
Abstract
Macrophages play a crucial role in tissue homeostasis and the innate immune system. They perform essential functions such as presenting antigens, regulating cytokines, and responding to inflammation. However, in diseases like cancer, cardiovascular disorders, and autoimmune conditions, macrophages undergo aberrant polarization, which disrupts tissue regulation and impairs their normal behavior. To address these challenges, there has been growing interest in developing customized targeted drug delivery systems specifically designed for macrophage-related functions in different anatomical locations. Nanomedicine, utilizing nanoscale drug systems, offers numerous advantages including improved stability, enhanced pharmacokinetics, controlled release kinetics, and precise temporal drug delivery. These advantages hold significant promise in achieving heightened therapeutic efficacy, specificity, and reduced side effects in drug delivery and treatment approaches. This review aims to explore the roles of macrophages in major diseases and present an overview of current strategies employed in targeted drug delivery to macrophages. Additionally, this article critically evaluates the design of macrophage-targeted delivery systems, highlighting limitations and discussing prospects in this rapidly evolving field. By assessing the strengths and weaknesses of existing approaches, we can identify areas for improvement and refinement in macrophage-targeted drug delivery.
Introduction
Macrophages, discovered by Metchnikoff, are crucial immune cells that maintain overall health by engulfing and digesting harmful microorganisms, dead cells, and foreign substances. They assist in tissue clearance and homeostasis.Citation1 Macrophages differentiate into tissue-resident macrophages (RTMs) or monocyte-derived macrophages based on the tissue environment and demand. RTMs establish long-term residence in tissues, while monocyte-derived macrophages are recruited to inflamed sites and exhibit functional differences.Citation2,Citation3 Macrophages have diverse functions, including tissue homeostasis, immune response induction and resolution, and tissue development and repair.Citation4–6 They can be activated into distinct cellular states in response to stimuli from different tissues or cues.Citation7 Typically, macrophages can differentiate into classically activated M1 macrophages or alternatively activated M2 macrophages. M1-type macrophages kill pathogens, while M2-type macrophages contribute to tissue remodeling and healing. Dysregulated expression of M1 or M2 is implicated in diseases such as tumors and rheumatoid arthritis.Citation8,Citation9
Targeting macrophages using drug delivery systems holds promise for regulating disease progression and improving therapeutic efficacy. These targeted delivery systems enable high selectivity in delivering nanomedicines, reducing distribution in healthy tissues, and minimizing toxicity.Citation10 Various nanomaterials have been employed to target macrophages through different routes.Citation11 The intense phagocytic activity of macrophages leads to the passive targeting of conventional nanoscale carriers due to their surface materials or serum conditioning-mediated effects.Citation12 The liver, which contains a significant number of macrophages as part of the reticuloendothelial system (RES), often accumulates a substantial proportion of nanoscale carriers, which are subsequently recognized and metabolized by macrophages.Citation13 However, if targeted macrophages are not resident in the liver, the carriers must cross the RES to reach the lesion site and bind with specific macrophages.Citation14–18 For example, clodronate, a specific macrophage-toxic drug, has been encapsulated in liposomes and shown promise in modulating various diseases, including cancer, cardiovascular diseases, and autoimmune diseases.Citation19 Although the concept of targeting macrophages with personalized drug delivery systems for immune-related diseases is still in its early stages, it holds great long-term potential.Citation20
This article summarizes active and passive targeting techniques for interaction between different carriers and macrophages, elucidating the role of macrophages in specific diseases. Additionally, the article examines the obstacles encountered by macrophage-targeted nanomedicine, including issues related to poor selectivity. Furthermore, it explores the potential prospects of nanocarriers in modulating macrophage activity.
Influence of Physiological Environment on Macrophage-Nanomedicine Interactions
Proteins, Biological Molecules
Proteins and other biomolecules play an important role in regulating the interactions between nanomaterials and macrophages. Plasma is the most important biological medium, in which the total plasma protein concentration is about 60–80 g/L.Citation21 When the nanomedicine enters into the blood vessel for about 30s, it adsorbs a large amount of proteins on its surface and forms a biocrown.Citation22 The composition of the biocrown significantly affects the in vivo biological properties of the carrier. First, the binding mode of nanodrugs to plasma proteins is a key factor affecting their recognition by macrophages. It has been shown that adsorption of non-modulin proteins such as serum albumin and apolipoprotein J (Apo J) reduces the carrier-cell membrane interaction and macrophage uptake, and incubation with serum albumin significantly inhibits uptake by dendritic cells.Citation23 On the contrary, when more conditioning agents, such as complement and immunoglobulin, were adsorbed on the carrier surface, phagocytes could rapidly recognize the nanomedicine, which in turn accelerated its clearance. Secondly, the adsorbed protein crowns can significantly affect the targeting of nanocarriers such as liposomes, eg, Guan et al showed that liposomes modified with cation-rich peptides adsorbed higher amounts of IgG and IgM, which inhibited the targeting of the formulations and enhanced the interaction with immune cells.Citation24 Therefore, reducing the content of protein crowns or adjusting the adsorbed species can effectively regulate the interaction between carriers and macrophages. One approach is to modify nanomedicine with polyethylene glycol (PEG).Citation25–27 PEG, as a hydrophilic long-chain polymer, effectively reduces protein adsorption on the surface of nanocarriers.Citation28 Increasing the density of PEG on gold nanoparticles was shown to significantly reduce serum protein adsorption and uptake by macrophages.Citation29 However, Schottler et al discovered that the adsorption of a specific quantity of serum proteins could actually enhance the carrier’s ability to inhibit macrophage uptake.Citation30 This finding introduces complexity and raises questions about how protein corona formation influences long-circulation properties. In addition to PEG modification, biomimetic modifications have gained attention. Incorporating cell membranes or exosomes into liposomes during preparation allows retention of self-recognition properties while achieving targeted delivery and high drug loading capacity.Citation31–33 CD47, which interacts with SIRPα proteins on macrophages, is considered a basis for achieving biocompatibility and efficient drug delivery in biomimetic systems. Interestingly, our preliminary validation confirms that this modification strategy does not decrease accessibility for cell recognition and internalization, and thus, there was no significant reduction in the distribution of carriers in the liver or on cell membrane surfaces.Citation34
In conclusion, proteins and other biomolecules play a key regulatory role in modulating the interactions between nanomaterials and macrophages. They can influence the interaction of nanomaterials with macrophages by way of adsorption, immunomodulation and biocompatibility, which can have a significant impact on biological effects.
Physiological Fluids
Physiological fluids play an important influence in regulating the interaction of nanomaterials with macrophages. First, the presence of physiological fluids changes the way nanomaterials move in biological systems. Nanomaterials exhibit different behaviors in fluids due to the mechanics of fluid motion, such as shear and turbulence. In blood, different tissues and organs in the body often have their own unique physiological microenvironments, allowing nanocarriers to have different motions. For example, in the hepatic sinusoids of the liver, the flow rate of the nanocarriers is reduced by ~1000 times relative to the blood vessels outside the liver, allowing them to passively aggregate in the liver, thus enhancing the interaction with macrophages in the liver.Citation35 However, as the particle size of the carriers increases to the micrometer scale, they can significantly break through the limitations of the hepatic endothelial system to achieve aggregation in the lungs.Citation36 Lymph is a colorless, transparent fluid composed of plasma, lymphocytes, and other solutes. Its circulation rate is significantly lower than in the blood. As a result, the interaction between nanomedicines and macrophages in lymph is significantly enhanced. For example, based on the role of lymph in immune defense, Wang et al constructed a nano vaccine targeting lymphatic sinus-associated SIGNR1+ macrophages, which could effectively present antigens to B cells after interaction with macrophages and promote the killing of chronic hepatitis B.Citation37
Second, physiological fluids can modulate macrophage response to nanomaterials. Through hydrodynamic stimulation, physiological fluids can affect macrophage morphology, degree of activation, and functional expression. One of the key effects is that mechanical stimulation under physiological flow conditions induces macrophages to secrete specific cytokines that modulate their interaction with nanomaterials. The work of Hyojae Son et al showed that shear stress induces monocyte/macrophage to undergo M1-type polarization and increase secretion of pro-inflammatory factors such as CCL2, TNF-α, and IL-1β. and IL-1β.Citation38 Yazdimamaghani et al produced four types of silica nanoparticles (NPs) with different densities and surface charges and tested the cytotoxicity and uptake on RAW 264.7 macrophage cells after 24 hours of incubation with the cells, in static or under flow conditions. They found that the cell viability is enhanced under flow conditions, compared to static culture. Authors also found that, cellular uptake of silica NPs was more in static conditions compared to dynamic conditions. Furthermore, low-density particles, showed lower uptake under flow conditions compared to high-density particles.Citation39
Local Pathology on Nanomedicine-Macrophages Interactions
When exposed to various microenvironmental stimuli, macrophages are highly plastic and differentiate predominantly into pro-inflammatory M1 and anti-inflammatory M2 types, both of which almost always have completely opposite functions. Due to this feature, macrophages perform different functions at different stages of immunity and inflammation.
The inflammatory state is a complex physiological response that is a nonspecific defense and repair process of the body against injury, infection, or other stimuli.Citation40 Undue chemokines, cytokines, and proinflammatory mediators from macrophages are closely associated with many human diseases, such as pneumonia and rheumatoid arthritis.Citation41 In an inflammatory environment, macrophages often present as pro-inflammatory M1-type macrophages. They constitute the first line of defense against intracellular pathogens and promote Th1 polarization of CD4 cells, in addition to releasing inflammatory mediators such as cytokines, chemokines, and oxygen free radicals, affecting the surface properties and stability of nanomaterials and promoting their interaction with macrophages.Citation42–45
M2-like polarized populations are particularly important during parasitic, helminthic, and fungal infections, where they are induced in response to a Th2 response. Macrophage colony-stimulating factor (M-CSF), IL-4, IL-10, IL-13, or a combination of these factors are able to polarize macrophages toward an M2-like phenotype.Citation46,Citation47 M2-like macrophages are identified primarily based on the expression of CD64 and CD209 (a C-type lectin).Citation45 In contrast to classically activated M1-like macrophages, M2-like macrophages have negatively regulated pro-inflammatory cytokines and induce the production of anti-inflammatory mediators such as IL-4, IL-10, and TGF-β.Citation48,Citation49 Indeed, they are highly endocytosed and partially phagocytosed and are involved in a wide range of functions, including repair mechanisms, homeostasis, metabolic processes, and pathogenesis.Citation50
Toxicity of Nanomaterials
The toxicity of macrophage-targeted nanoparticles (NPs) is an important consideration in their clinical application. Macrophages play a crucial role in the immune response and are responsible for the phagocytosis of foreign substances, including nanoparticles. However, the interaction of nanoparticles with macrophages may lead to potential toxicity issues. First, when macrophages phagocytose nanoparticles, they may trigger cellular stress and inflammatory responses, which in turn produce reactive oxygen radicals (ROS) and pro-inflammatory cytokines. Excessive ROS production may lead to oxidative stress and damage cellular structures, while high levels of pro-inflammatory cytokines may lead to tissue damage and immune dysregulation. For example, Aaron Lee et al demonstrated that polylysine modification increased TNF-α secretion.Citation51
In addition, macrophages are important regulators of the immune system. The interaction of nanoparticles with macrophages may modulate their normal immune functions, such as antigen presentation, cytokine production, and phagocytic activity. Depending on the nature of the nanoparticles, this altered immune response may have systemic effects and influence overall immune homeostasis. For example, inhalation of carbon nanotubes can suppress the humoral immune response through the production of transforming growth factor β (TGF-β) by alveolar macrophages and the subsequent production of prostaglandins by splenocytes, leading to systemic immunosuppression.Citation52
Targeting Strategy
Nanomedicine targeted delivery involves precise drug transportation to specific body sites by manipulating nanoscale drug carriers. It allows drugs to bypass tissue barriers and accumulate selectively in diseased tissues or organs. Targeted delivery operates on two main mechanisms: the affinity between target receptors and drug carriers for precise drug localization, and strategies like surface modification to enhance targeting delivery. This approach enhances drug efficacy, reduces side effects, and improves patient quality of life. As a result, targeted delivery has emerged as a major focus of research in the field of drug delivery. In this context, we aim to explore the interaction between nanocarriers and macrophages from both passive and active targeting perspectives.
Passive Targeting
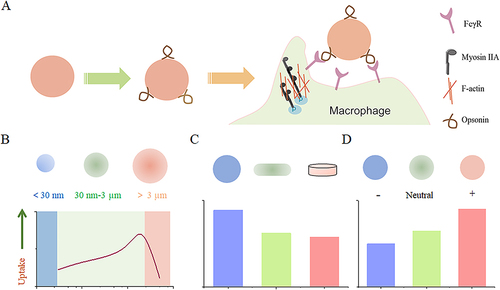
Passive targeting is one of the important strategies for nanodrug-targeted therapy. It is a non-specific targeting approach that does not require specific targeting molecules. Instead, Passive targeting relies on the preferential accumulation of NPs modified with specific physical properties (such as size, shape, and charge), which significantly impact the rate and range of macrophage uptake.Citation53 Therefore, we will provide an overview of these specific conditions ().
Figure 1 The impact of nanoparticle size, shape, and surface charge on interactions with macrophages. (A) Nanoparticle carriers quickly adsorb complement components from the plasma upon entering the vascular system, enhancing their recognition by macrophages. (B) Spherical particles of different sizes affect their interaction with macrophages. Particle sizes ranging from 30 to 3000 nm have been shown to increase macrophage recognition. Conversely, smaller nanoparticles below 30 nm or larger nanoparticles above 3 μm tend to reduce their interaction with macrophages. (C) Distinct nanoparticle shapes exhibit unique properties that significantly affect interaction with cell membranes, and uptake by macrophages. (D) The charge carried by nanoparticles, resulting from various surface chemistries, also affects their interaction with macrophages. Positively charged particles have a greater tendency to be captured by macrophages.
Particle Size
The shape of nano-carriers plays a significant role in macrophage uptake, influencing the mechanism and rate of macrophage engulfment. Generally, particles with sizes ranging from 30 nm to 3 µm can be specifically recognized and phagocytosed by macrophages, while outside this range, the phagocytic activity towards the particles decreases.Citation54–56 Moreover, within a certain range, macrophages seem to preferentially internalize larger-sized carriers. For instance, a previous study demonstrated that in cultured primary peritoneal macrophages, they exhibited higher uptake of approximately 500 nm polymer nanoparticles compared to those sized 150 or 300 nm.Citation57 In other studies, J774A.1 macrophages showed the highest uptake of nanoparticles with a diameter of 1.9 μm compared to sizes of 430 nm and 4.8 μm.Citation58
Macrophages are capable of internalizing and engulfing particles of various sizes through different mechanisms. For larger particles, their micrometer scale allows for active binding and phagocytosis, while smaller particles can be taken up by macrophages through various mechanisms. Therefore, the particle size of nanocarriers not only affects their uptake efficiency but also determines the internalization mechanisms of nanoparticles. Experimental studies have compared the absorption mechanisms of particles by macrophages. Kuhn et al conducted cellular experiments using different-sized polystyrene particles in macrophages (J774A.1).Citation59 In J774A.1 cells, 1 μm particles were internalized via phagocytosis/macropinocytosis, while 40 nm polystyrene particles were taken up through clathrin-mediated or caveolin-mediated endocytosis as well as phagocytosis. Furthermore, Yue et al demonstrated that macrophages solely employ the lysosomal pathway when engulfing nanosized carriers.Citation58 These findings underscore the significance of considering particle size in comprehending the mechanisms of nanoparticle uptake by macrophages.
