
Abstract
Breast cancer is a leading killer of women worldwide. Cyclodextrin-based estrogen receptor-targeting drug-delivery systems represent a promising direction in cancer therapy but have rarely been investigated. To seek new targeting therapies for membrane estrogen receptor-positive breast cancer, an estrogen-anchored cyclodextrin encapsulating a doxorubicin derivative Ada-DOX (CDE1-Ada-DOX) has been synthesized and evaluated in human breast cancer MCF-7 cells. First, we synthesized estrone-conjugated cyclodextrin (CDE1), which formed the complex CDE1-Ada-DOX via molecular recognition with the derivative adamantane-doxorubicin (Ada-DOX) (Kd =1,617 M−1). The structure of the targeting vector CDE1 was fully characterized using 1H- and 13C-nuclear magnetic resonance, mass spectrometry, and electron microscopy. CDE1-Ada-DOX showed two-phase drug-release kinetics with much slower release than Ada-DOX. The fluorescence polarization analysis reveals that CDE1-Ada-DOX binds to recombinant human estrogen receptor α fragments with a Kd of 0.027 µM. Competition assay of the drug complex with estrogen ligands demonstrated that estrone and tamoxifen competed with CDE1-Ada-DOX for membrane estrogen receptor binding in MCF-7 cells. Intermolecular self-assembly of CDE1 molecules were observed, showing tail-in-bucket and wire-like structures confirmed by transmission electronic microscopy. CDE1-Ada-DOX had an unexpected lower drug uptake (when the host–guest ratio was >1) than non-targeting drugs in MCF-7 cells due to ensconced ligands in cyclodextrins cavities resulting from the intermolecular self-assembly. The uptake of CDE1-Ada-DOX was significantly increased when the host–guest ratio was adjusted to be less than half at the concentration of CDE1 over 5 µM due to the release of the estrone residues. CDE1 elicited rapid activation of mitogen-activated protein kinases (p44/42 MAPK, Erk1/2) in minutes through phosphorylation of Thr202/Tyr204 in MCF-7 cells. These results demonstrate a targeted therapeutics delivery of CDE1-Ada-DOX to breast cancer cells in a controlled manner and that the drug vector CDE1 can potentially be employed as a molecular tool to differentiate nongenomic from genomic mechanism.
Introduction
The targeted drug-delivery systems formed through various types of intermolecular forces have attracted great interest because of their therapeutic potential in drug development and cancer treatment.Citation1,Citation2 Many elegant drug-delivery systems based on such non-bonding host–guest interactions have been developed.Citation3–Citation6 Cyclodextrins (CDs) are among the best nonimmunogenic vector candidates for self-assembly of targeted drug-delivery systemsCitation7,Citation8 due to their unique hydrophobic hollow-cavity-containing structure, excellent biocompatibility, chemical modifiability of hydroxyl groups in the primary face, and strong ability to entrap drug molecules through molecular recognition.Citation7–Citation9 Ligand-attached CDs facilitate drug delivery to tumor cells that often express abundant target receptors on the tumor cell membrane. For example, multivalent CD-based glycoclusters specifically target clinically relevant sugar receptors;Citation10 a recently synthesized series of drug complexes based on CDs functionalized with folic acid, arginylglycylaspartic acid (RGD tripeptide), and hyaluronic acid, in which hydrophobic therapeutics were entrapped, have been found to possess higher targeting efficiency and better drug affinity to target receptor-positive cancer cells (data from arginylglycylaspartic acid- and hyaluronic acid-conjugated CDs are unpublished).Citation11 Ligand-attached CDs represent promising multimodal drug-delivery vectors.
However, CD-based estrogen-anchored therapeutic delivery systems have never been investigated, even though estrogens are crucial in the initiation, development, and progression of human breast cancer.Citation12–Citation14 Overexpression of estrogen receptors (ERs) is observed in approximately 70% of human breast cancers.Citation12,Citation15 In addition to residing in the nucleus (“nuclear ERs” [nERs], ie, ERα and ERβ), ERs can also reside on the membrane and endoplasmic reticulum (so-called “membrane ERs” [mERs]).Citation16–Citation19 Targeting the overexpressed mERs for the delivery of therapeutics represents a promising and effectual strategy for breast cancer therapy.Citation20–Citation22 In addition, the estrogen-conjugated CD vector is expected to preserve and elicit the estrogen response and act as a new antiestrogen for additional adjuvant endocrine treatments of ER-dependent breast cancer via targeting the ER-dependent pathways besides the targeting drug-delivery modality.Citation21
Herein, we report an mER targeting estrone (E1)-conjugated CD vector CDE1 and prepared drug-delivery complex CDE1-Ada-DOX () through molecular recognition of a doxorubicin derivative adamantane-doxorubicin (Ada-DOX) and CDE1. In this system, E1 was first conjugated to CDs (CDE1) to vectorize the encapsulated drug Ada-DOX which comprises an adamantane (Ada) molecule covalently linked to doxorubicin.Citation11 The molecular moiety of adamantine forms stable drug inclusion with CDs with high affinity and low cytotoxicity.Citation23,Citation24 The drug uptake of CDE1-Ada-DOX and non-targeting CD-Ada-DOX have been examined in time course altered with host–guest ratios; notably, the drug release and uptake of Ada-DOX from CDE1-Ada-DOX in MCF-7 cells was in a controllable manner due to the unique intermolecular self-assembly of and the resulting stealth or release of the E1 residues in CDE1. Finally, we investigated whether the synthesized “estrogen-like” CDE1 could activate nonclassical rapid response through activated ER-mediated pathway in MCF-7 cells, and the potential application in differentiation of genomic and nongenomic pathways.Citation25–Citation27
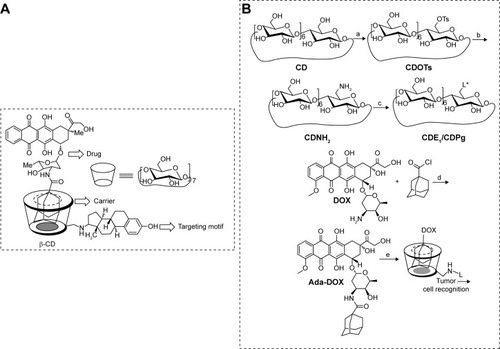
Figure 1 Schematic of estrone targeting vector based on cyclodextrin.
Notes: (A) The chemical structure of the CDE1-Ada-DOX inclusion complex. (B) Reaction scheme for the synthesis of ligand-attached cyclodextrin vectors (CDE1 and CDPg), intermediates (CDOTs, CDNH2Citation28), and mER-targeting supramolecule CDE1-Ada-DOX. (a) Dissolve β-CD in H2O at the presence of NaOH (3eq) followed by adding acetonitrile solution of p-TsCl (1 eq), react for 6 hours at rt. (b) (1) Neutralized by adding 2 eq of HCl, followed by recrystallization with cold acetone; (2) React in NH3·H2O for 3–7 days at 70°C. (c) (1) React in pyridine (or DMF) for 48 hours with ligand (2) excess NaBH4 in methanol solution. (d) React in anhydrous CH2Cl2 at presence of Et3N and N2 overnight at rt. (e) Molecular recognition with CDE1 or CDPg. L* interpreted as: ligand as estrone (E1); progesterone (Pg). The CDE1-Ada-DOX inclusion complex is a supermolecule generated though hydrophobic interaction and molecular recognition.
