
Abstract
It has been reported that increased levels and activity of the heme oxygenase-1 (HO-1) protein ameliorate tissue injuries. In the present study, we investigated the effects and mechanisms of action of gold nanoparticles (AuNPs) on HO-1 protein expression in human vascular endothelial cells (ECs). The AuNPs induced HO-1 protein and mRNA expression in a concentration- and time-dependent manner. The induction was reduced by the thiol-containing antioxidants, including N-acetylcysteine and glutathione, but not by the non-thiol-containing antioxidants and inhibitors that block the enzymes for intracellular reactive oxygen species generation. The AuNPs enhanced Nrf2 protein levels but did not affect Nrf2 mRNA expression. In response to the AuNP treatment, the cytosolic Nrf2 translocated to the nucleus, and, concomitantly, Bach1 exited the nucleus and its tyrosine phosphorylation increased. The chromatin immunoprecipitation assay revealed that the translocated Nrf2 bound to the antioxidant-response element located in the E2 enhancer region of the HO-1 gene promoter and acted as a transcription factor. Although N-acetylcysteine inhibited the AuNP-induced Nrf2 nuclear translocation, the AuNPs did not promote intracellular reactive oxygen species production or endoplasmic reticulum stress in the ECs. Knockdown of Nrf2 expression by RNA interference significantly inhibited AuNP-induced HO-1 expression at the protein and mRNA levels. In summary, AuNPs enhance the levels and nuclear translocation of the Nrf2 protein and Bach1 export/tyrosine phosphorylation, leading to Nrf2 binding to the HO-1 E2 enhancer promoter region to drive HO-1 expression in ECs. This study, together with our parallel findings, demonstrates that AuNPs can act as an HO-1 inducer, which may partially contribute to their anti-inflammatory bioactivity in human vascular ECs.
Introduction
The endothelium plays a pivotal role in vasoregulation.Citation1 Vascular dysfunction or injury induced by aging, smoking, inflammation, trauma, hyperlipidemia, and hyperglycemia are among a myriad of risk factors that may contribute to the pathogenesis of many cardiovascular diseases, such as hypertension, diabetes, and atherosclerosis.Citation2 These factors also produce reactive oxygen species (ROS), which regulate fundamental cellular functions such as growth (hyperplastic or hypertrophic), endothelial dysfunction, migration, and inflammation, which have been demonstrated to play a role in atherogenesis.Citation3 Therefore, cells have evolved endogenous defense mechanisms to counteract oxidative stress. Among these, the redox-sensitive transcription factor nuclear factor erythroid 2-related factor 2 (Nrf2) serves as a “master regulator” of cell survival through the coordinated induction of the phase II and antioxidant defense enzymes via a cis-acting element, designated the antioxidant-response element (ARE), in the promoter of the target genes.Citation4 These include a myriad of genes encoding chemo-preventive proteins, including detoxifying enzymes NAD(P) H:quinone oxidoreductases (NQO1 and NQO2), the glutathione (GSH) S-transferase Ya subunit, γ-glutamylcysteine synthetase, and heme oxygenase (HO)-1.Citation5
HO is the rate-limiting enzyme in the catabolism of heme, a process that leads to formation of equimolar amounts of the bile pigment biliverdin, free iron, and carbon monoxide (CO).Citation6 There are two isoenzymes of HO: the original enzyme, designated HO-1, and the second isoenzyme, designated HO-2. These two enzymes share approximately 40% amino acid sequence homology.Citation7,Citation8 It is apparent that HO-2 is constitutively expressed, whereas HO-1 is inducible by a large number of structurally unrelated pharmacological and natural agents as well as by a variety of environmental influences, such as heat shock, and other forms of cellular stress.Citation6 Heme degradation is now considered critical for cellular defense, because the prooxidant heme is removed and the increased production of bilirubin and CO is beneficial and critical to the cellular defense mechanisms.Citation6 Major pathological alterations of the endothelium have been observed in HO-1 knockout mice, in which the endothelial cells (ECs) were more susceptible to apoptotic cell death and denudation from the extracellular matrix.Citation9 In addition, anti-inflammatory endothelial protection via HO-1 has been shown to be mediated by its ability to downregulate TNF-α-induced expression of various adhesion molecules.Citation10,Citation11
