological functions including information transmission, immune regulation, repair, and regeneration. As discussed in this review, a number of studies provide evidence for the important role played by exosomes in the pathological process of ocular diseases, including inflammation, neuronal degeneration, oxidative stress, and neovascularization. In addition, exosomes have the ability to cross biological barriers. Using exosomes as biomarkers or therapeutic carriers may bring better and personalized treatment to patients with eye diseases. Taken together, this review advances the understanding of the role of exosomes in ocular diseases. However, based on the complexity and diversity of exosomes, the functions and mechanisms of exosomes in ocular diseases requires further investigation. The role of exosomes as biomarkers or therapeutic vehicles in ocular disease diagnosis and clinical treatments needs further study. Moreover, there is no established standard for separating and purifying exosomes, let alone the mass production of exosomes, which needs urgently require research. Therefore, significant studies are needed to develop such therapeutic treatments in ocular diseases.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (81970826) to Zhuhong Zhang, Talent Induction Program for Youth Innovation Teams in Colleges and University of Shandong Province to Yanping Zhu, and National Natural Science Foundation of China (81800853) to Feng Jiang.
Disclosure
The authors report no conflicts of interest for this work.
References
- Mathieu M, Martin-Jaular L, Lavieu G, Thery C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat Cell Biol. 2019;21(1):9–17. doi:10.1038/s41556-018-0250-930602770
- Hessvik NP, Llorente A. Current knowledge on exosome biogenesis and release. Cell Mol Life Sci. 2018;75(2):193–208. doi:10.1007/s00018-017-2595-928733901
- Pan BT, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: selective externalization of the receptor. Cell. 1983;33(3):967–978. doi:10.1016/0092-8674(83)90040-56307529
- Pan BT, Teng K, Wu C, Adam M, Johnstone RM. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J Cell Biol. 1985;101(3):942–948. doi:10.1083/jcb.101.3.9422993317
- Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem. 1987;262(19):9412–9420.3597417
- Yu B, Zhang X, Li X. Exosomes derived from mesenchymal stem cells. Int J Mol Sci. 2014;15(3):4142–4157. doi:10.3390/ijms1503414224608926
- Mueller SK, Nocera AL, Bleier BS. Exosome function in aerodigestive mucosa. Nanomedicine. 2018;14(2):269–277. doi:10.1016/j.nano.2017.10.00829127037
- Colombo M, Raposo G, Thery C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014;30:255–289. doi:10.1146/annurev-cellbio-101512-12232625288114
- Li S, Li Y, Chen B, et al. exoRBase: a database of circRNA, lncRNA and mRNA in human blood exosomes. Nucleic Acids Res. 2018;46(D1):D106–D112. doi:10.1093/nar/gkx89130053265
- Klingeborn M, Dismuke WM, Bowes Rickman C, Stamer WD. Roles of exosomes in the normal and diseased eye. Prog Retin Eye Res. 2017;59:158–177. doi:10.1016/j.preteyeres.2017.04.00428465248
- Kang X, Zuo Z, Hong W, Tang H, Geng W. Progress of research on exosomes in the protection against ischemic brain injury. Front Neurosci. 2019;13:1149. doi:10.3389/fnins.2019.0114931736691
- Yamashita T, Takahashi Y, Takakura Y. Possibility of exosome-based therapeutics and challenges in production of exosomes eligible for therapeutic application. Biol Pharm Bull. 2018;41(6):835–842. doi:10.1248/bpb.b18-0013329863072
- Akbari A, Jabbari N, Sharifi R, et al. Free and hydrogel encapsulated exosome-based therapies in regenerative medicine. Life Sci. 2020. doi:10.1016/j.lfs.2020.117447
- Zhang L, Liu H, Jia L, et al. Exosomes mediate hippocampal and cortical neuronal injury induced by hepatic ischemia-reperfusion injury through activating pyroptosis in rats. Oxid Med Cell Longev. 2019;2019:1–17. doi:10.1155/2019/3753485
- Li N, Zhao L, Wei Y, Ea VL, Nian H, Wei R. Recent advances of exosomes in immune-mediated eye diseases. Stem Cell Res Ther. 2019;10(1):278. doi:10.1186/s13287-019-1372-031470892
- Tkach M, Thery C. Communication by extracellular vesicles: where we are and where we need to go. Cell. 2016;164(6):1226–1232. doi:10.1016/j.cell.2016.01.04326967288
- Edgar JR. Q&A: what are exosomes, exactly? BMC Biol. 2016;14:46. doi:10.1186/s12915-016-0268-z27296830
- Mulcahy LA, Pink RC, Carter DR. Routes and mechanisms of extracellular vesicle uptake. J Extracell Vesicles. 2014;3doi 10.3402/jev.v3.24641.