Particle Shape
The shape of nanocarriers is a factor that is easily overlooked, as conventional nanocarriers such as liposomes, polymer micelles, and polymer nanoparticles are spherical in shape. However, emerging nanocarrier preparation technologies, such as lipid nanodiscs and membrane-encapsulated carriers, offer the possibility to modulate carrier morphology and optimize carrier performance.
In general, spherical particles are more readily phagocytosed by macrophages compared to elliptical, rod-shaped, or cylindrical particles. When the curvature of a particle exceeds 45 degrees, it cannot be completely internalized by cells.Citation60,Citation61 Some studies have found that worm-like elongated nanoparticles or ellipsoids can reduce or inhibit the rate of phagocytosis, allowing them to evade immune responses.Citation62,Citation63 For instance, Champion et al conducted studies on the phagocytic mechanisms using different shapes of polystyrene nanoparticles, including spheres, ellipsoids, and oval or rectangular disks.Citation64 The results showed that, the curvature of the particles plays a crucial role in the internalization process, independent of their size. Furthermore, other studies indicate that a high aspect ratio leads to reduced internalization. Different morphologies of carriers may have different hemodynamic parameters, and their behavior in the vasculature can influence their interactions with macrophages.Citation35 Indirectly, the interaction between carriers and organ macrophages in the hemodynamic environment of the liver is also affected. Additionally, highly curved regions such as carbon nanotubes or graphene edges have penetrating abilities and can puncture the cell membrane to enter the cell. These differences in uptake mechanisms enable macrophages to selectively internalize particles of different shapes.Citation65,Citation66
Particle Stiffness
Stiffness can affect the mechanical properties and rigidity of nanoparticles, thereby influencing their interactions with cells.Citation67 For example, Aaron Lee et al evaluated the uptake of poly(N-isopropylacrylamide) with different degrees of crosslinking by primary human monocyte-derived macrophages (MDM) and observed a preference of MDM for removing highly crosslinked (stiffer) particles over less crosslinked (softer) particles.Citation51 Furthermore, there are variations in the response of different shapes of carriers to stiffness. In reducing macrophage internalization of spherical particles, increasing size was more effective than reducing the stiffness of immunoglobulin G-functionalized poly(acrylic acid) (PAA) particles. On the other hand, the internalization of rod-shaped but non-spherical PAA particles critically depended on stiffness.Citation64 In addition, Palomba et al demonstrate that compared to rigid nanostructures, softer nanostructures achieved up to five-fold higher efficiency in evading cellular uptake by bone marrow-derived monocytes. Soft circular and square nanostructures were similarly absorbed by professional phagocytic cells (<15%); however, soft elliptical particles were more easily internalized (<60%), possibly due to their larger size and elongated shape, while any shape and size of rigid nanostructures were taken up by over 70% of macrophages.Citation68
Particle Charge
The charge associated with nanoparticles, which arises from diverse surface chemistries, influences opsonization, circulation times, and interactions with macrophages in organs. Typically, cationic nanoparticles are more easily taken up by phagocytes compared to neutral or negatively charged nanoparticles. The positive charge on nanoparticles can interact with the negatively charged phospholipid compositions of cell membranes, leading to increased nonspecific interactions with biological cells and circulating proteins. However, it is crucial to acknowledge that the positive charge of nanoparticles can also induce adverse biological reactions including enhanced complement system activation and cytotoxicity. Therefore, careful consideration must be given to balancing the benefits of improved cellular uptake against potential harmful effects when utilizing cationic nanoparticles for macrophage targeting.Citation69–71 The role of charge in macrophage uptake remains a topic of debate. For instance, Oleg Lunov et al conducted an analysis involving THP-1 cells and macrophages differentiated from human monocytes using nanoparticles with different surface charges: PS-COOH (negative charge) and PS-NH2 (positive charge). After incubation with these nanoparticles in a buffer for 3 to 6 hours, both types of nanoparticles were taken up by the cells. However, THP-1 cells absorbed approximately 38% more PS-COOH nanoparticles than macrophages did, whereas both cell types displayed similar levels of cellular internalization with PS-NH2 nanoparticles. Interestingly, when the analysis was conducted in HBSS solution, both cell types exhibited lower uptake of PS-NH2 nanoparticles compared to PS-COOH nanoparticles.Citation72 Additionally, in the case of RAW 264.7 cells, a specific type of consuming cells, the intake level of negatively charged AuNPs is similar to that of positively charged AuNPs.Citation73
Active Target
Active targeted therapy relies on specific molecular recognition to deliver substances to specific tissues and cells while minimizing accumulation in healthy tissues. In the case of targeting macrophages, challenges arise due to varying receptor expression among different macrophage phenotypes. For example, in tumor-associated macrophages (TAMs), it is essential to target M2-type TAMs specifically. However, distinguishing receptor expression across macrophages throughout the body can be difficult. Research has identified highly expressed receptors in macrophage populations, providing insights into ligands used for macrophage targeting and the receptors involved ( and ). These findings contribute to understanding the strength and specificity of receptor-ligand interactions.
Figure 2 Graphical representation of surface proteins utilized for macrophage targeting.
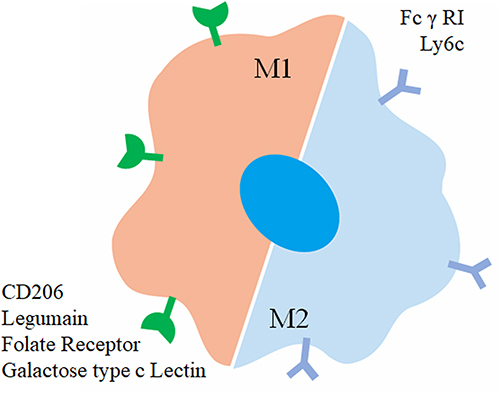
Table 1 Affinity Analysis Between Receptors and Ligands
Antibody Antigen Interaction
Antibodies are highly specific and diverse protein molecules produced by the immune system. They play a crucial role in targeted therapy for macrophages in two ways: 1) Regulating macrophage signaling pathways: Antibodies selectively bind to receptors or antigens on macrophages, modulating their functions including inflammation regulation, immune responses, phagocytosis, and cytokine production. 2) Serving as targeting ligands: Antibodies recognize and bind to specific antigens on macrophages, enhancing drug delivery specificity and allowing efficient and targeted delivery of therapeutic agents. However, challenges such as systemic toxicity and limited specificity exist with current antibody-based targeting strategies.
CSF-1 is a signaling protein that binds to its receptor CSF-1R and plays a crucial role in regulating macrophages.Citation85 Inhibiting CSF-1R has shown promise in preclinical cancer models but can cause systemic toxicity.Citation86–88 The CD47/ signal regulatory proteins-α (SIRP α) axis is another therapeutic target where blocking CD47 using anti-CD47 antibodies enhances macrophage function against cancer cells.Citation89–92 Antibodies targeting macrophage surface proteins facilitate targeted delivery of nanoparticles, but their efficacy can be hindered by overlapping expression and non-specific uptake by macrophages.Citation93–95 Nanobodies, smaller fragments without the Fc domain, have been developed to overcome limitations but still face challenges.Citation96,Citation97 Considering these limitations, research is focused on finding alternative ligands that are more efficient and specific in enhancing drug delivery to macrophages.
Peptide Ligands
Peptide ligands offer specific recognition of their corresponding receptors and come with several advantages over antibodies, including smaller size, lower immunogenicity, and reduced manufacturing costs. These ligands often target macrophage surface receptors that are highly expressed. Cieslewicz et al conducted a study where they discovered a unique peptide sequence called M2pep. This peptide demonstrated a preference for binding to M2 macrophages rather than M1/M0 macrophages or other white blood cells.Citation98 In a subsequent study, Conde et al conjugated M2pep to gold nanoparticles to achieve targeted delivery of small interfering RNA (siRNA) for the inhibition of VEGF expression. The researchers demonstrated the high selectivity of this delivery system towards M2 macrophages in lung tissue and lavage fluid.Citation99 Qian et al also utilized M2pep as a targeting ligand for TAMs by delivering siRNA against anti-CSF-1R. This approach resulted in anti-tumor activity in mouse tumor models.Citation100 Furthermore, Ngambenjawong et al synthesized bivalent and tetravalent display bodies ([M2pep]2-Biotin and [M2pep]4-Biotin) based on M2pep. Compared to the monovalent form, [M2pep]2-Biotin showed higher affinity and selective binding to M2 macrophages. However, [M2pep]4-Biotin decreases the binding activity and selectivity. Interestingly, both the bivalent and tetravalent forms of M2pep displayed selective toxicity towards M2 macrophages without any cytotoxic drug loading, while this effect was not observed in monovalent M2pep treatment.Citation101
In the case of CD206, also known as the Mannose Receptor (MMR), the peptide UNO has shown selective binding to this receptor and has been used for targeted drug delivery. The UNO-mediated delivery system has exhibited significant therapeutic efficacy in various tumor models such as breast cancer, melanoma, glioblastoma, and gastric cancer.Citation75 This system outperforms other CD206-targeting peptides or mannose-like compounds by avoiding binding to additional receptors like SIRPα or CD209. Another peptide, CRV, which interacts with the extracellular retinoid X receptor β (RXRB) found on CD11b+F4/80+ macrophages, specifically binds to macrophages and can rapidly extravasate into tumors.Citation83 Nevertheless, peptide delivery systems face challenges such as reduced binding affinity compared to antibodies and increased susceptibility to protein degradation.Citation102
Carbohydrate Ligands
Carbohydrate-targeting ligands indeed offer advantages such as high specificity, high affinity, high water solubility, and low cost. The macrophage mannose receptor (MMR/CD206) is prominently expressed on M2 macrophages, making it an attractive target for specific delivery. Modifying the surface of carriers with mannose can effectively facilitate recognition by M2 macrophages, leading to significant colocalization of mannose-modified carriers with M2 macrophages.Citation103,Citation104 Besides monosaccharides, polysaccharides related to mannose can also exhibit similar recognition effects. These longer chain polysaccharides can effectively reduce plasma protein-mediated inhibition of carrier targeting. An example is Beta-glucan from Grifola frondosa (BSP) and its conjugates, which have been shown to reduce tumor angiogenesis through their effects on macrophages, exhibiting inhibition of sarcoma development.Citation78 In addition, we have reported the targeting of galactose-type C-type lectins, which are highly expressed receptors on M2 macrophages. By utilizing galactose-modified liposomes, the recognition of M2 macrophages can be enhanced.Citation105
However, it’s important to note that carbohydrates can be recognized by various lectins, and their antibody counterparts exhibit high specificity towards their homologous receptors. For instance, the mannose fraction can be recognized by other mannose-binding receptors, including DC-SIGN, L-SIGN, Endo180, or mannose-binding lectin.Citation106 While carbohydrate ligands offer advantages such as high solubility and low cost, they may lack the specificity exhibited by other ligands toward their target receptors.
Nucleic Acid Aptamers
Nucleic acid aptamers, including both RNA and DNA oligonucleotides, exhibit unique secondary structures that enable them to interact with target molecules effectively. These aptamers possess distinct advantages such as simple structure, easy synthesis, and good chemical and biological stability.Citation107,Citation108 Sylvestre et al, identified a specific aptamer selectively bound to CD14+ monocytes rather than CD16+ monocytes and was rapidly internalized by these cells.Citation109 Additionally, Qian et al constructed a DNA tetrahedral framework nucleic acid (tFNA) nano-delivery system to convert TAMs from an M2 phenotype to an M1 phenotype within the tumor microenvironment, thereby exerting significant anti-tumor effects.Citation110 Moreover, they also proposed an aptamer-engineered M1 macrophage (ApEn-M1) which demonstrated outstanding anti-tumor efficacy.Citation111
Interactions Between Different Carriers and Macrophages
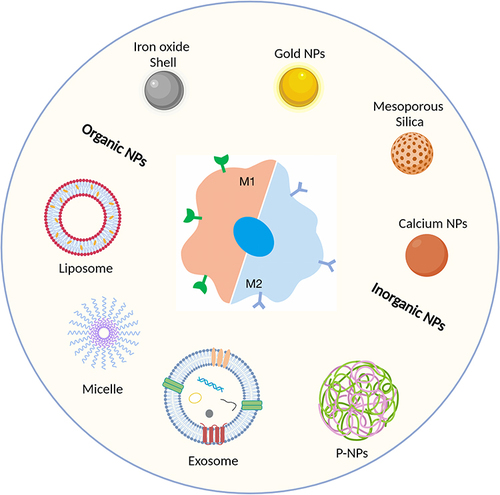
In this context, we delve into the complex interplay between carriers and macrophages, with a primary focus on the diverse types of carriers employed in drug delivery research ().
Figure 3 Overview of commonly used carrier types: classified as organic or inorganic nanoparticles. Created with BioRender.com.
Liposomes
Liposomes are artificial vesicles that mimic the structure of cell membranes and can effectively deliver drugs.Citation25,Citation112,Citation113 The interaction between liposomes and macrophages is mainly related to the nature of their carrier materials. Tang et al compared the effects of liposome size, charge, and lipid composition on their interaction with macrophages, and their results showed that compared with 100-nm neutral 1,2-dipalmitoyl-sn-glucose-3-phosphatidylcholine (DPPC) liposomes, macrophages internalized the cationic liposomes and 1.2-di olive acyl-sn-glucose-3-phosphatidylcholine (DSPC) liposomes to a greater extent, and the primary mechanisms were phagocytosis and lattice protein-mediated endocytosis.Citation114 In addition, phosphatidylserine (PS) is a common major cell membrane component and acts as a “phagocytic signal” for macrophages. Therefore, phosphatidylserine-constructed liposomes can mimic this exposed phosphatidylserine and trigger phagocytosis by macrophages.Citation115 However, Riki Toita et al showed that Phosphatidylserine liposome has the potential to promote the polarization of M1-type macrophages to M2-type macrophages in bone regeneration and repair.Citation116
Exosomes
Cellular communication primarily occurs through chemical messengers, among which extracellular vesicles (EVs) are a common form. Exosomes are the smallest type of EVs, with diameters ranging from approximately 30 to 150 nm, and they play important roles in intercellular interactions.Citation117 Based on the protein composition anchored on the surface of exosomes, they can achieve various biological functions. Firstly, exosomes can achieve long-circulating delivery through the expression of immune recognition proteins such as CD47 on their surfaces.Citation118 Furthermore, although both liposomes and exosomes consist of phospholipid bilayers, liposomes are often composed of synthetic materials, which have been found to have a potential risk of inducing hypersensitivity reactions.Citation119 In addition, exosomes derived from endogenous cells can maintain the drug-loading capabilities of liposomes while achieving targeted delivery, thus improving the efficiency of targeted drug delivery.