The common pathway for inducing HO-1 expression depends on the activation of redox-dependent transcriptional activators, such as Nrf2, NF-κB, and AP-1, along with the transcription repressor BTB and CNC homolog 1 (Bach1).Citation12 Activation of Nrf2 by oxidative stress is primarily controlled by the cytosolic inhibitor Kelch-like ECH-associated protein 1 (Keap1).Citation13 Numerous prooxidant stimuli and electrophiles cause the dissociation of Nrf2 from Keap1, which permits the subsequent nuclear translocation of Nrf2 and its interaction with AREs of HO-1 promoter.Citation12,Citation14 However, it was recently reported that Nrf2 can also be regulated and activated independent of Keap1, including the phosphorylation of Nrf2 by several signal transduction pathways, the involvement of epigenetic factors such as microRNAs, or the interaction of Nrf2 with other proteins.Citation12 Interestingly, an antioxidant, tert-butylhydroquinone, also induces tyrosine486 phosphorylation of Bach1 and leads to rapid nuclear export of Bach1, which allows Nrf2 to bind to the ARE and activate defensive gene expression.Citation15
Gold-based compounds have been paid much attention and have a long history of being used for therapeutic purposes in arthritis and cancer. Most commercially available colloidal gold solutions are obtained either by chemical reduction or physical synthesis. Previous studies have shown that gold nanoparticles (AuNPs) can affect vascular endothelial growth factor (VEGF)- and basic fibroblast growth factor (bFGF)-induced cell proliferation in vitro and angiogenesis in vivo.Citation16,Citation17 In addition, AuNPs bind to VEGF in arthritic synovial fluid, exert antiangiogenic activities, and reduce inflammation and arthritis.Citation18 However, some studies have also shown that nanoparticles, particularly high concentrations of silver nanoparticles, exhibit cell cytotoxicity and, therefore, induce HO-1 expression.Citation19,Citation20
In our parallel study, we observed that AuNPs are able to inhibit cell adhesion molecule (CAM) expression and have anti-inflammatory effects on human vascular ECs (unpublished data). Given that HO-1 expression has been associated with anti-inflammation and injury in some tissues,Citation6 we hypothesized that AuNPs can affect HO-1 expression. Therefore, the ability of AuNPs to induce HO-1 expression was evaluated and their mechanism of action in human vascular ECs was investigated in this study.
Materials and methods
AuNPs
AuNPs were obtained from Gold NanoTech Inc. (Taipei, Taiwan), and their preparation has been previously described.Citation21,Citation22 Briefly, the gold was cut into the target material and evaporated to the atomic level by an electrical gasification method. The evaporated gold was slowly deposited as AuNPs in distilled water under high vacuum (10 Citation8 Pa). The sizes of the AuNPs were controlled by the evaporation time and electric current used. The AuNPs, ranging from 3 to 5 nm, were dissolved in distilled water and gently agitated before being applied to the ECs. Their basic physical properties have been characterized, including their size distribution and average size. The average size was estimated to be 3.38±0.974 nm by transmission electronic microscopy (TEM) (unpublished data).
Materials
N-acetylcysteine (NAC), reduced GSH, vitamin C (vit C), Trolox, dithiothreitol (DTT), and the MAPK and PI-3K inhibitors were purchased from Sigma-Aldrich Co. (St Louis, MO, USA). Rotenone was obtained from Tocris Cookson Ltd. (Bristol, UK). Leptomycin B (LMB), apocynin, and thapsigargin were from Cayman Chemicals (Ann Arbor, MI, USA). CM-H2DCFDA was purchased from Thermo Fisher Scientific (Waltham, MA, USA). The antibody (Ab) for HO-1 was from Abcam (Cambridge, MA, USA). The endoplasmic reticulum (ER) stress sampler kit with Abs raised against stress proteins was purchased from Cell Signaling, Inc. (Danvers, MA, USA). The Abs for Bach1 and lamin B were obtained from Santa Cruz Biotechnology Inc. (Dallas, TX, USA). The Ab for α-tubulin was purchased from EMD Millipore (Billerica, MA, USA). The Ab raised against phosphotyrosine (4G10) was from EMD Milllipore (Billerica, MA, USA).
