Abstract
Evidence continues to accumulate showing that tumors contain a minority population of cells responsible for tumor initiation, growth, and recurrence. These are termed “cancer stem cells” (CSCs). Functional assays have identified the self-renewal and tumor-initiation capabilities of CSCs. Moreover, recent studies have revealed that these CSCs is responsible for chemotherapy resistance within a tumor. Several mechanisms of chemoresistance have been proposed, including increased Wnt/â-catenin and Notch signaling, as well as high expression levels of adenosine triphosphate-binding cassette transporters, an active DNA repair capacity, and slow rate of self-renewal. Nanoscale drug-delivery systems, which transport therapeutically active molecules, prolong circulation, and improve biodistribution in the body, may allow more effective and specific therapies to address the challenges posed by CSCs. In particular, some nanovehicles are being exploited for selective drug delivery to CSCs and show promising results. In this review, we highlight the mechanisms of drug resistance and the novel strategies using nanoscale drugs to eliminate CSCs.
Introduction
Cancer is becoming more recognized as a heterogeneous disease with hierarchies of cellular populations that demonstrate a range of differentiation phenotypes. The majority of cells in bulk tumors may be non-tumorigenic end cells, and only a small subpopulation of cells within tumors is responsible for tumor initiation, growth, and recurrence. These are called “cancer stem cells” (CSCs).Citation1,Citation2 CSCs possess both self-renewal and differentiation capabilities.Citation3 Several signaling pathways are involved in the self-renewal behavior of CSCs, including Wnt/â-catenin, Notch, and hedgehog signaling, which mediate the resistances against radiotherapy and chemotherapy.Citation4 Despite the moderate success of currently available therapeutic approaches to tumors, they have several limitations. One of the main therapy drawbacks is that there is insufficient elimination of CSCs. Further, frequently there is multiple drug resistance (MDR) with advanced tumors.Citation5 Surviving CSCs will lead to tumor recurrence. Therefore, attention has been focused on defining new agents and novel therapies for cancer prevention and therapy by eliminating CSCs.
Nanoscale drug-delivery systems for cancer therapeutics are rapidly evolving and may offer an innovative approach to overcome the drug resistances of CSCs. Recently, nanoparticle-based strategies have demonstrated enhanced therapeutic efficacy and reduced adverse side effects, compared with those of classical therapeutic methods. Properly designed nanoparticles have the ability to significantly accumulate in tumor tissues by extravasation of nanoparticulates through fenestrated tumor vasculature via either passive or active targeting.Citation6 Moreover, loaded-drug formulations show excellent tumor cell uptake characteristics, block drug efflux from cancer cells, and reverse the MDR of tumors.Citation7,Citation8
To improve the outcome of cancer treatments, we need to comprehend characteristics of CSCs, and propose new strategies to eliminate CSCs based on the available literature.
CSCs
Despite the ongoing debate over CSCs exist, there is no denying that most cancers are heterogeneous and show functional and phenotypical differences at the cell population level. These observations may result from clonal evolution driven by the differentiation of CSCs or from genetic instability.Citation9,Citation10 Moreover, CSCs can vary between different patient tumors and can constantly change as the disease progresses. Therefore, for cancer prevention and treatment, we need to identify and characterize these subpopulations. CSCs may display certain properties; CSC subpopulations: can be isolated based on cell surface marker profiles,Citation11 exhibit increased resistance against conventional radiotherapy and chemotherapy,Citation12,Citation13 and may initiate tumors at limiting dilutions in animals.Citation14 These characteristics imply the existence of a distinct fraction of cancer cells that have a self-renewal property and the potential to cause tumors with only a limited number of cells.
CSCs were first observed in acute myeloid leukemia (AML). There is a small fraction of AML cells with a surface marker phenotype of CD34+CD38- that is able to recapitulate the phenotypes of the original patient tumor in immune-deficient mice.Citation15 Although new studies have revealed additional unexpected heterogeneity of severe combined immunodeficiency leukemia-initiating cells, CD34–, Lin+, CD38+, and CD45RA+ fractions have the capacity to form xenografted tumors. Subsequently, CSCs were demonstrated to exist in solid tumors, including those formed by brain, breast, colon, prostate, pancreatic, lung, liver, melanoma and ovarian cancers ().Citation16–Citation31 These cells express markers of stemness and are capable of reproducing the cancer in mouse models. In breast cancer, CD44+CD24− and aldehyde dehydrogenase 1 (ALDH1)+ have been demonstrated to be selective phenotypes that enrich CSCs.Citation26,Citation32 Recently, the CD133+ cell subpopulation was found to harbor brain CSCs.Citation33 However, several studies suggest that the glycosylation status of CD133 molecule, rather than the expression of the CD133 protein itself, appears to be a marker for CSC phenotypes. Additionally, CD44 is often expressed as a variety of isoforms. CD44v is highly expressed in certain cancers and CD44v6 has been targeted for cancer therapy.
Table 1 Identification of CSCs using surface markers
CSCs have also been identified by tumorsphere culture.Citation34 Similar to the forming of mouse embryonic fibroblasts and neurospheres in suspension culture, CSCs may grow in the absence of serum and without attachment to culture plates, whereas differentiated cells fail to survive under the same conditions.Citation35 Moreover, CSCs are capable of forming subsequent passages of tumorspheres and multi-lineage differentiation. These properties can be used to identify the self-renewal capacity of CSCs for treatment with or without drugs in vitro.