Abbreviations: CDE1, estrone-conjugated cyclodextrin; CDNH2, mono-6-deoxy-6-aminob-cyclodextrin; CDOTs, mono-6-deoxy-6-(p-tolylsulfonyl)-b-cyclodextrin; CDPg, progesterone-conjugated cyclodextrin; mER, membrane estrogen receptor; Ada-Dox, adamantane-doxorubicin; rt, room temperature.
Methods
For chemical synthesis, characterization, drug release, TEM microscopy, drug uptake, and Western blot, see Supplementary materials.
Binding affinity of CDE1 with Ada-DOX in aqueous solution determined using the fluorescence titration method
We next determined how strong the host molecule CDE1 interacted with the guest molecule Ada-DOX using the fluorescence titration method as described previously.Citation29 The effect of varying CDE1 concentrations on the fluorescence intensity of Ada-DOX was examined in order to determine the association constant (Kd) between CDE1 and Ada-DOX. The concentration of Ada-DOX was set at 50 µM in the presence of CDE1 at escalating concentrations of 0, 0.17, 0.26, 0.35, 0.44, 0.53, 0.62, 0.71, 0.79, and 0.88 mM, and the fluorescence intensity was monitored using a Synergy™ H4 Hybrid Microplate Reader (BioTek Inc., Winooski, VT, USA). The solvent used was dimethylformamide:H2O (50/50, v/v). The fluorescence of Ada-DOX was measured with λex at 490 nm and λem ranging from 500 to 700 nm with an escalating step of 2 nm. The Kd value was calculated using the above-mentioned approach by nonlinear fitting for various models.
Effects of the phosphorylation of p44/42 mitogen-activated protein kinase (Erk1/2) at Thr202/Tyr204 in MCF-7 cells
To examine whether CDE1 elicited nongenomic events in MCF-7 cells, the cells were treated with CDE1 or other drugs at 1 µM for 5, 10, 15, 30, and 60 minutes to detect p44/42 mitogen-activated protein kinase (MAPK) (Erk1/2) phosphorylation at Thr202/Tyr204 using Western blotting analysis in MCF-7 cells. The experiments were repeated in triplicate.
Binding determination of the CDE1-Ada-DOX inclusion complex to recombinant human ERα fragments using the fluorescence polarization method
Fluorescence polarization (also called fluorescence anisotropy) is a versatile solution-based technique that has been widely used to investigate molecular interactions (eg, ligand– receptor binding), enzymatic activity, and nucleic acid hybridization.Citation48 Quantitatively, fluorescence polarization (mP) is defined as the difference of the emission light intensity parallel and perpendicular to the excitation light plane normalized by the total fluorescence emission intensity. The binding of the CDE1-Ada-DOX inclusion complex to recombinant human ERα fragments consisting of amino acid residues 1–116 at the C-terminus (His tag C-terminus, Molecular Mass =12,200 Da) (catalogue number: ab153776; Abcam Plc, Cambridge, UK) was investigated using the fluorescence polarization method as described previously.Citation31 Briefly, human ERα fragments were reconstituted in phosphate-buffered saline to the final concentration of 0.8 µM, and CDE1-Ada-DOX complex samples at concentrations from 0.04 µM to 1.26 µM were added to the protein solution. The samples were mixed well at room temperature and subject to analysis immediately. mP was measured using the Synergy H4 Hybrid Multi-Mode Microplate reader at λEx =485/20 nm and λEm =620/10 nm with xenon flash as the light source. The acquisition parameters were set as follows: 200 flashes and positioning delay 100 msec. The experiment was repeated in triplicate. The mP values were plotted against an increasing concentration of CDE1-Ada-DOX and the equilibrium binding constant (Kd) was calculated using nonlinear least squares that fit the curve data by Prism 6.03 program (GraphPad Software, Inc., La Jolla, CA, USA).
Results and discussion
In this study, E1 was successfully conjugated with β-CD to generate CDE1 as a new drug vector to target mERs of breast cancer cells. At the same time, progesterone, having a similar structure to E1, was also conjugated to CDs as the substrate extension of the conjugation reaction (). CDE1 accommodates the hydrophobic Ada-DOX through host–guest interactions to form the CDE1-Ada-DOX supramolecule for drug delivery since the geometry and hydrophobicity of the adamantyl group allows an excellent fit into the CD’s torus inner cavity. The structures of the synthetic compounds have been confirmed using multiple spectral methods which include 1H nuclear magnetic resonance, 13C nuclear magnetic resonance, high-resolution matrix-assisted laser desorption/ionization-time of flight mass spectroscopy, fluorescence spectroscopy, and circular dichroism spectroscopy (). The 1:1 stoichiometric inclusion complex CDE1-Ada-DOX was readily prepared by the coprecipitation method through molecular recognition.
Figure 2 Characterization of CDE1.
Notes: (A) The 1H-NMR spectrum of CDE1 (800 MHz, d4-DMSO, temperature =298 K). (B) The HR-MALDI-TOF spectrum of CDE1-Ada-DOX. (C) Fluorescence emission spectra of CDE1-Ada-DOX (ConAda-DOX =50 µM) in DMSO at different CDE1 concentrations of 0.17, 0.26, 0.35, 0.44, 0.53, 0.62, 0.71, 0.79, and 0.88 mM, with increasing step of 0.08 mM. (D) Circular dichroism spectra of CDE1-Ada-DOX. Samples in fluorescence measurements are excited at λex =485 nm.*103 indicates the y-axis value needs to be multiplied by 1,000.
Abbreviations: CDE1, estrone-conjugated cyclodextrin; DMSO, dimethyl sulfoxide; HR-MALDI-TOF, high-resolution matrix-assisted laser desorption/ionization-time of flight; NMR, nuclear magnetic resonance; Ada-Dox, adamantane-doxorubicin.
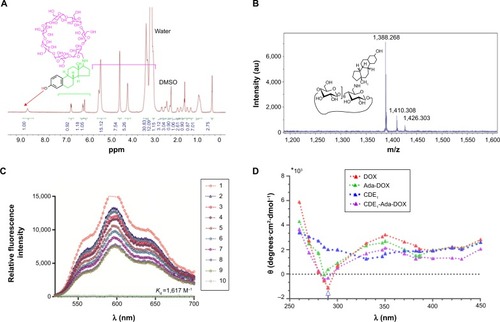
The host–guest binding affinity of CDE1 with Ada-DOX (Kd) was measured using the fluorescence titration method as described previously.Citation29 With an increasing amount of CDE1 added to the dimethylformamide and H2O (1:1) solution containing Ada-DOX, the fluorescence intensity was measured at varying emission wavelengths from 500 to 700 nm with 600 nm as the maximum peak, which decreased in a concentration-dependent manner (). The Kd values were calculated using the fluorescence intensity versus CDE1 concentration curve. This data showed that the one-phase and two-phase exponential decay models were the best fit. The calculated Kd was 0.93, 1.09, 0.77, and 0.73 mM when the emission wavelengths were 550, 574, 600, and 634 nm, respectively. The fluorescence quenching effect during the titration of CDE1 to Ada-DOX indicated that host–guest binding occurs.