Cell cultures
The use of human umbilical cord veins for preparing human vascular ECs was approved by the Institutional Review Board of Cathay General Hospital (Taipei, Taiwan). The ECs were prepared and characterized by a previously described method.Citation23,Citation24 Isolated ECs were maintained in M199 containing 20% fetal bovine serum, 30 μg/mL endothelial cell growth supplement, 4 mM L-glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, and 250 ng/mL Fungizone (Thermo Fisher Scientific). The cells were used between the second and fourth passages in this study.
Cell lysate preparation, Western blot analysis, and immunoprecipitation
The cell lysates were prepared as previously described.Citation23 The total proteins were separated by electrophoresis through sodium dodecyl sulfate polyacrylamide gels, electroblotted onto poly-vinylidene fluoride membranes, and then probed using a primary Ab. The immunoblots were developed using Immobilon Western Chemiluminescent HRP Substrate (EMD Millipore, Billerica, MA, USA). To perform the immunoprecipitations (IPs), equal amounts of the total proteins were incubated with anti-Bach1 or phosphotyrosine Abs (1 μg Ab/200 μg of the total proteins) at 4°C for 24 hours, followed by incubation with Protein A/G PLUS-agarose (Santa Cruz Biotechnology Inc.) at 4°C for 2 hours. The beads were sedimented by brief centrifugation after the IP and washed extensively with RIPA lysis buffer. The proteins were released by boiling in Laemmli sample buffer and analyzed by Western blotting.
Enzyme-linked immunosorbent assay measurement of the HO-1 levels
The absolute intracellular levels of HO-1 were determined using a human HO-1 enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Inc., Minneapolis, MN, USA), according to the manufacturer’s protocol. Briefly, the ECs were treated with vehicle (ddH2O) or the AuNPs. The cell lysates were prepared, and the level of HO-1 was measured by ELISA at 450 nm. The absolute HO-1 concentrations were calculated from a standard curve.
Reverse transcription polymerase chain reaction analysis of HO-1, Nrf2, and β-actin mRNA expression
Oligonucleotide polymerase chain reaction (PCR) primers targeting human HO-1, Nrf2, and β-actin were synthesized and are listed in . Briefly, the total RNA from the ECs was extracted with TRIzol (Thermo Fisher Scientific) and the reverse transcription (RT) reaction was performed by using the iScript cDNA Synthesis Kit (Bio-Rad Laboratories Inc., Hercules, CA, USA). The cDNAs were synthesized from 1 μg of total RNA and the iScript reaction mix according to the manufacturer’s protocol. Aliquots of transcribed cDNAs were subjected to PCR in a 25 μL reaction mixture containing the GoTaq green master mix (Promega Corporation, Fitch-burg, WI, USA). PCR was performed with a hot start at 94°C for 5 minutes and then with 30 cycles of denaturation at 94°C for 1 minute, annealing at 51°C for 1 minute, and elongation at 72°C for 1.5 minutes on the ABI 7200 Thermal Cycler (Thermo Fisher Scientific). The amplification products were then analyzed by 2% agarose gel electrophoresis.
Table 1 Primers for the reverse transcription polymerase chain reaction analysis
Western blot analysis of Nrf2 translocation and Bach1 export
The cytosolic and nuclear fractions of the ECs were prepared using the NE-PER™ nuclear and cytoplasmic extraction reagents (Thermo Fisher Scientific) according to the manufacturer’s protocol. The Nrf2 translocation and Bach1 export were determined by Western blot analysis of their distributions in the cytosolic and nuclear fractions.