Self-renewal pathways of CSCs
CSCs produce tumors through self-renewal and differentiation regulated by several signaling pathways ().Citation36–Citation42 Understanding the mechanisms of self-renewal in CSCs is of great importance for drug discovery and development. Wnt/â-catenin signaling is one of the key pathways that modulates CSC self-renewal.Citation43,Citation44 Activation of Wnt-target genes depends on mediation by â-catenin, which enters the nucleus from the cytoplasm, then cooperates with the TCF/LEF transcription factor, eventually resulting in the activation of Wnt-target genes such as cyclin D1, c-Jun, and c-Myc.Citation44,Citation45 Beta-catenin protein is degraded by the ubiquitin–proteasome pathway through phosphorylation at Ser33/Ser37/Thr41 by GSK3â.Citation46,Citation47 The Wnt/â-catenin pathway is implicated in the maintenance of CSC self-renewal in leukemia, melanoma, and breast, lung, and liver cancers.Citation48–Citation52 It has been reported that a high level of â-catenin increases the drug resistance of numerous tumor types,Citation53 indicating that dysregulation of â-catenin plays a crucial role in cancer treatment. If the transcriptional activity of â-catenin can be significantly inhibited, cancer growth will be suppressed. Therefore, it is critical that agents be found that can directly target â-catenin and its downstream molecules.
Table 2 Signaling pathways involved in CSCs self-renewal
Notch signaling is another important signaling pathway involved in modulation of CSC self-renewal.Citation54–Citation57 Four Notch genes (Notch 1 to 4) have been identified in mammals, which act as transmembrane receptors for Jagged 1 and 2, and Delta-like 1, 3, and 4.Citation58 The binding of ligands to Notch results in its cleavage by A disintegrin and metalloprotease (ADAM) family members and ã-secretase.Citation59 The intracellular domain of Notch translocates to the nucleus, where it activates downstream target genes such as cyclin D1, c-Myc, and nuclear factor kappa B(NF-kB).Citation60 Recent studies have shown that the Notch pathway is upregulated in CSCs, leading to uncontrolled self-renewal. Specifically, Notch 1 has been reported to cross-talk with Wnt/â-catenin signaling in diverse cellular situations and the interaction between Notch and Wnt/â-catenin pathways suggests that Notch is probably involved in CSC-related tumor recurrence following therapy.
Other pathways, such as the hedgehog-signaling pathway, can also maintain the self-renewal of CSCs.Citation61–Citation65 The hedgehog pathway has been identified to be involved in CSC self-renewal and tumorigenicity in human breast cancer.Citation39 Further, a previous report showed that the hedgehog pathway is associated with NF-kB signaling, indicating that sonic hedgehog might be activated by the transcription factor NF-kB.Citation66
CSCs and drug resistance
To explain why chemotherapies ultimately fail to cure cancer, the CSC hypothesis suggests that the therapeutic resistance of CSCs in tumors might be the mechanism. Such CSC characteristics might result from several situations, including high expression levels of adenosine triphosphate-binding cassette (ABC) transporters, resistance against apoptosis, an active DNA repair capacity, and slow rate of self-renewal. Studies of cancer cells have revealed that CSCs commonly express high levels of drug efflux pumps.Citation67,Citation68 Such drug pumps are responsible for protecting cancer cells from damage by cytotoxic chemotherapies via efflux pumping mechanisms.Citation69 Therefore, as a result of these biophysical and biological properties, CSCs are rendered resistant against chemotherapeutic agents.
Additional studies have revealed that CSCs extrude the fluorescent dye rhodamine 123 and Hoechst 33342. The cells that efflux Hoechst 33342 can be detected by flow cytometry and are called “side population” (SP) cells.Citation70 The Hoechst efflux assay has successfully identified SP cells in various solid tissues including breast, pancreatic, and liver.Citation71–Citation73 Moreover, studies have confirmed that chemoresistant cancer cells contain a higher proportion of SP cells than chemosensitive cells.Citation74 Recent findings suggest that other transporters, including octreotide (Oct) 1, also contribute to CSC resistance against certain drugs. Maddox et al showed that Oct1 controls multiple stem cell phenotypes in both normal and tumor cells.Citation75 Elevated Oct1 protein expression correlates with elevated CD24–CD44+ or ALDHhi CSC populations in breast cancer tissues. Genes associated with drug efflux pumps, such as Abcg2, Abcb1 and Abcb4, are directly regulated by Oct1. Furthermore, Oct1 knockdown specifically decreases the number of SP cells among A549 cells.
In addition to possessing an increased capacity for cytotoxic agent efflux, CSCs are identified by their characteristic slow-cycling and quiescent properties.Citation76 These cells, also termed “label-retaining” cells, can be purified by functional assays.Citation77,Citation78 Such a small subset of CSCs mostly remains quiescent in the G0 phase. Over time, CSCs are induced to divide and produce transit-ampliflying cells by stimuli. Subsequently, some of these transit-ampliflying cells differentiate into new mature cancer cells with a chemoresistant phenotype.Citation79 These observations have been confirmed in CSCs derived from AML and solid tumors.Citation80,Citation81 The acquired chemoresistance of cancer, which corresponds with the presence of CSCs, increases greatly after chemotherapy in the clinic. Ultimately, patients at this stage will develop recurrent tumors and fail to be responsive to further treatment by chemotherapy.
In a third model of acquired resistance, drug-resistant variants of CSCs or their close descendants arise, which produce a population of DNA-repairing tumor cells.Citation82,Citation83 A previous study has revealed that CD133+ cells express > 30-fold higher levels of the DNA repair protein O6-methylguanine-DNA methyltransferase (MGMT) than matched CD133− gliomas.Citation13 Because of the increased DNA repair capacity, CD133+ cells are more resistant to radiotherapy than CD133− cells. Moreover, patients with high expression levels of MGMT demonstrate significantly reduced median survival times compared with patients with low MGMT expression.Citation84,Citation85 These results suggest that DNA repair may be a target for the elimination of CSCs to facilitate the survival of patients.