In targeted delivery to macrophages, exosomes can play two main roles: 1) direct targeting of macrophages, and 2) targeting the communication processes between macrophages and other cells (eg, tumor cells). Engineered therapeutic exosomes developed by researchers can provide targeted control of macrophage phenotypes through transcription factor ASO (exoASO-STAT6), effectively reprogramming tumor-associated macrophages (TAMs) into an M1 phenotype.Citation120,Citation121 Another approach involves bacterial outer membrane vesicles (OMVs), which are highly immunogenic nanovesicles that can stimulate macrophages. To create a “tunable bidirectional adapter” based on OMVs, Feng et al fused CD47 nanobody onto the surface of OMVs and encapsulated them with polyethylene glycol (PEG) containing diselenide bonds (PEG/Se). They named this construct PEG/Se @ OMV-CD47nb. Through various stimulation pathways, this adapter activates tumor-associated macrophages by inducing M1 polarization or blocking the “do not eat me” signal. Consequently, TAMs are stimulated to engulf tumor cells.Citation122
Micelles
Micelles prepared from amphiphilic block copolymers have become a research hotspot in drug delivery due to their unique core-shell structure. In recent years, there have been numerous studies utilizing hydrophilic polymers as shell materials for micelles. Among these, polyethylene glycol (PEG) has emerged as the most commonly used hydrophilic polymer due to its excellent biocompatibility, stealth behavior associated with its molecular structure, and ability to bind a large amount of water through hydrogen bonding.Citation123,Citation124
Similar to other nanoparticles, the micelles themselves interact with macrophages mainly originating from the carrier potential. Faezeh Sabzehei et al showed that conjugated branched PEI-2k and oleic acid (OL) as the building blocks of the cationic micelle significantly increased uptake in RAW264.7 cells.Citation125 Hou et al evaluated the effect of PCL Chain Length on the interaction of Poly(ε-Caprolactone)-Methoxypolyethylene Glycol (PCL-MPEG)-based micelles with macrophages, confirming that compared to PCL-MPEG, the uptake of the cationic micelle was significantly increased. The effect of chain length on the interaction between PCL-MPEG-based micelles and macrophages confirmed that protein adsorption was affected and dependent on chain conformation compared to PCL40-MPEG and PCL20-MPEG, with the lowest adsorption and the least uptake in RAW264.7 cells.Citation126 Similar to liposomes, the incorporation of phosphatidylserine into the carrier material can contribute to enhanced macrophage recognition.Citation127
In addition, the micellar can be simultaneously modified with the corresponding ligand to improve targeting efficiency. Sun et al developed hemoglobin-conjugated doxorubicin-loaded micelles (Hb-DOXM) that could bind with endogenous plasma thrombin and achieve specific targeting of M2 TAMs.Citation128 Liu et al developed zinc protoporphyrin IX (ZnPP)-grafted poly(l-lysine)-b-poly(ethylene glycol) peptide micelles (ZnPP-PM) for galactose-functionalized targeting of macrophages.Citation129
Polymer Nanoparticles
Polymer nanoparticles (P-NPs) are nanoscale particles that can encapsulate or adsorb active compounds. They offer advantages such as easy manufacturing, surface functionalization, biodegradability, and biocompatibility.Citation130–132 Moreover, P-NPs have shown promise as carriers for photodynamic agents, photothermal agents, and chemotherapeutic drugs.Citation133–135
Some materials can modulate macrophage polarization, and their effects depend on various modifications and residues. Carboxyl-modified and amino-modified polystyrene nanoparticles could inhibit M2 macrophage polarization by downregulating CD200R, CD163, and IL-10 expression without affecting M1 markers.Citation136 Similarly, hydrophilic polyurethane nanoparticles could suppress M1 macrophage polarization by reducing inflammatory mediators like TNF-α and IL-1β.Citation137
Inorganic Nanoparticles
As mentioned previously, some nanoparticles have the ability to reprogram macrophages, and one notable example is iron oxide nanoparticles. In a study by Saeid Zanganeh et al, it was discovered that clinically approved iron oxide nanoparticles, specifically ferumoxytol, could polarize macrophages towards the M1 phenotype, leading to the inhibition of tumor growth.Citation138 Interestingly, subsequent research revealed that both feracarbotran and ferumoxytol, two clinically approved iron oxide nanoparticles, could induce autophagy in macrophages and trigger inflammation through TLR4-mediated signaling and oxidative stress.Citation139 Additionally, other iron oxide nanoparticles have been reported to effectively convert macrophages into anti-tumor phenotypes.Citation140–145 Furthermore, studies have indicated that iron-related nanoparticles and calcium ions, could promote the production of pro-inflammatory cytokines such as IL-1β.Citation146 Consequently, the use of calcium-based nanoparticle drugs may have a positive impact on M1 repolarization. For instance, He et al successfully increased the expression of several M1 markers in J774A.1 macrophages by delivering plasmid DNA using peptide-modified calcium carbonate nanoparticle.Citation147 Nanomedicines have demonstrated the ability to induce repolarization of bone marrow-derived macrophages, leading to an increased expression of M1 markers and a decreased expression of M2 markers.Citation148 Additionally, another study achieved similar effects in macrophage repolarization by coating hyaluronic acid (HA) onto manganese dioxide nanoparticles.Citation103 Furthermore, black phosphorus nanoparticles and mesoporous Prussian blue have been utilized to polarize M2 macrophages through the loading of low molecular weight HA.Citation149
The Regulatory Role of Macroscopic Targeted Nanomedicines in Different Diseases
Macrophages have a pivotal role in maintaining physiological balance within the body, and any dysregulation or impairment of their functions can contribute to the development of systemic diseases.Citation5,Citation150 For instance, in non-alcoholic fatty liver disease, M1-polarized macrophages create an inflammatory microenvironment by releasing cytokines like TGF-β, thereby promoting hepatocyte steatosis and liver fibrosis. Conversely, in tumors, M2-polarized tumor-associated macrophages are known to support tumor growth by participating in angiogenesis and aiding in tumor cell metastasis. Therefore, Effective therapeutic interventions can be devised by understanding the state of macrophages in diverse disease contexts. Additionally, we also summarize some of the nanomedicines that have entered clinical trials from the relevant literature and the US National Library of MedicineCitation151 ().
Table 2 Examples of Clinical Trials on Macrophage-Targeted Nanomedicine for Disease Treatment and Diagnosis
Tumors
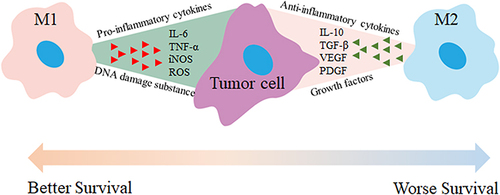
Tumors are a severe disease, causing harmful effects on organs, mortality, and societal implications.Citation152 Tumor-associated macrophages (TAMs) play a critical role in cancer progression.Citation153,Citation154 They are classified into M1 and M2 subgroups. M1 TAMs promote an inflammatory response and exhibit anti-tumor effects during early stages, while M2 TAMs possess anti-inflammatory properties that contribute to tumor growth and invasion. Targeting M2 TAMs can offer therapeutic benefits (). Therefore, targeting M2 TAMs offers therapeutic benefits and can be achieved through strategies such as inhibiting recruitment, eliminating or depleting TAMs, and reprogramming them into an M1 phenotype.Citation153,Citation155,Citation156
Figure 4 M1 and M2 macrophages have distinct roles in tumor therapy. M1 macrophages secrete pro-inflammatory cytokines like TNF-α, IL-6, and IL-12, which activate immune responses, enhance anti-tumor effects, and promote tumor cell apoptosis. In contrast, M2 macrophages release anti-inflammatory or immunosuppressive cytokines such as TGF-β, IL-10, IL-13, and IL-4, which create an immunosuppressive environment, inhibit anti-tumor responses, and contribute to angiogenesis and tissue remodeling. Therefore, activating M1 macrophages and inhibiting M2 macrophage activation can reduce tumor growth, metastasis, and angiogenesis.
Monocytes/macrophages play diverse roles in immune regulation, angiogenesis, tumor metastasis, and invasion.Citation153 Macrophages in different tumor regions have distinct origins. Classical monocytes are the main source of macrophages in tumor epithelial, perivascular, and hypoxic areas, while non-classical monocytes contribute predominantly to macrophages in perivascular regions of tumors.Citation157 Recruitment of monocytes/macrophages by tumors and other tissues is facilitated by cytokines and chemokines such as CCL2, CCL5, VEGF, CSF-1.Citation153 Modulating monocytes through a lipid nanoparticle platform that silences CCR2 with siRNA has shown therapeutic effects in mouse cancer models.Citation158 Macrophage development relies on the interaction between macrophage colony-stimulating factor (M-CSF) and its receptor M-CSFR or CD115, with recruitment dependent on the CCL2-CCR2 axis.Citation159 Drugs like PF-04136309, a CCR2 antagonist, could block the migration of CCR2+ monocytes from bone marrow to tumors, resulting in reduced numbers of M2 TAMs.Citation160
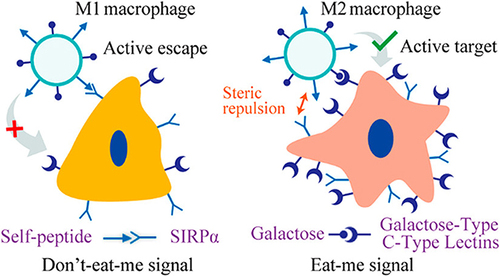
Targeted clearance of macrophages is another major modality, and modification of the surface of the carrier with ligands specifically expressed on the surface of M2-type macrophages is effective in increasing the selectivity of nanomedicines.Citation161 However, carriers targeting M2-type macrophages also have high selectivity for M1-type macrophages. To address this issue, we propose to combine “eat me” and “don’t eat me” signaling to improve selectivity for M2 TAMs. We have confirmed that “do not eat me” signaling is more inclined to suppress the uptake of M1 macrophages compared to M2 macrophages. Therefore, surface modification of macrophages with CD47 and the utilization of macrophage galactose c-type lectin as targeting ligands have shown promise in achieving precise selection of M2 macrophages ().Citation42,Citation105,Citation162 In addition, while the non-specific uptake of nanoparticles by macrophages has traditionally been considered a ‘drawback’ in circulation. Macrophages could actively migrate towards and engulf nanoparticles extravasating from blood vessels, contributing to their accumulation within the tumor stroma.Citation163
Figure 5 Enhancing precise recognition of M2 macrophages through combined Eat Me and Do not Eat Me signals. The interaction between the CD47 protein-derived Self peptide and the SIRPα protein on macrophage surfaces activates the Do not Eat Me signaling pathway, resulting in the inhibition of macrophage phagocytosis. Furthermore, the Do not Eat Me signal exhibits a tendency to suppress phagocytosis specifically in M1 macrophages. Conversely, the recognition of lactose by the highly expressed lactose receptor (MGL) on M2 macrophages triggers the Eat Me signal. The recognition between receptors and ligands shows selective range specificity, wherein the interaction between lactose and MGL indirectly creates spatial hindrance against the Self peptide-SIRPα protein interaction, consequently inhibiting the response of M2 macrophages to the Do not Eat Me signal. Reprinted from Tang Y, Tang Z, Li P et al. Precise delivery of nanomedicines to M2 macrophages by combining “eat me/don’t eat me” signals and its anticancer application. ACS Nano. 2021; 15(11): 18,100–18,112. Copyright 2021, with permission from ACS publications.Citation105
Targeting tumor-associated macrophages (TAMs) by repolarizing M2 anti-inflammatory macrophages into M1 pro-inflammatory macrophages is an effective strategy for tumor therapy. This process, known as TAM reprogramming or education, can convert the pro-tumor phenotype into an anti-tumor one. Small molecules and nanoparticle formulations, such as toll-like receptor agonists, cytokines, antibodies, and RNA, have been investigated to achieve macrophage repolarization. For example, β-cyclodextrin nanoparticles loaded with R848 (CDNP-R848) can activate TLR7 and TLR8 in TAMs, converting them into an M1 phenotype.Citation164 Some of the vectors can also assist in realizing the reprogramming of M2 TAM by nanomedicines, such as biomimetic nanovesicles called phagocytosable cell wall-like (PCCW) systems which could target TAMs and promote immune destruction of tumor cells.Citation161,Citation165
Obesity
Obesity is a metabolic disorder characterized by excessive accumulation of white adipose tissue (WAT), leading to low-grade systemic inflammation and associated complications. The presence of macrophages within WAT contributes to this inflammation, with increased pro-inflammatory M1-like macrophages in obese individuals.Citation166–168
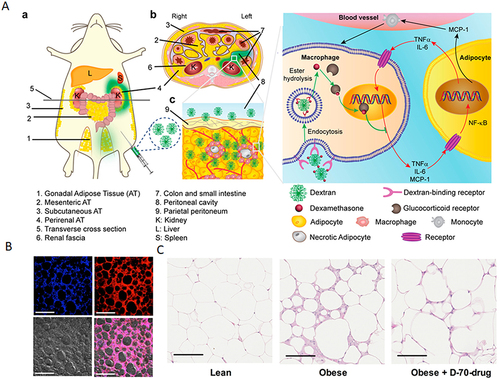
M1-type macrophages contribute to tissue inflammation and insulin resistance, while M2-type macrophages are more prevalent in lean individuals. Su et al, utilizing a combined treatment approach involving PHB P3 peptide-modified RHDL to enhance the expression of brown adipocyte markers, reduce adipocyte size, and inhibit weight gain caused by a high-fat diet.Citation169 Additionally, ASXL2 gene plays an important role in regulating macrophage response to dietary factors. Therefore, deletion of the ASXL2 gene has been shown to increase caloric expenditure in obese mice.Citation170 In a study by Liang et al, using polysaccharide nanocarriers spliced with anti-inflammatory drugs (D-70-drug) and demonstrated that more than 63% of the administered nanoparticles accumulated in visceral adipose tissue and then taken up by M1 macrophages through receptor-mediated endocytosis ().Citation171
Figure 6 (A) Proposed mechanism of dextran-dexamethasone conjugate accumulation in obese Visceral Adipose Tissue (VAT), macrophage uptake, and uncoupling of the paracrine loop between M1 macrophages and adipocytes: (a) In obese mice following intraperitoneal left-side injection, dextran conjugates (highlighted in green) accumulate in the left perirenal adipose tissue (AT) and left gonadal AT. The anatomical depiction shows mice with different fat depots: one mesenteric, two perirenal, and two gonadal AT depots. (b) A transverse cross-section of the mouse abdomen illustrates the localization of the green dextran solution after administration into the peritoneal cavity. (cInflamed adipose tissue enables rapid association of dextran conjugates with M1 macrophages by transport across the peritoneum to directly access interstitial cells. (B) Dextran nanocarrier uptake by myeloid cells in the adipose tissue of obese mice. Obese mice were intraperitoneally injected with D-70-fluor. Twenty-four hours later, the left gonadal adipose tissue was isolated, sectioned, and stained with Hoechst dye. Confocal laser scanning microscopy images show the Hoechst channel (top left), the D-fluor channel (top right), bright-field microscopy (bottom left), and the overlay (bottom right). D-70 was observed within the interstitial space between adipocytes. Scale bars: 200 μm. (C) In vivo anti-inflammatory effects of D-70-drug in obese mouse VAT. Representative hematoxylin-and-eosin-stained images of paraffin-embedded gonadal adipose tissue isolated from each mouse group. Scale bar: 100 μm. Reprinted from Ma L, Liu TW, Wallig MA et al. Efficient targeting of adipose tissue macrophages in obesity with polysaccharide nanocarriers. ACS Nano. 2016;10(7): 6952–6962. Copyright 2016, with permission from ACS publications.Citation171
Non-Alcoholic Fatty Liver Disease
NAFLD is a prevalent metabolic syndrome in obese populations, with non-alcoholic fatty liver (NAFL) and non-alcoholic steatohepatitis (NASH) as its subtypes. Hepatic resident macrophages have an important role in the progression of NAFLD. First, MCP-1 secreted by non-parenchymal liver cells recruits monocytes, which differentiate into macrophages and promote fibroblast activation.Citation172–174 Secondly, inflammation plays a crucial role, triggered by LPS binding to Kupffer cell receptors, resulting in the release of cytokines and chemokines like TGF-β and IL-1β that contribute to NAFLD pathogenesis.Citation175–177 Therefore, targeting lipid metabolism, inflammation, and macrophages can offer effective treatment strategies for NAFLD ().Citation178,Citation179
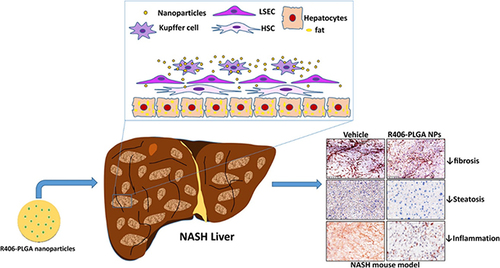
Figure 7 PLGA nanoparticle-based delivery of SYK pathway inhibitors for NASH treatment. This study utilized R406, a small molecule inhibitor, and synthesized PLGA nanoparticles to block the Fc receptor signaling pathway, specifically inhibiting the SYK pathway and reducing immune complex-mediated inflammation. Reprinted from Kurniawan DW, Jajoriya AK, Dhawan G et al. Therapeutic inhibition of spleen tyrosine kinase in inflammatory macrophages using PLGA nanoparticles for the treatment of non-alcoholic steatohepatitis. J Control Release. 2018; 288: 227–238. Creative Commons.Citation180
Since liver macrophages are the largest group of phagocytes in the body, nanoparticles tend to co-localize with these cells. This characteristic can be utilized to deliver therapeutic agents specifically to the liver and macrophages, potentially improving the treatment outcomes for NAFLD.Citation35,Citation178,Citation179 For example, the results of Muralidhara et al showed that liposome-encapsulated lipophilic curcumin significantly attenuated the inflammatory polarization of macrophages.Citation181 And results from Celia Martínez-Sánchez et al also show that the treatment with dextran–dexamethasone reduced the secretion of inflammatory chemokines and directly attenuated the pro-fibrogenic response in HSCs and liver spheroids.Citation182 In addition, in the context of liver iron deposition, a nano-antioxidant named Snoman-HSA has been developed to target Kupffer cells. By delivering nitric oxide (NO) and thiol groups to the liver, this nanodrug exhibits hepatoprotective effects comparable to combined treatment with other nano-antioxidants and NO donors.Citation183,Citation184 Traber and Zomer used galactin-3 inhibitors to target inflammatory macrophage functions in liver diseases in mice. The treatment with galectin-3 resulted in regression of NASH and fibrosis.Citation185 In conclusion, targeting macrophages holds promise as a treatment option for the management of NAFLD and steatohepatitis.