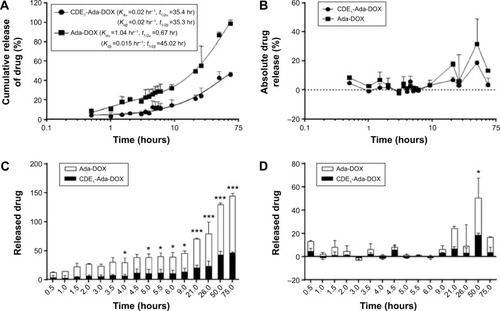
Moreover, we examined the releasing kinetics of Ada-DOX from the CDE1-Ada-DOX inclusion complex in phosphate-buffered saline against 50% fetal bovine serum solution using a validated fluorescence method.Citation11 The release kinetics of Ada-DOX and CDE1-Ada-DOX from the formulations clearly demonstrated that the drug released from CDE1-Ada-DOX was significantly slower (>50%) than Ada-DOX within the incubation time period (). Detachment of Ada-DOX from the CDE1-Ada-DOX inclusion complex showed sustained releasing kinetics. By fitting the released drug data versus time, the kinetic parameters were calculated. For Ada-DOX, the Kdα and Kdβ were 1.04 and 0.015 hours−1, with elimination half-life (t1/2)α and t1/2β of 0.67 and 45.02 hours, respectively. For CDE1-Ada-DOX, the Kdα and Kdβ were 0.02 and 0.02 hours−1, with t1/2α and t1/2β of 35.40 and 35.30 hours, respectively. The accumulative and absolute released drug for Ada-DOX and CDE1-Ada-DOX were also compared using two-way analysis of variance. At 2 hours, the cumulated drug release was 6.5% and 20.5% for CDE1-Ada-DOX and Ada-DOX, respectively (P>0.05). At 4 hours, the values were almost unchanged (6.6% versus 23.5%, P<0.05). At 6 hours, the values were increased to 11.1% and 28.4%, respectively (P<0.05). At 21 hours, drug-release values were increased to 20.9% and 49.1%, respectively (P<0.001). The measured drug released was 46.1% for CDE1-Ada-DOX and 99.0% for Ada-DOX over 75 hours of dialysis (P<0.001). These data clearly show that the release of Ada-DOX from CDE1-Ada-DOX is sustained compared with Ada-DOX over 75 hours. The sustained release of Ada-DOX from the inclusion complex would facilitate continuous drug uptake and long-term proliferation inhibition of the cancer cells.
Figure 3 Drug release profile of CDE1-Ada-DOX.
Notes: (A) The cumulative drug-releasing profile of Ada-DOX and CDE1-Ada-DOX over 75 hours. The best fit for the released drug level over time curve is a two-phase association model and the Kdα, Kdβ, t1/2α, and t1/2β values were obtained. (B) The absolute drug releasing profile of Ada-DOX and CDE1-Ada-DOX over 75 hours. (C) The cumulative drug-releasing profile of Ada-DOX and CDE1-Ada-DOX over 75 hours. (D) The absolute drug-releasing profile of Ada-DOX and CDE1-Ada-DOX over 75 hours. The release of Ada-DOX was determined by a dialysis method against 50% fetal bovine serum and 50% phosphate buffered saline solution, and the released Ada-DOX was quantified using a microplate reader at λex =490 nm and λem =600 nm. A calibration curve was prepared using different concentrations of free Ada-DOX. *P<0.05, ***P<0.001. Data are presented as the mean ± SD from three independent experiments.
Abbreviations: CDE1, estrone-conjugated cyclodextrin; t1/2, elimination half-life; Ada-Dox, adamantane-doxorubicin.
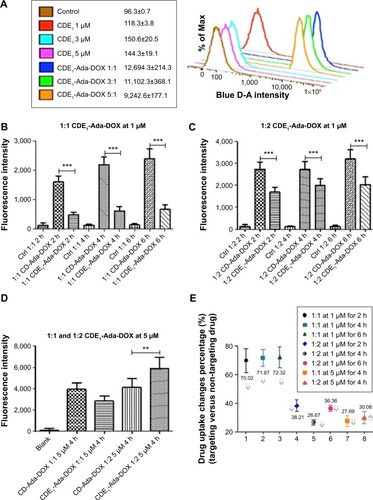
In addition to the sustained drug release of CDE1-Ada-DOX, it has been clearly demonstrated that anchored ligands in the drug-delivery systems providing specific drug–cell surface interactions are crucial in determining the ultimate drug internalization by cancer cells.Citation8 The cellular uptake of the mER-targeting drug complex CDE1-Ada-DOX and non-targeting compound CD-Ada-DOX was investigated to check for the targeting efficiency; flow cytometric analysis was performed in MCF-7 cells by taking advantage of intrinsic fluorescence emission from Ada-DOX. MCF-7 cells were treated with CDE1-Ada-DOX and non-targeting drug CD-Ada-DOX at different host–guest molecular molar ratios and drug concentrations in time course (). The control (MCF-7 cells with no drug treated) and CDE1 itself did not elicit obvious fluorescence and showed very low levels of autofluorescence. The addition of CDE1 at 1, 3, or 5 µM to the MCF-7 cells slightly increased the fluorescence. Interestingly, excess CDE1 quenched the fluorescence intensity of Ada-DOX at 1 µM when formulated in ratios of 1:1, 1:3, and 1:5 (Ada-DOX:CDE1) with 12.5% and 27.2% decrease for 1:3 and 1:5, respectively, in comparison to 1:1 CDE1-Ada-DOX complex (). CDE1-Ada-DOX had an unexpected lower cellular uptake than non-targeting CD-Ada-DOX complex in MCF-7 cells at 1 µM with a 1:1 host–guest molar ratio (), while the uptake of CDE1-Ada-DOX was enhanced when the host–guest molecule molar ratio and total concentration were altered. The cells were exposed to different CD or CDE1 inclusion complexes for 2, 4, and 6 hours, respectively, with host and guest molar ratio set at 1:1. The drug uptake in MCF-7 cells treated with CD-Ada-DOX with host–guest ratio of 1:1 (CDE1 concentration =1 µM) for 2, 4, and 6 hours was 3.34-, 3.55-, and 3.61-fold of that in cells treated with CDE1-Ada-DOX with host–guest ratio of 1:1 with CDE1 concentration at 1 µM, respectively (P<0.001). The internalization efficacy of the targeting complex CDE1-Ada-DOX rebounded greatly compared with the drug without ligand attached (CD-Ada-DOX) as the host–guest molecular molar ratio increased to 1:2 with CDE1 concentration of 1 µM (). The drug uptake in MCF-7 cells treated with CD-Ada-DOX with host–guest ratio of 1:2 at 1 µM of CDE1 for 2, 4, and 6 hours was 1.62-, 1.36-, and 1.57-fold of that in cells treated with CDE1-Ada-DOX with host–guest ratio of 1:1 (CDE1 concentration =1 µM), respectively (P<0.001). The drug uptake of CDE1-Ada-DOX exceeded that of CD-Ada-DOX when the concentration of the guest drug was continually raised. When the host–guest molecular ratio was altered to 1:2 with a CDE1 concentration of 5 µM, the drug uptake of MCF-7 cells treated with CDE1-Ada-DOX was higher (69.9%) than cells treated with CD-Ada-DOX (P<0.01) (). These results suggest that the targeted CDE1-Ada-DOX inclusion complex improved the uptake of Ada-DOX in comparison to the non-targeted CD-Ada-DOX inclusion complex when CDE1 and Ada-DOX were formulated in appropriate ratios and drug concentrations to release ample E1 ligands from CDE1.
