
Abstract
Breast cancer stem cells (BCSCs), also known as breast cancer initiating cells, are reported to be responsible for the initiation, progression, therapeutic resistance, and relapse of breast cancer. Conventional therapeutic agents mainly kill the bulk of breast tumor cells and fail to eliminate BCSCs, even enhancing the fraction of BCSCs in breast tumors sometimes. Therefore, it is essential to develop specific and effective methods of eliminating BCSCs that will enhance the efficacy of killing breast tumor cells and thereby, increase the survival rates and quality of life of breast cancer patients. Despite the availability of an increasing number of anti-BCSC agents, their clinical translations are hindered by many issues, such as instability, low bioavailability, and off-target effects. Nanosized drug delivery systems (NDDSs) have the potential to overcome the drawbacks of anti-BCSC agents by providing site-specific delivery and enhancing of the stability and bioavailability of the delivered agents. In this review, we first briefly introduce the strategies and agents used against BCSCs and then highlight the mechanism of action and therapeutic efficacy of several state-of-the-art NDDSs that can be used to treat breast cancer by eliminating BCSCs.
Introduction
Despite many advances in breast cancer therapy with the development of drugs targeting breast cancer cells, breast cancer still remains one of the major causes of patient deaths worldwide.Citation1 Therapeutic resistance, recurrence, and metastasis are the leading challenges in breast cancer treatment.Citation2 Accumulating evidence has demonstrated that breast cancer stem cells (BCSCs), also known as breast cancer initiating cells, are responsible for the poor prognosis of breast cancer; as they play key roles in the initiation, progression, therapeutic resistance, and recurrence of breast cancer.Citation3–Citation8 BCSCs possess specific markers that distinguish them from bulk tumor cells, such as the high expression of surface antigen cluster of differentiation 44/low or negative expression of cluster of differentiation 24 (CD44+/CD24low/-),Citation9 high expression of CD133 (CD133+)Citation10 and positive expression of aldehyde dehydrogenase 1 (ALDH1+).Citation11 Moreover, BCSCs are intrinsically drug resistant and often display high expression levels of drug efflux transporters and over-activation of anti-apoptotic signaling pathways.Citation12–Citation14 Besides, breast cancer treatment with conventional chemotherapy or radiotherapy can kill only bulk tumor cells and fail to eliminate BCSCs, possibly even enhancing the fraction of BCSCs in breast tumors. These residual BCSCs will become cancer cells in the future, and consequently lead to tumor recurrence.Citation15–Citation17 In addition, BCSCs facilitate the metastasis of breast cancer by inducing the transformation of epithelial cancer cells to mesenchymal cancer cells, allowing them to spread easily to distant lesions.Citation18 Therefore, it is crucial to eliminate BCSCs to enhance the efficacy of breast cancer treatment.
In recent years, many strategies have been proposed to eradicate BCSCs by blocking the signaling pathways related to BCSCs self-renewal such as Wnt/β-catenin,Citation19 Hedgehog (Hh),Citation20 Notch,Citation21 HippoCitation22 and transforming growth factor-β (TGF-β).Citation23 Blockage of these pathways is designed to inhibit the proliferation and growth of breast tumors, target the breast tumor microenvironment to destroy the communication between BCSCs and cytokines,Citation24–Citation26 target the BCSC surface markers to locate and destroy BCSCs,Citation27–Citation30 and interfere with the differentiationCitation31 or metabolismCitation32,Citation33 of BCSCs to render them more sensitive to conventional therapy. An increasing number of anti-BCSC agents have been proposed to treat breast cancer based on the above strategies, such as quercetin,Citation34 sulforaphane,Citation35 curcumin,Citation36 salinomycin (SAL),Citation37 nuclear factor-kappa B (NF-κB) short hairpin ribonucleic acid (shRNA),Citation38 octamer 4 (Oct-4) small interfering RNA (siRNA),Citation39 and microRNA-100 (miR-100).Citation6 However, similar to conventional chemotherapeutic drugs, most currently reported anti-BCSC agents have disadvantages such as poor solubility, low stability, high toxicity, unfavorable pharmacokinetics, and lack of tissue selective distribution,Citation3 that restrict their clinical applications. In addition, these agents are potentially toxic to normal stem cells as BCSCs share properties with normal stem cells and conventional anti-BCSC agents cannot distinguish them from normal stem cells.
Because of their site-specific delivery and enhanced drug stabilization, nanosized drug delivery systems (NDDSs) have shown significant promise in the delivery of anti-BCSC agents and have the potential to overcome the limitations of conventional anti-BCSC agents mentioned above. There are an increasing number of NDDSs used to deliver anti-BCSC agents, including polymeric nanoparticles,Citation40 micelles,Citation41 liposomes,Citation42 nanocomplexes,Citation38 nanoprodrugs,Citation43 aptamer–conjugated deoxyribonucleic acid (DNA) nanotrains,Citation16 single-walled carbon nanotube (SWCNT) nanocarriers,Citation44 nanoexosomes,Citation45 lipid–polymer hybrid nanoparticles,Citation46 and nanocages.Citation47 However, the clinical translation of these NDDSs is tough, as many challenges still remain unaddressed. In this review, we briefly introduce the strategies and agents used against BCSCs and then highlight the mechanism of action and therapeutic efficacy of several state-of-the-art NDDSs that can be used to treat breast cancer by eliminating BCSCs.
Current Treatment Strategies Against Breast Cancer by Inhibition of BCSCs
A growing body of research has shown that BCSCs account for breast cancer initiation, progression, recurrence, and therapeutic resistance. In addition, BCSCs can self-renew and give rise to non-tumorigenic cancer cells.Citation48,Citation49 Therefore, it is necessary to completely eliminate BCSCs to successfully eradicate breast cancer. In recent years, many strategies to treat breast cancer by targeting BCSCs have been proposed, such as targeting BCSC surface markers,Citation27,Citation28,Citation50 inhibiting BCSC-dependent signaling pathways,Citation19–Citation21,Citation23 interfering with BCSC differentiation,Citation31,Citation51 targeting metabolism in BCSCs,Citation52–Citation54 and targeting the breast tumor microenvironment.Citation55,Citation56 Therapeutic strategies against breast cancer according to the characteristics of BCSCs are shown in .
Figure 1 Strategies against breast cancer stem cells: (1) targeting BCSC surface markers; (2) inhibition of BCSC-dependent signaling pathways; (3) interfering the BCSC differentiation; (4) targeting metabolisms in BCSCs; (5) targeting the breast tumor microenvironments.
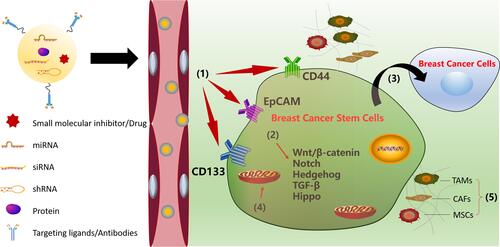
Targeting BCSC Surface Markers
The surface markers on cancer stem cells play a crucial role in the isolation, identification, diagnosis, and targeted therapy of cancer stem cells. Commonly used surface markers of BCSCs include CD44, CD133, and epithelial cell adhesion molecule (EpCAM). CD44 is a transmembrane protein that has been identified in many cancer stem cells, including BCSCs, and it plays a very important role in regulating the properties of BCSCs that involve self-renewal, tumor initiation, therapeutic resistance, and metastasis.Citation27,Citation50 The characteristic CD44 overexpression in BCSCs indicates that CD44 is a potential target in the treatment of BCSCs. Therefore, anti-CD44 with monoclonal antibodies or ligands may be promising strategies for eliminating BCSCs. P245, an anti-CD44 antibody, has been demonstrated to inhibit breast cancer growth and eliminate BCSCs in xenograft mouse models.Citation57 Treatment with P245 prevented tumor recurrence in human breast cancer xenografts after treatment with doxorubicin (DOX) and cyclophosphamide.Citation57 CD133, also known as prominin-1, is a five-transmembrane glycoprotein that is overexpressed in several types of cancers, such as breast cancer, ovarian cancer, and gastric carcinoma.Citation28,Citation58 As one among several surface markers of BCSCs, CD133 is critical for the survival and growth of BCSCs, and antibodies against CD133 can reduce the growth of BCSCs. It was demonstrated that the fusion protein dCD133KDEL represents a new biological assessment tool that can be used to determine the clinical significance of eradicating CD133+ cells.Citation59 EpCAM (also known as CD326/ESA), a glycoprotein, is another molecular target of BCSCs, and its overexpression may facilitate the proliferation, metastasis, and drug resistance of breast cancer cells.Citation29,Citation30 Kubo et alCitation30 found that catumaxomab, an EpCAM antibody, combined with activated T-cells could eliminate EpCAM-positive triple-negative breast cancer cells and overcome the chemoresistance induced by these cells in vitro.
Inhibition of BCSC-Dependent Signaling Pathways
Dysregulation of signaling pathways plays an important role in rendering BCSCs capable of maintaining stem cell characteristics and facilitates the role of BCSCs in the development and progression of breast tumors.Citation60,Citation61 The major signaling pathways related to the maintenance, self-renewal, survival, and differentiation of BCSCs are Wnt/β-catenin,Citation19 Hh,Citation20 Notch,Citation21 HippoCitation22 and TGF-β.Citation23 Blocking these signaling pathways using inhibitors may be a potential strategy for BCSC-targeted therapies.
Wnt/β-Catenin Pathway
The Wnt/β-catenin signaling pathway regulates many physiological processes, such as self-renewal, growth, and regeneration of cells.Citation62 After a Wnt ligand binds to a Wnt receptor, such as heterodimeric frizzled-7 (Fzd7) or low-density lipoprotein receptor-related protein 6 (LRP6), the Wnt pathway is activated and the signal is transferred to β-catenin through several downstream processes. Upon activation, the dephosphorylated β-catenin enters the nucleus to activate Wnt target genes.Citation63 This signaling pathway is upregulated in many cancers, including breast cancer, and is considered to be a key factor in the maintenance and self-renewal of BCSCs. Thus, selective targeting of Wnt/β-catenin signaling may be a strategy to eliminate BCSCs and reduce breast cancer aggressiveness. For instance, Jang et alCitation7 demonstrated that Wnt/β-catenin signaling is relatively more active in BCSCs than in bulk tumor cells, which results in the therapeutic resistance of BCSCs. These investigators also designed CWP232228 (a small-molecule inhibitor) to antagonize β-catenin that binds to T-cell factor in the nucleus. Their results showed that CWP232228 can inhibit the growth of both BCSCs and bulk tumor cells. However, an interesting observation was that the BCSCs exhibited decreased growth than the bulk tumor cells, indicating that CWP23228 has some degree of selectivity toward BCSCs. In addition to small-molecule inhibitors, macromolecular agents such as antibodies have been reported to block the Wnt/β-catenin pathway. For example, Gurney et alCitation64 reported that vantictumab (a monoclonal antibody) inhibits tumor growth and reduces tumor-initiating cell frequency by interacting with five Fzd receptors to block canonical Wnt signaling.
Notch Signaling Pathway
An activated pathway in breast cancer cells namely, Notch signaling, plays a vital role in stem cell retention and differentiation and has attracted much attention in recent times, as a target to inhibit breast cancer relapse and metastasis by eradicating BCSCs.Citation48,Citation65 The Notch signaling pathway is activated by ligands (eg, Delta-like [DLL]1, 3, and 4; and Jagged [JAG]1 and 2) that binding to Notch receptors (Notch1-4).Citation66,Citation67 This binding results in cleavage of the Notch receptor by the enzyme γ-secretase to release the Notch intracellular domain (NICD). The released NICD translocates to the nucleus to activate Notch target genes. Therefore, inhibitors of γ-secretase and antagonists of Notch receptors or ligands have the potential to inhibit Notch activity in BCSCs. For example, Grudzien et alCitation67 reported that BCSCs possess greater Notch signaling than bulk tumor cells, and they demonstrated a reduction in sphere formation, proliferation, and/or colony formation in soft agar by blocking Notch signaling using pharmacological and genomic approaches (eg, by using MRK003, a γ-secretase inhibitor). Moreover, McClements et alCitation68 reported that specific peptides (ALM201 and AD-01) inhibited BCSCs in both ER+ and ER− breast cancer by downregulating DLL4 and Notch 4;Citation68 this study was the first to demonstrate the preclinical systemic activity of ALM201 and AD-01on breast cancer.