Immunofluorescence microscopy of the subcellular distribution of Nrf2 and Bach1
The cells were seeded on chamber slides and treated with the AuNPs. The cells were then washed, fixed with 1% paraformaldehyde for 20 minutes, and then permeabilized with 0.1% Triton X-100 for 10 minutes. After blocking with 3% BSA, the cells were incubated with control and specific Abs for Nrf2 (1:150; Abcam) or Bach1 (1:175; Santa Cruz Biotechnology Inc.) at 4°C overnight. Next, the chamber slides were incubated with DAPI (1:3,000; Thermo Fisher Scientific), analyzed under a Nikon Eclipse Ti-S fluorescence microscope (Nikon Corporation, Tokyo, Japan), and photographed using a digital camera. The images were merged using Adobe Photoshop 7.0.
Chromatin IP assay
The chromatin IP (ChIP) assay was performed using the EZ-Magna ChIP kit (EMD Milllipore) according to the manufacturer’s protocol. Briefly, the fixed cells were sonicated and centrifuged, and the supernatant fraction was used as the cell lysate in the ChIP analysis. Anti-Nrf2 Ab (Abcam) and normal rabbit IgG (EMD Milllipore), as a negative control, were used. These Abs were immobilized to magnetic protein G beads and used for the IP reactions. The precipitated DNA fragments were detected using RT-PCR and real-time PCR with the following previously described primers:Citation25 HO-1 E2 enhancer region: 5′-CCCTGCTGAGTAATCCTTTCC-3′ and 5′-GGCGGTGACTTAGCGAAAAT-3′; and HO-1 gene promoter region: 5′-GCCAGAAAGTGGGCATCAG-3′ and 5′-CTGAGGACGCTCGAGGGAG-3′. The real-time PCR was performed using a 7500 StepOne Real-Time PCR System (Thermo Fisher Scientific). The relative gene expression was determined by the ΔΔCt method, where Ct was the threshold cycle. All experiments were performed in duplicate or triplicate.
Measurement of intracellular ROS production
The ECs were seeded in 96-well plates and cultured until the cell density reached 90% confluency. The cells were incubated with M199 containing CM-H2DCFDA (5 μM) at 37°C for 30 minutes, followed by a brief wash. H2O (control), AuNPs, or H2O2 (100 μM) was added to ECs for the indicated time intervals, and the fluorescence intensity was determined by fluorometry at 485 nm (excitation) and 525 nm (emission).
Nrf2 siRNA transfection
ON-TARGET plus SMARTpool control and Nrf2 siRNAs were purchased from Thermo Fisher Scientific. The transfection was performed according to the manufacturer’s protocol, with some modifications. The ECs were seeded in six-well plates and incubated overnight in complete medium. The cell cultures were transfected with control or Nrf2 siRNAs (150 nM, 2 mL) for 3 days using the DharmaFECT transfection reagent. After being treated with the AuNPs, the cells were collected and prepared for Western blotting and RT-PCR analysis.
Statistical analysis
The data are expressed as the means ± standard error of the means. A comparison of the means of two groups of data was performed using the unpaired, two-tailed Student’s t-test. *P<0.05, **P<0.01, and ***P<0.001 are considered significant.
Results
AuNPs induce HO-1 protein and mRNA expression in ECs
Western blotting was performed to determine whether the AuNPs induced HO-1 expression in the ECs. As shown in , HO-1 was expressed at a low level in unstimulated cells, but was apparently increased by the AuNP treatment. A quantitative analysis indicated that the AuNPs induced HO-1 expression in a concentration- and time-dependent manner. A similar concentration-dependent result was obtained by ELISA, which showed that the HO-1 levels were significantly induced by the AuNPs at 1 and 2 ppm (). The AuNPs were dissolved in ddH2O. Therefore, to exclude the possibility that the HO-1 induction resulted from endotoxin contamination, a similar experiment was performed in the presence of polymyxin B, an antibiotic that can bind to the lipid A portion of lipopolysaccharide (LPS) and interferes with LPS function.Citation26 As shown in , LPS alone did not cause HO-1 expression, but induced intercellular adhesion molecule-1 (ICAM-1) expression. Polymyxin B did not interfere with the AuNP-induced HO-1 expression; however, it inhibited LPS-induced ICAM-1 expression.