- McKelvey KJ, Powell KL, Ashton AW, Morris JM, McCracken SA. Exosomes: mechanisms of uptake. J Circ Biomark. 2015;4:7. doi:10.5772/6118628936243
- Nedaeinia R, Manian M, Jazayeri M, et al. Circulating exosomes and exosomal microRNAs as biomarkers in gastrointestinal cancer. Cancer Gene Ther. 2017;24(2):48–56. doi:10.1038/cgt.2016.7727982021
- Cho S, Yang H, Rhee W. Simultaneous multiplexed detection of exosomal microRNAs and surface proteins for prostate cancer diagnosis. Biosens Bioelectron. 2019;146:111749. doi:10.1016/j.bios.2019.11174931600625
- Xia X, Wang Y, Huang Y, Zhang H, Lu H, Zheng JC. Exosomal miRNAs in central nervous system diseases: biomarkers, pathological mediators, protective factors and therapeutic agents. Prog Neurobiol. 2019;183:101694. doi:10.1016/j.pneurobio.2019.10169431542363
- Betzer O, Perets N, Angel A, et al. In vivo neuroimaging of exosomes using gold nanoparticles. ACS Nano. 2017;11(11):10883–10893. doi:10.1021/acsnano.7b0449528960957
- Haney MJ, Klyachko NL, Zhao Y, et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J Control Release. 2015;207:18–30. doi:10.1016/j.jconrel.2015.03.03325836593
- Ha D, Yang N, Nadithe V. Exosomes as therapeutic drug carriers and delivery vehicles across biological membranes: current perspectives and future challenges. Acta Pharm Sin B. 2016;6(4):287–296. doi:10.1016/j.apsb.2016.02.00127471669
- Kibria G, Ramos EK, Wan Y, Gius DR, Liu H. Exosomes as a drug delivery system in cancer therapy: potential and challenges. Mol Pharm. 2018;15(9):3625–3633. doi:10.1021/acs.molpharmaceut.8b0027729771531
- Bourne RRA, Jonas JB, Bron AM, et al. Prevalence and causes of vision loss in high-income countries and in Eastern and Central Europe in 2015: magnitude, temporal trends and projections. Br J Ophthalmol. 2018;102(5):575–585. doi:10.1136/bjophthalmol-2017-31125829545417
- Sabanayagam C, Cheng C-Y. Global causes of vision loss in 2015: are we on track to achieve the Vision 2020 target? The Lancet Global Health. 2017;5(12):e1164–e1165. doi:10.1016/s2214-109x(17)30412-629032196
- Flaxman SR, Bourne RRA, Resnikoff S, et al. Global causes of blindness and distance vision impairment 1990–2020: a systematic review and meta-analysis. The Lancet Global Health. 2017;5(12):e1221–e1234. doi:10.1016/S2214-109X(17)30393-529032195
- Fricke TR, Tahhan N, Resnikoff S, et al. Global prevalence of presbyopia and vision impairment from uncorrected presbyopia: systematic review, meta-analysis, and modelling. Ophthalmology. 2018;125(10):1492–1499. doi:10.1016/j.ophtha.2018.04.01329753495
- DelMonte D, Kim T. Anatomy and physiology of the cornea. J Cataract Refract Surg. 2011;37(3):588–598. doi:10.1016/j.jcrs.2010.12.03721333881
- Vallabh NA, Romano V, Willoughby CE. Mitochondrial dysfunction and oxidative stress in corneal disease. Mitochondrion. 2017;36:103–113. doi:10.1016/j.mito.2017.05.00928549842
- Forrester JV, Kuffova L, Dick AD. Autoimmunity, autoinflammation, and infection in uveitis. Am J Ophthalmol. 2018;189:77–85. doi:10.1016/j.ajo.2018.02.01929505775
- Mathew B, Ravindran S, Liu X, et al. Mesenchymal stem cell-derived extracellular vesicles and retinal ischemia-reperfusion. Biomaterials. 2019;197:146–160. doi:10.1016/j.biomaterials.2019.01.01630654160
- Yu B, Li XR, Zhang XM. Mesenchymal stem cell-derived extracellular vesicles as a new therapeutic strategy for ocular diseases. World J Stem Cells. 2020;12(3):178–187. doi:10.4252/wjsc.v12.i3.17832266050
- Mead B, Tomarev S. Extracellular vesicle therapy for retinal diseases. Prog Retin Eye Res. 2020;79:100849. doi:10.1016/j.preteyeres.2020.10084932169632
- Li SF, Han Y, Wang F, Su Y. Progress in exosomes and their potential use in ocular diseases. Int J Ophthalmol. 2020;13(9):1493–1498. doi:10.18240/ijo.2020.09.2332953591
- Villarroya-Beltri C, Baixauli F, Gutierrez-Vazquez C, Sanchez-Madrid F, Mittelbrunn M. Sorting it out: regulation of exosome loading. Semin Cancer Biol. 2014;28:3–13. doi:10.1016/j.semcancer.2014.04.00924769058
- Chen L, Chen R, Kemper S, Brigstock DR. Pathways of production and delivery of hepatocyte exosomes. J Cell Commun Signal. 2018;12(1):343–357. doi:10.1007/s12079-017-0421-729063370
- Gurunathan S, Kang MH, Jeyaraj M, Qasim M, Kim JH. Review of the isolation, characterization, biological function, and multifarious therapeutic approaches of exosomes. Cells. 2019;8(4):Apr. doi:10.3390/cells8040307
- Alenquer M, Amorim MJ. Exosome biogenesis, regulation, and function in viral infection. Viruses. 2015;7(9):5066–5083. doi:10.3390/v709286226393640
- Colombo M, Moita C, van Niel G, et al. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J Cell Sci. 2013;126(Pt 24):5553–5565. doi:10.1242/jcs.12886824105262
- Juan T, Fürthauer M. Biogenesis and function of ESCRT-dependent extracellular vesicles. Semin Cell Dev Biol. 2018;74:66–77. doi:10.1016/j.semcdb.2017.08.02228807885
- Baietti MF, Zhang Z, Mortier E, et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat Cell Biol. 2012;14(7):677–685. doi:10.1038/ncb250222660413
- Hikita T, Kuwahara A, Watanabe R, Miyata M, Oneyama C. Src in endosomal membranes promotes exosome secretion and tumor progression. Sci Rep. 2019;9(1):3265. doi:10.1038/s41598-019-39882-z30824759