Hh Signaling Pathway
The Hh signaling pathway essentially regulates the maintenance, self-renewal, survival, and proliferation of BCSCs.Citation12,Citation69,Citation70 In this pathway, the binding of Hh ligand to the patched receptor of a neighboring cell reduces the inhibition of the transmembrane receptor protein Smoothened (Smo). Smo activation then leads to the release of the glioma-associated oncogene (Gli) family of transcription factors (Gli1/2/3), which undergo nuclear translocation to regulate the expression of Hh target genes. The overexpression of both Smo and Gli has been found in the BCSC subpopulation, therefore, Smo and Gli are potential targets for inhibiting the Hh signaling pathway to eliminate BCSCs.Citation12,Citation71 In this regard, GANT61, a Gli1/2 inhibitor, was reported to decrease the percentage of cancer stem cells and enhance the anti-mitogenic activity of paclitaxel in several triple-negative breast cancer (TNBC) cell lines.Citation72 This result implicated that GANT61 as a potential therapeutic agent in TNBC. In addition, cyclopamine, the first Hh inhibitor to be identified, reduced the growth of breast cancer cells by binding to and inactivating Smo to suppress Gli1 expression.Citation73 These results demonstrate that therapeutic agents with the ability to inhibit Smo and Gli exhibit the potential to reduce the percentage of BCSCs in breast tumor tissue.
Hippo Signaling Pathway
Hippo signaling is regulated by a network of core kinase cascades and is a key regulator of tumorigenesis and stem cell renewal.Citation74 It was demonstrated that the dephosphorylation of yes-associated protein 1 (YAP1) and transcriptional co-activator with PDZ-binding motif (TAZ) results in their nuclear translocation, leading to the activation of Hippo target gene transcription.Citation74 YAP1 and TAZ are reported to be overexpressed in BCSCs of metastatic breast cancer, and thus, could be potential targets for inhibiting the Hippo signaling pathway and reduce the percentage of BCSCs.Citation75 Interestingly, Li et alCitation76 reported that inhibiting the transcriptional activities of YAP1 and TAZ with apigenin, a naturally occurring compound, reduced the stemness of TNBC cells. This indicates that apigenin is a promising therapeutic agent for the treatment of TNBC patients showing high YAP/TAZ activity.
TGF-β Signaling Pathway
TGF-β is the prototype of the TGF-β family of growth and differentiation factors. TGF-β can facilitate the transformation of cancer cells into cancer stem cells by the activation of epithelial–mesenchymal transition (EMT)–inducing transcription factors, resulting in drug resistance.Citation77–Citation80 Li et alCitation78 showed that the pleiotropic effects of TGF-β influence chemotherapeutic drug resistance by modulating EMT, stemness, and apoptosis. TGF-β signaling has also been reported to play a vital role in the maintenance and functioning of BCSCs.Citation62,Citation81 Therefore, targeting TGF-β signaling may be an effective strategy to treat breast cancer by inhibiting BCSCs. It was demonstrated by Liu et alCitation81 that Gd–metallofullerenol-based nanomaterial could eradicate BCSCs by inhibiting TGF-β signaling under normoxic conditions and suppressing both hypoxia-inducible factor (HIF)-1α and TGF-β activities under hypoxic conditions. An even more exciting fact is that the metallofullerenol nanomaterial Gd@C82(OH)22 is essentially nontoxic to normal mammary epithelial cells and can inhibit breast tumor initiation and metastasis by eliminating BCSCs.
Inducing BCSC Differentiation
Differentiation was demonstrated by Warrel et alCitation82 as an effective method of treating acute promyelocytic leukemia. In this study, after the patients were treated with all-trans retinoic acid (ATRA), it was found that their leukemic promyelocytes failed to differentiate into mature granulocytes, indicating that preventing cancer stem cell differentiation maybe useful in the treatment of other cancers. Differentiation therapy targets cancer stem cells and alters their stemness to reduce the therapeutic resistance of cancer. Sun et alCitation31 demonstrated that a drug delivery system encapsulating ATRA and DOX effectively suppressed breast cancer by inducing BCSCs to differentiate into non-BCSCs, reducing their tumor initiation abilities and enhancing their sensitivity to DOX under the effect of ATRA. Pham et alCitation51 showed that the knockdown of CD44 with shRNA using lentivirus particles differentiated BCSCs into non-BCSCs, increasing their susceptibility to both chemotherapy and radiation. This indicated that CD44 knockdown is an effective strategy to eliminate the stemness of BCSCs and can be a potential strategy to treat breast cancer by targeting these cells.
Targeting Metabolism in BCSCs
Cancer stem cells exhibit specific metabolic properties, such as the metabolism of glucose and mevalonate.Citation32,Citation52,Citation70 For example, hexokinase 2 (HK2), a very important kinase involved in glucose metabolism, is overexpressed in BCSCs.Citation33,Citation70 Thus, inhibiting HK2 is a potential method to eradicate BCSCs. In studies conducted previously, metformin (MET) has displayed anti-BCSC activity and enhanced the efficacy of chemotherapy in breast cancer by inhibiting HK2.Citation53 Moreover, inhibiting the mevalonate metabolic pathway with hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase blockers apparently suppressed the growth of BCSCs.Citation32 As another example, SAL was reported to reduce the stemness of BCSCs by inhibiting the Wnt pathway, which is a known regulator of cell metabolism.Citation83 It has also been reported that iron metabolism plays an essential role in cancer stem cells; therefore targeting iron metabolism may improve the therapeutic effect against many cancers, including breast cancer.Citation54 These examples demonstrate that the characteristic metabolic properties of BCSCs could be used as targets to treat them.
Targeting the Tumor Microenvironment
The microenvironment in which BCSCs are located plays an essential role in maintaining the functions of these cells.Citation24,Citation84 This specific microenvironment is regulated by many factors such as mesenchymal stem cells (MSCs), immune cells, cancer-associated fibroblasts (CAFs), autocrine signals, the extracellular matrix and vascular network, oxygen pressure, and nutrient levels. In addition, this microenvironment can generate BCSCs by inducing the characteristics of cancer stem cells in non-BCSC.Citation55 It was reported that MSCs in breast tissue could expand from bone marrow-derived MSCs to regulate BCSCs by cytokine loops involving interleukin (IL)-6 and C-X-C motif ligand 7 (CXCL7) to expedite the growth of breast cancer.Citation85 CAFs were found to produce high levels of chemokine (C-C motif) ligand 2 (CCL2) to stimulate stem cell-specific features in breast cancer cells,Citation25 and CAFs were also found to regulate BCSCs via IL-6 and IL-8.Citation26 Tumor-associated macrophages (TAMs), a type of immune cell, were reported to facilitate the generation of cancer stem cells by secreting tumor necrosis factor-α (TNF-α) and TGF-β,Citation56 and promote BCSC phenotypes in murine breast cancer cells by affecting the epidermal growth factor receptor (EGFR)/signal transducer and activator of transcription 3 (STAT3)/sex-determining region Y-box 2 (SOX2) signaling pathway.Citation86 Mammary adipose tissues secrete adipokines to augment the features and proliferation of BCSCs.Citation87 Thus, factors affecting the BCSC microenvironment are potential targets for eliminating BCSCs to reduce the relapse, therapeutic resistance, and metastasis of breast cancer.
Agents Against BCSCs
Considering the effects of BCSCs on the initiation, maintenance, development, relapse, therapeutic resistance, and metastasis of breast cancer, many agents have been proposed to eliminate BCSCs. These agents are divided into three categories according to their physicochemical properties: small-molecule inhibitors/drugs, nucleic acid drugs, and protein drugs ().
Table 1 Agents Against BCSCs
Small-Molecule Inhibitors/Drugs
Small-molecule inhibitors/drugs account for the majority of current anti-BCSC agents. Quercetin suppressed the proliferation, self-renewal, and invasiveness of BCSCs in MDA-MB-231 cells by downregulating the expression of ALDH1 family, member A1 (ALDH1A1), C-X-C chemokine receptor type 4 (CXCR4), mucin 1 (MUC1), and EpCAM.Citation34 Sulforaphane inhibited BCSCs both in vitro and in vivo, as demonstrated by a mammosphere formation assay, ALDEFLUOR assay, and secondary tumor growth in mice; one potential mechanism is by downregulation of the Wnt/β-catenin pathway in BCSCs.Citation35 It was reported that curcumin inhibited BCSCs by suppressing both the sonic hedgehog (SHH) and Wnt/β-catenin pathways, which play essential roles in maintaining the stemness of BCSCs.Citation36 A noncanonical Hh inhibitor, GANT61, was reported to downregulate the expression of GLi1, GLi2, and/or SHH to decrease the levels of BCSCs induced by estradiol in ER-positive breast cancer cells.Citation20
Vitamin D compounds (1α,25[OH]2D3; BXL0124) were found to effectively reduce BCSCs in TNBC by repressing Notch1, Notch2, Notch3, JAG1, and JAG2 to inhibit the Notch signaling pathway in BCSCsCitation106 Apigenin suppresses the stemness of BCSCs by targeting YAP and TAZ, two main downstream effectors of the Hippo pathway. This inhibits YAP/TAZ-TEAD complex activity, which is essential for tumor initiation and maintenance of the self-renewal ability of BCSCs.Citation76 Simvastatin, an inhibitor of HMG-CoA reductase, blocks the biosynthesis of mevalonic acid to reduce BCSCs both in vitro and in vivo.Citation32
Chloroquine, an autophagic inhibitor, reduces BCSCs in TNBC by inhibiting the Janus-activated kinase 2 (JAK2)-STAT3 signaling pathway, which sensitizes TNBC stem cells to paclitaxel by inhibiting autophagy.Citation102 By activating protein kinase A, forskolin induces EMT in mammary epithelial cells, which causes them to lose the ability to initiate tumors and sensitizes them (in vitro) to conventional chemotherapeutic agents, such as DOX.Citation93 SAL was shown to inhibit mammary tumor growth and reduce the proportion of BCSCs, and its effect was more than 100 times that of paclitaxel.Citation37 A clinical pilot study by Scatena et alCitation107 found that doxycycline, an Food and Drug Administration (FDA)-approved antibiotic, could reduce BCSC markers (CD44 and ALAH1) and eliminate BCSCs in breast cancer patients by inhibiting the mitochondrial-related proteins that were overexpressed in many cancer stem cells, including BCSCs. However, the authors emphasized that additional clinical studies with larger numbers of patients will be needed to validate this promising pilot study. In general, there have been many reports about small-molecule agents with anti-BCSC capabilities, but most of these studies were in the preclinical stages. More research is needed to confirm the anti-BCSC effects of these agents and reduce the toxicities induced by their nonselective distribution in vivo.
Nucleic Acid Drugs
Gene therapy is a potential method to target BCSCs. In theory, downregulation of BCSC surface markers or blockade of signaling pathways using siRNA, shRNA, or miRNA agents has the potential to suppress the function of BCSCs. For example, NF-κB shRNA was reported to decrease the percentage of ALDH+ BCSCs and mammosphere colony formations.Citation38 Hu et alCitation39 found that Oct-4 gene suppression by Otc-4 siRNA induced BCSC apoptosis via inhibition of the Oct-4/Tcl1/Akt1 signaling pathway. miRNAs, post-transcriptional regulators of various cellular functions, have also been reported as potential anticancer agents. Elevating the expression of miR-100 was shown to decrease the production of BCSCs by attenuating the expression of the cancer stem cell regulatory gene (SWI/SNF-Related Matrix-Associated Actin-Dependent Regulator of Chromatin Subfamily D Member 1, SMARCD1) and bone morphogenetic protein receptor type 2 (BMPR2).Citation6 miR-34a, a tumor-suppressor miRNA, has the capacity to affect the properties of BCSCs and enhance their susceptibility to doxorubicin treatment by targeting Notch1.Citation119
In addition to siRNA, shRNA, and miRNA, gene editing technologies, especially the CRISPR/Cas9 genome-editing system, have generated enormous interest in the field of gene therapy.Citation128,Citation129 This technology provides a robust tool to generate knockout cells or animal models quickly, exhibiting great potential for applications in the treatment of cancer.Citation130 For instance, Wang et alCitation120 knocked out integrin α 9 (ITGA9) in TNBC cells using CRISPR/Cas9 technology and found that ITGA9 knockout noticeably attenuated the properties of BCSCs in TNBC cells and the angiogenesis, growth, and metastasis of tumors by promoting β-catenin degradation. Yang et alCitation121 using CRISPR/Cas9 to knock out the Y-box binding protein 1 (YB-1) gene, found that YB-1 deletion inhibited the proliferation of BCSCs, leading to cell cycle arrest and apoptosis, and induced irreversible differentiation of cancer stem cells. This indicates that YB-1 plays an important role in maintaining the stemness of BCSCs and reverting the differentiated tumor cells back to cancer stem cells.