CSCs expressing elevated levels of ALDH1, (a molecular metabolic mediator) show resistance against cytotoxic agents.Citation86 ALDH1 is a detoxification enzyme involved in catalyzing the oxidation of acetaldehydes produced from ethanol.Citation87 As a detoxification enzyme, overexpression of ALDH1 in CSCs has implications in the resistances against chemotherapeutic drugs such as cyclophosphamide. Furthermore, high ALDH1 activity in cancer is associated with a poor outcome,Citation88 suggesting that chemoresistant molecules expressed by CSCs will directly affect patient prognoses.
Nanomedical strategies for cancer stem cell therapy
The existence of CSCs has important implications for chemo-prevention and treatment of cancer. CSCs are more resistant to treatment than bulk cells, meaning that conventional chemotherapies for cancer often fail. Strategies to address this concern include new approaches and therapeutic agents for the reversal of chemotherapy resistance by targeting CSCs. Nanomedicine offers an innovative approach to overcome these hurdles. The potential of nanomedicine lies in the ability to engineer formulations at the nanometer scale for loading chemotherapeutics or active molecules. In addition, the designed vehicles may sensitize or enhance therapeutic strategies that cater to the unique dynamics of cancer.
The nanovehicles that transport therapeutic drugs and facilitate cellular uptake based on self-assembled supramolecular differ according to the drug and nano-sized carrier. Their development depends on several key factors that govern interactions with the body, including the size, polarity, numbers and hydrophobic or hydrophilic nature of nanoparticles.Citation89 Nanovehicles prolong circulation and improve the biodistribution of the incorporated drug, yielding superior accumulation in tumors via a process known as “enhanced permeability and retention.”Citation90 The virtue of nanovehicles is that they can be adjusted using molecules without loss of activity. Moreover, nanovehicles are used to encapsulate chemotherapeutics, which can hide unfavorable domains between the drug and the body. Based on these advantages, the objective of nanomedicine is to develop new agents to provide beneficial pharmacological properties for eliminating CSCs.
Accordingly, nanomedicine for the targeting of CSCs requires that there be multidisciplinary cooperation to develop new agents as well as accurate interpretation of the data obtained from different disciplines. In particular, to harness the potential of nanobiology and nanomedicine, the properties of CSCs and their role in cancer progression need to be carefully understood. Therefore, novel nanomedicines will need to be created for the development of therapeutic strategies against CSCs.
Development of nanomedicine for CSCs
Drug-delivery systems for CSCs
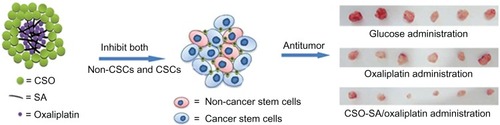
To improve the therapeutic effect on CSCs, nanoscaled drugs have enabled development of many novel strategies to overcome the known shortcomings of many anticancer drugs, such as drug extrusion, low aqueous solubility and stability, and high nonspecific toxicity.Citation91,Citation92 These nanoparticles include polymeric micelles and non-polymeric systems. Polymeric micelles with a core–shell structure can be formed by the self-aggregation of amphiphilic grafts in water, providing a significant advantage for delivery of cytotoxic agents to cancer.Citation93,Citation94 Previous studies have demonstrated that cell uptake of drugs is increased using nanovehicles compared with the free drug.Citation95 For example, we have developed a novel micelle formulation of oxaliplatin encapsulated in a chitosan vesicle (CSO-SA/OXA micelles).Citation96 These CSO-SA/OXA micelles show an excellent internalization ability that targets the tumor cell nucleus and increases the oxaliplatin concentration in tumor cells, which can reverse the drug resistance of CSCs and effectively eliminate CSCs in vitro and in vivo (). The uptake of nanovehicles may be via endocytosis in which the free drug is internalized into cancer cells by molecular diffusion. Using drug-loaded nanovehicles, an efficient route for drug delivery is penetration of the cell membrane, especially in chemoresistant tumor cells.
Figure 1 Oxaliplatin-incorporating micelles are effective for the eradication of cancer stem cells (CSCs).
Note: The drug-loaded CSO-SA micelles suppress both colorectal CSCs and bulk cancer cells, resulting in enhancement of antitumor efficacy.
Abbreviations: CSO, chitosan oligosaccharide; SA, stearic acid.
In another example, Zhang et al demonstrated the strong therapeutic potential of salinomycin-loaded polyethylene glycol-b-polycaprolactone (PEG-b-PCL) polymeric micelles (M-SAL) and octreotide (Oct)-modified paclitaxel (PTX)-loaded PEG-b-PCL polymeric micelles (Oct-M-PTX) in the treatment of breast CSCs.Citation97 Oct is an octapeptide analog of endogenous somatostatin and mainly binds to somatostatin receptors (SSTRs) that are overexpressed in many cancers. By coupling Oct, copolymer micelles can enhance their binding to SSTR-positive cancer cells and increase intracellular delivery of drugs. Combinatorial therapy using Oct-M-PTX plus M-SAL may eradicate breast cancer cells together with breast CSCs via receptor-mediated endocytosis.Citation97
Similarly, CSC persistence in chronic myeloid leukemia (CML) can also be targeted by vectorized nanocarriers. Bcr-Abl tyrosine-kinase inhibitors (TKIs) are the first-line therapy for most patients with CML. However, imatinib (a TKI) has been shown to be a substrate of ABCG2, and fails to cure end-stage patients. Zhou et al described that the resistance of CML CD34+ and primitive CD34+CD38- cells can be overcome using synthetic low-density lipoprotein (sLDL) particles.Citation98 sLDL is prepared using a solvent evaporation method involving a mixture consisting of phosphatidylcholine, triolein, cholesterol, and cholesteryl oleate at a molar ratio of 3:2:1:1, respectively. Low-density lipoprotein recep-tor-specific lipophilic synthetic peptides have been used to target CML cells. The results indicated that Bcr-Abl-positive cell lines show increased and preferential uptake of sLDL compared with Bcr-Abl negative cells.Citation98
Nanomedicine has the potential to overcome the known shortcomings of many anticancer agents, including low water-solubility, instability, and nonspecific toxicity. For example, curcumin has been reported to eliminate colorectal CSCs in vitro. However, its application in cancer treatment is limited by high hydrophobicity, instability, and poor pharmacokinetics. Nanoscale drugs offer an innovative approach to overcome such problems. For example, we have prepared a nanotechnology-based curcumin formulation and confirmed that it can self-assemble to form nanoscale micelles in an aqueous medium.Citation99 Nanoparticles increase the stability of curcumin by protecting the encapsulated curcumin against hydrolysis and biotransformation. Moreover, the formulation effectively inhibits CSCs and marginally suppresses tumor growth.