- Hyenne V, Labouesse M, Goetz JG. The Small GTPase Ral orchestrates MVB biogenesis and exosome secretion. Small GTPases. 2018;9(6):445–451. doi:10.1080/21541248.2016.125137827875100
- Murrow L, Debnath J. Atg12-Atg3 coordinates basal autophagy, endolysosomal trafficking, and exosome release. Mol Cell Oncol. 2018;5(5):e1039191. doi:10.1080/23723556.2015.103919130263931
- Yoon S, Kovalenko A, Bogdanov K, Wallach D. MLKL, the protein that mediates necroptosis, also regulates endosomal trafficking and extracellular vesicle generation. Immunity. 2017;47(1):51–65 e7. doi:10.1016/j.immuni.2017.06.00128666573
- Zhang J, Li S, Li L, et al. Exosome and exosomal microRNA: trafficking, sorting, and function. Genomics Proteomics Bioinformatics. 2015;13(1):17–24. doi:10.1016/j.gpb.2015.02.00125724326
- Skotland T, Sandvig K, Llorente A. Lipids in exosomes: current knowledge and the way forward. Prog Lipid Res. 2017;66:30–41. doi:10.1016/j.plipres.2017.03.00128342835
- Pegtel DM, Gould SJ. Exosomes. Annu Rev Biochem. 2019;88:487–514. doi:10.1146/annurev-biochem-013118-11190231220978
- Yim N, Ryu S-W, Choi K, et al. Exosome engineering for efficient intracellular delivery of soluble proteins using optically reversible protein–protein interaction module. Nat Commun. 2016;7. doi:10.1038/ncomms12277
- He C, Zheng S, Luo Y, Wang B. Exosome theranostics: biology and translational medicine. Theranostics. 2018;8(1):237–255. doi:10.7150/thno.2194529290805
- Valadi H, Ekstrom K, Bossios A, Sjostrand M, Lee JJ, Lotvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9(6):654–659. doi:10.1038/ncb159617486113
- Cheng J, Meng J, Zhu L, Peng Y. Exosomal noncoding RNAs in Glioma: biological functions and potential clinical applications. Mol Cancer. 2020;19(1):66. doi:10.1186/s12943-020-01189-332213181
- Farooqi AA, Desai NN, Qureshi MZ, et al. Exosome biogenesis, bioactivities and functions as new delivery systems of natural compounds. Biotechnol Adv. 2018;36(1):328–334. doi:10.1016/j.biotechadv.2017.12.01029248680
- Abels ER, Breakefield XO. Introduction to extracellular vesicles: biogenesis, RNA cargo selection, content, release, and uptake. Cell Mol Neurobiol. 2016;36(3):301–312. doi:10.1007/s10571-016-0366-z27053351
- Horibe S, Tanahashi T, Kawauchi S, Murakami Y, Rikitake Y. Mechanism of recipient cell-dependent differences in exosome uptake. BMC Cancer. 2018;18(1):47. doi:10.1186/s12885-017-3958-129306323
- Ren R, Tan X-H, Zhao J-H, et al. Bone marrow mesenchymal stem cell-derived exosome uptake and retrograde transport can occur at peripheral nerve endings. Artif Cells, Nanomed Biotechnol. 2019;47(1):2918–2929. doi:10.1080/21691401.2019.164071331317777
- McKay TB, Hutcheon AEK, Zieske JD, Ciolino JB. Extracellular vesicles secreted by corneal epithelial cells promote myofibroblast differentiation. Cells. 2020;9(5):1080. doi:10.3390/cells9051080
- Samaeekia R, Rabiee B, Putra I, et al. Effect of human corneal mesenchymal stromal cell-derived exosomes on corneal epithelial wound healing. Invest Ophthalmol Vis Sci. 2018;59(12):5194–5200. doi:10.1167/iovs.18-2480330372747
- Mead B, Tomarev S. Bone marrow-derived mesenchymal stem cells-derived exosomes promote survival of retinal ganglion cells through miRNA-dependent mechanisms. Stem Cells Transl Med. 2017;6(4):1273–1285. doi:10.1002/sctm.16-042828198592
- Thery C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7(1):1535750. doi:10.1080/20013078.2018.153575030637094
- Tkach M, Kowal J, Thery C. Why the need and how to approach the functional diversity of extracellular vesicles. Philos Trans R Soc Lond B Biol Sci. 2018;373(1737):20160479. doi:10.1098/rstb.2016.047929158309
- Witwer KW, Thery C. Extracellular vesicles or exosomes? On primacy, precision, and popularity influencing a choice of nomenclature. J Extracell Vesicles. 2019;8(1):1648167. doi:10.1080/20013078.2019.164816731489144
- Li P, Kaslan M, Lee SH, Yao J, Gao Z. Progress in exosome isolation techniques. Theranostics. 2017;7(3):789–804. doi:10.7150/thno.1813328255367
- Lucchetti D, Fattorossi A, Sgambato A. Extracellular vesicles in oncology: progress and pitfalls in the methods of isolation and analysis. Biotechnol J. 2019;14(1):e1700716. doi:10.1002/biot.20170071629878510
- Théry C, Amigorena S, Raposo G, Clayton A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Current Protocols Cell Biol. 2006;Unit 3.22. doi:10.1002/0471143030.cb0322s30
- Purushothaman A. Exosomes from cell culture-conditioned medium: isolation by ultracentrifugation and characterization. Methods Mol Biol. 2019;1952:233–244. doi:10.1007/978-1-4939-9133-4_1930825179
- Livshits MA, Khomyakova E, Evtushenko EG, et al. Isolation of exosomes by differential centrifugation: theoretical analysis of a commonly used protocol. Sci Rep. 2015;5:17319. doi:10.1038/srep1731926616523
- Momen-Heravi F. Isolation of extracellular vesicles by ultracentrifugation. Methods Mol Biol. 2017;1660:25–32. doi:10.1007/978-1-4939-7253-1_328828645
- Mincheva‐Nilsson L, Baranov V, Nagaeva O, Dehlin E. Isolation and characterization of exosomes from cultures of tissue explants and cell lines. Curr Protoc Immunol. 2016;115(1):1442 1–14 42 21. doi:10.1002/cpim.17