Protein Drugs
Protein drugs, especially antibodies, are being explored to suppress BCSCs by targeting their surface markers or various signaling pathways. For instance, OMP-185R, a monoclonal antibody, suppressed canonical Wnt signaling by blocking the Fzd receptor family to reduce the growth of many tumors (including breast cancer tumors) and the frequency of tumor-initiating cells.Citation64 P245 mAb, which targets CD44, reduced the growth and prevented recurrence of tumors in human breast cancer xenografts. These effects were believed to be attributed to the induction of antiproliferative cytokines.Citation57 The recombinant protein TmSm (T34A) was demonstrated to prevent the proliferation and growth of BCSCs by downregulating the expression of cyclin D1 and significantly inducing the apoptosis of BCSCs.Citation125
Problems of Current BCSC-Specific Agents
Although obvious progress has been achieved with BCSC-specific agents, some serious challenges still remain. First, similar to conventional chemotherapeutic drugs, BCSC-specific agents may possess characteristics that are undesirable in vivo. For instance, accumulating evidence has demonstrated that miRNA, siRNA, and shRNA have great potential to be used as anti-BCSC agents; however, RNA-based therapies are limited by many obstacles in vivo including their degradation in blood, poor cellular uptake, and potential systemic toxicity, the latter of which results from poor tissue targeting.Citation38,Citation132,Citation133 The clinical applications of certain anti-BCSC agents, such as curcumin and quercetin, have been limited because of their poor absorption and rapid metabolism.Citation34,Citation41,Citation89,Citation134 Second, BCSCs share many characteristics (such as self-renewal and quiescence) with normal stem cells. Moreover, none of the presently reported anti-BCSC agents can distinguish BCSCs from normal stem cells; therefore, these agents are potentially toxic to normal stem cells. For instance, although most γ-secretase inhibitors show anti-BCSC properties, they may concomitantly damage normal stem cells.Citation12,Citation60,Citation63,Citation135,Citation136
NDDSs Against BCSCs
There is an urgent need to solve the problems of current anti-BCSC agents, such as poor solubility, instability, unfavorable biodistribution, and high toxicity induced by off-target effects.Citation3,Citation130,Citation137 NDDSs have the potential to address this need. NDDSs can passively target tumor tissues owing to their enhanced permeability and retention (EPR) effects. Moreover, the BCSC-targeted effects of NDDSs can be further enhanced by their surface modification with suitable ligands that interact with overexpressed receptors on the surface of BCSCs. A deeper understanding of the biology of BCSCs and numerous advances in nanotechnology have resulted in increasing numbers of NDDSs being developed to treat breast cancer by eliminating BCSCs. The delivery strategies of NDDSs against BCSCs mainly include: delivery of anti-BCSC agents to tumors; combinational delivery of chemotherapeutics and anti-BCSC agents to tumors; active-targeted delivery of anti-BCSC agents and/or chemotherapeutics agents to tumors. In this section, NDDS-targeted BCSCs are summarized and categorized in –, according to their cargo delivery and modifications.
Table 2 Delivery of BCSC-Specific Small Molecular Agents
Table 3 Delivery of BCSC-Specific Nucleic Acid Drugs
Table 4 Combinational Delivery of Chemotherapeutics and CSC-Specific Agents
Delivery of Chemotherapeutic Agents to BCSCs
Most properties of the currently reported BCSC-specific chemotherapeutic agents are undesirable in vivo and are similar to those of traditional chemotherapeutic agents. One application of NDDSs is to qualify therapeutic agents and drug candidates; displays an overview of the NDDSs that have been used to deliver BCSC-specific chemotherapeutic agents. For example, curcumin, a polyphenol derived from the ancient Asian spice turmeric, has been reported to target cancer stem cells by downregulating signaling pathways such as Wnt, Notch1, and NF-κB, and reducing the expression of ALDH, a marker of cancer stem cells.Citation171–Citation173 However, its clinical development has been restricted by its hydrophobicity, poor in vivo stability, and rapid metabolism. Gülçür et alCitation41 developed a novel nanomicellar formulation of curcumin to overcome these shortcomings. Encapsulating curcumin in sterically stabilized micelles (C-SSM) significantly enhanced its aqueous solubility and stability. Furthermore, curcumin-encapsulated C-SSM clearly enhanced the efficacy of curcumin against both breast cancer cells and BCSCs.
Wedelolactone, an active polyphenolic compound of Sphagneticola calendulacea and Eclipta prostrata,Citation174 was demonstrated to kill many cancer cells—including breast cancer cells—but its disadvantages, such as poor solubility and bioavailability, restrict its clinical application.Citation40,Citation175 Das et alCitation40 formulated wedelolactone-encapsulated poly(lactic-co-glycolic acid) (PLGA) nanoparticles (nWdl) to target BCSCs and overcome their shortcomings. nWdl was shown to sensitize BCSCs to the effects of wedelolactone by downregulating SOX2 and adenosine-triphosphate (ATP)-binding cassette super-family G member 2 (ABCG2).
SAL, a polyether ionophore antibiotic, has demonstrated great potential in eliminating BCSCs, but its clinical application is hindered by its poor aqueous solubility and severe systemic toxicity.Citation83,Citation139,Citation176,Citation177 Considering the need for an efficient drug while reducing potential damage to normal tissues, Zhao et alCitation139 developed biocompatible gold nanoparticles coated with poly(ethylene glycol) (PEG) that were conjugated with SAL. These SAL-loaded gold nanoparticles enhanced the therapeutic efficacy of SAL against BCSCs derived from a CD24low/CD44high subpopulation.
MET, an anti-type 2 diabetic drug, was reported to affect breast cancer at low dosages by targeting BCSCs; however, its anti-breast cancer efficacy is hindered by its low bioavailability and nonselective biodistribution. Lee et alCitation42 demonstrated that MET-encapsulated trastuzumab-conjugated immunoliposomes (Her-LP-MET) could target BCSCs and inhibit both their proliferation and migration. The combination of Her-LP-MET with free DOX resulted in better breast tumor remission than treatment with only free DOX.
It was suggested by Sun et alCitation13 that rationally designed drug delivery systems could significantly enhance the cancer stem cell therapy of conventional chemotherapeutic drugs such as DOX by delivering these drugs into cancer stem cells. They formulated DOX-tethered gold nanoparticles (DOX-Hyd@AuNPs) and demonstrated that DOX-Hyd@AuNPs could inhibit the growth of breast cancer without increasing the BCSC subpopulation in the tumor by delivering more DOX into the BCSCs. This process overcame the intrinsic resistance of BCSCs arising from P-glycoprotein (P-gp) drug efflux.
Delivery of Nucleic Acid Therapeutics to BCSCs
In addition to increasing the solubilization of low-solubility drugs, NDDSs have the capacity to enhance the stability and cellular uptake of macromolecules such as siRNA, shRNA, and miRNA which could potentially treat cancer.Citation130,Citation178 AKT2, a major downstream effector of the phosphatidylinositol 3-kinase (PI3K) pathway, was reported to be associated with cancer stem cell tumorigenicity and the malignant phenotype of cancer cells.Citation179–Citation181 The silencing of AKT2 with siRNA has the potential to inhibit the development and recurrence of tumors. Nevertheless, the rapid degradation and poor cellular uptake of siRNA are challenges for siRNA-based therapies. Using NDDSs to deliver siRNA may be a promising strategy to increase the stability and cellular delivery of siRNA. Rafael et alCitation152 developed an innovative nanocarrier system composed of Pluronic® F127-based micelles associated with polyethylenimine (PEI)-based polyplexes to deliver AKT2 siRNA. This AKT2-siRNA delivery system displayed strong suppressive effects on BCSCs migration and invasion in breast cancer.
NF-κB plays an important role in maintaining the properties of BCSCs in various types of breast cancer.Citation182,Citation183 Therefore, it is possible to target BCSCs by downregulating the expression of NF-κB proteins using RNA interference, including siRNA and shRNA. Compared to siRNA, shRNA is more stable; it is a double-stranded RNA molecule with a tight hairpin structure.Citation184 Ke et alCitation38 synthesized a carbamate-mannose-modified PEI (CMP) for the targeted delivery of NF-κB shRNA to BCSCs. These CMP/NF-κB-targeted shRNA nanocomplexes were demonstrated to reduce the percentage of BCSCs, inhibit the formation of mammospheres and colonies, suppress the migration and invasion of breast cancer cells, and sensitize breast cancer cells to treatment with doxorubicin-loaded micellar nanoparticles.
miRNAs are essential post-transcriptional regulators of many cellular functions. miR-34a is a tumor-suppressor miRNA that has been reported to have the capacity to attenuate the properties of BCSCs.Citation119 Lin et alCitation151 established a human telomerase reverse transcriptase (hTERT) promoter-driven VP16-Gal4-WPRE integrated systemic amplifier (VISA) delivery system for miR-34a (TV-miR-34a) plasmid. They demonstrated that TV-miR-34a clearly eliminated BCSCs both in vitro and in vivo in a safe and efficient way and showed increased therapeutic efficacy toward breast cancer cells in combination with docetaxel. Further mechanistic studies revealed that TV-miR-34a attenuated BCSC properties, promoted adherence, and boosted the differentiation of BCSCs by directly targeting chromosome 22 open reading frame 28 (C22ORF28).
Combinational Delivery of Chemotherapeutics and Cancer Stem Cell-Specific Agents
An increasing amount of evidence indicates that tumors are heterogeneous tissues with different types of cells, such as cancer stem cells and non-cancer stem cells.Citation137 Some therapeutic agents have been reported to eliminate cancer stem cells; however, cancer cells can also spontaneously transition to cancer stem cells; thus, the depletion of cancer stem cells alone is neither sufficient nor effective as a therapeutic measure.Citation3,Citation130,Citation185 Instead of targeting only cancer stem cells or non-cancer stem cells, combination strategies designed to simultaneously eradicate both cell types may have the potential to improve therapeutic outcomes. shows representative reports about combination strategies that target both cancer and non-cancer stem cells.
To eradicate both breast cancer cells and BCSCs, Kim et alCitation159 developed cross-linked multilamellar liposomal vesicles (cMLVs) to co-deliver DOX (a conventional chemotherapeutic drug) and SAL (an inhibitor of BCSCs), producing cMLV(DOX+SAL) particles. The antitumor results of cMLV(DOX+SAL) in vitro and in vivo demonstrated that the co-delivery of DOX and SAL in a single cMLV greatly inhibited both breast cancer cells and BCSCs, which may be attributed to the simultaneous delivery of the two drugs to the tumor tissue by cMLV(DOX+SAL).Citation159 Similarly, Zhang et alCitation160 evaluated the therapeutic efficacy of micelles that were co-loaded with the cytotoxic drug epirubicin (EPI) and the BCSC inhibitor staurosporine (STS) to treat breast cancers, especially when the tumors recurred after traditional chemotherapy. Theses results demonstrated that the STS/EPI-loaded micelles can potentially treat naïve orthotopic 4T1-luc breast tumors and their recurrent EPI-resistant counterparts by suppressing breast cancer cells together with the BCSC-associated subpopulation, such as the ALDH+ and CD44+/CD24− subpopulations.
NDDSs have co-delivered various traditional chemotherapeutic drugs with different antitumor mechanisms and exhibited the potential to eliminate both BCSCs and non-BCSCs. For instance, Sun et alCitation43 developed a cargo-free and pH-responsive nanomedicine for the co-delivery of a pH-responsive prodrug of DOX and 7-Ethyl-10-hydroxycamptothecin (SN38) to target breast cancer. The results showed that this nanomedicine significantly increased drug accumulation at the tumor site and simultaneously eradicated both BCSCs and non-BCSCs to achieve a superior antitumor efficacy in vivo. The excellent anti-BCSC capability of this developed nanomedicine may be attributed to the suppression of topoisomerase I (TOP I) and TOP II by DOX and SN38, respectively. Similarly, Wang et alCitation167 developed a nanoparticle loaded with DOX and irinotecan to inhibit both TOP I and TOP II, and the results showed that this co-delivery system noticeably enhanced the eradication of BCSCs with no evident systemic toxicity both in vitro and in vivo.
NDDSs can also be used for the co-delivery of traditional chemotherapeutic agents and nucleic acid agents such as siRNA, shRNA, and miRNA. For example, Chen et alCitation17 designed cationic-lipid-assisted nanoparticles to co-encapsulate docetaxel (DTXL, a traditional chemotherapeutic agent) and an siRNA targeting BMI-1 (siBMI-1, a nucleic acid agent) by the double emulsion method, producing DTXLNPsiBMI-1 nanoparticles. These nanoparticles could effectively deliver therapeutic agents to both bulk cancer cells and BCSCs to produce combinational effects in the treatment of breast cancer. The bulk cancer cells were killed by DTXL and the expression of BMI-1 in the BCSCs was downregulated by siBMI-1, thereby eliminating them by enhancing their chemosensitivity to DTXL by reducing stemness. In the MDA-MB-231 xenograft model, DTXLNPsiBMI-1 completely inhibited tumor growth and prevented recurrence, which was attributed to its capacity to kill both bulk cancer cells and BCSCs. Similarly, Samson et alCitation157 developed glucose-regulated protein 78 (GRP78)-targeted 1,2-dioleoyloxy-3-trimethylammoniumpropane (DOTAP) liposomes to deliver either camptothecin (CPT) and GRP78 siRNA (named DOTAP-CPT-siGRP78) or CPT and clusterin (CLU) siRNA (named DOTAP-CPT-siCLU). Both DOTAP-CPT-siGRP78 and DOTAP-CPT-siCLU exhibited stronger breast cancer cell- and BCSC-targeted activities than free CPT, confirming the synergistic effects of co-delivering anticancer drugs and siRNAs.