Figure 1 Effect of the AuNPs on HO-1 protein expression.
Notes: (A and B) The cells were treated with (a) the indicated concentrations of the AuNPs for 16 hours or (b) 2 ppm of the AuNPs for the indicated time intervals. The cells were collected and 1 expression was analyzed by (A) Western blotting and (B) ELISA. (C) The cells were treated with vehicle (ddH2O), LPSor the AuNPs (2 ppm) in the presence or absence of polymyxin B for 16 h. ICAM-1 and HO-1 expression were determined by Western blotting (n=3). (D) The cells were treated with (a) the indicated concentrations of the AuNPs for 4 hours or (b) 2 ppm of the AuNPs for the indicated time intervals. The total RNAs were prepared, and 1 mNexpression was analyzed by RT-PCR(n=3). *P<0.05, **P<0.01, and ***P<0.001 versus the control.
Abbreviations: AuNPs, gold nanoparticles; ELISA, enzyme-linked immunosorbent assay; HO-1, heme oxygenase-1; ICAM-1, intercellular adhesion molecule; LPS, lipopolysaccharide; NS, non significant; RT-PCR, reverse transcription polymerase chain reaction.
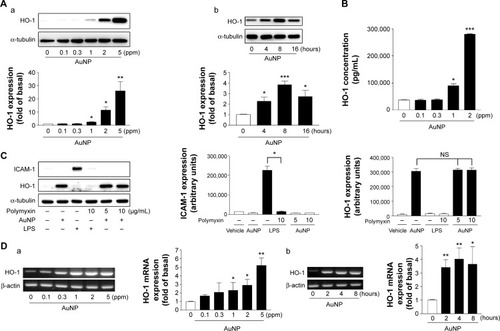
Next, we examined whether the AuNPs affected the HO-1 mRNA level in the ECs. As shown in , the AuNPs induced HO-1 mRNA expression in a concentration-and time-dependent manner. In contrast, the β-actin level remained unchanged. The HO-1 mRNA induction could be observed at 1 ppm of AuNPs and after 2 hours of AuNP treatment.
Effect of signaling inhibitors on AuNP-induced HO-1 expression
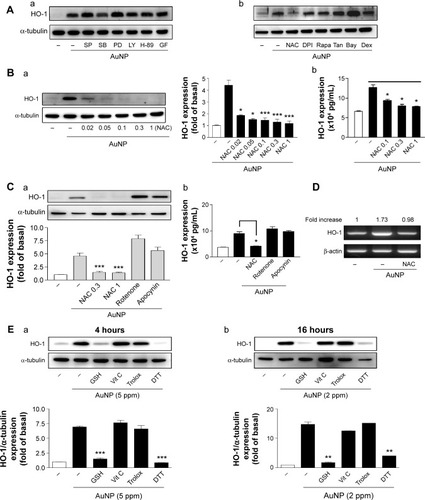
In an attempt to investigate the possible signaling pathways involved in the induction of HO-1 expression by the AuNPs, the effects of several pharmacological inhibitors targeting intracellular signaling pathways were determined. The inhibitors included SP600125 for JNK, SB202190 for p38 MAPK, PD98059 for MAPKK, LY294002 for PI-3K, H-89 for protein kinase A (PKA), GF109203X for PKC, NAC for scavenging ROS or destroying disulfide linkages,Citation27 Bay117085 for NF-κB, diphenyleneiodonium for NADPH oxidase inhibitor, rapamycin for the mTOR pathway, tan-shinone IIA for AP-1, and steroidal dexamethasone. In , the Western blot analysis shows that only NAC could abolish the AuNP-induced HO-1 expression. As shown in , the effects of different NAC doses and other ROS inhibitors on the ECs were further determined. NAC inhibited the AuNP-induced HO-1 expression in a concentration-dependent manner, as assayed by Western blotting and ELISA (). However, the AuNP-induced HO-1 expression was not affected by the other ROS-related inhibitors, including rotenone (a mitochondrial ROS inhibitor) and apocynin (a NADPH oxidase inhibitor) (). Moreover, the RT-PCR analysis indicated that NAC could suppress the AuNP-induced HO-1 mRNA expression (). As NAC have ROS scavenging and reducing activity, next, two ROS scavengers (vit C and Trolox) and two reducing agents (GSH and DTT) were used to clarify which types of antioxidants could affect the AuNP-induced HO-1 expression. As shown in , only GSH and DTT could abolish the AuNP-induced HO-1 expression at 4- and 16-hour incubation, suggesting that the AuNPs affect HO-1 expression via a redox reaction.