- Gupta S, Rawat S, Arora V, et al. An improvised one-step sucrose cushion ultracentrifugation method for exosome isolation from culture supernatants of mesenchymal stem cells. Stem Cell Res Ther. 2018;9(1):180. doi:10.1186/s13287-018-0923-029973270
- Greening DW, Xu R, Ji H, Tauro BJ, Simpson RJ. A protocol for exosome isolation and characterization: evaluation of ultracentrifugation, density-gradient separation, and immunoaffinity capture methods. Methods Mol Biol. 2015;1295:179–209. doi:10.1007/978-1-4939-2550-6_1525820723
- Li K, Wong DK, Hong KY, Raffai RL. Cushioned-Density Gradient Ultracentrifugation (C-DGUC): a refined and high performance method for the isolation, characterization, and use of exosomes. Methods Mol Biol. 2018;1740:69–83. doi:10.1007/978-1-4939-7652-2_729388137
- Baranyai T, Herczeg K, Onodi Z, et al. Isolation of exosomes from blood plasma: qualitative and quantitative comparison of ultracentrifugation and size exclusion chromatography methods. PLoS One. 2015;10(12):e0145686. doi:10.1371/journal.pone.014568626690353
- An M, Wu J, Zhu J, Lubman DM. Comparison of an optimized ultracentrifugation method versus size-exclusion chromatography for isolation of exosomes from human serum. J Proteome Res. 2018;17(10):3599–3605. doi:10.1021/acs.jproteome.8b0047930192545
- Lobb R, Moller A. Size exclusion chromatography: a simple and reliable method for exosome purification. Methods Mol Biol. 2017;1660:105–110. doi:10.1007/978-1-4939-7253-1_928828651
- Monguio-Tortajada M, Galvez-Monton C, Bayes-Genis A, Roura S, Borras FE. Extracellular vesicle isolation methods: rising impact of size-exclusion chromatography. Cell Mol Life Sci. 2019;76(12):2369–2382. doi:10.1007/s00018-019-03071-y30891621
- Oksvold MP, Neurauter A, Pedersen KW. Magnetic bead-based isolation of exosomes. Methods Mol Biol. 2015;1218:465–481. doi:10.1007/978-1-4939-1538-5_2725319668
- Soares Martins T, Catita J, Martins Rosa I, Henriques AG. Exosome isolation from distinct biofluids using precipitation and column-based approaches. PLoS One. 2018;13(6):e0198820. doi:10.1371/journal.pone.019882029889903
- Weng Y, Sui Z, Shan Y, et al. Effective isolation of exosomes with polyethylene glycol from cell culture supernatant for in-depth proteome profiling. Analyst. 2016;141(15):4640–4646. doi:10.1039/c6an00892e27229443
- Kalluri R. The biology and function of exosomes in cancer. J Clin Invest. 2016;126(4):1208–1215. doi:10.1172/JCI8113527035812
- Kourembanas S. Exosomes: vehicles of intercellular signaling, biomarkers, and vectors of cell therapy. Annu Rev Physiol. 2015;77:13–27. doi:10.1146/annurev-physiol-021014-07164125293529
- Mathivanan S, Ji H, Simpson RJ. Exosomes: extracellular organelles important in intercellular communication. J Proteomics. 2010;73(10):1907–1920. doi:10.1016/j.jprot.2010.06.00620601276
- French KC, Antonyak MA, Cerione RA. Extracellular vesicle docking at the cellular port: extracellular vesicle binding and uptake. Semin Cell Dev Biol. 2017;67:48–55. doi:10.1016/j.semcdb.2017.01.00228104520
- Lerner N, Chen I, Schreiber-Avissar S, Beit-Yannai E. Extracellular vesicles mediate anti-oxidative response-in vitro study in the ocular drainage system. Int J Mol Sci. 2020;21(17):6105. doi:10.3390/ijms21176105
- Li W, Jin L, Cui Y, Nie A, Xie N, Liang G. Bone marrow mesenchymal stem cells-induced exosomal microRNA-486-3p protects against diabetic retinopathy through TLR4/NF-κB axis repression. J Endocrinol Invest. 2020. doi:10.1007/s40618-020-01405-3
- Kurywchak P, Tavormina J, Kalluri R. The emerging roles of exosomes in the modulation of immune responses in cancer. Genome Med. 2018;10(1):23. doi:10.1186/s13073-018-0535-429580275
- Greening DW, Gopal SK, Xu R, Simpson RJ, Chen W. Exosomes and their roles in immune regulation and cancer. Semin Cell Dev Biol. 2015;40:72–81. doi:10.1016/j.semcdb.2015.02.00925724562
- Kim SH, Lechman ER, Bianco N, et al. Exosomes derived from IL-10-treated dendritic cells can suppress inflammation and collagen-induced arthritis. J Immunol. 2005;174(10):6440–6448. doi:10.4049/jimmunol.174.10.644015879146
- Kojima M, Costantini TW, Eliceiri BP, Chan TW, Baird A, Coimbra R. Gut epithelial cell-derived exosomes trigger posttrauma immune dysfunction. J Trauma Acute Care Surg. 2018;84(2):257–264. doi:10.1097/TA.000000000000174829194317
- Whiteside TL. Exosomes and tumor-mediated immune suppression. J Clin Invest. 2016;126(4):1216–1223. doi:10.1172/JCI8113626927673
- Whiteside TL. Tumor-derived exosomes and their role in cancer progression. Adv Clin Chem. 2016;74:103–141. doi:10.1016/bs.acc.2015.12.00527117662
- Knickelbein JE, Liu B, Arakelyan A, et al. Modulation of immune responses by extracellular vesicles from retinal pigment epithelium. Invest Ophthalmol Vis Sci. 2016;57(10):4101–4107. doi:10.1167/iovs.15-1835327537259
- Chen B, Li Q, Zhao B, Wang Y. Stem cell-derived extracellular vesicles as a novel potential therapeutic tool for tissue repair. Stem Cells Transl Med. 2017;6(9):1753–1758. doi:10.1002/sctm.16-047728653443