Additionally, NDDSs were explored to co-deliver protein agents targeting BCSCs and traditional chemotherapeutic agents to treat breast cancer. For instance, Xu et alCitation16 designed and prepared an aptamer-conjugated DNA nanotrain for the co-delivery of DOX and AKTin (an AKT inhibitor peptide); this drug delivery system was named TA6NT-AKTin-DOX. The efficacy of TA6NT-AKTin-DOX was evaluated on MCF-7 BCSCs and tumors generated by injecting BCSCs into nude mice. The results demonstrated that TA6NT-AKTin-DOX exhibited better efficacy than free DOX and various DNA nanotrains both in vitro and in vivo. The synergistic response of TA6NT-AKTin-DOX may be explained by AKTin, which can overcome the drug resistance of BCSCs via inhibition of the AKT signaling pathway.Citation16
Moreover, multiple therapeutic agents with various anticancer mechanisms can be co-encapsulated in a single NDDSs with a high capacity to synergistically kill tumors. For example, paclitaxel (PTX, a chemotherapeutic agent), thioridazine (THZ, an anti-BCSC agent), and HY1991 (HY, a programmed cell death 1 [PD-1]/programmed cell death ligand 1 [PD-L1] inhibitor) were incorporated into an enzyme/pH dual-sensitive nanoparticle with a micelle-liposome double-layer structure. This PTX/THZ/HY-co-loaded drug delivery system displayed excellent anti-breast cancer efficacy and prolonged the lifespan of tumor-bearing mice. This observation was attributed to the combination of chemotherapy, anti-BCSC therapy, and immune checkpoint blockade therapy.Citation163
Active-Targeting NDDSs
The BCSC-targeting capacity of NDDSs can be further increased by surface modification of the NDDSs with suitable ligands that can interact with overexpressed cell-surface proteins on the BCSCs, such as CD44, EpCAM, integrin α5 (ITGA5), extra domain B of fibronectin (EDB-FN), transferrin receptor 1 (TfR1), and scavenger receptor class A membrane 5 (SCARA5). This should achieve active-targeting drug delivery, by enhancing the accumulation of drugs in targeted cells and reducing off-target effects.
For instance, Al Faraj et alCitation44 demonstrated that modification with CD44 antibody significantly enhanced SAL and PTX-conjugated SWCNT nanocarrier accumulation in both breast cancer cells and BCSCs in a xenograft murine model, providing a potential method for the effective treatment of breast cancer by targeting and eliminating both tumor cell and BCSC populations. Hyaluronic acid and chitosan were also reported to be CD44 ligands and could be used for the active targeting of CD44-overexpressing cells. For example, Yang et alCitation169 fabricated a hyaluronic acid lipoid by attaching a lipoid to the surface of PLGA nanoparticles to construct a vehicle for co-delivering PTX and curcumin to BCSCs. Owing to the interaction between hyaluronic acid of the fabricated lipoid and the cell-surface CD44 receptors on the BCSCs, it was shown that the BCSC-targeting ability of the fabricated hyaluronic acid lipoid were significantly enhanced, which allowed the fabricated lipoid to suppress the proliferation and migration of BCSCs. Moreover, the fabricated hyaluronic acid lipoid displayed excellent anticancer effects against MCF-7 xenograft tumor models by simultaneously suppressing the growth of breast cancer cells and BCSCs. Similarly, Rao et alCitation145 formulated DOX-loaded polymeric nanoparticles decorated with chitosan on their surface to target the overexpressed CD44 receptors on tumor reinitiating cancer stem-like cells. The nanoparticles enhanced six times of cytotoxicities compared with free doxorubicin for the eradication of CD44+ cancer stem-like cells in 3D mammary tumor spheroids, and reduced tumor size with no obvious systemic toxicity in an orthotopic xenograft model by significantly increasing DOX accumulation in the tumors while reducing it in normal organs.
EpCAM is a surface marker on cancer stem cells that can be used to target them.Citation29,Citation30 For instance, Tran et alCitation45 comprehensively studied the antitumor effects of nanoamorphous aspirin-loaded exosomes and showed that they had an unprecedented cancer stem cell eradication capacity. These authors also modified the exosomes with an aptamer specifically targeting EpCAM, which they found could further enhance the active-targeting ability of the exosomes.
The canonical Wnt/β-catenin pathway plays essential roles in the generation and maintenance of both cancer and normal stem cells. Thus, cancer therapy using inhibitors of the Wnt/β-catenin pathway may be toxic to normal stem cells. To achieve the specific inhibition of β-catenin in cancer cells, Li et alCitation46 proposed a strategy to suppress the stemness and metastasis of TNBC by developing an ITGA5-targeting lipid–polymer hybrid (LPH) nanoparticle modified with an ITGA5 ligand (a commercialized RGD motif, Arg-Gly-Asp) for the TNBC-targeted delivery of diacidic norcantharidin (NCTD). The developed RGD-decorated LPH nanoparticles significantly enhanced the accumulation of the delivered drug in orthotopic mammary TNBC tumors and lung metastatic tumors in nude mice. The nanoparticles also reduced the growth and metastasis of TNBC compared to that of free NCTD and non-modified LPH nanoparticles by downregulating β-catenin.
EDB-FN plays essential roles in the maintenance and growth of BCSCs, as well as in the expression of genes encoding surface markers on BCSCs and controlling their self-renewal.Citation154 Therefore, EDB-FN may be used as a biomarker for both targeting and treating BCSCs. APTEDB, an EDB-FN-specific peptide, was used as a cancer-targeting ligand by Sun et alCitation154 to modify liposomes encapsulating EDB-FN siRNAs, forming a liposomal system (APTEDB-LS-siRNAEDB) with the potential to simultaneously target and knockdown EDB-FN in breast cancer treatment. It was found that APTEDB-LS-siRNAEDB significantly increased the accumulation and cellular uptake of the delivered EDB-FN siRNA in EDB-FN-positive BCSCs in cultured cells and tissues compared to those in non-targeted liposomes. Moreover, APTEDB-LS-siRNAEDB could knockdown EDB-FN both in vitro and in vivo, effectively treating EDB-FN-positive BCSC-derived tumors by eradicating the self-renewal ability of BCSCs.
Because of specific binding to the highly expressed TfR1 and SCARA5, ferritin is a promising nanoplatform for the efficient targeting and deep penetration of tumors.Citation47 Recently, Tan et alCitation47 formulated ferritin nanocages loaded with 1.1-dioctadecyl-3,3,3,3-tetramethylindotricarbocyanine iodide (DBN) and EPI for the targeted treatment of breast cancer. Both DBN and EPI collected in large numbers at the tumor site, permeated throughout the tumor mass, and accessed the BCSCs in a metastatic 4T1-induced tumor model, which was attributed to the tumor-homing and biomimetic properties of the ferritin nanocages. It was noted that the DBN and EPI-loaded, BCSC-accessing nanocages considerably suppressed primary tumor growth with the notable elimination of BCSCs in the tumor mass, and significantly suppressed lung metastasis under the combined effect of photothermal and chemo therapies.
In addition to BCSC-specific surface proteins, some membrane proteins that are overexpressed on both BCSCs and non-BCSCs could be utilized to enhance the targeting ability of NDDSs. The identification of surface receptors overexpressed on both cancer and non-cancer stem cells is important for successful cancer treatment because the latter can transform into cancer stem cells via plastic EMT. For example, Fonseca et alCitation150 demonstrated that nucleolin receptors are overexpressed in both the BCSCs and non-BCSCs found in TNBC. These authors developed DOX and C6-ceramide co-loaded liposomes coated with nucleolin-binding F3 peptides to reduce the plasticity and adaptability associated with BCSCs and non-BCSCs. The developed F3 peptide-targeted liposomes exhibited significantly increased cellular toxicity against both BCSCs and non-BCSCs compared to that of the liposomes without F3 peptides.Citation150 Similarly, glucose transporter (GLUT) is overexpressed in many cancer cells and cancer stem cells owing to the Warburg effect.Citation146,Citation186,Citation187 Thus, GLUT is a potential target for cancer treatment. Recently, Yi et alCitation155 created a glucose-installed nanocarrier by conjugating unimer polyion complexes on Au nanoparticles for the targeted delivery of polo-like kinase 1 (PLK1) siRNA to BCSCs via the interaction of glucose ligands with the glucose transporter 1 (GLUT1) overexpressed on their surface. Glucose nanoparticles loaded with PLK1 siRNA have the capacity to efficiently reduce the percentage of BCSCs in cancer stem cell-rich orthotopic MDA-MB-231 tumor tissue by enhancing gene silencing, indicating that modification with glucose could significantly enhance the BCSC-targeted delivery of nanocarriers.
Surface ligand density also plays an essential role in cancer cell-specific delivery. Thus, dual-targeted NDDSs appear to be an efficient strategy for enhancing the BCSC-targeting capacity of NDDSs. For example, Qiao et alCitation147 developed an NDDS for the highly selective targeting of BCSCs by binding hyaluronic acid and doublecortin-like kinase 1 (DCLK1) monoclonal antibody on the surface of PEG–PLGA nanoparticles (PEG–PLGA NPs). The formed NPs were named DCLK1–HA–PEG–PLGA NPs, and they targeted BCSCs both in vitro and in vivo by the specific interaction of the DCLK1 monoclonal antibodies and HA molecules with the DCLK1 protein and CD44 receptors overexpressed on the surface of BCSCs, respectively. Similarly, nucleus-targeted drug delivery systems hold great potential to reverse cancer stem cell-mediated drug resistance. Because of this, Li et alCitation148 designed and synthesized a nanosystem with a core/shell structure of mesoporous silica nanoparticles loaded with the anticancer drug tirapazamine (TPZ). The surface of the silica was also modified with anti-CD133 antibody and TAT peptide. This nanosystem possessed three stages of drug delivery: 1) target BCSCs with the anti-CD133 antibody; 2) target the nucleus with the TAT peptide; and 3) release the TPZ in the nucleus to eliminate hypoxic BCSCs. It was revealed that the synthesized nanosystem significantly inhibited BCSC survival in vitro and suppressed tumor growth in a breast tumor xenograft model without obvious side effects. Mechanistically, this nanosystem was found to attenuate the hypoxia signaling pathway by suppressing the expression of HIF-1α.Citation148
Advantages and Limitations of Current NDDSs Against BCSCs
The overview of the current state suggests that NDDSs are an effective treatment solution that can overcome the disadvantages of conventional therapeutic agents against BCSCs and speed up the development of potential drugs against breast cancer via anti-BCSCs. Compared to the conventional agents, NDDSs have some potential advantages against BCSCs. For example, NDDSs can overcome many of the undesirable properties of conventional agents against BCSCs.Citation3,Citation130,Citation137 Moreover, although normal stem cells and BCSCs may have similar properties, such as self-renewal,Citation80 NDDSs could reduce, at least to some extent, the toxicity on normal stem cells by selectively accumulating in tumor tissue with the help of the EPR effect to minimize any impact on normal stem cells.Citation131 In addition, the targeting capacity of NDDSs to BCSCs could be further enhanced by the modification of NDDSs with BCSCs surface marker-specific ligands/antibodies, resulting in a further increase in the anti-BCSC ability of NDDSs while reducing toxicity on normal tissue.Citation131 Furthermore, NDDSs could encapsulate agents used against the large numbers of breast cancer cells, agents against the less abundant BCSCs, and/or agents targeting the breast tumor microenvironment into the same nanoparticle, enabling these drugs to target the tumor tissue as a single drug. This can solve the potential problems arising from these drugs due to the fact that they possess different biopharmaceutical parameters in vivo, which prevents them from producing the desired synergistic effect.Citation163 Nevertheless, the research and development of NDDSs against BCSCs is in its infancy, and many problems need to be overcome. Further research on the biological characteristics of BCSCs and the design of more efficient NDDSs is needed to overcome the limitations identified during the practical application of NDDSs. First, to eliminate BCSCs within breast cancer tissue, the targeted NDDSs need to penetrate the sites where BCSCs are located. Indeed, some subpopulations of BCSCs are located in poorly vascularized regions, which are extremely difficult for NDDSs to reach.Citation3 Another limitation of NDDSs is that although many solutions have been proposed to reduce the reticuloendothelial system (RES) uptake of NDDSs, the retention of NDDSs in bypassing organs and their cellular uptake by RES macrophages is still a significant problem.Citation163
Conclusion
Through their involvement in the relapse, metastasis, and therapeutic resistance of breast cancer, BCSCs can make its treatment challenging. However, with a greater understanding of the biological properties of BCSCs, an increasing number of strategies, such as targeting surface markers, specific signaling pathways, metabolism, and the microenvironment of BCSCs, have been proposed to treat breast cancer by eradicating BCSCs. In this review, we summarized the current development of anti-BCSC strategies to treat breast cancer using conventional agents and NDDSs against BCSCs. Although many agents, including small-molecule inhibitors/drugs, proteins, and nucleic acids, have the potential to eliminate BCSCs, their clinical translation is limited because of their poor solubility, instability, unfavorable biodistribution, and high toxicity induced by off-target effects. This overview of the recent utilization of NDDSs to target BCSCs suggests that NDDSs have the capacity to address many shortcomings of current anti-BCSC agents and exhibit various advantages to treat breast cancer by eliminating BCSCs. However, the application of NDDSs for BCSC targeting is in its infancy, and many issues require further elucidation to develop more efficient NDDSs for targeting BCSCs with low systemic toxicities.