Figure 2 Effect of signaling inhibitors on the AuNP-induced HO-1 expression.
Notes: (A) The cells were pretreated with various inhibitors for 0.5 hours, followed by AuNP treatment (2 ppm) for 16 hours. (a) SP, SB, PD, and LY: 10 μM; H-89: 5 μM; GF: 2 μM. (b) NAC: 1 mM; Rapa; DPI: 1 μM; 1 μg/mL; Tan: 10 μM; Bay: 5 μM; Dex: 100 nM. The HO-1 expression was analyzed by Western blotting. (B and C) The cells were pretreated with (B) NAC (0.02–1 mM) or (C) the indicated inhibitors, followed by AuNP treatment (2 ppm) for 16 hours. The HO-1 expression level in (B) and (C) was analyzed by (a) Western blotting and (b) ELISA. NAC: 0.3 and 1 mM; rotenone: 100 μM; apocynin: 2 μM. (D) The cells were pretreated with NAC (1 mM), followed by AuNP treatment (2 ppm) for 4 hours. The HO-1 mRNA level was analyzed by RT-PCR (n=3–5). (E) Cells were treated with AuNPs in the presence or absence of GSH (5 mM), vit C (0.5 mM), Trolox (0.1 mM), or DTT (0.5 mM) for (a) 4 or (b) 16 hours. The HO-1 expression was determined by Western blotting (n=3). *P<0.05, **P<0.01, and ***P<0.001 versus the control. The NAC values are concentrations in mM.
Abbreviations: AuNPs, gold nanoparticles; Dex, dexamethasone; DPI, diphenyleneiodonium; DTT, dithiothreitol; ELISA; enzyme-linked immunosorbent assay; GSH, glutathione; HO-1, heme oxygenase-1; NAC, N-acetylcysteine; Rapa, rapamycin; RT-PCR, reverse transcription polymerase chain reaction; Tan, tanshinone IIA; vit C, vitamin C; SP, SP600125; SB, SB202190; PD, PD098059; LY, LY294002; GF, GF1009203X.
Effect of the AuNPs on the Nrf2-mediated signaling pathway
We have shown that NF-κB and AP-1 did not participate in the AuNP-induced HO-1 expression in the ECs (). Next, we tested whether the AuNPs influenced the Nrf2 signaling pathway. As shown in , treatment of ECs with the AuNPs led to a significant increase of the Nrf2 protein but not mRNA expression in a time- and concentration-dependent manner, suggesting the involvement of posttranslational modifications of Nrf2 in this induction.
Figure 3 Effect of the AuNPs on Nrf2 expression and translocation and Bach1 export.
Notes: (A) The cells were treated with (a) the indicated concentrations of the AuNPs for 4 hours or (b) 2 ppm of the AuNPs for the indicated time intervals, and Nrf2 expression was analyzed by Western blotting and RT-PCR (n=3). (B) The cells were treated with the AuNPs (2 ppm) for the indicated time intervals. The subcellular distribution (translocation/export) of Nrf2 and Bach1 was analyzed by Western blotting (n=3–4). (C) Immunofluorescence microscopy of subcellular distribution of (a) Nrf2 and (b) Bach1. The cells were treated with the AuNPs (5 ppm) for 3 hours. After fixation and permeabilization, the cells were stained with anti-Nrf2 and anti-Bach1 Abs, followed by staining with DAPI. The cells were photographed under the microscope. The nuclear-positive stained cells (green and indigo colors, arrows) and the total cells in the “Merge” panels were counted (n=4). The number of cells with Nrf2 translocation and Bach1 export was expressed as the nuclear-positive stained cells/total cells. ***P<0.001 versus the control.