- Bjorge IM, Kim SY, Mano JF, Kalionis B, Chrzanowski W. Extracellular vesicles, exosomes and shedding vesicles in regenerative medicine - a new paradigm for tissue repair. Biomater Sci. 2017;6(1):60–78. doi:10.1039/c7bm00479f29184934
- Yue Y, Garikipati VNS, Verma SK, Goukassian DA, Kishore R. Interleukin-10 deficiency impairs reparative properties of bone marrow-derived endothelial progenitor cell exosomes. Tissue Eng Part A. 2017;23(21–22):1241–1250. doi:10.1089/ten.TEA.2017.008428471299
- Zhang S, Chuah SJ, Lai RC, Hui JHP, Lim SK, Toh WS. MSC exosomes mediate cartilage repair by enhancing proliferation, attenuating apoptosis and modulating immune reactivity. Biomaterials. 2018;156:16–27. doi:10.1016/j.biomaterials.2017.11.02829182933
- Hyung S, Jeong J, Shin K, et al. Exosomes derived from chemically induced human hepatic progenitors inhibit oxidative stress induced cell death. Biotechnol Bioeng. 2020;117(9):2658–2667. doi:10.1002/bit.2744732484909
- Hu C, Li L. Improvement of mesenchymal stromal cells and their derivatives for treating acute liver failure. J Mol Med (Berl). 2019;97(8):1065–1084. doi:10.1007/s00109-019-01804-x31197403
- Nojima H, Freeman CM, Schuster RM, et al. Hepatocyte exosomes mediate liver repair and regeneration via sphingosine-1-phosphate. J Hepatol. 2016;64(1):60–68. doi:10.1016/j.jhep.2015.07.03026254847
- Atienzar-Aroca S, Flores-Bellver M, Serrano-Heras G, et al. Oxidative stress in retinal pigment epithelium cells increases exosome secretion and promotes angiogenesis in endothelial cells. J Cell Mol Med. 2016;20(8):1457–1466. doi:10.1111/jcmm.1283426999719
- Barile L, Vassalli G. Exosomes: therapy delivery tools and biomarkers of diseases. Pharmacol Ther. 2017;174:63–78. doi:10.1016/j.pharmthera.2017.02.02028202367
- Li Y, Zheng Q, Bao C, et al. Circular RNA is enriched and stable in exosomes: a promising biomarker for cancer diagnosis. Cell Res. 2015;25(8):981–984. doi:10.1038/cr.2015.8226138677
- Xiao Y, Li Y, Yuan Y, et al. The potential of exosomes derived from colorectal cancer as a biomarker. Clin Chim Acta. 2019;490:186–193. doi:10.1016/j.cca.2018.09.00730194933
- Melo SA, Luecke LB, Kahlert C, et al. Glypican-1 identifies cancer exosomes and detects early pancreatic cancer. Nature. 2015;523(7559):177–182. doi:10.1038/nature1458126106858
- Brinton LT, Sloane HS, Kester M, Kelly KA. Formation and role of exosomes in cancer. Cell Mol Life Sci. 2015;72(4):659–671. doi:10.1007/s00018-014-1764-325336151
- Hesari A, Golrokh Moghadam SA, Siasi A, et al. Tumor-derived exosomes: potential biomarker or therapeutic target in breast cancer? J Cell Biochem. 2018;119(6):4236–4240. doi:10.1002/jcb.2636428833502
- Skotland T, Ekroos K, Kauhanen D, et al. Molecular lipid species in urinary exosomes as potential prostate cancer biomarkers. Eur J Cancer. 2017;70:122–132. doi:10.1016/j.ejca.2016.10.01127914242
- Dismuke WM, Challa P, Navarro I, Stamer WD, Liu Y. Human aqueous humor exosomes. Exp Eye Res. 2015;132:73–77. doi:10.1016/j.exer.2015.01.01925619138
- Liu Y, Bailey JC, Helwa I, et al. A common variant in MIR182 is associated with primary open-angle glaucoma in the NEIGHBORHOOD consortium. Invest Opthalmol Visual Sci. 2016;57(10):4528. doi:10.1167/iovs.16-19688
- Zhao Y, Weber SR, Lease J, et al. Liquid biopsy of vitreous reveals an abundant vesicle population consistent with the size and morphology of exosomes. Transl Vis Sci Technol. 2018;7(3):6. doi:10.1167/tvst.7.3.6
- Klingeborn M, Dismuke WM, Skiba NP, Kelly U, Stamer WD, Bowes Rickman C. Directional exosome proteomes reflect polarity-specific functions in retinal pigmented epithelium monolayers. Sci Rep. 2017;7(1):4901. doi:10.1038/s41598-017-05102-928687758
- Kang GY, Bang JY, Choi AJ, et al. Exosomal proteins in the aqueous humor as novel biomarkers in patients with neovascular age-related macular degeneration. J Proteome Res. 2014;13(2):581–595. doi:10.1021/pr400751k24400796
- Klingeborn M, Skiba NP, Stamer WD, Bowes Rickman C. Isolation of retinal exosome biomarkers from blood by targeted immunocapture. Adv Exp Med Biol. 2019;1185:21–25. doi:10.1007/978-3-030-27378-1_431884583
- Luan X, Sansanaphongpricha K, Myers I, Chen H, Yuan H, Sun D. Engineering exosomes as refined biological nanoplatforms for drug delivery. Acta Pharmacol Sin. 2017;38(6):754–763. doi:10.1038/aps.2017.1228392567
- Ran N, Gao X, Dong X, et al. Effects of exosome-mediated delivery of myostatin propeptide on functional recovery of mdx mice. Biomaterials. 2020;236:119826. doi:10.1016/j.biomaterials.2020.11982632028167
- Gao X, Ran N, Dong X, et al. Anchor peptide captures, targets, and loads exosomes of diverse origins for diagnostics and therapy. Sci Transl Med. 2018;10(444):eaat0195. doi:10.1126/scitranslmed.aat019529875202
- Sterzenbach U, Putz U, Low LH, Silke J, Tan SS, Howitt J. Engineered exosomes as vehicles for biologically active proteins. Mol Ther. 2017;25(6):1269–1278. doi:10.1016/j.ymthe.2017.03.03028412169
- Chen R, Huang H, Liu H, et al. Friend or Foe? Evidence indicates endogenous exosomes can deliver functional gRNA and Cas9 protein. Small. 2019;15(38):e1902686. doi:10.1002/smll.20190268631271518