Acknowledgments
This work was supported by Fundamental Research Funds for the Central Universities (Grant No. 18ykpy19); Science and Technology Program of Guangzhou, China (Grant No. 201904010270); Yixian Scientific Research Project Set Sail (Grant No. YXQH201706).
Disclosure
The authors report no conflicts of interest for this work.
References
- Tao Z, Shi A, Lu C, Song T, Zhang Z, Zhao J. Breast cancer: epidemiology and etiology. Cell Biochem Biophys. 2015;72(2):333–338. doi:10.1007/s12013-014-0459-6
- Sousa B, Ribeiro AS, Paredes J. Heterogeneity and plasticity of breast cancer stem cells. Adv Exp Med Biol. 2019;1139:83–103.
- Zhao Y, Alakhova DY, Kabanov AV. Can nanomedicines kill cancer stem cells? Adv Drug Deliv Rev. 2013;65(13–14):1763–1783.
- Kai M, Kanaya N, Wu SV, et al. Targeting breast cancer stem cells in triple-negative breast cancer using a combination of LBH589 and salinomycin. Breast Cancer Res Treat. 2015;151(2):281–294. doi:10.1007/s10549-015-3376-5
- Tian J, Raffa FA, Dai M, et al. Dasatinib sensitises triple negative breast cancer cells to chemotherapy by targeting breast cancer stem cells. Br J Cancer. 2018;119(12):1495–1507. doi:10.1038/s41416-018-0287-3
- Deng L, Shang L, Bai S, et al. MicroRNA100 inhibits self-renewal of breast cancer stem-like cells and breast tumor development. Cancer Res. 2014;74(22):6648–6660. doi:10.1158/0008-5472.CAN-13-3710
- Jang GB, Hong IS, Kim RJ, et al. Wnt/β-catenin small-molecule inhibitor CWP232228 preferentially inhibits the growth of breast cancer stem-like cells. Cancer Res. 2015;75(8):1691–1702. doi:10.1158/0008-5472.CAN-14-2041
- Yang F, Xu J, Tang L, Guan X. Breast cancer stem cell: the roles and therapeutic implications. Cell Mol Life Sci. 2017;74(6):951–966. doi:10.1007/s00018-016-2334-7
- Zhang P, Liu L, Zhang L, et al. Runx2 is required for activity of CD44(+)/CD24(-/low) breast cancer stem cell in breast cancer development. Am J Transl Res. 2020;12(5):2305–2318.
- Joseph C, Arshad M, Kurozomi S, et al. Overexpression of the cancer stem cell marker CD133 confers a poor prognosis in invasive breast cancer. Breast Cancer Res Treat. 2019;174(2):387–399. doi:10.1007/s10549-018-05085-9
- Louhichi T, Ziadi S, Saad H, Dhiab MB, Mestiri S, Trimeche M. Clinicopathological significance of cancer stem cell markers CD44 and ALDH1 expression in breast cancer. Breast Cancer. 2018;25(6):698–705. doi:10.1007/s12282-018-0875-3
- Nilendu P, Kumar A, Kumar A, Pal JK, Sharma NK. Breast cancer stem cells as last soldiers eluding therapeutic burn: a hard nut to crack. Int J Cancer. 2018;142(1):7–17. doi:10.1002/ijc.30898
- Sun TM, Wang YC, Wang F, et al. Cancer stem cell therapy using doxorubicin conjugated to gold nanoparticles via hydrazone bonds. Biomaterials. 2014;35(2):836–845. doi:10.1016/j.biomaterials.2013.10.011
- Nayak D, Tripathi N, Kathuria D, et al. Quinacrine and curcumin synergistically increased the breast cancer stem cells death by inhibiting ABCG2 and modulating DNA damage repair pathway. Int J Biochem Cell Biol. 2020;119:105682. doi:10.1016/j.biocel.2019.105682
- Yuan JD, ZhuGe DL, Tong MQ, et al. pH-sensitive polymeric nanoparticles of mPEG-PLGA-PGlu with hybrid core for simultaneous encapsulation of curcumin and doxorubicin to kill the heterogeneous tumour cells in breast cancer. Artif Cells, Nanomed Biotechnol. 2018;46(sup1):1–12. doi:10.1080/21691401.2017.1423495
- Xu Z, Ni R, Chen Y. Targeting breast cancer stem cells by a self-assembled, aptamer-conjugated DNA nanotrain with preloading doxorubicin. Int J Nanomedicine. 2019;14:6831–6842. doi:10.2147/IJN.S200482
- Chen K, Shen S, Zhao G, Cao Z, Yang X, Wang J. Simultaneous elimination of cancer stem cells and bulk cancer cells by cationic-lipid-assisted nanoparticles for cancer therapy. Nano Res. 2018;11(8):4183–4198. doi:10.1007/s12274-018-2007-y
- Chen JH, Huang WC, Bamodu OA, Chang PMH, Chao TY, Huang TH. Monospecific antibody targeting of CDH11 inhibits epithelial-to-mesenchymal transition and represses cancer stem cell-like phenotype by up-regulating miR-335 in metastatic breast cancer, in vitro and in vivo. BMC Cancer. 2019;19(1):634. doi:10.1186/s12885-019-5811-1
- Xie T, Jiang C, Dai T, et al. Knockdown of XB130 restrains cancer stem cell-like phenotype through inhibition of Wnt/β-Catenin signaling in breast cancer. Mol Carcinog. 2019;58(10):1832–1845. doi:10.1002/mc.23071
- Kurebayashi J, Koike Y, Ohta Y, et al. Anti-cancer stem cell activity of a hedgehog inhibitor GANT61 in estrogen receptor-positive breast cancer cells. Cancer Sci. 2017;108(5):918–930. doi:10.1111/cas.13205
- Kim DH, Yoon HJ, Cha YN, Surh YJ. Role of heme oxygenase-1 and its reaction product, carbon monoxide, in manifestation of breast cancer stem cell-like properties: notch-1 as a putative target. Free Radic Res. 2018;52(11–12):1336–1347. doi:10.1080/10715762.2018.1473571
- Cordenonsi M, Zanconato F, Azzolin L, et al. The Hippo transducer TAZ confers cancer stem cell-related traits on breast cancer cells. Cell. 2011;147(4):759–772. doi:10.1016/j.cell.2011.09.048
- Woosley AN, Dalton AC, Hussey GS, et al. TGFβ promotes breast cancer stem cell self-renewal through an ILEI/LIFR signaling axis. Oncogene. 2019;38(20):3794–3811. doi:10.1038/s41388-019-0703-z
- Elia I, Rossi M, Stegen S, et al. Breast cancer cells rely on environmental pyruvate to shape the metastatic niche. Nature. 2019;568(7750):117–121. doi:10.1038/s41586-019-0977-x
- Tsuyada A, Chow A, Wu J, et al. CCL2 mediates cross-talk between cancer cells and stromal fibroblasts that regulates breast cancer stem cells. Cancer Res. 2012;72(11):2768–2779. doi:10.1158/0008-5472.CAN-11-3567
- Korkaya H, Liu S, Wicha MS. Breast cancer stem cells, cytokine networks, and the tumor microenvironment. J Clin Invest. 2011;121(10):3804–3809. doi:10.1172/JCI57099
- Ryoo IG, Choi BH, Ku SK, Kwak MK. High CD44 expression mediates p62-associated NFE2L2/NRF2 activation in breast cancer stem cell-like cells: implications for cancer stem cell resistance. Redox Biol. 2018;17:246–258. doi:10.1016/j.redox.2018.04.015
- Brugnoli F, Grassilli S, Al-Qassab Y, Capitani S, Bertagnolo V. CD133 in breast cancer cells: more than a stem cell marker. J Oncol. 2019;2019:7512632. doi:10.1155/2019/7512632
- Naujokat C. Monoclonal antibodies against human cancer stem cells. Immunotherapy. 2014;6(3):290–308. doi:10.2217/imt.14.4
- Kubo M, Umebayashi M, Kurata K, et al. Catumaxomab with activated T-cells efficiently lyses chemoresistant EpCAM-positive triple-negative breast cancer cell lines. Anticancer Res. 2018;38(7):4273–4279. doi:10.21873/anticanres.12724
- Sun R, Liu Y, Li SY, et al. Co-delivery of all-trans-retinoic acid and doxorubicin for cancer therapy with synergistic inhibition of cancer stem cells. Biomaterials. 2015;37:405–414. doi:10.1016/j.biomaterials.2014.10.018
- Ginestier C, Monville F, Wicinski J, et al. Mevalonate metabolism regulates basal breast cancer stem cells and is a potential therapeutic target. Stem Cells. 2012;30(7):1327–1337. doi:10.1002/stem.1122
- Patra Krushna C, Wang Q, Bhaskar Prashanth T, et al. Hexokinase 2 is required for tumor initiation and maintenance and its systemic deletion is therapeutic in mouse models of cancer. Cancer Cell. 2013;24(2):213–228. doi:10.1016/j.ccr.2013.06.014
- Wang R, Yang L, Li S, et al. Quercetin inhibits breast cancer stem cells via downregulation of aldehyde dehydrogenase 1A1 (ALDH1A1), chemokine receptor type 4 (CXCR4), mucin 1 (MUC1), and epithelial cell adhesion molecule (EpCAM). Med Sci Monit. 2018;24:412–420. doi:10.12659/MSM.908022
- Li Y, Zhang T, Korkaya H, et al. Sulforaphane, a dietary component of broccoli/broccoli sprouts, inhibits breast cancer stem cells. Clin Cancer Res. 2010;16(9):2580–2590. doi:10.1158/1078-0432.CCR-09-2937
- Li X, Wang X, Xie C, et al. Sonic hedgehog and Wnt/β-catenin pathways mediate curcumin inhibition of breast cancer stem cells. Anticancer Drugs. 2018;29(3):208–215. doi:10.1097/CAD.0000000000000584
- Gupta PB, Onder TT, Jiang G, et al. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell. 2009;138(4):645–659. doi:10.1016/j.cell.2009.06.034
- Ke X, Yang C, Cheng W, Yang YY. Delivery of NF-κB shRNA using carbamate-mannose modified PEI for eliminating cancer stem cells. Nanomedicine: NBM. 2018;14(2):405–414. doi:10.1016/j.nano.2017.11.015
- Hu T, Liu S, Breiter DR, Wang F, Tang Y, Sun S. Octamer 4 small interfering RNA results in cancer stem cell–like cell apoptosis. Cancer Res. 2008;68(16):6533–6540. doi:10.1158/0008-5472.CAN-07-6642
- Das S, Mukherjee P, Chatterjee R, Jamal Z, Chatterji U. Enhancing chemosensitivity of breast cancer stem cells by downregulating SOX2 and ABCG2 using wedelolactone-encapsulated nanoparticles. Mol Cancer Ther. 2019;18(3):680–692. doi:10.1158/1535-7163.MCT-18-0409
- Gülçür E, Thaqi M, Khaja F, Kuzmis A, Önyüksel H. Curcumin in VIP-targeted sterically stabilized phospholipid nanomicelles: a novel therapeutic approach for breast cancer and breast cancer stem cells. Drug Deliv Transl Res. 2013;3(6):562–574. doi:10.1007/s13346-013-0167-6
- Lee JY, Shin DH, Kim JS. Anticancer effect of metformin in herceptin-conjugated liposome for breast cancer. Pharmaceutics. 2019;12(1):11. doi:10.3390/pharmaceutics12010011
- Sun N, Zhao CY, Cheng R, et al. Cargo-free nanomedicine with pH sensitivity for codelivery of DOX conjugated prodrug with SN38 to synergistically eradicate breast cancer stem cells. Mol Pharm. 2018;15(8):3343–3355. doi:10.1021/acs.molpharmaceut.8b00367
- Al Faraj A, Shaik AS, Ratemi E, Halwani R. Combination of drug-conjugated SWCNT nanocarriers for efficient therapy of cancer stem cells in a breast cancer animal model. J Control Release. 2016;225:240–251. doi:10.1016/j.jconrel.2016.01.053
- Tran PHL, Wang T, Yin W, et al. Aspirin-loaded nanoexosomes as cancer therapeutics. Int J Pharm. 2019;572:118786. doi:10.1016/j.ijpharm.2019.118786
- Li Y, Xiao Y, Lin HP, et al. In vivo beta-catenin attenuation by the integrin alpha5-targeting nano-delivery strategy suppresses triple negative breast cancer stemness and metastasis. Biomaterials. 2019;188:160–172. doi:10.1016/j.biomaterials.2018.10.019
- Tan T, Wang H, Cao H, et al. Deep tumor-penetrated nanocages improve accessibility to cancer stem cells for photothermal-chemotherapy of breast cancer metastasis. Adv Sci. 2018;5(12):1801012. doi:10.1002/advs.201801012
- Dittmer J. Breast cancer stem cells: features, key drivers and treatment options. Semin Cancer Biol. 2018;53:59–74.