Targeting of signaling pathways in CSCs
Nanovehicles loaded with small molecules to target signaling pathways are another avenue toward the eradication of CSCs. Although surface markers are partly shared with normal stem cells, there are still many differences, including signaling pathway and metabolic alterations in CSCs, which may be exploited for selective targeted delivery of nanoscale drugs. The molecular targeting of deregulated signaling pathways, which may contribute to the chemoresistance of cancer, is currently under concerted investigation. The potential pathways include Wnt/â-catenin, hedgehog, and Notch signaling.Citation100,Citation101 Zhou et al recently designed an N-(2-hydroxy-propyl)methacrylamide (HPMA)-based delivery system for delivery of the hedgehog-signaling inhibitor cyclopamine that is a selective macromolecular therapeutic against CSCs.Citation102 However, the clinical use of cyclopamine is restricted by its high hydrophobicity and systemic toxicity. A HPMA copolymer has been synthesized by reversible addition-fragmentation chain transfer copolymerization of HPMA and 3-(N-methacryloyl- glycylphenylalanylleucylglycyl)-thiazolidine-2-thione (MA-GFLG-TT), followed by conjugation of cyclopamine to glycylphenylalanylleucylglycyl side chains. The HPMA copolymer-cyclopamine conjugate binds to cells via the smoothened (SMO) membrane receptor. The authors reported that the HPMA copolymer-cyclopamine conjugate shows a selective inhibitory effect on prostate CSCs in comparison with that on bulk cancer cells.
In another example, the application of ã-secretase inhibitors (GSIs) in cancer treatment to block Notch signaling is limited by their high hydrophobicity and side effects. Mamaeva et al developed mesoporous silica nanoparticles (MSNPs) as vehicles for targeted delivery of GSIs to block Notch signaling.Citation103 The folate receptor is overexpressed in many tumors, therefore, a GSI is encapsulated in MSNPs, and folate is covalently conjugated to the outside to target folate receptor-enriched cancer cells. The average size of synthesized MSNPs is 200 to 350 nm. GSI-loaded MSNPs efficiently target and block Notch activity, inhibit tumor growth, and CSC functions in vivo. These biocompatible and biodegradable MSNPs provide a novel platform for efficient small-molecule drug delivery for the development of refined Notch therapy.
Targeting of CSC regulatory pathways by RNA interference (RNAi) has been reported using nanodelivery systems to treat cancers. Lo et al developed nanodelivery of double-stranded DNA (dsDNA) encoding siRNA to efficiently downregulate the activity of EZH2 and Oct4 associated with CSC properties, which directly led to an anticancer effect.Citation104 By conjugating nuclear localization signal peptides, the efficacy of dsDNA encoding siRNA against EZH2 or Oct4 is enhanced because of the facilitated nuclear delivery. Treatment of head and neck squamous cancer cell xenografts with this formulation remarkably represses CSCs and enhances radiosensitivity, which may involve the Wnt pathway.
In the treatment of glioblastoma (GBM), promising therapeutic approaches include miR145 incorporated with polyurethane-short branch polyethylenimine (PU-PEI) to block key signal transduction pathways, which has been found to effectively mediate downregulation of Oct4 and Sox2 by targeting the Oct4 and Sox2 3′ untranslated regions in GBM CD133+ cells.Citation104 Moreover, real-time polymerase chain reaction analysis has shown that the expression of other stemness genes, such as Nanog, c-Myc, and the oncogene Bmi-1, are also downregulated by PU-PEI-miR145 treatment. These results suggest that PU-PEI-miR145 might suppress the self-renewal and tumor-initiating properties of GBM cells. Notably, miR145 delivery with a combination of radiotherapy (2 Gy) and temozolomide (200 mM) can eliminate tumor formation in vivo.Citation104 Thus, PU-PEI-miR145 treatment for CSC eradication is a potential therapeutic approach to improve current tumor treatments, especially for tumors that have developed a resistance against conventional therapy. Importantly, polymer-based gene delivery systems are considered to induce low immune responses and are potentially safer than viral-mediated delivery.Citation104
Destruction of CSCs and their niches
Apart from direct targeting of CSCs, various agents are being developed for targeting the microenvironments (niches) of cancer cells.Citation105 Maintenance of CSC self-renewal involves cross-talk between CSCs and their supporting stroma or vasculature. New evidence has revealed that CSC cross-talk with their supporting stroma favors tumor progression by promoting cell growth, proliferation, and drug resistance.Citation106 As such, disruption of the cross-talk between CSCs and their niches is an attractive approach for cancer treatment. An emerging strategy may be employed to design new nanoparticle-based combinatorial therapies for interference of the supportive microenvironmental cross-talk. Currently, many physicochemical treatment methods are being developed to enhance CSC-directed therapy to interfere with CSC niches. Wang et al prepared anti-CD133 monoclonal antibody-conjugated single-walled carbon nanotubes that selectively eradicate CD133+ GMB cells by releasing substantial heat in the nanoenvironment after irradiation with near-infrared laser light.Citation107 Anti-CD133 monoclonal antibody-conjugated single-walled carbon nanotubes have been demonstrated to be promising heat absorbers, and are used in photothermolysis of malignant cells.Citation107 After conjugation with an anti-CD133-Phycoerythrin (PE) antibody, nanoparticles retained their photonic features and targeted GBM CD133+ cells. In addition, the in vitro tumorigenic, spheroid body formation, and self-renewal capabilities of GBM CD133+ cells are effectively inhibited because of the localized hyperthermia. Similarly, Burke et al used the efficient heating rates of amide-functionalized multi-walled carbon under irradiation.Citation108 Stem and bulk breast cancer cells are equally sensitive to nanotube-mediated thermal treatment.Citation107 The mechanisms of nanotube thermal therapeutic effect are promotion of rapid membrane permeabilization and necrosis of CSCs.