Rheumatoid Arthritis (RA)
Rheumatoid arthritis (RA) is a chronic inflammatory autoimmune disease characterized by macrophage aggregation in synovial tissue, autoantibody production, synovitis, joint swelling, and cartilage and bone destruction.Citation186,Citation187 The increased number of macrophages in the synovium of RA patients sustains chronic inflammation by releasing pro-inflammatory cytokines (TNF-α, IL-1β, IL-6) and mediators like prostaglandins and chemokines. This process leads to pain, increased infiltration of inflammatory cells, synovial angiogenesis, and joint destruction.Citation188 Targeting macrophages in the treatment of RA can be achieved through two strategies: directly killing macrophages or reprogramming pro-inflammatory M1 macrophages into anti-inflammatory M2 macrophages.Citation189,Citation190
One approach involves using clodronate, a bisphosphonate that induces apoptosis in macrophages. To target macrophages specifically, clodronate can be encapsulated in liposomes. Animal experiments using low-dose liposomal clodronate injections have shown a temporary reduction in joint swelling and slowed the progression of RA. However, clodronate may cause synovitis, and rapid clearance of nanoparticles by macrophages in the reticuloendothelial system (RES) can lead to reduced drug availability. Overcoming these challenges in selectively delivering nanodrugs to targeted organs remains a significant task.Citation19,Citation191
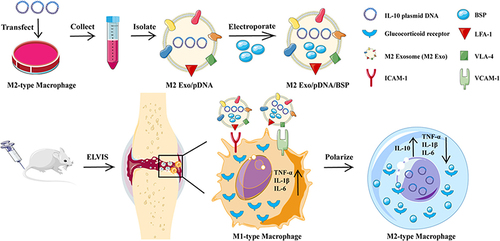
Alternatively, modulation of the inflammatory microenvironment within joints can convert M1 macrophages to anti-inflammatory M2 macrophages to alleviate inflammation. Delivering IL-10 plasmid DNA nanoparticles can promote macrophage polarization from an M1 to an M2 phenotype ().Citation192 Folate-modified silver nanoparticles (FA AgNPs) have been developed to actively target M1 macrophages and induce their reversion and polarization to M2 macrophages, effectively treating RA.Citation193 pH/ROS dual-responsive carriers loaded with glucocorticoids and surface-modified with RGD peptide have also been used and demonstrated that these nanomedicines were readily phagocytosed by macrophages and synovial cells, promoting the transformation of M1 macrophages into an M2 phenotype, thereby reducing inflammation, swelling, and joint destruction in mice.Citation194
Figure 8 Engaging molecularly engineered M2 macrophage-derived exosomes with inflammation tropism and anti-inflammatory capabilities for co-delivery of IL-10 pDNA and GCs to achieve rheumatoid arthritis (RA) treatment via M1-to-M2 macrophage repolarization. Reprinted from J Control Release, 341, Li H, Feng Y, Zheng X et al. M2-type exosomes nanoparticles for rheumatoid arthritis therapy via macrophage re-polarization. 16–30. Copyright 2022, with permission from Elsevier.Citation195
Atherosclerosis
Atherosclerosis is a progressive inflammatory disease characterized by lipid accumulation and macrophage infiltration in the arterial wall. It can lead to the formation of atherosclerotic plaques, narrowing the arteries. Macrophages play a crucial role in atherosclerosis by absorbing lipoproteins, contributing to plaque formation, and secreting inflammatory mediators.Citation196,Citation197
Nanomaterial-based therapies targeting macrophages in atherosclerosis show promise. Lowering lipid levels can promote regression of atherosclerosis by inducing an M2-like phenotype in plaques. Therefore, strategies that induce macrophages to transition into an M2 polarization state may be beneficial in treating atherosclerosis.Citation198,Citation199 Such as amino-functionalized hyaluronic acid nanoparticles (HA-NPs) could selectively target macrophages associated with atherosclerotic plaques, reducing inflammation, and transforming M1-type macrophages into M2-type macrophages.Citation200 In addition, inducing M2 polarization by upregulating fatty acid oxidation (FAO) has shown the potential to reduce atherosclerosis occurrence. Targeting scavenger receptor B type I (SR-BI) using a peptide ligand and a macrophage-targeted lipid nanoparticle loaded with an inhibitor of the CD40-induced inflammatory NF-kB pathway could suppress inflammation in plaques.Citation201,Citation202 In addition, specific drug delivery systems have been developed to promote cholesterol efflux in macrophages/foam cells. This strategy could reduce cellular cholesterol accumulation, improving mobility, decreasing lipid storage, and redirecting macrophages towards an M2 phenotype.Citation203
The design of biomimetic nanocarriers based on cellular biological actions also has great potential in atherosclerosis treatment. Gao et al designed ROS-responsive nanoparticles with lovastatin as a model drug and macrophage membranes as the outer layer, which could target atherosclerosis based on macrophage inflammatory chemotaxis and also improve the therapeutic effect on mouse atherosclerosis ().Citation204 While Wang et al coated macrophage membranes (MM) on the surface of rapamycin-carrying PLGA nanoparticles (RAPNPs) to prepare bionic nanoparticles (MM/RAPNPs), MM/RAPNPs can be effectively targeted and accumulated in atherosclerotic lesions, thereby slowing down the progression of atherosclerosis.Citation205 Second, the macrophage’s tropism for atherosclerosis makes it an effective carrier to target atherosclerosis. Hu et al constructed macrophages loaded with the Tetrapod PdH Nanoenzyme, which could be targeted to the arterial plaques, thereby decreasing the level of ROS, and significantly inhibiting inflammation-associated pathological processes. In addition, the tip-like morphology of the PdH nanoenzyme could further trigger a strong autophagic response in macrophages, which could synergistically maintain intracellular homeostasis.Citation206
Figure 9 Schematic illustration of the combinational regulation of plaque microenvironment by plaque macrophage-targeted nanosystems: 37pALNP/6877002 and 37pA-PtLNP. During atherosclerosis (AS) therapy, 37pA-PtLNP effectively scavenges intracellular reactive oxygen species (ROS) in inflammatory macrophages within the plaque. On the other hand, 37pA-LNP/6877002 inhibits the activities of TRAF6, leading to a modest decrease in the proportion of the proinflammatory macrophage phenotype (reducing the expression of CD86 and iNOS). More importantly, the combination of 37pA-PtLNP and 37pA-LNP/6877002 regulates the atherosclerotic plaque microenvironment and achieves satisfactory stabilization of plaques with minimal progression. Reprinted from Yang Q, Jiang H, Wang Y et al. Plaque Macrophage-Targeting Nanosystems with Cooperative Co-Regulation of ROS and TRAF6 for Stabilization of Atherosclerotic Plaques. Adv. Funct. Mater. 2018;33: 2,301,053. © 2023 Wiley-VCH GmbH.Citation201
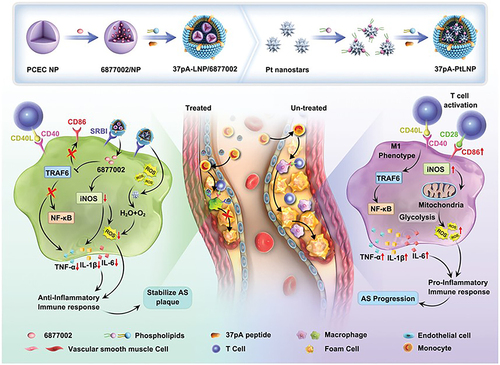
Inflammatory Bowel Disease (IBD)
Inflammatory bowel disease (IBD), including ulcerative colitis (UC) and Crohn’s disease (CD), is a chronic inflammatory disorder of the gastrointestinal tract. Macrophages play a protective role in the healthy intestinal epithelium by defending against pathogens. Disrupted cytokine balance leads to monocyte infiltration and differentiation into tissue-resident macrophages, releasing pro-inflammatory cytokines that exacerbate inflammation.Citation207 During steady-state, monocytes recruited to the gut differentiate into mature tissue-resident macrophages with an anti-inflammatory M2-like phenotype. These macrophages secrete cytokines like IL-10 and TGF-β, enhancing the epithelial barrier integrity and promoting stem cell renew.Citation207 Therefore, IBD treatment methods always impact macrophage function by inhibiting inflammatory signaling or inducing M2 polarization.Citation208,Citation209 Approaches like adoptive transfer of M2 macrophages,Citation210,Citation211 oral administration of Lupeol, tumor necrosis factor α stimulated gene 6, IL-33, and probiotic butyric acid bacteria to adoptive transfer of M2 macrophages have shown benefits for IBD treatment.Citation212–214
Nanomedicines offer a potential solution for targeted drug delivery to macrophages in inflammatory bowel disease (IBD), effectively reducing inflammation and reversing the actions of macrophages. The interaction between nanoscale carriers and macrophages facilitates their recognition and phagocytosis, enabling targeted imaging. In the context of IBD, tumor necrosis factor-alpha (TNF-α) plays a crucial role in its pathogenesis. Zhen et al have constructed nanocomplexes using konjac glucan (cKGM) combined with anti-TNF-α antisense oligodeoxynucleotides, which effectively reduced local TNF-α levels in the colon and suppressed inflammatory symptoms. These findings demonstrate the potential of using nanocomplexes as an approach to alleviate inflammation and counteract the actions of macrophages in IBD ().Citation215 Another approach involves using mannose-glycosylated cationic bioreducible polymers encapsulating RNA interference (RNAi) against TNF-α. Targeting macrophages with this method significantly reduces TNF-α levels in colonic tissues.Citation216 Hyaluronic acid (HA)-bilirubin nanomedicine (HABN) has been developed as a drug specifically targeting CD44 receptors. This nanomedicine helps clear reactive oxygen species and modulate the gut microbiota for mucosal repair.Citation217 In addition, gelatin microspheres loaded with IL-10 have also been employed to enhance the effectiveness of IBD treatment.Citation218
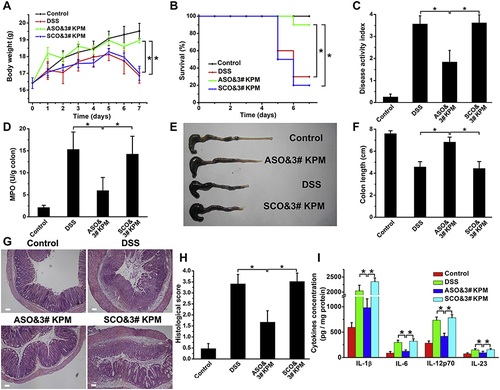
Figure 10 The microspheric vehicle, 3# KPM, composed of cationic konjac glucomannan (cKGM), phytagel, and an antisense oligonucleotide against TNF-α, showed anti-colitis activity. (A) Body weight changes were observed over time. (B) Survival analysis. (C) Disease activity index was calculated to evaluate the severity of colitis. (D) myeloperoxidase (MPO) activity determination. (E and F) Colon length was measured as an indicator of inflammation and tissue damage. (G and H) Colon sections were stained with H&E and histopathological scoring was conducted to assess inflammatory cell infiltration and tissue damage. Scale bar: 100 μm. (I) The levels of colonic inflammatory cytokines (IL-1β, IL-6, IL-12p70, and IL-23) were determined in DSS colitic mice with different treatments. Results are expressed as the mean (SEM). *p<0.05. Reprinted from Biomaterials; 48, Huang Z, Gan J, Jia L et al. An orally administrated nucleotide-delivery vehicle targeting colonic macrophages for the treatment of inflammatory bowel disease. 26–36. Copyright 2015, with permission from Elsevier.Citation215
Conclusion
In conclusion, macrophages play a pivotal role in immune responses and have emerged as crucial therapeutic targets in various diseases. Macrophages are an important part of the body’s immune system. Therefore, the development of tumors and inflammatory and immune-related diseases such as NAFLD, IBD and RA is accompanied by macrophage dysfunction and mis-polarization. Therefore, it is an important therapeutic target in these diseases. Relevant drug formulations are already undergoing clinical trials. Compared with conventional drugs, nanomedicines show unique advantages in improving the stability and solubility of drugs, and based on the characteristic of macrophage recognition of phagocytosis of foreign substances, it makes nanomedicines can easily contact with macrophages and exert therapeutic effects.