Figure 4 Flow cytometric analysis of the drug uptake by MCF-7 cells after incubation with various drug formulations for 2–6 hours.
Notes: (A) The fluorescence intensity of various samples of CDE1 and CDE1-Ada-DOX with host–guest molar concentration ratio of 1:1, 3:1, and 5:1 at 1 µM of Ada-DOX in MCF-7 cells. The control is obtained by the autofluorescence from the MCF-7 cells without any drug exposure. (B–E) The drug uptake indicated as the fluorescence intensity of various samples of CDE1-Ada-DOX and the non-targeting drug complex CD-Ada-DOX with 1:1, 1:2, or 1:5 of host–guest molar concentration ratio in MCF-7 cells under CDE1 concentration of 1 or 5 µM. Data are presented as the mean ± standard deviation from three independent experiments. **P<0.01; ***P<0.001. The fluorescence is tested with Ex of 490 nm and Em of 600 nm.
Abbreviations: CD, cyclodextrin; CDE1, estrone-conjugated cyclodextrin; Ctrl, control; h, hours; Ada-Dox, adamantane-doxorubicin.
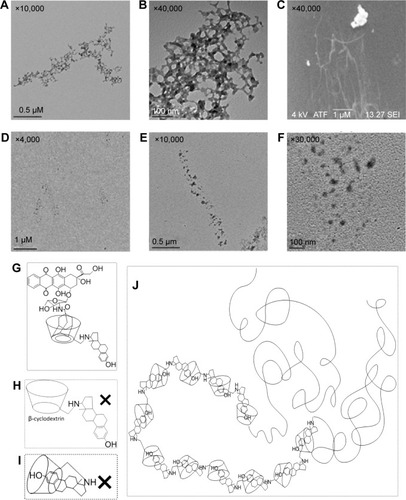
With higher guest drug concentrations or altered host–guest molecular molar ratios, the release of the targeting moieties from the complex CDE1-Ada-DOX have been enhanced through binding to the mERs and have consequently facilitated the drug internalization process in a controlled manner. These interesting findings led us to propose that estrogen residues covalently bonded with the CD are stealthy under certain circumstances. In order to further substantiate the presence of stealthy ligands in CDE1 and CDE1-Ada-DOX, transmission electronic microscopy (TEM) and scanning electronic microscopy (SEM) examinations were conducted. Representative TEM and SEM images of CDE1 and CDE1-Ada-DOX are shown in . The intermolecular assembly of CDE1 exhibited a tail-in-bucket structure and wire-like morphology for CDE1, which shows the conjugated estrogen residing inside the CD cavity of the adjacent CDE1 molecule. illustrate the long, tangled, uniform CDE1 wires. The estrogen residues in CDE1 act as linkers of CDE1 molecules due to intermolecular recognition. show the TEM images of CDE1-Ada-DOX under the same experimental conditions as CDE1. CDE1-Ada-DOX particles showed an unorganized structure under the same preparation conditions compared with the CDE1 containing no drug payload. The observation was consistent and reproducible in the TEM/SEM examinations. These findings indicate that the conjugated E1 residues of CDE1 were entrapped in the CD cavities of CDE1 in the absence of a guest molecule. Furthermore, when the guest molecule such as Ada-DOX was added into CDE1, the competition between exogenous and the intermolecular recognition with the CD cavity resulted in the consequent release of a certain amount of E1 residues, in turn disrupting intramolecular self-assembling, dissembling the wire-like and tail-in-bucket structure of CDE1. Moreover, circular dichroism analysis indicated that Ada-DOX interacted with CDE1 () and caused conformational changes at the CD cavity binding site followed by chiral microenvironment changes for the whole drug complex supramolecule. The circular dichroism spectra provide supportive evidence for the morphology differences of CDE1 and CDE1-Ada-DOX complexes.
Figure 5 The TEM/SEM characterization of CDE1 and CDE1-Ada-DOX.
Notes: A solution of CDE1 or CDE1-Ada-DOX at 1.8 mM in water:dimethylformamide (1:1, v/v) was dried in vacuum oven at 35°C overnight and then subject to TEM/SEM analysis. (A and B) The TEM image of CDE1 (magnification: ×10,000 for A; ×40,000 for B) shows the wire-like morphology for CDE1 due to self-assembly. The estrogen residue in CDE1 acts as the linker in intermolecular recognition. (C) The SEM image of CDE1 (magnification: ×40,000) shows the long, tangled, and uniform CDE1 wires resulting from self-assembly. (D–F) The TEM image of CDE1-Ada-DOX (magnification: ×4,000 for D; ×10,000 for E; ×30,000 for F). (G) The structure of CDE1-Ada-DOX. (H and I) Presumable structures of CDE1 which do not exist. (J) The tail-in-bucket and wire-like structure of CDE1 resulting from self-assembly, which were confirmed by TEM and SEM examinations.
Abbreviations: CDE1, estrone-conjugated cyclodextrin; SEM, scanning electron microscope; TEM, transmission electron microscope; Ada-Dox, adamantane-doxorubicin.
Herein, we consider that targeting mERs expressed on MCF-7 cells is an effective means of affecting the uptake of the drug complex into these cells since the drug complex is taken up by ER-mediated endocytosis. Our results showed that CDE1 preserved the binding ability after E1 conjugated to CD. More importantly, drug uptake can be augmented significantly if the estrogen molecules in the complex are released to ensure the estrogen residue tailed outside the CD cavity. The intermolecular recognition between the covalent attached estrogen residue of one CDE1 and the CD cavity of another CDE1 molecule results in the host–guest molar ratio-dependent difference in drug uptake since the targeting moiety has been entrapped, and in turn reduces or loses its affinity to mERs. When an appropriate guest molecule approaches the CDE1, the estrogen residues are pushed out and released. The competition between complexation of the drug and intermolecular inclusion of the estrogen in the CD cavity leads to altered drug uptake. In other words, the equilibrium of the host–guest molecules and the complex is the critical factor for controlling and optimizing the release kinetics of the targeting and drug moieties, mER binding, drug uptake efficiency, dissimilarity, and biological responses.