Telomerase-based therapy of CSCs
Finally, it is noteworthy to mention telomerase-based approaches, as these have potential in nanomedicine-based therapeutics against CSCs. Telomerase is expressed in both bulk tumor cells and CSCs, but has only limited activity in normal tissues and acts as an immortalizing agent. Joseph et alCitation109 showed that imetelstat (a potent telomerase inhibitor) decreases telomerase activity and suppresses the self-renewal potential of breast CSCs. In addition, imetelstat treatment inhibits tumorigenicity of PANC1 and MDA-MB231 cells in vivo. However, telomerase inhibitors have biopharmaceutical problems such as high hydrophobicity, permeability, and instability. Thus, efficient delivery to target cells and tumors is required. Nanomedicine can overcome their biopharmaceutical shortcomings and ensure that sufficient bioavailability is provided. Although, as far as the authors are aware, no related articles have reported nanoparticles containing a telomerase inhibitor for CSC therapy, such an approach will no doubt be studied in the future.
Conclusion and future perspectives
New concepts of chemoresistance in cancer have been proposed, which involve the contribution of CSCs to treatment failure. Although the mechanisms responsible for chemotherapy resistance by CSCs have not been clearly identified, overexpression of ABC transporters, a slow rate of self-renewal, and an active DNA repair capacity are all possible pathological mechanisms. In particular, interpreting the cellular heterogeneity in tumors may help to delineate the resistance of cancers to conventional therapies.
Nonetheless, designing nanomedical therapies against CSCs has proven to be complex, possibly because CSCs in the same type of tumor are phenotypically and functionally heterogeneous and because of the nonspecific nature of CSC markers used for targeting. Moreover, CSCs are protected by multiple resistance mechanisms that make them less susceptible to conventional therapies. Nanoparticle-based drugs have the potential to enhance treatments by overcoming chemoresistance or targeting CSCs. We would like to emphasize that elucidating the signaling pathways in CSCs may drive the development of new targeting therapies. Furthermore, Ginsburg and Willard have reported that chemoresistance and treatment effects depend on the distinct patterns of genes associated with stemness/differentiation pathways.Citation110 Genomic signature (DNA or RNA) differences have recently been exploited to personalize medicine and CSCs, which may facilitate individual-specific nanomedicine and dose selection for better cancer treatment efficacy and patient prognoses.
Acknowledgment
This study was supported in part by the Zhejiang Provincial Natural Science Foundation of China (grant nos Z2100366, Y2100414, and Y2090386), Science and Technology Bureau of Zhejiang Province (Grant No. 2011C37004), Zhejiang Provincial Program for the Cultivation of High-Level Innovative Health Talents (JH), Stem Cell Engineering and Clinical Translational Medicine (Zhejiang Medical innovation disciplines, JH) and Doctoral Fund of Ministry of Education of China (20100101110124).
Disclosure
The authors declare no conflicts of interest in this work.