In the physiological environment, carriers will adsorb serum proteins soon after entering the body, and the conditioning elements in serum will enhance the recognition of carriers by macrophages. However, different carrier materials have different properties, so their interaction with macrophages will be different. Here we have compiled a list of the effects of physical and chemical properties of carriers such as particle size, hardness, traits and potential on their interaction with macrophages, in general, in the range of 10~5000 nm, carriers with larger particle size, softer surfaces, higher roundness and stronger positive potentials will enhance interaction with macrophages.
In addition, targeted drug delivery systems offer a promising approach to delivering drugs specifically to macrophages, thereby enhancing efficacy while minimizing toxicity. Passive targeting exploits the phagocytic activity of macrophages, allowing nanoscale carriers to accumulate in tissues abundant in macrophages. However, when macrophages need to be targeted outside these tissues, barriers must be overcome for carriers to reach the desired sites. Active targeting strategies employ macrophage-specific ligands on carrier surfaces to enhance recognition and uptake by target macrophages, thus improving drug delivery efficiency. While liposomal clodronate formulations have already found their way into clinical applications, further research is needed to fully grasp macrophage targeting and develop personalized drug delivery systems. Overcoming challenges such as heterogeneous macrophage populations, fluctuating marker expression, and real-time monitoring of changes in macrophage phenotypes and functions remains critical.Citation219
The role of nanomedicines in modulating the role of macrophages to enhance disease treatment is generally divided into three main blocks, including 1) inhibition of macrophage recruitment, 2) direct killing of macrophages and 3) repolarization (re-education) of macrophages. For the inhibition of macrophage recruitment, some drugs have been clinically validated and have achieved some efficacy. However, the strategy is focused on the control of the disease. Direct killing of macrophages is generally carried out by liposomes encapsulating clodronate, which is effective and simple. However, there are greater safety concerns with this approach, as this killing is not selective. Re-education of macrophages, on the other hand, has better expectations and has received a lot of attention because of its higher safety profile and its ability to convert pathogenic macrophages into therapeutic macrophages. In this regard, it is worth noting that some of the nanomedicine carrier materials themselves have certain macrophage re-education properties, and thus the organic combination of carrier material design and encapsulated drugs may be a more valuable breakthrough direction in the future.
Disclosure
The authors declare no conflicts of interest in this work.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 32001001), the Natural Science Foundation of Shandong Province (No. ZR2021MH118), Shandong Province Postdoctoral Innovation Project (202102065), Special funds for Taishan Scholars Project (tsqnz20221156, tsqn202306383), China Postdoctoral Science Foundation (2022M711972), and academic promotion program of Shandong First Medical University (2019LJ003).
References
- Davies LC, Jenkins SJ, Allen JE, Taylor PR. Tissue-resident macrophages. Nat Immunol. 2013;14(10):986–995. doi:10.1038/ni.2705
- Gordon S, Taylor PR. Monocyte and macrophage heterogeneity. Nat Rev Immunol. 2005;5(12):953–964. doi:10.1038/nri1733
- Ginhoux F, Guilliams M. Tissue-Resident Macrophage Ontogeny and Homeostasis. Immunity. 2016;44(3):439–449. doi:10.1016/j.immuni.2016.02.024
- Park MD, Silvin A, Ginhoux F, Merad M. Macrophages in health and disease. Cell. 2022;185(23):4259–4279. doi:10.1016/j.cell.2022.10.007
- Wynn TA, Chawla A, Pollard JW. Macrophage biology in development, homeostasis and disease. Nature. 2013;496(7446):445–455. doi:10.1038/nature12034
- Murray PJ, Wynn TA. Protective and pathogenic functions of macrophage subsets. Nat Rev Immunol. 2011;11(11):723–737. doi:10.1038/nri3073
- Martinez FO, Gordon S. The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000prime Rep. 2014;6:13. doi:10.12703/P6-13
- Mantovani A, Sozzani S, Locati M, Allavena P, Sica A. Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002;23(11):549–555. doi:10.1016/S1471-4906(02)02302-5
- Fukui S, Iwamoto N, Takatani A, et al. M1 and M2 Monocytes in Rheumatoid Arthritis: a Contribution of Imbalance of M1/M2 Monocytes to Osteoclastogenesis. Front Immunol. 2017;8:1958. doi:10.3389/fimmu.2017.01958
- Shields CW, Wang LL, Evans MA, Mitragotri S. Materials for Immunotherapy. Adv Mater. 2020;32(13):1901633. doi:10.1002/adma.201901633
- Jain RK, Martin JD, Stylianopoulos T. The role of mechanical forces in tumor growth and therapy. Annu Rev Biomed Eng. 2014;16:321–346. doi:10.1146/annurev-bioeng-071813-105259
- Papini E, Tavano R, Mancin F. Opsonins and Dysopsonins of Nanoparticles: facts, Concepts, and Methodological Guidelines. Front Immunol. 2020;11:567365. doi:10.3389/fimmu.2020.567365
- Zhang YN, Poon W, Tavares AJ, McGilvray ID, Chan WCW. Nanoparticle–liver interactions: cellular uptake and hepatobiliary elimination. J Controlled Release. 2016;240:332–348. doi:10.1016/j.jconrel.2016.01.020
- Wei X, Ying M, Dehaini D, et al. Nanoparticle Functionalization with Platelet Membrane Enables Multifactored Biological Targeting and Detection of Atherosclerosis. ACS Nano. 2018;12(1):109–116. doi:10.1021/acsnano.7b07720
- Zhao Y, He Z, Gao H, et al. Fine Tuning of Core–Shell Structure of Hyaluronic Acid/Cell-Penetrating Peptides/siRNA Nanoparticles for Enhanced Gene Delivery to Macrophages in Antiatherosclerotic Therapy. Biomacromolecules. 2018;19(7):2944–2956. doi:10.1021/acs.biomac.8b00501
- Lee GY, Kim JH, Choi KY, et al. Hyaluronic acid nanoparticles for active targeting atherosclerosis. Biomaterials. 2015;53:341–348. doi:10.1016/j.biomaterials.2015.02.089
- Poh S, Putt KS, Low PS. Folate-Targeted Dendrimers Selectively Accumulate at Sites of Inflammation in Mouse Models of Ulcerative Colitis and Atherosclerosis. Biomacromolecules. 2017;18(10):3082–3088. doi:10.1021/acs.biomac.7b00728
- He H, Yuan Q, Bie J, et al. Development of mannose functionalized dendrimeric nanoparticles for targeted delivery to macrophages: use of this platform to modulate atherosclerosis. Transl Res. 2018;193:13–30. doi:10.1016/j.trsl.2017.10.008
- van Rooijen N, van Kesteren-Hendrikx E. Clodronate liposomes: perspectives in research and therapeutics. J Liposome Res. 2002;12(1–2):81–94. doi:10.1081/lpr-120004780
- He H, Lu Y, Qi J, Zhu Q, Chen Z, Wu W. Adapting liposomes for oral drug delivery. Acta Pharm Sin B. 2019;9(1):36–48. doi:10.1016/j.apsb.2018.06.005
- Tenzer S, Docter D, Kuharev J, et al. Rapid formation of plasma protein Corona critically affects nanoparticle pathophysiology. Nat Nanotechnol. 2013;8(10):772–781. doi:10.1038/nnano.2013.181
- Milani S, Baldelli Bombelli F, Pitek AS, Dawson KA, Rädler J. Reversible versus Irreversible Binding of Transferrin to Polystyrene Nanoparticles: soft and Hard Corona. ACS Nano. 2012;6(3):2532–2541. doi:10.1021/nn204951s
- Nguyen VH, Lee BJ. Protein Corona: a new approach for nanomedicine design. Int J Nanomedicine. 2017;12:3137–3151. doi:10.2147/IJN.S129300
- Guan J, Shen Q, Zhang Z, et al. Enhanced immunocompatibility of ligand-targeted liposomes by attenuating natural IgM absorption. Nat Commun. 2018;9(1):2982. doi:10.1038/s41467-018-05384-1
- Zabielska-Koczywąs K, Lechowski R. The Use of Liposomes and Nanoparticles as Drug Delivery Systems to Improve Cancer Treatment in Dogs and Cats. Molecules. 2017;22(12):2167. doi:10.3390/molecules22122167
- Lamichhane N, Udayakumar T, D’Souza W, et al. Liposomes: clinical Applications and Potential for Image-Guided Drug Delivery. Molecules. 2018;23(2):288. doi:10.3390/molecules23020288
- Shaheen SM, Shakil Ahmed FR, Hossen MN, Ahmed M, Amran MS, Ul-Islam MA. Liposome as a Carrier for Advanced Drug Delivery. Pak J Biol Sci. 2006;9(6):1181–1191. doi:10.3923/pjbs.2006.1181.1191
- Poirier VJ, Thamm DH, Kurzman ID, et al. Liposome-Encapsulated Doxorubicin (Doxil) and Doxorubicin in the Treatment of Vaccine-Associated Sarcoma in Cats. J Vet Intern Med. 2002;16(6):726–731. doi:10.1111/j.1939-1676.2002.tb02415.x
- Walkey CD, Olsen JB, Guo H, Emili A, Chan WCW. Nanoparticle size and surface chemistry determine serum protein adsorption and macrophage uptake. J Am Chem Soc. 2012;134(4):2139–2147. doi:10.1021/ja2084338
- Schöttler S, Becker G, Winzen S, et al. Protein adsorption is required for stealth effect of poly(ethylene glycol)- and poly(phosphoester)-coated nanocarriers. Nat Nanotechnol. 2016;11(4):372–377. doi:10.1038/nnano.2015.330
- Cao H, Dan Z, He X, et al. Liposomes Coated with Isolated Macrophage Membrane Can Target Lung Metastasis of Breast Cancer. ACS Nano. 2016;10(8):7738–7748. doi:10.1021/acsnano.6b03148
- He H, Guo C, Wang J, et al. Leutusome: a Biomimetic Nanoplatform Integrating Plasma Membrane Components of Leukocytes and Tumor Cells for Remarkably Enhanced Solid Tumor Homing. Nano Lett. 2018;18(10):6164–6174. doi:10.1021/acs.nanolett.8b01892
- Belhadj Z, He B, Deng H, et al. A combined “eat me/don’t eat me” strategy based on extracellular vesicles for anticancer nanomedicine. J Extracell Vesicles. 2020;9(1):1806444. doi:10.1080/20013078.2020.1806444
- Tang Y, Wang X, Li J, et al. Overcoming the Reticuloendothelial System Barrier to Drug Delivery with a “Don’t-Eat-Us” Strategy. ACS Nano. 2019;13(11):13015–13026. doi:10.1021/acsnano.9b05679
- Tsoi KM, MacParland SA, Ma XZ, et al. Mechanism of hard-nanomaterial clearance by the liver. Nat Mater. 2016;15(11):1212–1221. doi:10.1038/nmat4718
- Cheng L, Niu MM, Yan T, et al. Bioresponsive micro-to-nano albumin-based systems for targeted drug delivery against complex fungal infections. Acta Pharm Sin B. 2021;11(10):3220–3230. doi:10.1016/j.apsb.2021.04.020
- Wang W, Zhou X, Bian Y, et al. Dual-targeting nanoparticle vaccine elicits a therapeutic antibody response against chronic hepatitis B. Nat Nanotechnol. 2020;15(5):406–416. doi:10.1038/s41565-020-0648-y
- Son H, Choi HS, Baek SE, et al. Shear stress induces monocyte/macrophage-mediated inflammation by upregulating cell-surface expression of heat shock proteins. Biomed Pharmacother. 2023;161:114566. doi:10.1016/j.biopha.2023.114566
- Yazdimamaghani M, Barber ZB, Hadipour Moghaddam SP, Ghandehari H. Influence of Silica Nanoparticle Density and Flow Conditions on Sedimentation, Cell Uptake, and Cytotoxicity. Mol Pharm. 2018;15(6):2372–2383. doi:10.1021/acs.molpharmaceut.8b00213
- Steinbach EC, Plevy SE. The Role of Macrophages and Dendritic Cells in the Initiation of Inflammation in IBD. Inflamm Bowel Dis. 2014;20(1):166–175. doi:10.1097/MIB.0b013e3182a69dca
- Singh R, Mishra MK, Aggarwal H. Inflammation, Immunity, and Cancer. Mediators Inflamm. 2017;2017:1. doi:10.1155/2017/6027305
- Qie Y, Yuan H, von Roemeling CA, et al. Surface modification of nanoparticles enables selective evasion of phagocytic clearance by distinct macrophage phenotypes. Sci Rep. 2016;6(1):26269. doi:10.1038/srep26269
- Schultze JL, Schmidt SV. Molecular features of macrophage activation. Semin Immunol. 2015;27(6):416–423. doi:10.1016/j.smim.2016.03.009
- Bonilla DL, Bhattacharya A, Sha Y, et al. Autophagy Regulates Phagocytosis by Modulating the Expression of Scavenger Receptors. Immunity. 2013;39(3):537–547. doi:10.1016/j.immuni.2013.08.026
- Tarique AA, Logan J, Thomas E, Holt PG, Sly PD, Fantino E. Phenotypic, Functional, and Plasticity Features of Classical and Alternatively Activated Human Macrophages. Am J Respir Cell Mol Biol. 2015;53(5):676–688. doi:10.1165/rcmb.2015-0012OC