Furthermore, the cholesterol levels have been monitored in both cancerous cells (MCF-7 and A549 cells) and normal cells (MCF-10A and T80 cells) to investigate cholesterol depletion of lipid rafts on the cell membrane after drug exposure of CDE1 and the drug complexes, since CD derivatives have been reported to be able to extract cholesterol from bilayer membrane by the CDs cavity, and modulate the activity of multiple signaling pathways.Citation30 Cholesterol and estrogens have structural similarity. It was shown that the cholesterol level was not significantly affected after CDE1 treatment due to the preoccupancy of the CD cavity by E1 residues from the intramolecular self-assembly ().
Figure 6 Cholesterol depletion from the cell membrane of human breast cancer MCF-7 cells, human normal lung T-80 cells and human normal breast MCF-10A cells, human lung cancer A549 cells.
Notes: Cholesterol depletion from the cell membrane ([A] human breast cancer MCF-7 cells, [B] human normal breast MCF-10A cells, [C] human normal lung T-80 cells, [D] human lung cancer A549 cells) after the treatment with saturated CDE1 solution; natural ligand E1 (40 µM); positive control Methyl-CD and beta-CD; DOX at 0.5, 2.5 µM; AD at 10 and 100 µM; and the drug complex CDE1-Ada-DOX in different host–guest ratio of 1:1, 1:3, and 1:5, respectively, at the host concentration of 10 µM. Cell cultures were treated for 2 hours at 37°C to deplete cholesterol. Cells were then washed twice in serum-free media and lysed. Cholesterol levels were detected using EnzyChrom™ cholesterol assay kit (BioAssay Systems, Hayward, CA, USA). CDE 1/10 means the CDE1 concentration is 1/10 of the saturated CDE1 solution. *P<0.Z<0.01, **P<0.01, ***P<0.001. Data are presented as the mean ± SD from three independent experiments.
Abbreviations: AD, adamantane; beta-CD, β-cyclodextrin; CD, cyclodextrin; CDE1, estrone-conjugated cyclodextrin; Ctrl, control; DOX, doxorubicin; E1, estrone; Methyl-CD, methyl-β-cyclodextrin; NS, nonsignificant; Ada-Dox, adamantane-doxorubicin.
![Figure 6 Cholesterol depletion from the cell membrane of human breast cancer MCF-7 cells, human normal lung T-80 cells and human normal breast MCF-10A cells, human lung cancer A549 cells.Notes: Cholesterol depletion from the cell membrane ([A] human breast cancer MCF-7 cells, [B] human normal breast MCF-10A cells, [C] human normal lung T-80 cells, [D] human lung cancer A549 cells) after the treatment with saturated CDE1 solution; natural ligand E1 (40 µM); positive control Methyl-CD and beta-CD; DOX at 0.5, 2.5 µM; AD at 10 and 100 µM; and the drug complex CDE1-Ada-DOX in different host–guest ratio of 1:1, 1:3, and 1:5, respectively, at the host concentration of 10 µM. Cell cultures were treated for 2 hours at 37°C to deplete cholesterol. Cells were then washed twice in serum-free media and lysed. Cholesterol levels were detected using EnzyChrom™ cholesterol assay kit (BioAssay Systems, Hayward, CA, USA). CDE 1/10 means the CDE1 concentration is 1/10 of the saturated CDE1 solution. *P<0.Z<0.01, **P<0.01, ***P<0.001. Data are presented as the mean ± SD from three independent experiments.Abbreviations: AD, adamantane; beta-CD, β-cyclodextrin; CD, cyclodextrin; CDE1, estrone-conjugated cyclodextrin; Ctrl, control; DOX, doxorubicin; E1, estrone; Methyl-CD, methyl-β-cyclodextrin; NS, nonsignificant; Ada-Dox, adamantane-doxorubicin.](/cms/asset/10f266a9-e002-4cdb-848e-eddf41b654e6/dijn_a_82255_f0006_c.jpg)
Therefore, the magnitude of drug binding and internalization in cancer cells could be modulated by disrupting the intramolecular self-assembly of CDE1, and changing the drug exposure levels and composition (eg, host–guest molecular molar ratio) in a controllable manner based on our findings.
To confirm the binding ability of CDE1-Ada-DOX to mERs in breast cancer cells, we performed the competition assay with CDE1-Ada-DOX using E1 and a selective ER modulator, tamoxifen, as the inhibitors in MCF-7 cells. The results are shown in . The flow cytometric analysis showed that E1 at 5−50 µM inhibited CDE1-Ada-DOX uptake in a concentration-dependent manner in MCF-7 cells. E1 at 5, 10, and 50 µM diminished the uptake of CDE1-Ada-DOX by 5.1%, 4.3%, and 8.3%, respectively, in MCF-7 cells (P<0.001). Furthermore, tamoxifen also concentration-dependently reduced the uptake of CDE1-Ada-DOX in MCF-7 cells. Tamoxifen at 1, 5, and 10 µM inhibited the uptake of CDE1-Ada-DOX by 7.0%, 20.5%, and 19.2%, respectively, in MCF-7 cells (P<0.001). The results demonstrate that both E1 and tamoxifen competed with CDE1-Ada-DOX for mER binding in MCF-7 cells in a concentration-dependent manner with higher competing potency for tamoxifen compared to E1.
Figure 7 Competition of drug uptake and binding experiment about CDE1-Ada-DOX.
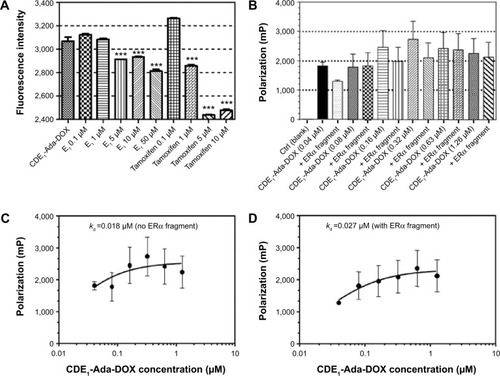
Notes: (A) The competition for the drug uptake of CDE1-Ada-DOX in the presence of E1 at 0.1–50 µM or tamoxifen at 0.1–10 µM in MCF-7 cells. Cells were incubated at 37°C with CDE1-Ada-DOX at 2 µM for 6 hours in the presence of E1 or tamoxifen at different concentrations. Data are presented as the mean ± SD from three independent experiments. ***P<0.001. (B) Binding of CDE1-Ada-DOX to recombinant human ERα fragments using the fluorescence polarization method. Recombinant human ERα fragments consisting of amino acid residues 1–116 at the C-terminus of ERα were reconstituted in phosphate buffered saline to the final concentration of 0.8 µM, and the CDE1-Ada-DOX inclusion complex samples at concentrations from 0.04 to 1.26 µM were added to the protein solution. The mP values were determined using a microplate reader at λEx =485/20 nm and λEm =620/10 nm. (C) The mP values were plotted against an increasing concentration of CDE1-Ada-DOX without adding human ERα fragments, and the Kd was calculated using nonlinear least squares fitting the curve data. The mP values were increased when the concentration of the CDE1-Ada-DOX inclusion complex was increased without adding the ERα fragments, with a Kd of 0.018 µM. (D) When the CDE1-Ada-DOX inclusion complex at escalating concentrations from 0.04 to 1.26 µM was mixed with human ERα fragments, the mP values were increased with a Kd of 0.027 µM. Data are the mean ± SD from three independent experiments.