- Wang QL, Zhuang X, Sriwastva MK, et al. Blood exosomes regulate the tissue distribution of grapefruit-derived nanovector via CD36 and IGFR1 pathways. Theranostics. 2018;8(18):4912–4924. doi:10.7150/thno.2760830429877
- Wassmer SJ, Carvalho LS, Gyorgy B, Vandenberghe LH, Maguire CA. Exosome-associated AAV2 vector mediates robust gene delivery into the murine retina upon intravitreal injection. Sci Rep. 2017;7:45329. doi:10.1038/srep4532928361998
- Zhang W, Wang Y, Kong Y. Exosomes derived from mesenchymal stem cells modulate miR-126 to ameliorate hyperglycemia-induced retinal inflammation via targeting HMGB1. Invest Ophthalmol Vis Sci. 2019;60(1):294–303. doi:10.1167/iovs.18-2561730657854
- Fu W, Lei C, Liu S, et al. CAR exosomes derived from effector CAR-T cells have potent antitumour effects and low toxicity. Nat Commun. 2019;10(1):4355. doi:10.1038/s41467-019-12321-331554797
- Chen L, Feng Z, Yue H, et al. Exosomes derived from HIV-1-infected cells promote growth and progression of cancer via HIV TAR RNA. Nat Commun. 2018;9(1):4585. doi:10.1038/s41467-018-07006-230389917
- Barclay RA, Schwab A, DeMarino C, et al. Exosomes from uninfected cells activate transcription of latent HIV-1. J Biol Chem. 2017;292(28):11682–11701. doi:10.1074/jbc.M117.79352128536264
- Cho YE, Seo W, Kim DK, et al. Exogenous exosomes from mice with acetaminophen-induced liver injury promote toxicity in the recipient hepatocytes and mice. Sci Rep. 2018;8(1):16070. doi:10.1038/s41598-018-34309-730375433
- Gao M, Dong Z, Sun J, et al. Liver-derived exosome-laden lncRNA MT1DP aggravates cadmium-induced nephrotoxicity. Environ Pollut. 2020;258:113717. doi:10.1016/j.envpol.2019.11371731864927
- Sardar Sinha M, Ansell-Schultz A, Civitelli L, et al. Alzheimer’s disease pathology propagation by exosomes containing toxic amyloid-beta oligomers. Acta Neuropathol. 2018;136(1):41–56. doi:10.1007/s00401-018-1868-129934873
- Huang C, Fisher KP, Hammer SS, Navitskaya S, Blanchard GJ, Busik JV. Plasma exosomes contribute to microvascular damage in diabetic retinopathy by activating the classical complement pathway. Diabetes. 2018;67(8):1639–1649. doi:10.2337/db17-158729866771
- Ke Y, Fan X, Rui H, et al. Exosomes derived from RPE cells under oxidative stress mediate inflammation and apoptosis of normal RPE cells through Apaf1/caspase-9 axis. J Cell Biochem. 2020;121(12):4849–4861. doi:10.1002/jcb.29713
- Sies H. Oxidative stress: a concept in redox biology and medicine. Redox Biol. 2015;4:180–183. doi:10.1016/j.redox.2015.01.00225588755
- Chettimada S, Lorenz DR, Misra V, et al. Exosome markers associated with immune activation and oxidative stress in HIV patients on antiretroviral therapy. Sci Rep. 2018;8(1):7227. doi:10.1038/s41598-018-25515-429740045
- Xiao J, Pan Y, Li XH, et al. Cardiac progenitor cell-derived exosomes prevent cardiomyocytes apoptosis through exosomal miR-21 by targeting PDCD4. Cell Death Dis. 2016;7(6):e2277. doi:10.1038/cddis.2016.18127336721
- Saeed-Zidane M, Linden L, Salilew-Wondim D, et al. Cellular and exosome mediated molecular defense mechanism in bovine granulosa cells exposed to oxidative stress. PLoS One. 2017;12(11):e0187569. doi:10.1371/journal.pone.018756929117219
- Hanus J, Anderson C, Wang S. RPE necroptosis in response to oxidative stress and in AMD. Ageing Res Rev. 2015;24(Pt B):286–298. doi:10.1016/j.arr.2015.09.00226369358
- Ung L, Pattamatta U, Carnt N, Wilkinson-Berka JL, Liew G, White AJR. Oxidative stress and reactive oxygen species: a review of their role in ocular disease. Clin Sci (Lond). 2017;131(24):2865–2883. doi:10.1042/CS2017124629203723
- Datta S, Cano M, Ebrahimi K, Wang L, Handa JT. The impact of oxidative stress and inflammation on RPE degeneration in non-neovascular AMD. Prog Retin Eye Res. 2017;60:201–218. doi:10.1016/j.preteyeres.2017.03.00228336424
- Stitt AW, Curtis TM, Chen M, et al. The progress in understanding and treatment of diabetic retinopathy. Prog Retin Eye Res. 2016;51:156–186. doi:10.1016/j.preteyeres.2015.08.00126297071
- Wong TY, Cheung CM, Larsen M, Sharma S, Simo R. Diabetic retinopathy. Nat Rev Dis Primers. 2016;2:16012. doi:10.1038/nrdp.2016.1227159554
- Walker R. Diabetic retinopathy: protecting the vision of people with diabetes. Br J Community Nurs. 2004;9(12):545–547. doi:10.12968/bjcn.2004.9.12.1724515655491
- Mu H, Wang Y, Wei H, et al. Collagen peptide modified carboxymethyl cellulose as both antioxidant drug and carrier for drug delivery against retinal ischaemia/reperfusion injury. J Cell Mol Med. 2018;22(10):5008–5019. doi:10.1111/jcmm.1376830030883
- Liu L, Sun Q, Wang R, et al. Methane attenuates retinal ischemia/reperfusion injury via anti-oxidative and anti-apoptotic pathways. Brain Res. 2016;1646:327–333. doi:10.1016/j.brainres.2016.05.03727208496
- Moisseiev E, Anderson JD, Oltjen S, et al. Protective effect of intravitreal administration of exosomes derived from mesenchymal stem cells on retinal ischemia. Curr Eye Res. 2017;42(10):1358–1367. doi:10.1080/02713683.2017.131949128636406
- Tokarz A, Szuścik I, Kuśnierz-Cabala B, et al. Extracellular vesicles participate in the transport of cytokines and angiogenic factors in diabetic patients with ocular complications. Folia Med Cracov. 2015;55(4):35–48.