- Verreck FAW, de Boer T, Langenberg DML, et al. Human IL-23-producing type 1 macrophages promote but IL-10-producing type 2 macrophages subvert immunity to (myco)bacteria. Proc Natl Acad Sci. 2004;101(13):4560–4565. doi:10.1073/pnas.0400983101
- Verreck FAW, de Boer T, Langenberg DML, van der Zanden L, Ottenhoff THM. Phenotypic and functional profiling of human proinflammatory type-1 and anti-inflammatory type-2 macrophages in response to microbial antigens and IFN-γ- and CD40L-mediated costimulation. J Leukoc Biol. 2005;79(2):285–293. doi:10.1189/jlb.0105015
- Jaguin M, Houlbert N, Fardel O, Lecureur V. Polarization profiles of human M-CSF-generated macrophages and comparison of M1-markers in classically activated macrophages from GM-CSF and M-CSF origin. Cell Immunol. 2013;281(1):51–61. doi:10.1016/j.cellimm.2013.01.010
- Schraufstatter IU, Zhao M, Khaldoyanidi SK, DiScipio RG. The chemokine CCL18 causes maturation of cultured monocytes to macrophages in the M2 spectrum: macrophage maturation by CCL18. Immunology. 2012;135(4):287–298. doi:10.1111/j.1365-2567.2011.03541.x
- Vogel DYS, Glim JE, Stavenuiter AWD, et al. Human macrophage polarization in vitro: maturation and activation methods compared. Immunobiology. 2014;219(9):695–703. doi:10.1016/j.imbio.2014.05.002
- Lee A, Septiadi D, Taladriz‐Blanco P, et al. Particle Stiffness and Surface Topography Determine Macrophage‐Mediated Removal of Surface Adsorbed Particles. Adv Healthc Mater. 2021;10(6):2001667. doi:10.1002/adhm.202001667
- Mitchell LA, Lauer FT, Burchiel SW, McDonald JD. Mechanisms for how inhaled multiwalled carbon nanotubes suppress systemic immune function in mice. Nat Nanotechnol. 2009;4(7):451–456. doi:10.1038/nnano.2009.151
- Kelly C, Jefferies C, Cryan SA. Targeted Liposomal Drug Delivery to Monocytes and Macrophages. J Drug Deliv. 2011;2011:1–11. doi:10.1155/2011/727241
- Ngambenjawong C, Gustafson HH, Pun SH. Progress in tumor-associated macrophage (TAM)-targeted therapeutics. Adv Drug Deliv Rev. 2017;114:206–221. doi:10.1016/j.addr.2017.04.010
- Tabata Y, Ikada Y. Effect of the size and surface charge of polymer microspheres on their phagocytosis by macrophage. Biomaterials. 1988;9(4):356–362. doi:10.1016/0142-9612(88)90033-6
- Inadomi JM, Vijan S, Janz NK, et al. Adherence to colorectal cancer screening: a randomized clinical trial of competing strategies. Arch Intern Med. 2012;172(7):575–582. doi:10.1001/archinternmed.2012.332
- He C, Hu Y, Yin L, Tang C, Yin C. Effects of particle size and surface charge on cellular uptake and biodistribution of polymeric nanoparticles. Biomaterials. 2010;31(13):3657–3666. doi:10.1016/j.biomaterials.2010.01.065
- Yue H, Wei W, Yue Z, et al. Particle size affects the cellular response in macrophages. Eur J Pharm Sci. 2010;41(5):650–657. doi:10.1016/j.ejps.2010.09.006
- Kuhn DA, Vanhecke D, Michen B, et al. Different endocytotic uptake mechanisms for nanoparticles in epithelial cells and macrophages. Beilstein J Nanotechnol. 2014;5:1625–1636. doi:10.3762/bjnano.5.174
- Gustafson HH, Holt-Casper D, Grainger DW, Ghandehari H. Nanoparticle uptake: the phagocyte problem. Nano Today. 2015;10(4):487–510. doi:10.1016/j.nantod.2015.06.006
- Champion JA, Mitragotri S. Role of target geometry in phagocytosis. Proc Natl Acad Sci. 2006;103(13):4930–4934. doi:10.1073/pnas.0600997103
- Champion JA, Walker A, Mitragotri S. Role of Particle Size in Phagocytosis of Polymeric Microspheres. Pharm Res. 2008;25(8):1815–1821. doi:10.1007/s11095-008-9562-y
- Champion JA, Mitragotri S. Shape Induced Inhibition of Phagocytosis of Polymer Particles. Pharm Res. 2009;26(1):244–249. doi:10.1007/s11095-008-9626-z
- Garapaty A, Champion JA. Tunable particles alter macrophage uptake based on combinatorial effects of physical properties: garapaty and Champion. Bioeng Transl Med. 2017;2(1):92–101. doi:10.1002/btm2.10047
- Albanese A, Tang PS, Chan WCW. The Effect of Nanoparticle Size, Shape, and Surface Chemistry on Biological Systems. Annu Rev Biomed Eng. 2012;14(1):1–16. doi:10.1146/annurev-bioeng-071811-150124
- Kinnear C, Moore TL, Rodriguez-Lorenzo L, Rothen-Rutishauser B, Petri-Fink A. Form Follows Function: nanoparticle Shape and Its Implications for Nanomedicine. Chem Rev. 2017;117(17):11476–11521. doi:10.1021/acs.chemrev.7b00194
- Anselmo AC, Mitragotri S. Impact of particle elasticity on particle-based drug delivery systems. Adv Drug Deliv Rev. 2017;108:51–67. doi:10.1016/j.addr.2016.01.007
- Palomba R, Palange AL, Rizzuti IF, et al. Modulating Phagocytic Cell Sequestration by Tailoring Nanoconstruct Softness. ACS Nano. 2018;12(2):1433–1444. doi:10.1021/acsnano.7b07797
- de Castro CE, Ribeiro CAS, Alavarse AC, et al. Nanoparticle–Cell Interactions: surface Chemistry Effects on the Cellular Uptake of Biocompatible Block Copolymer Assemblies. Langmuir. 2018;34(5):2180–2188. doi:10.1021/acs.langmuir.7b04040
- Verma A, Stellacci F. Effect of Surface Properties on Nanoparticle–Cell Interactions. Small. 2010;6(1):12–21. doi:10.1002/smll.200901158
- Rattan R, Bhattacharjee S, Zong H, et al. Nanoparticle-macrophage interactions: a balance between clearance and cell-specific targeting. Bioorg Med Chem. 2017;25(16):4487–4496. doi:10.1016/j.bmc.2017.06.040
- Lunov O, Syrovets T, Loos C, et al. Differential Uptake of Functionalized Polystyrene Nanoparticles by Human Macrophages and a Monocytic Cell Line. ACS Nano. 2011;5(3):1657–1669. doi:10.1021/nn2000756
- Liu X, Huang N, Li H, Jin Q, Ji J. Surface and Size Effects on Cell Interaction of Gold Nanoparticles with Both Phagocytic and Nonphagocytic Cells. Langmuir. 2013;29(29):9138–9148. doi:10.1021/la401556k
- Szolnoky G, Bata-Csörgö Z, Kenderessy AS, et al. A Mannose-Binding Receptor is Expressed on Human Keratinocytes and Mediates Killing of Candida albicans. J Invest Dermatol. 2001;117(2):205–213. doi:10.1046/j.1523-1747.2001.14071.x
- Scodeller P, Simón-Gracia L, Kopanchuk S, et al. Precision Targeting of Tumor Macrophages with a CD206 Binding Peptide. Sci Rep. 2017;7(1):14655. doi:10.1038/s41598-017-14709-x
- Jaynes JM, Sable R, Ronzetti M, et al. Mannose receptor (CD206) activation in tumor-associated macrophages enhances adaptive and innate antitumor immune responses. Sci Transl Med. 2020;12(530):eaax6337. doi:10.1126/scitranslmed.aax6337
- Parker CC, Bin Salam A, Song PN, et al. Evaluation of a CD206-Targeted Peptide for PET Imaging of Macrophages in Syngeneic Mouse Models of Cancer. Mol Pharm. 2023;20(5):2415–2425. doi:10.1021/acs.molpharmaceut.2c00977
- Zhan X, Jia L, Niu Y, et al. Targeted depletion of tumour-associated macrophages by an alendronate–glucomannan conjugate for cancer immunotherapy. Biomaterials. 2014;35(38):10046–10057. doi:10.1016/j.biomaterials.2014.09.007
- Vyas MN, Vyas NK, Quiocho FA. Crystallographic Analysis of the Epimeric and Anomeric Specificity of the Periplasmic Transport/Chemosensory Protein Receptor for D-Glucose and D-Galactose. Biochemistry. 1994;33(16):4762–4768. doi:10.1021/bi00182a003
- Müller C, Schibli R. Prospects in Folate Receptor-Targeted Radionuclide Therapy. Front Oncol. 2013;3. doi:10.3389/fonc.2013.00249
- Skytthe MK, Graversen JH, Moestrup SK. Targeting of CD163+ Macrophages in Inflammatory and Malignant Diseases. Int J Mol Sci. 2020;21(15):5497. doi:10.3390/ijms21155497
- Barclay AN. Signal regulatory protein alpha (SIRPα)/CD47 interaction and function. Curr Opin Immunol. 2009;21(1):47–52. doi:10.1016/j.coi.2009.01.008
- Tang T, Wei Y, Kang J, et al. Tumor-specific macrophage targeting through recognition of retinoid X receptor beta. J Controlled Release. 2019;301:42–53. doi:10.1016/j.jconrel.2019.03.009
- Ngambenjawong C, Gustafson HH, Pineda JM, Kacherovsky NA, Cieslewicz M, Pun SH. Serum Stability and Affinity Optimization of an M2 Macrophage-Targeting Peptide (M2pep). Theranostics. 2016;6(9):1403–1414. doi:10.7150/thno.15394
- Ries CH, Hoves S, Cannarile MA, Rüttinger D. CSF-1/CSF-1R targeting agents in clinical development for cancer therapy. Curr Opin Pharmacol. 2015;23:45–51. doi:10.1016/j.coph.2015.05.008
- Ries CH, Cannarile MA, Hoves S, et al. Targeting Tumor-Associated Macrophages with Anti-CSF-1R Antibody Reveals a Strategy for Cancer Therapy. Cancer Cell. 2014;25(6):846–859. doi:10.1016/j.ccr.2014.05.016
- Stafford JH, Hirai T, Deng L, et al. Colony stimulating factor 1 receptor inhibition delays recurrence of glioblastoma after radiation by altering myeloid cell recruitment and polarization. Neuro-Oncol. 2016;18(6):797–806. doi:10.1093/neuonc/nov272
- Pyonteck SM, Akkari L, Schuhmacher AJ, et al. CSF-1R inhibition alters macrophage polarization and blocks glioma progression. Nat Med. 2013;19(10):1264–1272. doi:10.1038/nm.3337
- Cannarile MA, Weisser M, Jacob W, Jegg AM, Ries CH, Rüttinger D. Colony-stimulating factor 1 receptor (CSF1R) inhibitors in cancer therapy. J Immunother Cancer. 2017;5(1):53. doi:10.1186/s40425-017-0257-y
- Matlung HL, Szilagyi K, Barclay NA, van den Berg TK. The CD47-SIRPα signaling axis as an innate immune checkpoint in cancer. Immunol Rev. 2017;276(1):145–164. doi:10.1111/imr.12527
- Chao MP, Alizadeh AA, Tang C, et al. Anti-CD47 Antibody Synergizes with Rituximab to Promote Phagocytosis and Eradicate Non-Hodgkin Lymphoma. Cell. 2010;142(5):699–713. doi:10.1016/j.cell.2010.07.044
- Weiskopf K, Ring AM, Ccm H, et al. Engineered SIRPα Variants as Immunotherapeutic Adjuvants to Anticancer Antibodies. Science. 2013;341(6141):88–91. doi:10.1126/science.1238856
- Chen WC, Kawasaki N, Nycholat CM, et al. Antigen Delivery to Macrophages Using Liposomal Nanoparticles Targeting Sialoadhesin/CD169. PLoS One. 2012;7(6):e39039. doi:10.1371/journal.pone.0039039
- Al Faraj A, Sultana Shaik A, Afzal S, Al Sayed B, Halwani R. MR imaging and targeting of a specific alveolar macrophage subpopulation in LPS-induced COPD animal model using antibody-conjugated magnetic nanoparticles. Int J Nanomedicine. 2014;1491. doi:10.2147/IJN.S59394
- Ecker DM, Jones SD, Levine HL. The therapeutic monoclonal antibody market. mAbs. 2015;7(1):9–14. doi:10.4161/19420862.2015.989042
- Nuhn L, Bolli E, Massa S, et al. Targeting Protumoral Tumor-Associated Macrophages with Nanobody-Functionalized Nanogels through Strain Promoted Azide Alkyne Cycloaddition Ligation. Bioconjug Chem. 2018;29(7):2394–2405. doi:10.1021/acs.bioconjchem.8b00319
- De Meyer T, Muyldermans S, Depicker A. Nanobody-based products as research and diagnostic tools. Trends Biotechnol. 2014;32(5):263–270. doi:10.1016/j.tibtech.2014.03.001
- Cieslewicz M, Tang J, Yu JL, et al. Targeted delivery of proapoptotic peptides to tumor-associated macrophages improves survival. Proc Natl Acad Sci. 2013;110(40):15919–15924. doi:10.1073/pnas.1312197110
- Qian Y, Qiao S, Dai Y, et al. Molecular-Targeted Immunotherapeutic Strategy for Melanoma via Dual-Targeting Nanoparticles Delivering Small Interfering RNA to Tumor-Associated Macrophages. ACS Nano. 2017;11(9):9536–9549. doi:10.1021/acsnano.7b05465
- Conde J, Bao C, Tan Y, et al. Dual Targeted Immunotherapy via In Vivo Delivery of Biohybrid RNAi-Peptide Nanoparticles to Tumor-Associated Macrophages and Cancer Cells. Adv Funct Mater. 2015;25(27):4183–4194. doi:10.1002/adfm.201501283
- Ngambenjawong C, Cieslewicz M, Schellinger JG, Pun SH. Synthesis and evaluation of multivalent M2pep peptides for targeting alternatively activated M2 macrophages. J Control Release. 2016;224:103–111. doi:10.1016/j.jconrel.2015.12.057
- Otvos L, Wade JD. Current challenges in peptide-based drug discovery. Front Chem. 2014;2. doi:10.3389/fchem.2014.00062
- Song M, Liu T, Shi C, Zhang X, Chen X. Bioconjugated Manganese Dioxide Nanoparticles Enhance Chemotherapy Response by Priming Tumor-Associated Macrophages toward M1-like Phenotype and Attenuating Tumor Hypoxia. ACS Nano. 2016;10(1):633–647. doi:10.1021/acsnano.5b06779
- Zhu S, Niu M, O’Mary H, Cui Z. Targeting of Tumor-Associated Macrophages Made Possible by PEG-Sheddable, Mannose-Modified Nanoparticles. Mol Pharm. 2013;10(9):3525–3530. doi:10.1021/mp400216r