References
- AbbottACancer: the root of the problemNature2006442710474274316915262
- ReyaTMorrisonSJClarkeMFWeissmanILStem cells, cancer, and cancer stem cellsNature2001414685910511111689955
- GangemiRPaleariLOrengoAMCancer stem cells: a new paradigm for understanding tumor growth and progression and drug resistanceCurr Med Chem200916141688170319442140
- McCubreyJAAbramsSLStadelmanKTargeting signal transduction pathways to eliminate chemotherapeutic drug resistance and cancer stem cellsAdv Enzyme Regul201050128530719895837
- RichJNBaoSChemotherapy and cancer stem cellsCell Stem Cell20071435335518371369
- LiuYMiyoshiHNakamuraMNanomedicine for drug delivery and imaging: a promising avenue for cancer therapy and diagnosis using targeted functional nanoparticlesInt J Cancer2007120122527253717390371
- ShapiraALivneyYDBroxtermanHJAssarafYGNanomedicine for targeted cancer therapy: towards the overcoming of drug resistanceDrug Resist Updat201114315016321330184
- XuYYDuYZYuanHLiuLNNiuYPHuFQImproved cytotoxicity and multidrug resistance reversal of chitosan based polymeric micelles encapsulating oxaliplatinJ Drug Target201119534435320854014
- DalerbaPChoRWClarkeMFCancer stem cells: models and conceptsAnnu Rev Med20075826728417002552
- ShackletonMQuintanaEFearonERMorrisonSJHeterogeneity in cancer: cancer stem cells versus clonal evolutionCell2009138582282919737509
- Ricci-VitianiLLombardiDGPilozziEIdentification and expansion of human colon-cancer-initiating cellsNature2007445712311111517122771
- TanSChenJSSunLJYa oHRSelective enrichment of hepatocellular cancer stem cells by chemotherapyJ Int Med Res20093741046105619761687
- BaoSWuQMcLendonREGlioma stem cells promote radioresistance by preferential activation of the DNA damage responseNature2006444712075676017051156
- VisvaderJELindemanGJCancer stem cells in solid tumours: accumulating evidence and unresolved questionsNat Rev Cancer200881075576818784658
- BonnetDDickJEHuman acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cellNat Med1997377307379212098
- FangJSDengYWLiMCIsolation and identification of brain tumor stem cells from human brain neuroepithelial tumorsZhonghua Yi Xue Za Zhi2007875298303 Chinese17456355
- DontuGEl-AshryDWichaMSBreast cancer, stem/progenitor cells and the estrogen receptorTrends Endocrinol Metab200415519319715223047
- CollinsATBerryPAHydeCStowerMJMaitlandNJProspective identification of tumorigenic prostate cancer stem cellsCancer Res20056523109461095116322242
- DalerbaPDyllaSJParkIKPhenotypic characterization of human colorectal cancer stem cellsProc Natl Acad Sci U S A200710424101581016317548814
- LiCLeeCJSimeoneDMIdentification of human pancreatic cancer stem cellsMethods Mol Biol200956816117319582426
- FangDNguyenTKLeishearKA tumorigenic subpopulation with stem cell properties in melanomasCancer Res200565209328933716230395
- BapatSAMaliAMKoppikarCBKurreyNKStem and progenitorlike cells contribute to the aggressive behavior of human epithelial ovarian cancerCancer Res20056583025302915833827
- EramoALottiFSetteGIdentification and expansion of the tumorigenic lung cancer stem cell populationCell Death Differ200815350451418049477
- YangZFHoDWNgMNSignificance of CD90+ cancer stem cells in human liver cancerCancer Cell200813215316618242515
- YeungTMGandhiSCWildingJLMuschelRBodmerWFCancer stem cells from colorectal cancer-derived cell linesProc Natl Acad Sci U S A201010783722372720133591
- HuangEHHynesMJZhangTAldehyde dehydrogenase 1 is a marker for normal and malignant human colonic stem cells (SC) and tracks SC overpopulation during colon tumorigenesisCancer Res20096983382338919336570
- HellstenRJohanssonMDahlmanASternerOBjartellAGaliellalactone inhibits stem cell-like ALDH-positive prostate cancer cellsPLoS One201167e2211821779382
- HermannPCHuberSLHerrlerTDistinct populations of cancer stem cells determine tumor growth and metastatic activity in human pancreatic cancerCell Stem Cell20071331332318371365
- HuangCPTsaiMFChangTHALDH-positive lung cancer stem cells confer resistance to epidermal growth factor receptor tyrosine kinase inhibitorsCancer Lett2013328114415122935675
- AlveroABChenRFuHHMolecular phenotyping of human ovarian cancer stem cells unravels the mechanisms for repair and chemoresistanceCell Cycle20098115816619158483
- LandenCNJrGoodmanBKatreAATargeting aldehyde dehydrogenase cancer stem cells in ovarian cancerMol Cancer Ther20109123186319920889728
- Al-HajjMWichaMSBenito-HernandezAMorrisonSJClarkeMFProspective identification of tumorigenic breast cancer cellsProc Natl Acad Sci U S A200310073983398812629218
- ErOCancer stem cells in solid tumorsOnkologie2009321060560919816080
- DontuGAbdallahWMFoleyJMIn vitro propagation and transcriptional profling of human mammary stem/progenitor cellsGenes Dev200317101253127012756227
- FanXOuyangNTengHYaoHIsolation and characterization of spheroid cells from the HT29 colon cancer cell lineInt J Colorectal Dis201126101279128521670985
- PalomeroTDominguezMFerrandoAAThe role of the PTEN/AKT Pathway in NOTCH1-induced leukemiaCell Cycle20087896597018414037
- KorkayaHPaulsonACharafe-JauffretERegulation of mammary stem/progenitor cells by PTEN/Akt/beta-catenin signalingPLoS Biol200976e100012119492080
- HambardzumyanDSquatritoMCarbajalEHollandECGlioma formation, cancer stem cells, and akt signalingStem Cell Rev20084320321018595010
- LiuSDontuGMantleIDHedgehog signaling and Bmi-1 regulate self-renewal of normal and malignant human mammary stem cellsCancer Res200666126063607116778178
- RaaphorstFMSelf-renewal of hematopoietic and leukemic stem cells: a central role for the Polycomb-group gene Bmi-1Trends Immunol2003241052252414552834