Abbreviations: AuNPs, gold nanoparticles; Bach1, BTB and CNC homolog 1; DAPI, 4′,6-diamidino-2-phenylindole; HPF, high-power field; Nrf2, nuclear factor erythroid 2-related factor 2; RT-PCR, reverse transcription polymerase chain reaction.
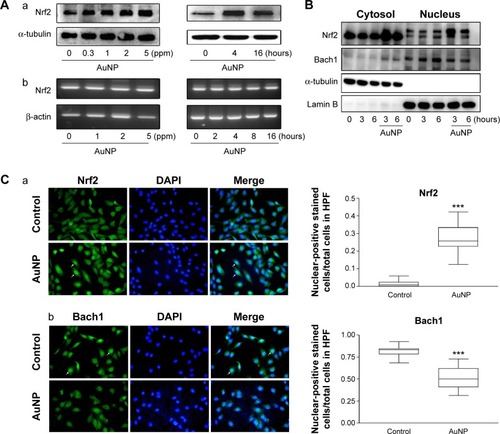
Next, we investigated whether the AuNPs affected sub-cellular distribution of Nrf2. As shown in , Nrf2 was preferentially expressed in the cytosol of the unstimulated ECs; however, Nrf2 expression in the nucleus was markedly increased in the AuNP-treated ECs. This indicates that Nrf2 translocated from the cytosol to nucleus. Interestingly, Bach1, a nuclear protein shown to associate with small MAF proteins and serve as a repressor of HO-1 gene expressionCitation28 was simultaneously increased in the cytosol of the ECs in response to the AuNP treatment, indicating that Bach1 was exported from the nucleus to cytosol (). This bidirectional transport of Nrf2 and Bach1 was further confirmed by immunofluorescence microscopy (). The location of nucleus was identified by DAPI (blue color), and the Nrf2 or Bach1 expression was identified by anti-Nrf2 and anti-Bach1 Abs (green color). The merger of these two images revealed more Nrf2-positive nuclear staining in the AuNP-treated cells and less Bach1-positive nuclear staining in the control group (indigo color). Taken together, the AuNP treatment significantly increased Nrf2 nuclear translocation and Bach1 cytosolic export in the ECs.
The phosphorylation of tyrosine486 can lead to rapid nuclear export of Bach1 and allows Nrf2 to bind to the ARE.Citation15 To examine the role of Bach1, LMB, a specific nuclear export inhibitor,Citation29 was used to determine its effect on the AuNP-induced HO-1 expression. shows that HO-1 protein and mRNA expression was notably reduced by LMB. Next, the alterations in Bach1 phosphorylation were examined in the presence of the AuNPs. shows that the tyrosine phosphorylation of Bach1 increased upon the AuNP treatment, as determined by IP. The internal controls indicated an equivalent loading and preparation of the samples.
Figure 4 Role of Bach1 export and tyrosine phosphorylation in the AuNP-induced HO-1 expression.
Notes: (A) Effect of the nucleocytoplasmic export inhibitor LMB on HO-1 expression. The cells were treated with LMB (20 ng/mL) and the AuNPs (2 ppm) for (a) 16 or (b) 4 hours. The HO-1 expression was determined by WB and RT-PCR (n=3). (B) The AuNPs induced Bach1 tyrosine phosphorylation. The cells were incubated with the AuNPs for 1.5 hours, and the tyrosine phosphorylation of Bach1 was determined by (a) IP of Bach1 followed by blotting with an anti-phosphotyrosine Ab or (b) IP of the tyrosine-phosphorylated proteins followed by blotting with an anti-Bach1 Ab. Both results show the tyrosine phosphorylation of Bach1 upon the AuNP treatment (n=3–4). The input was the cell lysates before the IP. b for basal. p-Bach1 for phospho-Bach1.