- Tang Y, Tang Z, Li P, et al. Precise Delivery of Nanomedicines to M2 Macrophages by Combining “Eat Me/Don’t Eat Me” Signals and Its Anticancer Application. ACS Nano. 2021;15(11):18100–18112. doi:10.1021/acsnano.1c06707
- Irache JM, Salman HH, Gamazo C, Espuelas S. Mannose-targeted systems for the delivery of therapeutics. Expert Opin Drug Deliv. 2008;5(6):703–724. doi:10.1517/17425247.5.6.703
- Keefe AD, Pai S, Ellington A. Aptamers as therapeutics. Nat Rev Drug Discov. 2010;9(7):537–550. doi:10.1038/nrd3141
- Zhou J, Rossi J. Aptamers as targeted therapeutics: current potential and challenges. Nat Rev Drug Discov. 2017;16(3):181–202. doi:10.1038/nrd.2016.199
- Sylvestre M, Saxby CP, Kacherovsky N, Gustafson H, Salipante SJ, Pun SH. Identification of a DNA Aptamer That Binds to Human Monocytes and Macrophages. Bioconjug Chem. 2020;31(8):1899–1907. doi:10.1021/acs.bioconjchem.0c00247
- Qian H, Zhou T, Fu Y, et al. Self-assembled tetrahedral framework nucleic acid mediates tumor-associated macrophage reprogramming and restores antitumor immunity. Mol Ther Nucleic Acids. 2022;27:763–773. doi:10.1016/j.omtn.2021.12.036
- Qian H, Fu Y, Guo M, et al. Dual-aptamer-engineered M1 macrophage with enhanced specific targeting and checkpoint blocking for solid-tumor immunotherapy. Mol Ther. 2022;30(8):2817–2827. doi:10.1016/j.ymthe.2022.04.015
- Johnston MJW, Semple SC, Klimuk SK, Ansell S, Maurer N, Cullis PR. Characterization of the drug retention and pharmacokinetic properties of liposomal nanoparticles containing dihydrosphingomyelin. Biochim Biophys Acta BBA - Biomembr. 2007;1768(5):1121–1127. doi:10.1016/j.bbamem.2007.01.019
- Filipczak N, Pan J, Yalamarty SSK, Torchilin VP. Recent advancements in liposome technology. Adv Drug Deliv Rev. 2020;156:4–22. doi:10.1016/j.addr.2020.06.022
- Tang J, Rakshit M, Chua HM, et al. Liposome interaction with macrophages and foam cells for atherosclerosis treatment: effects of size, surface charge and lipid composition. Nanotechnology. 2021;32(50):505105. doi:10.1088/1361-6528/ac2810
- Vorselen D. Dynamics of phagocytosis mediated by phosphatidylserine. Biochem Soc Trans. 2022;50(5):1281–1291. doi:10.1042/BST20211254
- Toita R, Kang JH, Tsuchiya A. Phosphatidylserine liposome multilayers mediate the M1-to-M2 macrophage polarization to enhance bone tissue regeneration. Acta Biomater. 2022;154:583–596. doi:10.1016/j.actbio.2022.10.024
- Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478):eaau6977. doi:10.1126/science.aau6977
- Ben XY, Wang YR, Zheng HH, et al. Construction of Exosomes that Overexpress CD47 and Evaluation of Their Immune Escape. Front Bioeng Biotechnol. 2022;10:936951. doi:10.3389/fbioe.2022.936951
- Szebeni J, Alving CR, Rosivall L, et al. Animal models of complement-mediated hypersensitivity reactions to liposomes and other lipid-based nanoparticles. J Liposome Res. 2007;17(2):107–117. doi:10.1080/08982100701375118
- Kanlikilicer P, Bayraktar R, Denizli M, et al. Exosomal miRNA confers chemo resistance via targeting Cav1/p-gp/M2-type macrophage axis in ovarian cancer. eBioMedicine. 2018;38:100–112. doi:10.1016/j.ebiom.2018.11.004
- Yang K, Xiao Q, Niu M, Pan X, Zhu X. Exosomes in atherosclerosis: convergence on macrophages. Int J Biol Sci. 2022;18(8):3266–3281. doi:10.7150/ijbs.71862
- Feng Q, Ma X, Cheng K, et al. Engineered Bacterial Outer Membrane Vesicles as Controllable Two‐Way Adaptors to Activate Macrophage Phagocytosis for Improved Tumor Immunotherapy. Adv Mater. 2022;34(40):2206200. doi:10.1002/adma.202206200
- Joralemon MJ, McRae S, Emrick T. PEGylated polymers for medicine: from conjugation to self-assembled systems. Chem Commun. 2010;46(9):1377. doi:10.1039/b920570p
- Kolate A, Baradia D, Patil S, Vhora I, Kore G, Misra A. PEG — a versatile conjugating ligand for drugs and drug delivery systems. J Controlled Release. 2014;192:67–81. doi:10.1016/j.jconrel.2014.06.046
- Sabzehei F, Taromchi AH, Danafar H, Rashidzadeh H, Ramazani A. In vitro Characterization of Polyethyleneimine–Oleic Acid Cationic Micelle as a Novel Protein Carrier. Adv Biomed Res. 2023;12(1):126. doi:10.4103/abr.abr_303_22
- Hou Z, Zhou W, Guo X, et al. Poly(ϵ-Caprolactone)-Methoxypolyethylene Glycol (PCL-MPEG)-Based Micelles for Drug-Delivery: the Effect of PCL Chain Length on Blood Components, Phagocytosis, and Biodistribution. Int J Nanomedicine. 2022;17:1613–1632. doi:10.2147/IJN.S349516
- Klein ME, Rieckmann M, Lucas H, Meister A, Loppnow H, Mäder K. Phosphatidylserine (PS) and phosphatidylglycerol (PG) enriched mixed micelles (MM): a new nano-drug delivery system with anti-inflammatory potential? Eur J Pharm Sci. 2020;152:105451. doi:10.1016/j.ejps.2020.105451
- Sun JH, Liang X, Cai M, et al. Protein-Crowned Micelles for Targeted and Synergistic Tumor-Associated Macrophage Reprogramming to Enhance Cancer Treatment. Nano Lett. 2022;22(11):4410–4420. doi:10.1021/acs.nanolett.2c00901
- Liu L, He H, Liang R, et al. ROS-Inducing Micelles Sensitize Tumor-Associated Macrophages to TLR3 Stimulation for Potent Immunotherapy. Biomacromolecules. 2018;19(6):2146–2155. doi:10.1021/acs.biomac.8b00239
- Lombardo D, Kiselev MA, Caccamo MT. Smart Nanoparticles for Drug Delivery Application: development of Versatile Nanocarrier Platforms in Biotechnology and Nanomedicine. J Nanomater. 2019;2019:1–26. doi:10.1155/2019/3702518
- Pinelli F, Perale G, Rossi F. Coating and Functionalization Strategies for Nanogels and Nanoparticles for Selective Drug Delivery. Gels. 2020;6(1):6. doi:10.3390/gels6010006
- Chenthamara D, Subramaniam S, Ramakrishnan SG, et al. Therapeutic efficacy of nanoparticles and routes of administration. Biomater Res. 2019;23(1):20. doi:10.1186/s40824-019-0166-x
- Fu X, Bai H, Lyu F, Liu L, Wang S. Conjugated Polymer Nanomaterials for Phototherapy of Cancer. Chem Res Chin Univ. 2020;36(2):237–242. doi:10.1007/s40242-020-0012-7
- Li S, Wang X, Hu R, et al. Near-Infrared (NIR)-Absorbing Conjugated Polymer Dots as Highly Effective Photothermal Materials for In Vivo Cancer Therapy. Chem Mater. 2016;28(23):8669–8675. doi:10.1021/acs.chemmater.6b03738
- Xiong Z, Timokhina I, Pereloma E. Clustering, nano-scale precipitation and strengthening of steels. Prog Mater Sci. 2021;118:100764. doi:10.1016/j.pmatsci.2020.100764
- Fuchs AK, Syrovets T, Haas KA, et al. Carboxyl- and amino-functionalized polystyrene nanoparticles differentially affect the polarization profile of M1 and M2 macrophage subsets. Biomaterials. 2016;85:78–87. doi:10.1016/j.biomaterials.2016.01.064
- Huang YJ, Hung KC, Hung HS, Hsu SH. Modulation of macrophage phenotype by biodegradable polyurethane nanoparticles: the possible relation between macrophage polarization and immune response of nanoparticles. ACS Appl Mater Interfaces. 2018;10(23):19436–19448. doi:10.1021/acsami.8b04718
- Zanganeh S, Hutter G, Spitler R, et al. Iron oxide nanoparticles inhibit tumour growth by inducing pro-inflammatory macrophage polarization in tumour tissues. Nat Nanotechnol. 2016;11(11):986–994. doi:10.1038/nnano.2016.168
- Jin R, Liu L, Zhu W, et al. Iron oxide nanoparticles promote macrophage autophagy and inflammatory response through activation of toll-like Receptor-4 signaling. Biomaterials. 2019;203:23–30. doi:10.1016/j.biomaterials.2019.02.026
- Chen L, Ma X, Dang M, et al. Simultaneous T Cell Activation and Macrophage Polarization to Promote Potent Tumor Suppression by Iron Oxide‐Embedded Large‐Pore Mesoporous Organosilica Core–Shell Nanospheres. Adv Healthc Mater. 2019;8(9):1900039. doi:10.1002/adhm.201900039
- Guo JC, An Q, Guo M, et al. Oxygen-independent free radical generation mediated by core-shell magnetic nanocomposites synergizes with immune checkpoint blockade for effective primary and metastatic tumor treatment. Nano Today. 2021;36:101024. doi:10.1016/j.nantod.2020.101024
- Kang H, Kim S, Wong DSH, et al. Remote Manipulation of Ligand Nano-Oscillations Regulates Adhesion and Polarization of Macrophages in Vivo. Nano Lett. 2017;17(10):6415–6427. doi:10.1021/acs.nanolett.7b03405
- Rojas JM, Sanz-Ortega L, Mulens-Arias V, Gutiérrez L, Pérez-Yagüe S, Barber DF. Superparamagnetic iron oxide nanoparticle uptake alters M2 macrophage phenotype, iron metabolism, migration and invasion. Nanomedicine Nanotechnol Biol Med. 2016;12(4):1127–1138. doi:10.1016/j.nano.2015.11.020
- Shah A, Dobrovolskaia MA. Immunological effects of iron oxide nanoparticles and iron-based complex drug formulations: therapeutic benefits, toxicity, mechanistic insights, and translational considerations. Nanomedicine Nanotechnol Biol Med. 2018;14(3):977–990. doi:10.1016/j.nano.2018.01.014
- Yang Y, Tian Q, Wu S, et al. Blue light-triggered Fe2+-release from monodispersed ferrihydrite nanoparticles for cancer iron therapy. Biomaterials. 2021;271:120739. doi:10.1016/j.biomaterials.2021.120739
- Pazár B, Ea HK, Narayan S, et al. Basic Calcium Phosphate Crystals Induce Monocyte/Macrophage IL-1β Secretion through the NLRP3 Inflammasome In Vitro. J Immunol. 2011;186(4):2495–2502. doi:10.4049/jimmunol.1001284
- He XY, Liu BY, Xu C, Zhuo RX, Cheng SX. A multi-functional macrophage and tumor targeting gene delivery system for the regulation of macrophage polarity and reversal of cancer immunoresistance. Nanoscale. 2018;10(33):15578–15587. doi:10.1039/C8NR05294H
- Chang -C-C, Dinh TK, Lee Y-A, et al. Nanoparticle Delivery of MnO2 and Antiangiogenic Therapy to Overcome Hypoxia-Driven Tumor Escape and Suppress Hepatocellular Carcinoma. ACS Appl Mater Interfaces. 2020;12(40):44407–44419. doi:10.1021/acsami.0c08473
- Zhang X, Tang J, Li C, Lu Y, Cheng L, Liu J. A targeting black phosphorus nanoparticle based immune cells nano-regulator for photodynamic/photothermal and photo-immunotherapy. Bioact Mater. 2021;6(2):472–489. doi:10.1016/j.bioactmat.2020.08.024
- Yang L, Zhang Y. Tumor-associated macrophages: from basic research to clinical application. J Hematol Oncol. 2017;10(1):58. doi:10.1186/s13045-017-0430-2
- Nirmala MJ, Kizhuveetil U, Johnson A, B G, Nagarajan R, Muthuvijayan V. Cancer nanomedicine: a review of nano-therapeutics and challenges ahead. RSC Adv. 2023;13(13):8606–8629. doi:10.1039/D2RA07863E
- Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer statistics, 2022. CA Cancer J Clin. 2022;72(1):7–33. doi:10.3322/caac.21708
- Mantovani A, Marchesi F, Malesci A, Laghi L, Allavena P. Tumour-associated macrophages as treatment targets in oncology. Nat Rev Clin Oncol. 2017;14(7):399–416. doi:10.1038/nrclinonc.2016.217
- Noy R, Pollard JW. Tumor-Associated Macrophages: from Mechanisms to Therapy. Immunity. 2014;41(1):49–61. doi:10.1016/j.immuni.2014.06.010
- Gustafson HH, Pun SH. Instructing macrophages to fight cancer. Nat Biomed Eng. 2018;2(8):559–561. doi:10.1038/s41551-018-0276-0
- Andón FT, Digifico E, Maeda A, et al. Targeting tumor associated macrophages: the new challenge for nanomedicine. Semin Immunol. 2017;34:103–113. doi:10.1016/j.smim.2017.09.004
- DeNardo DG, Ruffell B. Macrophages as regulators of tumour immunity and immunotherapy. Nat Rev Immunol. 2019;19(6):369–382. doi:10.1038/s41577-019-0127-6
- Leuschner F, Courties G, Dutta P, et al. Silencing of CCR2 in myocarditis. Eur Heart J. 2015;36(23):1478–1488. doi:10.1093/eurheartj/ehu225
- Hughes R, Qian BZ, Rowan C, et al. Perivascular M2 Macrophages Stimulate Tumor Relapse after Chemotherapy. Cancer Res. 2015;75(17):3479–3491. doi:10.1158/0008-5472.CAN-14-3587
- Yang Z, Li H, Wang W, et al. CCL2/CCR2 Axis Promotes the Progression of Salivary Adenoid Cystic Carcinoma via Recruiting and Reprogramming the Tumor-Associated Macrophages. Front Oncol. 2019;9:231. doi:10.3389/fonc.2019.00231
- Xu X, Li R, Dong R, et al. In vitro characterization and cellular uptake profiles of TAMs-targeted lipid calcium carbonate nanoparticles for cancer immunotherapy. Acta Mater Medica. 2022;1(3). doi:10.15212/AMM-2022-0030