- Li X, Meng Y, Xie C, et al. Diallyl Trisulfide inhibits breast cancer stem cells via suppression of Wnt/β-catenin pathway. J Cell Biochem. 2018;119(5):4134–4141. doi:10.1002/jcb.26613
- Yan Y, Zuo X, Wei D. Concise review: emerging role of CD44 in cancer stem cells: a promising biomarker and therapeutic target. Stem Cells Transl Med. 2015;4(9):1033–1043. doi:10.5966/sctm.2015-0048
- Pham PV, Phan NL, Nguyen NT, et al. Differentiation of breast cancer stem cells by knockdown of CD44: promising differentiation therapy. J Transl Med. 2011;9:209. doi:10.1186/1479-5876-9-209
- Snyder V, Reed-Newman TC, Arnold L, Thomas SM, Anant S. Cancer stem cell metabolism and potential therapeutic targets. Front Oncol. 2018;8. doi:10.3389/fonc.2018.00203
- Hirsch HA, Iliopoulos D, Struhl K. Metformin inhibits the inflammatory response associated with cellular transformation and cancer stem cell growth. Proc Natl Acad Sci. 2013;110(3):972–977. doi:10.1073/pnas.1221055110
- El Hout M, Dos Santos L, Hamaï A, Mehrpour M. A promising new approach to cancer therapy: targeting iron metabolism in cancer stem cells. Semin Cancer Biol. 2018;53:125–138. doi:10.1016/j.semcancer.2018.07.009
- Borovski T, De sousa EMF, Vermeulen L, Medema JP. Cancer stem cell niche: the place to be. Cancer Res. 2011;71(3):634–639. doi:10.1158/0008-5472.CAN-10-3220
- Plaks V, Kong N, Werb Z. The cancer stem cell niche: how essential is the niche in regulating stemness of tumor cells? Cell Stem Cell. 2015;16(3):225–238. doi:10.1016/j.stem.2015.02.015
- Marangoni E, Lecomte N, Durand L, et al. CD44 targeting reduces tumour growth and prevents post-chemotherapy relapse of human breast cancers xenografts. Br J Cancer. 2009;100(6):918–922. doi:10.1038/sj.bjc.6604953
- Grosse-Gehling P, Fargeas CA, Dittfeld C, et al. CD133 as a biomarker for putative cancer stem cells in solid tumours: limitations, problems and challenges. J Pathol. 2013;229(3):355–378.
- Ohlfest JR, Zellmer DM, Panyam J, et al. Immunotoxin targeting CD133(+) breast carcinoma cells. Drug Deliv Transl Res. 2013;3(2):195–204. doi:10.1007/s13346-012-0066-2
- Das PK, Rakib MA, Khanam JA, Pillai S, Islam F. Novel therapeutics against breast cancer stem cells by targeting surface markers and signaling pathways. Curr Stem Cell Res Ther. 2019;14(8):663–676. doi:10.2174/1574888X14666190628104721
- Nalla LV, Kalia K, Khairnar A. Self-renewal signaling pathways in breast cancer stem cells. Int J Biochem Cell Biol. 2019;107:140–153. doi:10.1016/j.biocel.2018.12.017
- Hong IS, Jang GB, Lee HY, Nam JS. Targeting cancer stem cells by using the nanoparticles. Int J Nanomedicine. 2015;10(Spec Iss):251–260. doi:10.2147/IJN.S88310
- Zhou J, Chen Q, Zou Y, Chen H, Qi L, Chen Y. Stem cells and cellular origins of breast cancer: updates in the rationale, controversies, and therapeutic implications. Front Oncol. 2019;9:820. doi:10.3389/fonc.2019.00820
- Gurney A, Axelrod F, Bond CJ, et al. Wnt pathway inhibition via the targeting of Frizzled receptors results in decreased growth and tumorigenicity of human tumors. Proc Natl Acad Sci U S A. 2012;109(29):11717–11722. doi:10.1073/pnas.1120068109
- Venkatesh V, Nataraj R, Thangaraj GS, et al. Targeting Notch signalling pathway of cancer stem cells. Stem Cell Investig. 2018;5:5. doi:10.21037/sci.2018.02.02
- Bray SJ. Notch signalling: a simple pathway becomes complex. Nat Rev Mol Cell Biol. 2006;7(9):678–689. doi:10.1038/nrm2009
- Grudzien P, Lo S, Albain KS, et al. Inhibition of Notch signaling reduces the stem-like population of breast cancer cells and prevents mammosphere formation. Anticancer Res. 2010;30(10):3853–3867.
- McClements L, Annett S, Yakkundi A, et al. FKBPL and its peptide derivatives inhibit endocrine therapy resistant cancer stem cells and breast cancer metastasis by downregulating DLL4 and Notch4. BMC Cancer. 2019;19(1):351. doi:10.1186/s12885-019-5500-0
- Butti R, Gunasekaran VP, Kumar TVS, Banerjee P, Kundu GC. Breast cancer stem cells: biology and therapeutic implications. Int J Biochem Cell Biol. 2019;107:38–52. doi:10.1016/j.biocel.2018.12.001
- Pindiprolu S, Krishnamurthy PT, Chintamaneni PK, Karri V. Nanocarrier based approaches for targeting breast cancer stem cells. Artif Cells Nanomed Biotechnol. 2018;46(5):885–898. doi:10.1080/21691401.2017.1366337
- Wang L, Duan W, Kang L, et al. Smoothened activates breast cancer stem-like cell and promotes tumorigenesis and metastasis of breast cancer. Biomed Pharmacother. 2014;68(8):1099–1104. doi:10.1016/j.biopha.2014.09.012
- Koike Y, Ohta Y, Saitoh W, et al. Anti-cell growth and anti-cancer stem cell activities of the non-canonical hedgehog inhibitor GANT61 in triple-negative breast cancer cells. Breast Cancer. 2017;24(5):683–693. doi:10.1007/s12282-017-0757-0
- Kubo M, Nakamura M, Tasaki A, et al. Hedgehog signaling pathway is a new therapeutic target for patients with breast cancer. Cancer Res. 2004;64(17):6071–6074. doi:10.1158/0008-5472.CAN-04-0416
- Ramos A, Camargo FD. The Hippo signaling pathway and stem cell biology. Trends Cell Biol. 2012;22(7):339–346. doi:10.1016/j.tcb.2012.04.006
- Stanger BZ. Quit your YAPing: a new target for cancer therapy. Genes Dev. 2012;26(12):1263–1267. doi:10.1101/gad.196501.112
- Li Y-W, Xu J, Zhu G-Y, et al. Apigenin suppresses the stem cell-like properties of triple-negative breast cancer cells by inhibiting YAP/TAZ activity. Cell Death Discovery. 2018;4:105. doi:10.1038/s41420-018-0124-8
- Morikawa M, Derynck R, Miyazono KTGF-Β. and the TGF-β family: context-dependent roles in cell and tissue physiology. Cold Spring Harb Perspect Biol. 2016;8(5):a021873. doi:10.1101/cshperspect.a021873
- Xu X, Zhang L, He X, et al. TGF-β plays a vital role in triple-negative breast cancer (TNBC) drug-resistance through regulating stemness, EMT and apoptosis. Biochem Biophys Res Commun. 2018;502(1):160–165. doi:10.1016/j.bbrc.2018.05.139
- Ghasemi F, Sarabi PZ, Athari SS, Esmaeilzadeh A. Therapeutics strategies against cancer stem cell in breast cancer. Int J Biochem Cell Biol. 2019;109:76–81. doi:10.1016/j.biocel.2019.01.015
- Najafi M, Farhood B, Mortezaee K. Cancer stem cells (CSCs) in cancer progression and therapy. J Cell Physiol. 2019;234(6):8381–8395. doi:10.1002/jcp.27740
- Liu Y, Chen C, Qian P, et al. Gd-metallofullerenol nanomaterial as non-toxic breast cancer stem cell-specific inhibitor. Nat Commun. 2015;6:5988. doi:10.1038/ncomms6988
- Warrell RP, de Thé H, Wang ZY, Degos L. Acute promyelocytic leukemia. N Engl J Med. 1993;329(3):177–189. doi:10.1056/NEJM199307153290307
- Jangamreddy JR, Ghavami S, Grabarek J, et al. Salinomycin induces activation of autophagy, mitophagy and affects mitochondrial polarity: differences between primary and cancer cells. Biochimica Et Biophysica Acta. 2013;1833(9):2057–2069. doi:10.1016/j.bbamcr.2013.04.011
- Bartoschek M, Oskolkov N, Bocci M, et al. Spatially and functionally distinct subclasses of breast cancer-associated fibroblasts revealed by single cell RNA sequencing. Nat Commun. 2018;9(1):5150. doi:10.1038/s41467-018-07582-3
- Liu S, Ginestier C, Ou SJ, et al. Breast cancer stem cells are regulated by mesenchymal stem cells through cytokine networks. Cancer Res. 2011;71(2):614–624. doi:10.1158/0008-5472.CAN-10-0538
- Yang J, Liao D, Chen C, et al. Tumor-associated macrophages regulate murine breast cancer stem cells through a novel paracrine EGFR/Stat3/Sox-2 signaling pathway. STEM CELLS. 2013;31(2):248–258. doi:10.1002/stem.1281
- Goto H, Shimono Y, Funakoshi Y, et al. Adipose-derived stem cells enhance human breast cancer growth and cancer stem cell-like properties through adipsin. Oncogene. 2019;38(6):767–779. doi:10.1038/s41388-018-0477-8
- Vásquez-Bochm LX, Velázquez-Paniagua M, Castro-Vázquez SS, et al. Transcriptome-based identification of lovastatin as a breast cancer stem cell-targeting drug. Pharmacol Rep. 2019;71(3):535–544. doi:10.1016/j.pharep.2019.02.011
- Li X, Zhou N, Wang J, et al. Quercetin suppresses breast cancer stem cells (CD44+/CD24−) by inhibiting the PI3K/Akt/mTOR-signaling pathway. Life Sci. 2018;196:56–62. doi:10.1016/j.lfs.2018.01.014
- Bak MJ, Furmanski P, Shan NL, et al. Tocopherols inhibit estrogen-induced cancer stemness and OCT4 signaling in breast cancer. Carcinogenesis. 2018;39(8):1045–1055. doi:10.1093/carcin/bgy071
- Aztopal N, Erkisa M, Erturk E, Ulukaya E, Tokullugil AH, Ari F. Valproic acid, a histone deacetylase inhibitor, induces apoptosis in breast cancer stem cells. Chem Biol Interact. 2018;280:51–58. doi:10.1016/j.cbi.2017.12.003
- Das T, Nair RR, Green R, et al. Actinomycin D down-regulates SOX2 expression and induces death in breast cancer stem cells. Anticancer Res. 2017;37(4):1655–1663.