- GodlewskiJNowickiMOBroniszATargeting of the Bmi-1 oncogene/stem cell renewal factor by microRNA-128 inhibits glioma proliferation and self-renewalCancer Res200868229125913019010882
- PeñuelasSAnidoJPrieto-SánchezRMTGF-beta increases glioma-initiating cell self-renewal through the induction of LIF in human glioblastomaCancer Cell200915431532719345330
- Kawaguchi-IharaNMurohashiINaraNTohdaSPromotion of the self-renewal capacity of human acute leukemia cells by Wnt3AAnticancer Res2008285A2701270419035298
- LiuSDontuGWichaMSMammary stem cells, self-renewal pathways, and carcinogenesisBreast Cancer Res200573869515987436
- KatohMWNT signaling pathway and stem cell signaling networkClin Cancer Res200713144042404517634527
- CleversHWnt/beta-catenin signaling in development and diseaseCell2006127346948017081971
- LiuCLiYSemenovMControl of beta-catenin phosphorylation/degradation by a dual-kinase mechanismCell2002108683784711955436
- YsebaertLChicanneGDemurCExpression of beta-catenin by acute myeloid leukemia cells predicts enhanced clonogenic capacities and poor prognosisLeukemia20062071211121616688229
- LiYWelmBPodsypaninaKEvidence that transgenes encoding components of the Wnt signaling pathway preferentially induce mammary cancers from progenitor cellsProc Natl Acad Sci U S A200310026158531585814668450
- ChienAJMooreECLonsdorfASActivated Wnt/beta-catenin signaling in melanoma is associated with decreased proliferation in patient tumors and a murine melanoma modelProc Natl Acad Sci U S A200910641193119819144919
- YangWYanHXChenLWnt/beta-catenin signaling contributes to activation of normal and tumorigenic liver progenitor cellsCancer Res200868114287429518519688
- TengYWangXWangYMaDWnt/beta-catenin signaling regulates cancer stem cells in lung cancer A549 cellsBiochem Biophys Res Commun2010392337337920074550
- DeanMFojoTBatesSTumour stem cells and drug resistanceNat Rev Cancer20055427528415803154
- Charafe-JauffretEMonvilleFGinestierCDontuGBirnbaumDWichaMSCancer stem cells in breast: current opinion and future challengesPathobiology2008752758418544962
- GurselDBBerryNBoockvarJAThe contribution of Notch signaling to glioblastoma via activation of cancer stem cell self-renewal: the role of the endothelial networkNeurosurgery2012702N19N2122251985
- SikandarSSPateKTAndersonSNOTCH signaling is required for formation and self-renewal of tumor-initiating cells and for repression of secretory cell differentiation in colon cancerCancer Res20107041469147820145124
- LeeC WSiminKLiuQA functional Notch-survivin gene signature in basal breast cancerBreast Cancer Res2008106R9719025652
- MummJSKopanRNotch signaling: from the outside inDev Biol2000228215116511112321
- WuJYRaoYFringe: defining borders by regulating the notch pathwayCurr Opin Neurobiol19999553754310508746
- BorggrefeTOswaldFThe Notch signaling pathway: transcriptional regulation at Notch target genesCell Mol Life Sci200966101631164619165418
- ClementVSanchezPde TriboletNRadovanovicIRuiz i AltabaAHEDGEHOG-GLI1 signaling regulates human glioma growth, cancer stem cell self-renewal, and tumorigenicityCurr Biol200717216517217196391
- HuangFTZhuan-SunYXZhuangYYInhibition of hedgehog signaling depresses self-renewal of pancreatic cancer stem cells and reverses chemoresistanceInt J Oncol20124151707171422923052
- Kawaguchi-IharaNOkuhashiYItohMMurohashiINaraNTohdaSPromotion of the self-renewal capacity of human leukemia cells by sonic hedgehog proteinAnticancer Res201131378178421498696
- FerruzziPMennilloFDe RosaAIn vitro and in vivo characterization of a novel Hedgehog signaling antagonist in human glioblastoma cell linesInt J Cancer20121312E33E4422072503
- MazumdarTDeVecchioJShiTJonesJAgyemanAHoughtonJAHedgehog signaling drives cellular survival in human colon carcinoma cellsCancer Res20117131092110221135115
- NakashimaHNakamuraMYamaguchiHNuclear factor-kappaB contributes to hedgehog signaling pathway activation through sonic hedgehog induction in pancreatic cancerCancer Res200666147041704916849549
- ScharenbergCWHarkeyMATorok-StorbBThe ABCG2 transporter is an efficient Hoechst 33342 efflux pump and is preferentially expressed by immature human hematopoietic progenitorsBlood200299250751211781231
- KimMTurnquistHJacksonJThe multidrug resistance transporter ABCG2 (breast cancer resistance protein 1) effluxes Hoechst 33342 and is overexpressed in hematopoietic stem cellsClin Cancer Res200281222811801536
- GottesmanMMFojoTBatesSEMultidrug resistance in cancer: role of ATP-dependent transportersNat Rev Cancer200221485811902585
- SummerRKottonDNSunXMaBFitzsimmonsKFineASide population cells and Bcrp1 expression in lungAm J Physiol Lung Cell Mol Physiol20032851L97L10412626330
- AlviAJClaytonHJoshiCFunctional and molecular characterisation of mammary side population cellsBreast Cancer Res200351R1R812559051
- LechnerALeechCAAbrahamEJNolanALHabenerJFNestin-positive progenitor cells derived from adult human pancreatic islets of Langerhans contain side population (SP) cells defined by expression of the ABCG2 (BCRP1) ATP-binding cassette transporterBiochem Biophys Res Commun2002293267067412054520
- HussainSZStromSCKirbyMRSide population cells derived from adult human liver generate hepatocyte-like cells in vitroDig Dis Sci200550101755176316187169
- Hirschmann-JaxCFosterAEWulfGGA distinct “side population” of cells with high drug efflux capacity in human tumor cellsProc Natl Acad Sci U S A200410139142281423315381773
- MaddoxJShakyaASouthSTranscription factor oct1 is a somatic and cancer stem cell determinantPLoS Genet2012811e100304823144633
- RothSFoddeRQuiescent stem cells in intestinal homeostasis and cancerCell Commun Adhes2011183334421913875