- Maisto R, Trotta MC, Petrillo F, et al. Resolvin D1 modulates the intracellular VEGF-related miRNAs of retinal photoreceptors challenged with high glucose. Front Pharmacol. 2020;11:235. doi:10.3389/fphar.2020.0023532210819
- Liu C, Ge HM, Liu BH, et al. Targeting pericyte-endothelial cell crosstalk by circular RNA-cPWWP2A inhibition aggravates diabetes-induced microvascular dysfunction. Proc Natl Acad Sci U S A. 2019;116(15):7455–7464. doi:10.1073/pnas.181487411630914462
- Zhang W, Dong X, Wang T, Kong Y. Exosomes derived from platelet-rich plasma mediate hyperglycemia-induced retinal endothelial injury via targeting the TLR4 signaling pathway. Exp Eye Res. 2019;189:107813. doi:10.1016/j.exer.2019.10781331560926
- Kamalden TA, Macgregor-Das AM, Kannan SM, et al. Exosomal MicroRNA-15a transfer from the pancreas augments diabetic complications by inducing oxidative stress. Antioxid Redox Signal. 2017;27(13):913–930. doi:10.1089/ars.2016.684428173719
- Katome T, Namekata K, Mitamura Y, et al. Expression of intraocular peroxisome proliferator-activated receptor gamma in patients with proliferative diabetic retinopathy. J Diabetes Complications. 2015;29(2):275–281. doi:10.1016/j.jdiacomp.2014.10.01025468312
- Hsu MY, Chiu CC, Wang JY, et al. Paper-based microfluidic platforms for understanding the role of exosomes in the pathogenesis of major blindness-threatening diseases. Nanomaterials (Basel). 2018;5. doi:10.3390/nano8050310
- Tong Y, Zhou YL, Wang YX, Zhao PQ, Wang ZY. Retinal pigment epithelium cell-derived exosomes: possible relevance to CNV in wet-age related macular degeneration. Med Hypotheses. 2016;97:98–101. doi:10.1016/j.mehy.2016.10.02727876140
- Shah N, Ishii M, Brandon C, et al. Extracellular vesicle-mediated long-range communication in stressed retinal pigment epithelial cell monolayers. Biochimica Et Biophysica Acta Mol Basis Dis. 2018;1864(8):2610–2622. doi:10.1016/j.bbadis.2018.04.016
- Hajrasouliha AR, Jiang G, Lu Q, et al. Exosomes from retinal astrocytes contain antiangiogenic components that inhibit laser-induced choroidal neovascularization. J Biol Chem. 2013;288(39):28058–28067. doi:10.1074/jbc.M113.47076523926109
- Atienzar-Aroca S, Serrano-Heras G, Freire Valls A, et al. Role of retinal pigment epithelium-derived exosomes and autophagy in new blood vessel formation. J Cell Mol Med. 2018;22(11):5244–5256. doi:10.1111/jcmm.1373030133118
- Wang AL, Lukas TJ, Yuan M, Du N, Tso MO, Neufeld AH. Autophagy, exosomes and drusen formation in age-related macular degeneration. Autophagy. 2009;5(4):563–564. doi:10.4161/auto.5.4.816319270489
- Aboul Naga SH, Dithmer M, Chitadze G, et al. Intracellular pathways following uptake of bevacizumab in RPE cells. Exp Eye Res. 2015;131:29–41. doi:10.1016/j.exer.2014.12.01025533679
- Elbay A, Ercan Ç, Akbaş F, Bulut H, Ozdemir H. Three new circulating microRNAs may be associated with wet age-related macular degeneration. Scand J Clin Lab Invest. 2019;79(6):388–394. doi:10.1080/00365513.2019.163793131277558
- Biasutto L, Chiechi A, Couch R, Liotta LA, Espina V. Retinal pigment epithelium (RPE) exosomes contain signaling phosphoproteins affected by oxidative stress. Exp Cell Res. 2013;319(13):2113–2123. doi:10.1016/j.yexcr.2013.05.00523669273
- Bai L, Shao H, Wang H, et al. Effects of mesenchymal stem cell-derived exosomes on experimental autoimmune uveitis. Sci Rep. 2017;7(1):4323. doi:10.1038/s41598-017-04559-y28659587
- Harrell CR, Simovic Markovic B, Fellabaum C, et al. Therapeutic potential of mesenchymal stem cell-derived exosomes in the treatment of eye diseases. Adv Exp Med Biol. 2018;1089:47–57. doi:10.1007/5584_2018_21929774506
- Davis BM, Crawley L, Pahlitzsch M, Javaid F, Cordeiro MF. Glaucoma: the retina and beyond. Acta Neuropathol. 2016;132(6):807–826. doi:10.1007/s00401-016-1609-227544758
- Razeghinejad MR, Myers JS, Katz LJ. Iatrogenic glaucoma secondary to medications. Am J Med. 2011;124(1):20–25. doi:10.1016/j.amjmed.2010.08.01121092926
- Weinreb RN, Aung T, Medeiros FA. The pathophysiology and treatment of glaucoma: a review. JAMA. 2014;311(18):1901–1911. doi:10.1001/jama.2014.319224825645
- Casson RJ, Chidlow G, Wood JP, Crowston JG, Goldberg I. Definition of glaucoma: clinical and experimental concepts. Clin Exp Ophthalmol. 2012;40(4):341–349. doi:10.1111/j.1442-9071.2012.02773.x22356435
- Almasieh M, Wilson A, Morquette B, Cueva Vargas J, Di Polo A. The molecular basis of retinal ganglion cell death in glaucoma. Prog Retin Eye Res. 2012;31(2):152–181. doi:10.1016/j.preteyeres.2011.11.00222155051