- Pattabiraman DR, Bierie B, Kober KI, et al. Activation of PKA leads to mesenchymal-to-epithelial transition and loss of tumor-initiating ability. Science. 2016;351(6277):aad3680. doi:10.1126/science.aad3680
- Manupati K, Debnath S, Goswami K, et al. Glutathione S-transferase omega 1 inhibition activates JNK-mediated apoptotic response in breast cancer stem cells. FEBS J. 2019;286(11):2167–2192. doi:10.1111/febs.14813
- Pan JX, Ding K, Wang CY. Niclosamide, an old antihelminthic agent, demonstrates antitumor activity by blocking multiple signaling pathways of cancer stem cells. Chin J Cancer. 2012;31(4):178–184. doi:10.5732/cjc.011.10290
- You J, Zhao J, Wen X, et al. Chemoradiation therapy using cyclopamine-loaded liquid-lipid nanoparticles and lutetium-177-labeled core-crosslinked polymeric micelles. J Control Release. 2015;202:40–48. doi:10.1016/j.jconrel.2015.01.031
- Cuyàs E, Gumuzio J, Verdura S, et al. The LSD1 inhibitor iadademstat (ORY-1001) targets SOX2-driven breast cancer stem cells: a potential epigenetic therapy in luminal-B and HER2-positive breast cancer subtypes. Aging. 2020;12(6):4794–4814. doi:10.18632/aging.102887
- Subramanian C, Grogan PT, Wang T, et al. Novel C-terminal heat shock protein 90 inhibitors target breast cancer stem cells and block migration, self-renewal, and epithelial–mesenchymal transition. Mol Oncol. 2020;14(9):2058–2068. doi:10.1002/1878-0261.12686
- Yu F, Zhao J, Hu Y, et al. The combination of NVP-BKM120 with trastuzumab or RAD001 synergistically inhibits the growth of breast cancer stem cells in vivo. Oncol Rep. 2016;36(1):356–364. doi:10.3892/or.2016.4799
- Xu L, Zhang L, Hu C, et al. WNT pathway inhibitor pyrvinium pamoate inhibits the self-renewal and metastasis of breast cancer stem cells. Int J Oncol. 2016;48(3):1175–1186. doi:10.3892/ijo.2016.3337
- Liu R, Shi P, Nie Z, et al. Mifepristone suppresses basal triple-negative breast cancer stem cells by down-regulating KLF5 expression. Theranostics. 2016;6(4):533–544. doi:10.7150/thno.14315
- Choi DS, Blanco E, Kim YS, et al. Chloroquine eliminates cancer stem cells through deregulation of Jak2 and DNMT1. Stem Cells. 2014;32(9):2309–2323. doi:10.1002/stem.1746
- Jung YC, Han S, Hua L, et al. Kazinol-E is a specific inhibitor of ERK that suppresses the enrichment of a breast cancer stem-like cell population. Biochem Biophys Res Commun. 2016;470(2):294–299. doi:10.1016/j.bbrc.2016.01.066
- Hirsch HA, Iliopoulos D, Tsichlis PN, Struhl K. Metformin selectively targets cancer stem cells, and acts together with chemotherapy to block tumor growth and prolong remission. Cancer Res. 2009;69(19):7507–7511. doi:10.1158/0008-5472.CAN-09-2994
- Sachlos E, Risueño Ruth M, Laronde S, et al. Identification of drugs including a dopamine receptor antagonist that selectively target cancer stem cells. Cell. 2012;149(6):1284–1297. doi:10.1016/j.cell.2012.03.049
- Shan NL, Wahler J, Lee HJ, et al. Vitamin D compounds inhibit cancer stem-like cells and induce differentiation in triple negative breast cancer. J Steroid Biochem Mol Biol. 2017;173:122–129. doi:10.1016/j.jsbmb.2016.12.001
- Scatena C, Roncella M, Di Paolo A, et al. Doxycycline, an inhibitor of mitochondrial biogenesis, effectively reduces cancer stem cells (CSCs) in early breast cancer patients: a clinical pilot study. Front Oncol. 2018;8:452. doi:10.3389/fonc.2018.00452
- Shimono Y, Zabala M, Cho RW, et al. Downregulation of miRNA-200c links breast cancer stem cells with normal stem cells. Cell. 2009;138(3):592–603. doi:10.1016/j.cell.2009.07.011
- Finlay-Schultz J, Cittelly DM, Hendricks P, et al. Progesterone downregulation of miR-141 contributes to expansion of stem-like breast cancer cells through maintenance of progesterone receptor and Stat5a. Oncogene. 2015;34(28):3676–3687. doi:10.1038/onc.2014.298
- Yu F, Jiao Y, Zhu Y, et al. MicroRNA 34c gene down-regulation via DNA methylation promotes self-renewal and epithelial-mesenchymal transition in breast tumor-initiating cells. J Biol Chem. 2012;287(1):465–473. doi:10.1074/jbc.M111.280768
- Zhu Y, Yu F, Jiao Y, et al. Reduced miR-128 in breast tumor-initiating cells induces chemotherapeutic resistance via Bmi-1 and ABCC5. Clin Cancer Res. 2011;17(22):7105–7115. doi:10.1158/1078-0432.CCR-11-0071
- Zhang Y, Eades G, Yao Y, Li Q, Zhou Q. Estrogen receptor α signaling regulates breast tumor-initiating cells by down-regulating miR-140 which targets the transcription factor SOX2. J Biol Chem. 2012;287(49):41514–41522. doi:10.1074/jbc.M112.404871
- Yu F, Yao H, Zhu P, et al. let-7 regulates self renewal and tumorigenicity of breast cancer cells. Cell. 2007;131(6):1109–1123. doi:10.1016/j.cell.2007.10.054
- Iliopoulos D, Lindahl-Allen M, Polytarchou C, Hirsch HA, Tsichlis PN, Struhl K. Loss of miR-200 inhibition of Suz12 leads to polycomb-mediated repression required for the formation and maintenance of cancer stem cells. Mol Cell. 2010;39(5):761–772. doi:10.1016/j.molcel.2010.08.013
- Yu F, Deng H, Yao H, Liu Q, Su F, Song E. Mir-30 reduction maintains self-renewal and inhibits apoptosis in breast tumor-initiating cells. Oncogene. 2010;29(29):4194–4204. doi:10.1038/onc.2010.167
- Tang W, Yu F, Yao H, et al. miR-27a regulates endothelial differentiation of breast cancer stem like cells. Oncogene. 2014;33(20):2629–2638. doi:10.1038/onc.2013.214
- Takahashi RU, Miyazaki H, Takeshita F, et al. Loss of microRNA-27b contributes to breast cancer stem cell generation by activating ENPP1. Nat Commun. 2015;6:7318. doi:10.1038/ncomms8318
- Zhang H, Cai K, Wang J, et al. MiR-7, inhibited indirectly by lincRNA HOTAIR, directly inhibits SETDB1 and reverses the EMT of breast cancer stem cells by downregulating the STAT3 pathway. Stem Cells. 2014;32(11):2858–2868. doi:10.1002/stem.1795
- Park EY, Chang E, Lee EJ, et al. Targeting of miR34a-NOTCH1 axis reduced breast cancer stemness and chemoresistance. Cancer Res. 2014;74(24):7573–7582. doi:10.1158/0008-5472.CAN-14-1140
- Wang Z, Li Y, Xiao Y, et al. Integrin α9 depletion promotes β-catenin degradation to suppress triple-negative breast cancer tumor growth and metastasis. Int J Cancer. 2019;145(10):2767–2780. doi:10.1002/ijc.32359
- Yang F, Cui P, Lu Y, Zhang X. Requirement of the transcription factor YB-1 for maintaining the stemness of cancer stem cells and reverting differentiated cancer cells into cancer stem cells. Stem Cell Res Ther. 2019;10(1):233. doi:10.1186/s13287-019-1360-4
- Korkaya H, Wicha MS. HER2 and breast cancer stem cells: more than meets the eye. Cancer Res. 2013;73(12):3489–3493. doi:10.1158/0008-5472.CAN-13-0260
- Damelin M, Bankovich A, Bernstein J, et al. A PTK7-targeted antibody-drug conjugate reduces tumor-initiating cells and induces sustained tumor regressions. Sci Transl Med. 2017;9(372):eaag2611. doi:10.1126/scitranslmed.aag2611
- Zhang S, Zhang H, Ghia EM, et al. Inhibition of chemotherapy resistant breast cancer stem cells by a ROR1 specific antibody. Proc Natl Acad Sci U S A. 2019;116(4):1370–1377. doi:10.1073/pnas.1816262116
- Ma X, Zhang Y, Kang Y, Li L, Zheng W. A recombinant protein TmSm(T34A) can inhibit proliferation and proapoptosis to breast cancer stem cells(BCSC) by down-regulating the expression of Cyclin D1. Biomed Pharmacother. 2016;84:373–381. doi:10.1016/j.biopha.2016.08.066
- Weigand S, Herting F, Maisel D, et al. Global quantitative phosphoproteome analysis of human tumor xenografts treated with a CD44 antagonist. Cancer Res. 2012;72(17):4329–4339. doi:10.1158/0008-5472.CAN-12-0136
- Willingham SB, Volkmer J-P, Gentles AJ, et al. The CD47-signal regulatory protein alpha (SIRPa) interaction is a therapeutic target for human solid tumors. Proc Natl Acad Sci. 2012;109(17):6662–6667. doi:10.1073/pnas.1121623109
- Givens BE, Naguib YW, Geary SM, Devor EJ, Salem AK. Nanoparticle-based delivery of CRISPR/Cas9 genome-editing therapeutics. Aaps j. 2018;20(6):108. doi:10.1208/s12248-018-0267-9
- Liu C, Zhang L, Liu H, Cheng K. Delivery strategies of the CRISPR-Cas9 gene-editing system for therapeutic applications. J Control Release. 2017;266:17–26. doi:10.1016/j.jconrel.2017.09.012
- Shen S, Xia JX, Wang J. Nanomedicine-mediated cancer stem cell therapy. Biomaterials. 2016;74:1–18. doi:10.1016/j.biomaterials.2015.09.037
- Gao J, Li W, Guo Y, Feng SS. Nanomedicine strategies for sustained, controlled and targeted treatment of cancer stem cells. Nanomedicine (Lond). 2016;11(24):3261–3282. doi:10.2217/nnm-2016-0261
- Liu C, Zhao G, Liu J, et al. Novel biodegradable lipid nano complex for siRNA delivery significantly improving the chemosensitivity of human colon cancer stem cells to paclitaxel. J Control Release. 2009;140(3):277–283. doi:10.1016/j.jconrel.2009.08.013
- Shi S, Han L, Gong T, Zhang Z, Sun X. Systemic delivery of microRNA-34a for cancer stem cell therapy. Angew Chem. 2013;52(14):3901–3905. doi:10.1002/anie.201208077
- Batra H, Pawar S, Bahl D. Curcumin in combination with anti-cancer drugs: a nanomedicine review. Pharmacol Res. 2019;139:91–105. doi:10.1016/j.phrs.2018.11.005
- Saeg F, Anbalagan M. Breast cancer stem cells and the challenges of eradication: a review of novel therapies. Stem Cell Investig. 2018;5:39. doi:10.21037/sci.22018.21010.21005
- Park SY, Choi JH, Nam JS. Targeting cancer stem cells in triple-negative breast cancer. Cancers. 2019;11(7):965. doi:10.3390/cancers11070965
- Misra R, Kandoi S, Varadaraj S, Vijayalakshmi S, Nanda A, Verma RS. Nanotheranostics: a tactic for cancer stem cells prognosis and management. J Drug Deliv Sci Technol. 2020;55:101457. doi:10.1016/j.jddst.2019.101457
- Stagni V, Kaminari A, Sideratou Z, Sakellis E, Vlahopoulos SA, Tsiourvas D. Targeting breast cancer stem-like cells using chloroquine encapsulated by a triphenylphosphonium-functionalized hyperbranched polymer. Int J Pharm. 2020;585:119465. doi:10.1016/j.ijpharm.2020.119465
- Zhao Y, Zhao W, Lim YC, Liu T. Salinomycin-loaded gold nanoparticles for treating cancer stem cells by ferroptosis-induced cell death. Mol Pharm. 2019;16(6):2532–2539. doi:10.1021/acs.molpharmaceut.9b00132
- Conley SJ, Baker TL, Burnett JP, et al. CRLX101, an investigational camptothecin-containing nanoparticle-drug conjugate, targets cancer stem cells and impedes resistance to antiangiogenic therapy in mouse models of breast cancer. Breast Cancer Res Treat. 2015;150(3):559–567. doi:10.1007/s10549-015-3349-8
- Gener P, Montero S, Xandri-Monje H, et al. Zileuton™ loaded in polymer micelles effectively reduce breast cancer circulating tumor cells and intratumoral cancer stem cells. Nanomedicine: NBM. 2020;24:102106. doi:10.1016/j.nano.2019.102106