- Barclay AN, van den Berg TK. The Interaction Between Signal Regulatory Protein Alpha (SIRPα) and CD47: structure, Function, and Therapeutic Target. Annu Rev Immunol. 2014;32(1):25–50. doi:10.1146/annurev-immunol-032713-120142
- Lin ZP, Nguyen LNM, Ouyang B, et al. Macrophages Actively Transport Nanoparticles in Tumors After Extravasation. ACS Nano. 2022;16(4):6080–6092. doi:10.1021/acsnano.1c11578
- Shime H, Matsumoto M, Oshiumi H, et al. Toll-like receptor 3 signaling converts tumor-supporting myeloid cells to tumoricidal effectors. Proc Natl Acad Sci. 2012;109(6):2066–2071. doi:10.1073/pnas.1113099109
- Shang Y, Lu H, Liao L, Li S, Xiong H, Yao J. Bioengineered Nanospores Selectively Blocking LC3-Associated Phagocytosis in Tumor-Associated Macrophages Potentiate Antitumor Immunity. ACS Nano. 2023;17(11):10872–10887. doi:10.1021/acsnano.3c02657
- Pan XF, Wang L, Pan A. Epidemiology and determinants of obesity in China. Lancet Diabetes Endocrinol. 2021;9(6):373–392. doi:10.1016/S2213-8587(21)00045-0
- Hernandez ED, Lee SJ, Kim JY, et al. A macrophage NBR1-MEKK3 complex triggers JNK-mediated adipose tissue inflammation in obesity. Cell Metab. 2014;20(3):499–511. doi:10.1016/j.cmet.2014.06.008
- Lumeng CN, Bodzin JL, Saltiel AR. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J Clin Invest. 2007;117(1):175–184. doi:10.1172/JCI29881
- Su Y, Wang W, Xiao Q, et al. Macrophage membrane-camouflaged lipoprotein nanoparticles for effective obesity treatment based on a sustainable self-reinforcement strategy. Acta Biomater. 2022;152:519–531. doi:10.1016/j.actbio.2022.08.055
- Zou W, Rohatgi N, Brestoff JR, et al. Myeloid-specific Asxl2 deletion limits diet-induced obesity by regulating energy expenditure. J Clin Invest. 2020;130(5):2644–2656. doi:10.1172/JCI128687
- Ma L, Liu TW, Wallig MA, et al. Efficient Targeting of Adipose Tissue Macrophages in Obesity with Polysaccharide Nanocarriers. ACS Nano. 2016;10(7):6952–6962. doi:10.1021/acsnano.6b02878
- Soccio RE. Galectin-3 in NAFLD: therapeutic Target or Noncausal Biomarker? J Clin Endocrinol Metab. 2021;106(9):e3773–e3774. doi:10.1210/clinem/dgab363
- Devisscher L, Verhelst X, Colle I, Van Vlierberghe H, Geerts A. The role of macrophages in obesity-driven chronic liver disease. J Leukoc Biol. 2016;99(5):693–698. doi:10.1189/jlb.5RU0116-016R
- Paré A, Mailhot B, Lévesque SA, et al. IL-1β enables CNS access to CCR2 hi monocytes and the generation of pathogenic cells through GM-CSF released by CNS endothelial cells. Proc Natl Acad Sci. 2018;115(6). doi:10.1073/pnas.1714948115
- Stöger JL, Gijbels MJJ, van der Velden S, et al. Distribution of macrophage polarization markers in human atherosclerosis. Atherosclerosis. 2012;225(2):461–468. doi:10.1016/j.atherosclerosis.2012.09.013
- Jenne CN, Kubes P. Immune surveillance by the liver. Nat Immunol. 2013;14(10):996–1006. doi:10.1038/ni.2691
- Tosello-Trampont AC, Landes SG, Nguyen V, Novobrantseva TI, Hahn YS. Kuppfer Cells Trigger Nonalcoholic Steatohepatitis Development in Diet-induced Mouse Model through Tumor Necrosis Factor-α Production. J Biol Chem. 2012;287(48):40161–40172. doi:10.1074/jbc.M112.417014
- Ngo W, Ahmed S, Blackadar C, et al. Why nanoparticles prefer liver macrophage cell uptake in vivo. Adv Drug Deliv Rev. 2022;185:114238. doi:10.1016/j.addr.2022.114238
- Ouyang B, Poon W, Zhang YN, et al. The dose threshold for nanoparticle tumour delivery. Nat Mater. 2020;19(12):1362–1371. doi:10.1038/s41563-020-0755-z
- Kurniawan DW, Jajoriya AK, Dhawan G, et al. Therapeutic inhibition of spleen tyrosine kinase in inflammatory macrophages using PLGA nanoparticles for the treatment of non-alcoholic steatohepatitis. J Controlled Release. 2018;288:227–238. doi:10.1016/j.jconrel.2018.09.004
- Maradana MR, Yekollu SK, Zeng B, et al. Immunomodulatory liposomes targeting liver macrophages arrest progression of nonalcoholic steatohepatitis. Metabolism. 2018;78:80–94. doi:10.1016/j.metabol.2017.09.002
- Martínez–Sánchez C, Bassegoda O, Deng H, et al. Therapeutic targeting of adipose tissue macrophages ameliorates liver fibrosis in non-alcoholic fatty liver disease. JHEP Rep. 2023;5(10):100830. doi:10.1016/j.jhepr.2023.100830
- Maeda H, Ishima Y, Saruwatari J, et al. Nitric oxide facilitates the targeting Kupffer cells of a nano-antioxidant for the treatment of NASH. J Controlled Release. 2022;341:457–474. doi:10.1016/j.jconrel.2021.11.039
- Song M, Schuschke DA, Zhou Z, et al. Kupffer cell depletion protects against the steatosis, but not the liver damage, induced by marginal-copper, high-fructose diet in male rats. Am J Physiol-Gastrointest Liver Physiol. 2015;308(11):G934–G945. doi:10.1152/ajpgi.00285.2014
- Traber PG, Zomer E. Therapy of Experimental NASH and Fibrosis with Galectin Inhibitors. PLoS One. 2013;8(12):e83481. doi:10.1371/journal.pone.0083481
- Akram MS, Pery N, Butler L, et al. Challenges for biosimilars: focus on rheumatoid arthritis. Crit Rev Biotechnol. 2021;41(1):121–153. doi:10.1080/07388551.2020.1830746
- Ye L, Wen Z, Li Y, et al. Interleukin-10 attenuation of collagen-induced arthritis is associated with suppression of interleukin-17 and retinoid-related orphan receptor γt production in macrophages and repression of classically activated macrophages. Arthritis Res Ther. 2014;16(2):R96. doi:10.1186/ar4544
- Cutolo M, Campitiello R, Gotelli E, Soldano S. The Role of M1/M2 Macrophage Polarization in Rheumatoid Arthritis Synovitis. Front Immunol. 2022;13:867260. doi:10.3389/fimmu.2022.867260
- Li S, Su J, Cai W, Liu JX. Nanomaterials Manipulate Macrophages for Rheumatoid Arthritis Treatment. Front Pharmacol. 2021;12:699245. doi:10.3389/fphar.2021.699245
- Nasra S, Bhatia D, Kumar A. Recent advances in nanoparticle-based drug delivery systems for rheumatoid arthritis treatment. Nanoscale Adv. 2022;4(17):3479–3494. doi:10.1039/D2NA00229A
- Gomez-Barrena E, Lindroos L, Ceponis A, et al. Cartilage oligomeric matrix protein (COMP) is modified by intra-articular liposomal clodronate in an experimental model of arthritis. Clin Exp Rheumatol. 2006;24(6):622–628.
- Jain S, Tran TH, Amiji M. Macrophage repolarization with targeted alginate nanoparticles containing IL-10 plasmid DNA for the treatment of experimental arthritis. Biomaterials. 2015;61:162–177. doi:10.1016/j.biomaterials.2015.05.028
- Yang Y, Guo L, Wang Z, et al. Targeted silver nanoparticles for rheumatoid arthritis therapy via macrophage apoptosis and Re-polarization. Biomaterials. 2021;264:120390. doi:10.1016/j.biomaterials.2020.120390
- Lu Y, Zhou J, Wang Q, et al. Glucocorticoid-loaded pH/ROS dual-responsive nanoparticles alleviate joint destruction by downregulating the NF-κB signaling pathway. Acta Biomater. 2023;164:458–473. doi:10.1016/j.actbio.2023.04.012
- Li H, Feng Y, Zheng X, et al. M2-type exosomes nanoparticles for rheumatoid arthritis therapy via macrophage re-polarization. J Controlled Release. 2022;341:16–30. doi:10.1016/j.jconrel.2021.11.019
- Moore KJ, Sheedy FJ, Fisher EA. Macrophages in atherosclerosis: a dynamic balance. Nat Rev Immunol. 2013;13(10):709–721. doi:10.1038/nri3520
- Peled M, Fisher EA. Dynamic Aspects of Macrophage Polarization during Atherosclerosis Progression and Regression. Front Immunol. 2014;5. doi:10.3389/fimmu.2014.00579
- Wang Y, Li L, Zhao W, et al. Targeted Therapy of Atherosclerosis by a Broad-Spectrum Reactive Oxygen Species Scavenging Nanoparticle with Intrinsic Anti-inflammatory Activity. ACS Nano. 2018;12(9):8943–8960. doi:10.1021/acsnano.8b02037
- Jiang C, Qi Z, Tang Y, et al. Rational Design of Lovastatin-Loaded Spherical Reconstituted High Density Lipoprotein for Efficient and Safe Anti-Atherosclerotic Therapy. Mol Pharm. 2019;16(7):3284–3291. doi:10.1021/acs.molpharmaceut.9b00445
- Beldman TJ, Senders ML, Alaarg A, et al. Hyaluronan Nanoparticles Selectively Target Plaque-Associated Macrophages and Improve Plaque Stability in Atherosclerosis. ACS Nano. 2017;11(6):5785–5799. doi:10.1021/acsnano.7b01385
- Yang Q, Jiang H, Wang Y, et al. Plaque Macrophage‐Targeting Nanosystems with Cooperative Co‐Regulation of ROS and TRAF6 for Stabilization of Atherosclerotic Plaques. Adv Funct Mater. 2023;33(28):2301053. doi:10.1002/adfm.202301053
- Ouimet M, Ediriweera HN, Gundra UM, et al. MicroRNA-33–dependent regulation of macrophage metabolism directs immune cell polarization in atherosclerosis. J Clin Invest. 2015;125(12):4334–4348. doi:10.1172/JCI81676
- He J, Zhang W, Zhou X, et al. Reactive oxygen species (ROS)-responsive size-reducible nanoassemblies for deeper atherosclerotic plaque penetration and enhanced macrophage-targeted drug delivery. Bioact Mater. 2023;19:115–126. doi:10.1016/j.bioactmat.2022.03.041
- Gao C, Huang Q, Liu C, et al. Treatment of atherosclerosis by macrophage-biomimetic nanoparticles via targeted pharmacotherapy and sequestration of proinflammatory cytokines. Nat Commun. 2020;11(1):2622. doi:10.1038/s41467-020-16439-7
- Wang Y, Zhang K, Li T, et al. Macrophage membrane functionalized biomimetic nanoparticles for targeted anti-atherosclerosis applications. Theranostics. 2021;11(1):164–180. doi:10.7150/thno.47841
- Hu R, Dai C, Dong C, et al. Living Macrophage-Delivered Tetrapod PdH Nanoenzyme for Targeted Atherosclerosis Management by ROS Scavenging, Hydrogen Anti-inflammation, and Autophagy Activation. ACS Nano. 2022;16(10):15959–15976. doi:10.1021/acsnano.2c03422
- Cordes T, Wallace M, Michelucci A, et al. Immunoresponsive Gene 1 and Itaconate Inhibit Succinate Dehydrogenase to Modulate Intracellular Succinate Levels. J Biol Chem. 2016;291(27):14274–14284. doi:10.1074/jbc.M115.685792
- Na YR, Stakenborg M, Seok SH, Matteoli G. Macrophages in intestinal inflammation and resolution: a potential therapeutic target in IBD. Nat Rev Gastroenterol Hepatol. 2019;16(9):531–543. doi:10.1038/s41575-019-0172-4
- Ma WT, Gao F, Gu K, Chen DK. The Role of Monocytes and Macrophages in Autoimmune Diseases: a Comprehensive Review. Front Immunol. 2019;10:1140. doi:10.3389/fimmu.2019.01140
- Haribhai D, Ziegelbauer J, Jia S, et al. Alternatively Activated Macrophages Boost Induced Regulatory T and Th17 Cell Responses during Immunotherapy for Colitis. J Immunol. 2016;196(8):3305–3317. doi:10.4049/jimmunol.1501956
- Lissner D, Schumann M, Batra A, et al. Monocyte and M1 Macrophage-induced Barrier Defect Contributes to Chronic Intestinal Inflammation in IBD. Inflamm Bowel Dis. 2015:1. doi:10.1097/MIB.0000000000000384
- Zhu Y, Li X, Chen J, et al. The pentacyclic triterpene Lupeol switches M1 macrophages to M2 and ameliorates experimental inflammatory bowel disease. Int Immunopharmacol. 2016;30:74–84. doi:10.1016/j.intimp.2015.11.031
- Song WJ, Li Q, Ryu MO, et al. TSG-6 Secreted by Human Adipose Tissue-derived Mesenchymal Stem Cells Ameliorates DSS-induced colitis by Inducing M2 Macrophage Polarization in Mice. Sci Rep. 2017;7(1):5187. doi:10.1038/s41598-017-04766-7
- Seo DH, Che X, Kwak MS, et al. Interleukin-33 regulates intestinal inflammation by modulating macrophages in inflammatory bowel disease. Sci Rep. 2017;7(1):851. doi:10.1038/s41598-017-00840-2
- Huang Z, Gan J, Jia L, et al. An orally administrated nucleotide-delivery vehicle targeting colonic macrophages for the treatment of inflammatory bowel disease. Biomaterials. 2015;48:26–36. doi:10.1016/j.biomaterials.2015.01.013
- Tannahill GM, Curtis AM, Adamik J, et al. Succinate is an inflammatory signal that induces IL-1β through HIF-1α. Nature. 2013;496(7444):238–242. doi:10.1038/nature11986
- Hayashi M, Sakata M, Takeda T, et al. Induction of glucose transporter 1 expression through hypoxia-inducible factor 1α under hypoxic conditions in trophoblast-derived cells. J Endocrinol. 2004;183(1):145–154. doi:10.1677/joe.1.05599
- Herfarth H. IL-10 therapy in Crohn’s disease: at the crossroads. Gut. 2002;50(2):146–147. doi:10.1136/gut.50.2.146
- Bae YH, Park K. Targeted drug delivery to tumors: myths, reality and possibility. J Controlled Release. 2011;153(3):198–205. doi:10.1016/j.jconrel.2011.06.001