- Harrell CR, Fellabaum C, Arsenijevic A, Markovic BS, Djonov V, Volarevic V. Therapeutic potential of mesenchymal stem cells and their secretome in the treatment of glaucoma. Stem Cells Int. 2019;2019:1–11. doi:10.1155/2019/7869130
- Lerner N, Schreiber-Avissar S, Beit-Yannai E. Extracellular vesicle-mediated crosstalk between NPCE cells and TM cells result in modulation of Wnt signalling pathway and ECM remodelling. J Cell Mol Med. 2020;24(8):4646–4658. doi:10.1111/jcmm.1512932168427
- Lerner N, Avissar S, Beit-Yannai E. Extracellular vesicles mediate signaling between the aqueous humor producing and draining cells in the ocular system. PLoS One. 2017;12(2):e0171153. doi:10.1371/journal.pone.017115328241021
- Liu Y, Allingham RR, Qin X, et al. Gene expression profile in human trabecular meshwork from patients with primary open-angle glaucoma. Invest Opthalmol Visual Sci. 2013;54(9):6382. doi:10.1167/iovs.13-12128
- Stamer WD, Hoffman EA, Luther JM, Hachey DL, Schey KL. Protein profile of exosomes from trabecular meshwork cells. J Proteomics. 2011;74(6):796–804. doi:10.1016/j.jprot.2011.02.02421362503
- Mead B, Ahmed Z, Tomarev S. Mesenchymal stem cell-derived small extracellular vesicles promote neuroprotection in a genetic DBA/2J mouse model of glaucoma. Invest Ophthalmol Vis Sci. 2018;59(13):5473–5480. doi:10.1167/iovs.18-2531030452601
- Pan D, Chang X, Xu M, et al. UMSC-derived exosomes promote retinal ganglion cells survival in a rat model of optic nerve crush. J Chem Neuroanat. 2019;96:134–139. doi:10.1016/j.jchemneu.2019.01.00630639447
- Coulson-Thomas VJ, Caterson B, Kao WWY. Transplantation of human umbilical mesenchymal stem cells cures the corneal defects of mucopolysaccharidosis VII mice. Stem Cells. 2013;31(10):2116–2126. doi:10.1002/stem.148123897660
- Han KY, Dugas-Ford J, Seiki M, Chang JH, Azar DT. Evidence for the Involvement of MMP14 in MMP2 processing and recruitment in exosomes of corneal fibroblasts. Invest Ophthalmol Vis Sci. 2015;56(9):5323–5329. doi:10.1167/iovs.14-1441725015352
- Kong L, Fry M, Al-Samarraie M, Gilbert C, Steinkuller PG. An update on progress and the changing epidemiology of causes of childhood blindness worldwide. J AAPOS. 2012;16(6):501–507. doi:10.1016/j.jaapos.2012.09.00423237744
- Fulton AB, Hansen RM, Moskowitz A, Akula JD. The neurovascular retina in retinopathy of prematurity. Prog Retin Eye Res. 2009;28(6):452–482. doi:10.1016/j.preteyeres.2009.06.00319563909
- Hansen RM, Moskowitz A, Akula JD, Fulton AB. The neural retina in retinopathy of prematurity. Prog Retin Eye Res. 2017;56:32–57. doi:10.1016/j.preteyeres.2016.09.00427671171
- Hellström A, Smith LEH, Dammann O. Retinopathy of prematurity. The Lancet. 2013;382(9902):1445–1457. doi:10.1016/s0140-6736(13)60178-6
- Xu W, Wu Y, Hu Z, et al. Exosomes from microglia attenuate photoreceptor injury and neovascularization in an animal model of retinopathy of prematurity. Mol Ther Nucleic Acids. 2019;16:778–790. doi:10.1016/j.omtn.2019.04.02931163320
- Salter MW, Beggs S. Sublime microglia: expanding roles for the guardians of the CNS. Cell. 2014;158(1):15–24. doi:10.1016/j.cell.2014.06.00824995975
- Ebneter A, Kokona D, Schneider N, Zinkernagel MS. Microglia activation and recruitment of circulating macrophages during ischemic experimental branch retinal vein occlusion. Invest Ophthalmol Vis Sci. 2017;58(2):944–953. doi:10.1167/iovs.16-2047428170538
- Lande K, Gupta J, Ranjan R, et al. Exosomes: insights from retinoblastoma and other eye cancers. Int J Mol Sci. 2020;21(19). doi:10.3390/ijms21197055
- Chattopadhyay C, Kim DW, Gombos DS, et al. Uveal melanoma: from diagnosis to treatment and the science in between. Cancer. 2016;122(15):2299–2312. doi:10.1002/cncr.2972726991400
- Carvajal RD, Schwartz GK, Tezel T, Marr B, Francis JH, Nathan PD. Metastatic disease from uveal melanoma: treatment options and future prospects. Br J Ophthalmol. 2017;101(1):38–44. doi:10.1136/bjophthalmol-2016-30903427574175
- Eldh M, Olofsson Bagge R, Lässer C, et al. MicroRNA in exosomes isolated directly from the liver circulation in patients with metastatic uveal melanoma. BMC Cancer. 2014;14:962. doi:10.1186/1471-2407-14-96225510783
- Ragusa M, Barbagallo C, Statello L, et al. miRNA profiling in vitreous humor, vitreal exosomes and serum from uveal melanoma patients: pathological and diagnostic implications. Cancer Biol Ther. 2015;16(9):1387–1396. doi:10.1080/15384047.2015.104602125951497