- DembinskiJLKraussSCharacterization and functional analysis of a slow cycling stem cell-like subpopulation in pancreas adenocarcinomaClin Exp Metastasis200926761162319421880
- HoranPKMelnicoffMJJensenBDSlezakSEFluorescent cell labeling for in vivo and in vitro cell trackingMethods Cell Biol1990334694902084480
- RoeschAFukunaga-KalabisMSchmidtECA temporarily distinct subpopulation of slow-cycling melanoma cells is required for continuous tumor growthCell2010141458359420478252
- IshikawaFYoshidaSSaitoYChemotherapy-resistant human AML stem cells home to and engraft within the bone-marrow endosteal regionNat Biotechnol200725111315132117952057
- TengCGuoYZhangHDingMDengHIdentification and characterization of label-retaining cells in mouse pancreasDifferentiation200775870271217381540
- JohannessenTCBjerkvigRTysnesBBDNA repair and cancer stemlike cells – potential partners in glioma drug resistance?Cancer Treat Rev200834655856718501520
- LiuGYuanXZengZAnalysis of gene expression and chemore-sistance of CD133+ cancer stem cells in glioblastomaMol Cancer200656717140455
- JaeckleKAEyreHJTownsendJJCorrelation of tumor O6 methylguanine-DNA methyltransferase levels with survival of malignant astrocytoma patients treated with bis-chloroethylnitrosourea: a Southwest Oncology Group studyJ Clin Oncol19981610331033159779706
- EstellerMGarcia-FoncillasJAndionEInactivation of the DNA-repair gene MGMT and the clinical response of gliomas to alkylating agentsN Engl J Med2000343191350135411070098
- EylerCERichJNSurvival of the fttest: cancer stem cells in therapeutic resistance and angiogenesisJ Clin Oncol200826172839284518539962
- MagniMShammahSSchiróRMelladoWDalla-FaveraRGianniAMInduction of cyclophosphamide-resistance by aldehyde-dehydrogenase gene transferBlood1996873109711038562935
- GinestierCHurMHCharafe-JauffretEALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcomeCell Stem Cell20071555556718371393
- ChoKWangXNieSChenZGShinDMTherapeutic nanoparticles for drug delivery in cancerClin Cancer Res20081451310131618316549
- LiSDHuangLPharmacokinetics and biodistribution of nanoparticlesMol Pharm20085449650418611037
- ChenBSunQWangXReversal in multidrug resistance by magnetic nanoparticle of Fe3O4 loaded with adriamycin and tetrandrine in K562/A02 leukemic cellsInt J Nanomedicine20083227728618686787
- LiuCZhaoGLiuJNovel biodegradable lipid nano complex for siRNA delivery significantly improving the chemosensitivity of human colon cancer stem cells to paclitaxelJ Control Release2009140327728319699770
- GaucherGDufresneMHSantVPKangNMaysingerDLerouxJCBlock copolymer micelles: preparation, characterization and application in drug deliveryJ Control Release20051091–316918816289422
- Hyung ParkJKwonSLeeMSelf-assembled nanoparticles based on glycol chitosan bearing hydrophobic moieties as carriers for doxorubicin: in vivo biodistribution and anti-tumor activityBiomaterials200627111912616023198
- DharSReddyEMPrabhuneAPokharkarVShirasAPrasadBLCytotoxicity of sophorolipid-gellan gum-gold nanoparticle conjugates and their doxorubicin loaded derivatives towards human glioma and human glioma stem cell linesNanoscale20113257558021069248
- WangKLiuLZhangTOxaliplatin-incorporated micelles eliminate both cancer stem-like and bulk cell populations in colorectal cancerInt J Nanomedicine201163207321822238509
- ZhangYZhangHWangXWangJZhangXZhangQThe eradication of breast cancer and cancer stem cells using octreotide modified paclitaxel active targeting micelles and salinomycin passive targeting micellesBiomaterials201233267969122019123
- ZhouPHatziieremiaSElliottMAUptake of synthetic Low Density Lipoprotein by leukemic stem cells – a potential stem cell targeted drug delivery strategyJ Control Release2010148338038720869412
- WangKZhangTLiuLNovel micelle formulation of curcumin for enhancing antitumor activity and inhibiting colorectal cancer stem cellsInt J Nanomedicine201274487449722927762
- GrudzienPLoSAlbainKSInhibition of Notch signaling reduces the stem-like population of breast cancer cells and prevents mammosphere formationAnticancer Res201030103853386721036696
- CurtinJCLorenziM VDrug discovery approaches to target Wnt signaling in cancer stem cellsOncotarget20101756357721317452
- ZhouYYangJKopecekJSelective inhibitory effect of HPMA copolymer-cyclopamine conjugate on prostate cancer stem cellsBiomaterials20123361863187222138033
- MamaevaVRosenholmJMBate-EyaLTMesoporous silica nanoparticles as drug delivery systems for targeted inhibition of Notch signaling in cancerMol Ther20111981538154621629222
- LoWLChienYChiouGYNuclear localization signal-enhanced RNA interference of EZH2 and Oct4 in the eradication of head and neck squamous cell carcinoma-derived cancer stem cellsBiomaterials201233143693370922361100
- YangZJWechsler-ReyaRJHit ’em where they live: targeting the cancer stem cell nicheCancer Cell20071113517222787
- ZhuXZhouXLewisMTXiaLWongSCancer stem cell, niche and EGFR decide tumor development and treatment response: A bio-computational simulation studyJ Theor Biol2011269113814920969880
- WangCHChiouSHChouCPChenYCHuangYJPengCAPhotothermolysis of glioblastoma stem-like cells targeted by carbon nanotubes conjugated with CD133 monoclonal antibodyNanomedicine201171697920620237
- BurkeARSinghRNCarrollDLThe resistance of breast cancer stem cells to conventional hyperthermia and their sensitivity to nanoparticle-mediated photothermal therapyBiomaterials201233102961297022245557
- JosephITresslerRBassettEThe telomerase inhibitor imetelstat depletes cancer stem cells in breast and pancreatic cancer cell linesCancer Res1115201070229494950421062983
- GinsburgGSWillardHFGenomic and personalized medicine: foundations and applicationsTransl Res2009154627728719931193