- Zhao Y, Alakhova DY, Zhao X, Band V, Batrakova EV, Kabanov AV. Eradication of cancer stem cells in triple negative breast cancer using doxorubicin/pluronic polymeric micelles. Nanomedicine: NBM. 2020;24:102124. doi:10.1016/j.nano.2019.102124
- Li R-J, Ying X, Zhang Y, et al. All-trans retinoic acid stealth liposomes prevent the relapse of breast cancer arising from the cancer stem cells. J Control Release. 2011;149(3):281–291. doi:10.1016/j.jconrel.2010.10.019
- Tan T, Wang Y, Wang H, et al. Apoferritin nanocages loading mertansine enable effective eradiation of cancer stem-like cells in vitro. Int J Pharm. 2018;553(1–2):201–209. doi:10.1016/j.ijpharm.2018.10.038
- Rao W, Wang H, Han J, et al. Chitosan-decorated doxorubicin-encapsulated nanoparticle targets and eliminates tumor reinitiating cancer stem-like cells. Acs Nano. 2015;9(6):5725–5740. doi:10.1021/nn506928p
- Mamaeva V, Niemi R, Beck M, et al. Inhibiting notch activity in breast cancer stem cells by glucose functionalized nanoparticles carrying γ-secretase inhibitors. Mol Ther. 2016;24(5):926–936. doi:10.1038/mt.2016.42
- Qiao S, Zhao Y, Geng S, et al. A novel double-targeted nondrug delivery system for targeting cancer stem cells. Int J Nanomedicine. 2016;11:6667–6678. doi:10.2147/IJN.S116230
- Li H, Yan W, Suo X, et al. Nucleus-targeted nano delivery system eradicates cancer stem cells by combined thermotherapy and hypoxia-activated chemotherapy. Biomaterials. 2019;200:1–14. doi:10.1016/j.biomaterials.2019.01.048
- Liu Y, Suo X, Peng H, et al. Multifunctional magnetic nanoplatform eliminates cancer stem cells via inhibiting the secretion of extracellular heat shock protein 90. Adv Healthcare Mater. 2019;8(13):1900160. doi:10.1002/adhm.201900160
- Fonseca NA, Rodrigues AS, Rodrigues-Santos P, et al. Nucleolin overexpression in breast cancer cell sub-populations with different stem-like phenotype enables targeted intracellular delivery of synergistic drug combination. Biomaterials. 2015;69:76–88. doi:10.1016/j.biomaterials.2015.08.007
- Lin X, Chen W, Wei F, Zhou BP, Hung MC, Xie X. Nanoparticle delivery of miR-34a eradicates long-term-cultured breast cancer stem cells via targeting C22ORF28 directly. Theranostics. 2017;7(19):4805–4824. doi:10.7150/thno.20771
- Rafael D, Gener P, Andrade F, et al. AKT2 siRNA delivery with amphiphilic-based polymeric micelles show efficacy against cancer stem cells. Drug Deliv. 2018;25(1):961–972. doi:10.1080/10717544.2018.1461276
- Reshadmanesh A, Rahbarizadeh F, Ahmadvand D, Jafari Iri Sofla F. Evaluation of cellular and transcriptional targeting of breast cancer stem cells via anti-HER2 nanobody conjugated PAMAM dendrimers. Artif Cells, Nanomed Biotechnol. 2018;46(sup3):S105–S115. doi:10.1080/21691401.2018.1489269
- Sun Y, Kim HS, Saw PE, Jon S, Moon WK. Targeted therapy for breast cancer stem cells by liposomal delivery of siRNA against fibronectin EDB. Adv Healthcare Mater. 2015;4(11):1675–1680. doi:10.1002/adhm.201500190
- Yi Y, Kim HJ, Zheng M, et al. Glucose-linked sub-50-nm unimer polyion complex-assembled gold nanoparticles for targeted siRNA delivery to glucose transporter 1-overexpressing breast cancer stem-like cells. J Control Release. 2019;295:268–277. doi:10.1016/j.jconrel.2019.01.006
- Borah A, Pillai SC, Rochani AK, et al. GANT61 and curcumin-loaded PLGA nanoparticles for GLI1 and PI3K/Akt-mediated inhibition in breast adenocarcinoma. Nanotechnology. 2020;31(18):185102. doi:10.1088/1361-6528/ab6d20
- Samson AAS, Park S, Kim SY, Min DH, Jeon NL, Song JM. Liposomal co-delivery-based quantitative evaluation of chemosensitivity enhancement in breast cancer stem cells by knockdown of GRP78/CLU. J Liposome Res. 2019;29(1):44–52. doi:10.1080/08982104.2017.1420081
- Liu J, Meng T, Yuan M, et al. MicroRNA-200c delivered by solid lipid nanoparticles enhances the effect of paclitaxel on breast cancer stem cell. Int J Nanomedicine. 2016;11:6713–6725. doi:10.2147/IJN.S111647
- Kim YJ, Liu Y, Li S, et al. Co-eradication of breast cancer cells and cancer stem cells by cross-linked multilamellar liposomes enhances tumor treatment. Mol Pharm. 2015;12(8):2811–2822. doi:10.1021/mp500754r
- Zhang J, Kinoh H, Hespel L, et al. Effective treatment of drug resistant recurrent breast tumors harboring cancer stem-like cells by staurosporine/epirubicin co-loaded polymeric micelles. J Control Release. 2017;264:127–135. doi:10.1016/j.jconrel.2017.08.025
- Gao J, Liu J. Co-delivery of docetaxel and salinomycin to target both breast cancer cells and stem cells by PLGA/TPGS nanoparticles. Int J Nanomedicine. 2019;14:9199–9216. doi:10.2147/IJN.S230376
- Ibiyeye KM, Zuki ABZ. Cockle shell-derived aragonite CaCO3 nanoparticles for co-delivery of doxorubicin and thymoquinone eliminates cancer stem cells. Int J Mol Sci. 2020;21(5):1900. doi:10.3390/ijms21051900
- Lang T, Liu Y, Zheng Z, et al. Cocktail strategy based on spatio-temporally controlled nano device improves therapy of breast cancer. Adv Mater. 2019;31(5):1806202. doi:10.1002/adma.201806202
- Muntimadugu E, Kumar R, Saladi S, Rafeeqi TA, Khan W. CD44 targeted chemotherapy for co-eradication of breast cancer stem cells and cancer cells using polymeric nanoparticles of salinomycin and paclitaxel. Colloids Surf B Biointerfaces. 2016;143:532–546. doi:10.1016/j.colsurfb.2016.03.075
- Pan Y, Zhou S, Li Y, Parshad B, Li W, Haag R. Novel dendritic polyglycerol-conjugated, mesoporous silica-based targeting nanocarriers for co-delivery of doxorubicin and tariquidar to overcome multidrug resistance in breast cancer stem cells. J Control Release. 2020.
- Chen D, Wang G, Song W, Zhang Q. Novel CD44 receptor targeting multifunctional “nano-eggs” based on double pH-sensitive nanoparticles for co-delivery of curcumin and paclitaxel to cancer cells and cancer stem cells. J Nanopart Res. 2015;17(10). doi:10.1007/s11051-015-3217-9
- Wang H, Agarwal P, Zhao S, et al. Hyaluronic acid-decorated dual responsive nanoparticles of Pluronic F127, PLGA, and chitosan for targeted co-delivery of doxorubicin and irinotecan to eliminate cancer stem-like cells. Biomaterials. 2015;72:74–89. doi:10.1016/j.biomaterials.2015.08.048
- Wang H, Agarwal P, Zhao ST, Yu JH, Lu XB, He XM. Combined cancer therapy with hyaluronan-decorated fullerene-silica multifunctional nanoparticles to target cancer stem-like cells. Biomaterials. 2016;97:62–73. doi:10.1016/j.biomaterials.2016.04.030
- Yang Z, Sun N, Cheng R, Zhao C, Liu J, Tian Z. Hybrid nanoparticles coated with hyaluronic acid lipoid for targeted co-delivery of paclitaxel and curcumin to synergistically eliminate breast cancer stem cells. J Mater Chem. 2017;5(33):6762–6775. doi:10.1039/C7TB01510K
- Hu K, Zhou H, Liu Y, et al. Hyaluronic acid functional amphipathic and redox-responsive polymer particles for the co-delivery of doxorubicin and cyclopamine to eradicate breast cancer cells and cancer stem cells. Nanoscale. 2015;7(18):8607–8618. doi:10.1039/C5NR01084E
- Yu Y, Kanwar SS, Patel BB, Nautiyal J, Sarkar FH, Majumdar AP. Elimination of colon cancer stem-like cells by the combination of curcumin and FOLFOX. Transl Oncol. 2009;2(4):321–328. doi:10.1593/tlo.09193
- Fong D, Yeh A, Naftalovich R, Choi TH, Chan MM. Curcumin inhibits the side population (SP) phenotype of the rat C6 glioma cell line: towards targeting of cancer stem cells with phytochemicals. Cancer Lett. 2010;293(1):65–72. doi:10.1016/j.canlet.2009.12.018
- Kakarala M, Brenner DE, Korkaya H, et al. Targeting breast stem cells with the cancer preventive compounds curcumin and piperine. Breast Cancer Res Treat. 2010;122(3):777–785. doi:10.1007/s10549-009-0612-x
- Mors WB, do Nascimento MC, Parente J, da Silva MH, Melo PA, Suarez-Kurtz G. Neutralization of lethal and myotoxic activities of South American rattlesnake venom by extracts and constituents of the plant Eclipta prostrata (Asteraceae). Toxicon. 1989;27(9):1003–1009. doi:10.1016/0041-0101(89)90151-7
- Benes P, Knopfova L, Trcka F, et al. Inhibition of topoisomerase IIα: novel function of wedelolactone. Cancer Lett. 2011;303(1):29–38. doi:10.1016/j.canlet.2011.01.002
- Fu YZ, Yan YY, He M, et al. Salinomycin induces selective cytotoxicity to MCF-7 mammosphere cells through targeting the Hedgehog signaling pathway. Oncol Rep. 2016;35(2):912–922. doi:10.3892/or.2015.4434
- An H, Kim JY, Lee N, Cho Y, Oh E, Seo JH. Salinomycin possesses anti-tumor activity and inhibits breast cancer stem-like cells via an apoptosis-independent pathway. Biochem Biophys Res Commun. 2015;466(4):696–703. doi:10.1016/j.bbrc.2015.09.108
- Kalota A, Shetzline SE, Gewirtz A. Progress in the development of nucleic acids therapeutics for cancer. Cancer Biol Ther. 2004;3(1):4–12. doi:10.4161/cbt.3.1.517
- Cheng GZ, Chan J, Wang Q, Zhang W, Sun CD, Wang LH. Twist transcriptionally up-regulates AKT2 in breast cancer cells leading to increased migration, invasion, and resistance to paclitaxel. Cancer Res. 2007;67(5):1979–1987. doi:10.1158/0008-5472.CAN-06-1479
- Umemura S, Mimaki S, Makinoshima H, et al. Therapeutic priority of the PI3K/AKT/mTOR pathway in small cell lung cancers as revealed by a comprehensive genomic analysis. J Thorac Oncol. 2014;9(9):1324–1331. doi:10.1097/JTO.0000000000000250
- Rafael D, Doktorovová S, Florindo HF, et al. EMT blockage strategies: targeting Akt dependent mechanisms for breast cancer metastatic behaviour modulation. Curr Gene Ther. 2015;15(3):300–312. doi:10.2174/1566523215666150126123642
- Kendellen MF, Bradford JW, Lawrence CL, Clark KS, Baldwin AS. Canonical and non-canonical NF-κB signaling promotes breast cancer tumor-initiating cells. Oncogene. 2014;33(10):1297–1305. doi:10.1038/onc.2013.64
- Liu M, Sakamaki T, Casimiro MC, et al. The canonical NF-kappaB pathway governs mammary tumorigenesis in transgenic mice and tumor stem cell expansion. Cancer Res. 2010;70(24):10464–10473. doi:10.1158/0008-5472.CAN-10-0732
- Rao DD, Vorhies JS, Senzer N, Nemunaitis J. siRNA vs. shRNA: similarities and differences. Adv Drug Deliv Rev. 2009;61(9):746–759. doi:10.1016/j.addr.2009.04.004
- Yang Z, Sun N, Cheng R, et al. pH multistage responsive micellar system with charge-switch and PEG layer detachment for co-delivery of paclitaxel and curcumin to synergistically eliminate breast cancer stem cells. Biomaterials. 2017;147:53–67. doi:10.1016/j.biomaterials.2017.09.013
- Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science. 2009;324(5930):1029–1033. doi:10.1126/science.1160809
- Hu C, Niestroj M, Yuan D, Chang S, Chen J. Treating cancer stem cells and cancer metastasis using glucose-coated gold nanoparticles. Int J Nanomed. 2015;10:2065–2077. doi:10.2147/IJN.S72144