Abbreviations: CDE1, estrone-conjugated cyclodextrin; Ctrl, control; E1, estrone; ER, estrogen receptor; SD, standard deviation; Ada-Dox, adamantane-doxorubicin.
Binding to mERs on tumor cells by tamoxifen or E1 prevented the entry of other ER ligands such as CDE1-Ada-DOX targeting complex to the binding pocket. These findings provide further evidence that estrogen residues from CDE1-Ada-DOX complex recognize and bind to mER/mERs on MCF-7 cells.
Additionally, we used a fragment of human ERα containing the ligand binding domain (recombinant human ERα fragments consisting of amino acid residues 1–116 at the C-terminus, His tag C-terminus, Mr =12,200 Da; Abcam Plc) to examine whether CDE1-Ada-DOX could bind to it using the fluorescence polarization approach.Citation31 The polarization (mP) data over the concentration of the CDE1-Ada-DOX inclusion complex in the absence or presence of human ERα fragments at a fixed concentration of 0.08 µM are shown in . The mP values were increased when the concentration of the CDE1-Ada-DOX inclusion complex was increased without adding the ERα fragments, with a Kd of 0.018 µM.
When the CDE1-Ada-DOX inclusion complex at escalating concentrations from 0.04 to 1.26 µM was mixed with human ERα fragments, the mP values were increased with a Kd of 0.027 µM. These results demonstrate the interaction of CDE1-Ada-DOX with human ERα fragments containing the ligand-binding domain.
In addition to the targeting drug-delivery modality of the novel “estrogen-like” molecule CDE1, the cellular response triggered by CDE1 in a manner different from the classical nER-mediated pathway was investigated by Western blotting assay (). Estrogens bind to nuclear ERα and ERβ, triggering the classical pathway of estrogen-dependent action and finally eliciting remarkable genomic responses. The action of nuclear ERs includes binding lipophilic hormone molecule in cytoplasm, translocation of the ligand–ER complex to the nucleus, dimerization, interaction with estrogen-specific response elements in the promoter areas of target genes, and finally initiating gene transcription.Citation18,Citation19,Citation32 The effects of steroid hormone action dependent on ER occur within hours or even days. On the other hand, some ligands can elicit rapid nongenomic signaling cascades in a much shorter time (from seconds to minutes) upon estrogen binding.Citation18,Citation19,Citation33,Citation34 These rapid nongenomic effects of estrogens result in calcium mobilization, cyclic adenosine monophosphate stimulation, phospholipase C activation, inositol phosphate generation, and activation of membrane-associated signaling pathways, including protein kinase A, phosphotidylinositol-3 kinase, and MAPK (p44/42 MAPK, also called extracellular-signal-regulated kinase [Erk1/2]) signaling pathways.Citation18,Citation19 Importantly, crosstalk via second messengers between mER- and nER-initiated signaling responses can regulate transcriptional activation of multiple target genes in a coordinated manner.Citation18,Citation19 The estrogens E1 and E2, the selective ER modulator tamoxifen, and the synthetic estrogen 17α-ethinyl estradiol were used as positive control to trigger the rapid response of Erk1/2; adamantane and 1-adamantanol were used as structural fraction controls to the drug complex CDE1-Ada-DOX. It was demonstrated that CDE1 resulted in rapid phosphorylation response of p44/42 MAPK (p-Erk1/2) within 30 minutes in ER-positive cancerous cells such as MCF-7 cells, A549 cells, OVCAR3 cells, and even in the normal lung T80 cells, while no significant response was observed in normal breast MCF-10A cells. Cells treated with CDE1 at 1 µM showed a maximum p44/p42 MAPK phosphorylation at Thr202/Tyr204 at 15 minutes drug exposure and increases by 22.2%, 25.5%, and 59.2% at 30 minutes for MCF-7, A549 and OVCAR3 cells respectively, compared with controls (P<0.05). These results show that treatment of MCF-7 cells with CDE1 activated the mER-mediated signaling pathway as indicated by the significantly increased phosphorylation of p44/42 MAPK (Erk1/2) rapidly within 5−30 minutes at Thr202/Tyr204 in MCF-7 cancer cells. Estrogens such as E2 induce a number of rapid signaling events in cells that express mERs such as GPR30,Citation19 except in classical pathways.Citation18,Citation19,Citation32 Different stimuli including mitogens, growth factors, cytokines, virus infection, ligands for heterotrimeric G protein-coupled receptors, transforming agents, and carcinogens can activate the p44/42 MAPK (Erk1/2) pathway.Citation35–Citation37 There is evidence that E2-induced p44/42 MAPK (Erk1/2) activation requires GPR30, and occurs via transactivation of the epidermal growth factor receptor.Citation38,Citation39 In the Raf–MEK–MAPK/Erk pathway, receptor tyrosine kinases and G protein-coupled receptors activate Ras, which in turn activates c-Raf.Citation35,Citation37 Activation of c-Raf involves phosphorylation at multiple residues including Ser338, Tyr341, Thr491, Ser494, Ser497, and Ser499. p21-activated protein kinase can phosphorylate c-Raf at Ser338 and the Src family phosphorylates Tyr341 of c-Raf.Citation35,Citation37 Activated c-Raf activates MAPK kinase (called MKK, MEK, or MAP2K) at Ser217/221 located in the activation loop of subdomain VIII, and MEK1/2 activate p44 and p42 through phosphorylation of activation loop residues Thr202/Tyr204 and Thr185/Tyr187, respectively.Citation37 p44/42 MAPK (Erk1/2) are negatively regulated by a family of dual-specificity (Thr/Tyr) MAPK phosphatases, along with MEK inhibitors such as U0126 and PD98059.Citation40
Figure 8 Western blot analysis of CDE1-Ada-DOX and its analogs on A549, MCF-7, OVCAR-3, T-80 and MCF-10A cells.
Notes: Western blot analysis shows the time course (0, 5, 10, 15, 30, and 60 minutes) of p44/42 MAPK (Erk1/2) phosphorylation at Thr202/Tyr204 by tamoxifen, Ada-DOX, 17α-ethinyl estradiol, CDE1-Ada-DOX, adamantane, E2, β-cyclodextrin, E1, and CDE1 at 1 µM in MCF-7, A549, and OVCAR cancerous cells (A); and in T-80 and MCF-10A human normal cells (B). In the experiments, Western immunoblotting was performed using rabbit antibodies against p-p44/42 MAPK (Erk1/2) at Thr202/Tyr204 and β-actin was used as the internal control. Data are from three independent experiments. Data are presented as the mean ± SD from three independent experiments.
Abbreviations: CDE1, estrone-conjugated cyclodextrin; E1, estrone; E2, estradiol; Ada-Dox, adamantane-doxorubicin.
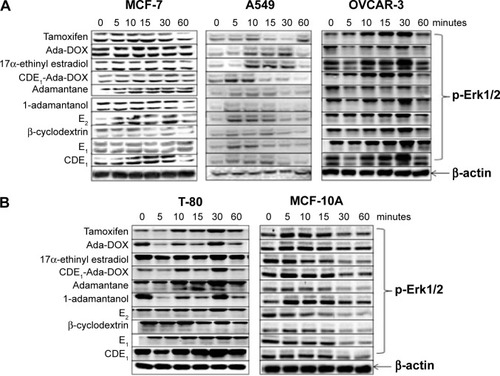
In this study, treatment of MCF-7 cells with the new synthetic CDE1 resulted in rapid phosphorylation of p44/42 MAPK (Erk1/2) in minutes. These findings suggest that CDE1 can interact with and activate mERs and might be used as a probe for studying mER-mediated nongenomic events in comparison with nER-mediated genomic responses that involve DNA binding and gene expression initiation; it also needs a much longer time. It was demonstrated that adamantane and 1-adamantanol molecules do not affect p44/42 MAPK phosphorylation in tumor cells significantly, while upregulation of p-Erk1/2 from the drug complex CDE1-Ada-DOX results from CDE1 and partially from the Ada-DOX since Ada-DOX treatment also cause slight pErk1/2 over-expression.
It is well known that estrogens activate the rapid, non-classical signaling cascades via mERs, and there is crosstalk between mER-mediated nonclassical and nER-mediated classical pathways.Citation39,Citation41,Citation42 Deregulation of both pathways plays important roles in the pathogenesis of cancer and other diseases.Citation43 It is recognized that selective ER ligands are needed to delineate the role of extranuclear and nuclear ERs in disease development and therapeutics where ERs represent important therapeutic targets. Several estrogen conjugates, including CD–E2, E2–BSA, E2–peroxidase, and E2–dendrimers, have been reported and used to probe the nonclassical and classical cascades.Citation44–Citation47 CD derivatives are generally considered cell-membrane impermeable. Therefore, CDE1 can be employed as a molecular tool to differentiate nongenomic response from genomic response based on the findings.
Conclusion
We synthesized the novel estrogen-anchored conjugate CDE1 and the corresponding drug inclusion with the doxorubicin derivative Ada-DOX (CDE1-Ada-DOX). The structures of these new compounds were confirmed with rigorous spectral methods. A tail-in-bucket and wire-like structure of CDE1 via intermolecular self-assembly was observed by TEM and SEM examination; in contrast, CDE1-Ada-DOX exhibited unorganized structure due to disruption of self-assembly in the presence of guest molecules. The binding of Kd between CDE1 and Ada-DOX through hydrophobic interactions was determined to be 0.77 mM by fluorescence titration. CDE1-Ada-DOX showed sustained and two-phase exponential drug-release kinetics over 75 hours. Notably, for the mER-targeted CDE1-Ada-DOX inclusion complex, the critical factor for drug uptake efficiency in MCF-7 cells relied on the equilibrium between the host–guest and drug complex. By altering the ratios between the host and the guest molecules and the breakdown of the self-assembly nanostructure, CDE1-Ada-DOX delivered the anticancer drug into MCF-7 cells in a controlled manner. E2 and tamoxifen suppressed the drug uptake in MCF-7 cells treated with CDE1-Ada-DOX through competition for mER binding. Moreover, CDE1-Ada-DOX binds to recombinant human ERα fragments with a Kd of 0.027 µM determined by fluorescence polarization. The treatment of MCF-7 cells with CDE1-Ada-DOX elicited rapid activation of MAPKs (p44/42 MAPK, Erk1/2) in minutes through phosphorylation of Thr202/Tyr204. These results demonstrate a targeted delivery of the DOX derivative Ada-DOX to mER-positive breast cancer cells using CDE1 as the drug carrier vectors in a controlled manner. The estrogen conjugates elicit nongenomic (but not genomic) events in MCF-7 cells. CDE1 can be used as a powerful probe to explore the classical and nonclassical steroid-mediated pathways that are critical in the initiation, development, and progression of certain type of cancer (breast and ovarian cancer).
Acknowledgments
The authors are grateful for the support from the National Natural Science Foundation of China (Grant No 81372383), and the startup fund from College of Pharmacy, University of South Florida. This work has been supported in part by the Florida Center of Excellence for Drug Discovery and Innovation at the University of South Florida. The authors thank Dr S Karoly and Dr A Garces at the Lisa Muma Weitz Advanced Microscopy Core Laboratory for their support in data acquisition of flow cytometry and electronic microscopy. We also thank Dr R Sprunt at Moffitt Cancer Center for the MALDI-TOF mass spectroscopy.
Disclosure
The authors report no conflicts of interest in this work.
References
- SchneiderHJBinding mechanisms in supramolecular complexesAngew Chem Int Ed Engl200948223924397719415701
- KubikSMolecular cages and capsules with functionalized inner surfacesTop Curr Chem201231913421952842
- Del ValleEMMCyclodextrins and their uses: a reviewProcess Biochem200439910331046
- ZhangXWangCSupramolecular amphiphilesChem Soc Rev20114019410120890490
- BenitoJMGómez-GarcíaMOrtiz MelletCBaussanneIDefayeJGarcía FernándezJMOptimizing saccharide-directed molecular delivery to biological receptors: design, synthesis, and biological evaluation of glycodendrimer-cyclodextrin conjugatesJ Am Chem Soc200412633103551036315315450
- PaliwalSRPaliwalRPalHCEstrogen-anchored pH-sensitive liposomes as nanomodule designed for site-specific delivery of doxo-rubicin in breast cancer therapyMol Pharm20129117618622091702
- ZhangJMaPXCyclodextrin-based supramolecular systems for drug delivery: recent progress and future perspectiveAdv Drug Deliv Rev20136591215123323673149
- DavisMEBrewsterMECyclodextrin-based pharmaceutics: past, present and futureNat Rev Drug Discov20043121023103515573101
- YinJJZhouZWZhouSFCyclodextrin-based targeting strategies for tumor treatmentDrug Deliv Transl Res20133436437425788282
- AndréSKaltnerHFuruikeTNishimuraSIGabiusHJPersubstituted cyclodextrin-based glycoclusters as inhibitors of protein-carbohydrate recognition using purified plant and mammalian lectins and wild-type and lectin-gene-transfected tumor cells as targetsBioconjug Chem2004151879814733587
- YinJJSharmaSShumyakSPSynthesis and biological evaluation of novel folic acid receptor-targeted, β-cyclodextrin-based drug complexes for cancer treatmentPloS One201385e6228923658721
- DebloisGGiguèreVOestrogen-related receptors in breast cancer: control of cellular metabolism and beyondNat Rev Cancer2013131273623192231
- ManavathiBDeyOGajulapalliVNBhatiaRSBugideSKumarRDerailed estrogen signaling and breast cancer: an authentic coupleEndocr Rev201334113222947396
- YueWYagerJDWangJPJupeERSantenRJEstrogen receptor-dependent and independent mechanisms of breast cancer carcinogenesisSteroids201378216117023178278
- ErolesPBoschAPérez-FidalgoJALluchAMolecular biology in breast cancer: intrinsic subtypes and signaling pathwaysCancer Treat Rev201238669870722178455
- LiangJShangYEstrogen and cancerAnnu Rev Physiol20137522524023043248
- Dahlman-WrightKCavaillesVFuquaSAInternational Union of Pharmacology. LXIV. Estrogen receptorsPharmacol Rev200658477378117132854
- KampaMPelekanouVNotasGStathopoulosENCastanasEThe estrogen receptor: two or more molecules, multiple variants, diverse localizations, signaling and functions. Are we undergoing a paradigm-shift as regards their significance in breast cancer?Hormones (Athens)2013121698523624133
- SoltysikKCzekajPMembrane estrogen receptors – is it an alternative way of estrogen action?J Physiol Pharmacol201364212914223756388
- RenoirJMMarsaudVLazennecGEstrogen receptor signaling as a target for novel breast cancer therapeuticsBiochem Pharmacol201385444946523103568
- AliSBuluwelaLCoombesRCAntiestrogens and their therapeutic applications in breast cancer and other diseasesAnnu Rev Med20116221723221054173
- NilssonSKoehlerKFGustafssonJADevelopment of subtype-selective oestrogen receptor-based therapeuticsNat Rev Drug Discov2011101077879221921919
- GrillaudMRussierJBiancoAPolycationic adamantane-based dendrons of different generations display high cellular uptake without triggering cytotoxicityJ Am Chem Soc2014136281081924354635
- BohmIIsenbügelKRitterHBranscheidRKolbUCyclodextrin and adamantane host-guest interactions of modified hyperbranched poly(ethylene imine) as mimetics for biological membranesAngew Chem Int Ed Engl201150347896789921710677
- SilvaEKabilAKortenkampACross-talk between non-genomic and genomic signalling pathways – distinct effect profiles of environmental estrogensToxicol Appl Pharmacol2010245216017020206645
- MarinoMGalluzzoPAscenziPEstrogen signaling multiple pathways to impact gene transcriptionCurr Genomics20067849750818369406
- ParveenSSahooSKLong circulating chitosan/PEG blended PLGA nanoparticle for tumor drug deliveryEur J Pharmacol20116702–337238321951969
- TangWNgSCFacile synthesis of mono-6-amino-6-deoxy-alpha-, beta-, gamma-cyclodextrin hydrochlorides for molecular recognition, chiral separation and drug deliveryNat Protoc20083469169718388952
- AnandRMalangaMManetICitric acid-γ-cyclodextrin cross-linked oligomers as carriers for doxorubicin deliveryPhotochem Photobiol Sci201312101841185423900688
- KabouridisPSJanzenJMageeALLeySCCholesterol depletion disrupts lipid rafts and modulates the activity of multiple signaling pathways in T lymphocytesEur J Immunol200030395496310741414
- ParkerGJLawTLLenochFJBolgerREDevelopment of high throughput screening assays using fluorescence polarization: nuclear receptor-ligand-binding and kinase/phosphatase assaysJ Biomol Screen200052778810803607
- NilssonSMäkeläSTreuterEMechanisms of estrogen actionPhysiol Rev20018141535156511581496
- ChaudhriRAOlivares-NavarreteRCuencaNHadadiABoyanBDSchwartzZMembrane estrogen signaling enhances tumorigenesis and metastatic potential of breast cancer cells via estrogen receptor-α36 (ERα36)J Biol Chem2012287107169718122247547
- AlbanitoLMadeoALappanoRG protein-coupled receptor 30 (GPR30) mediates gene expression changes and growth response to 17beta-estradiol and selective GPR30 ligand G-1 in ovarian cancer cellsCancer Res20076741859186617308128
- MelocheSPouysségurJThe ERK1/2 mitogen-activated protein kinase pathway as a master regulator of the G1- to S-phase transitionOncogene200726223227323917496918
- JohnsonGLLapadatRMitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinasesScience200229856001911191212471242
- DentPCrosstalk between ERK, AKT, and cell survivalCancer Biol Ther201415324524624424114
- FilardoEJQuinnJABlandKIFrackeltonARJrEstrogen-induced activation of Erk-1 and Erk-2 requires the G protein-coupled receptor homolog, GPR30, and occurs via trans-activation of the epidermal growth factor receptor through release of HB-EGFMol Endocrinol200014101649166011043579
- BanerjeeSChamblissKLMineoCShaulPWRecent insights into non-nuclear actions of estrogen receptor-alphaSteroids201481646924252382
- BöttnerMThelenPJarryHEstrogen receptor-beta: tissue distribution and the still largely enigmatic physiological functionJ Steroid Biochem Mol Biol201413924525123523517
- WehlingMLöselRNon-genomic steroid hormone effects: membrane or intracellular receptors?J Steroid Biochem Mol Biol20061021–518018317113980
- WatsonCSAlyeaRAJengYJKochukovMYNongenomic actions of low concentration estrogens and xenoestrogens on multiple tissuesMol Cell Endocrinol20072741–21717601655
- DerooBJKorachKSEstrogen receptors and human diseaseJ Clin Invest2006116356157016511588
- KimHYSohnJWijewickramaGTClick synthesis of estradiol-cyclodextrin conjugates as cell compartment selective estrogensBioorg Med Chem201018280982120031420
- ClarkSRainvilleJZhaoXKatzenellenbogenBSPfaffDVasudevanNEstrogen receptor-mediated transcription involves the activation of multiple kinase pathways in neuroblastoma cellsJ Steroid Biochem Mol Biol2014139455324121066
- BulayevaNNGametchuBWatsonCSQuantitative measurement of estrogen-induced ERK 1 and 2 activation via multiple membrane-initiated signaling pathwaysSteroids200469318119215072920
- AdlanmeriniMSolinhacRAbotAMutation of the palmitoylation site of estrogen receptor-α in vivo reveals tissue-specific roles for membrane versus nuclear actionsProc Natl Acad Sci U S A20141112E283E29024371309
- LeaWASimeonovAFluorescence polarization assays in small molecule screeningExpert Opin Drug Discov201161173222328899