Abbreviations: Ab, antibody; AuNPs, gold nanoparticles; Bach1, BTB and CNC homolog 1; HO-1, heme oxygenase-1; IP, immunoprecipitation; LMB, leptomycin B; RT-PCR, reverse transcription polymerase chain reaction; WB, Western blotting.

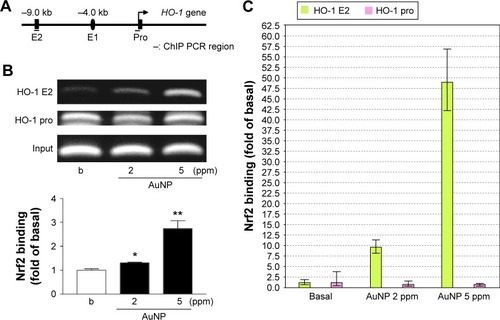
Effect of the AuNPs on Nrf2 binding to the HO-1 promoter region
In the human HO-1 gene locus, the regions at4 and9 kb upstream of the transcription start site contain multiple ARE sites.Citation30 To investigate whether the translocated Nrf2 interacted with the HO-1 promoter region, the primer sets for the enhancer region (E2) and promoter region near transcription start site were synthesized for ChIP assays (, upper panel schematic). The ChIP analyses demonstrated that AuNP treatment significantly induced Nrf2 binding to the human HO-1 E2 enhancer region, but not to the promoter region near the transcription start site. The binding to the enhancer region was concentration dependent, as determined by RT-PCR and real-time PCR ().
Figure 5 AuNPs induce Nrf2 binding to the ARE in the HO-1 promoter region.
Notes: (A) Schematic diagram of the primer design for the ChIP assay of the HO-1 promoter region. (B and C) The cells were incubated with vehicle or the AuNPs for 2 hours and then harvested for the ChIP assay. The precipitated DNA was analyzed by (B) RT-PCR and (C) real-time PCR (n=4). *P<0.05, **P<0.01, versus the Basil (b).
Abbreviations: ARE, antioxidant-response element; AuNPs, gold nanoparticles; ChIP, chromatin immunoprecipitation; HO-1, heme oxygenase-1; Nrf2, nuclear factor erythroid 2-related factor 2; PCR, polymerase chain reaction; RT-PCR, reverse transcription PCR.
Effect of the AuNPs on intracellular ROS production and ER stress
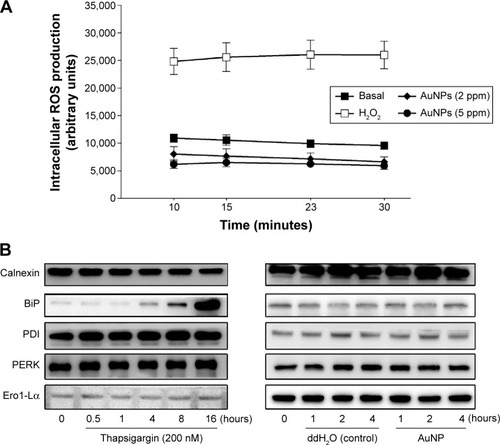
It has been reported that modification of one or more critical cysteine residues in Keap1 represents a likely chemico-biological trigger for the activation of Nrf2.Citation12 To determine whether AuNPs are a stressor, we examined ROS production and ER stress in the ECs upon the AuNP treatment. As shown in , the AuNPs did not cause intracellular ROS production, which was in contrast to the exogenous H2O2. The AuNPs also did not induce ER stress in the ECs. However, thapsigargin, a compound known to cause ER stress by emptying the ER of calcium ions, decreased calnexin and increased the BiP and PDI proteins after 4 hours of treatment ().
Figure 6 Effect of the AuNPs on intracellular ROS production and ER stress-related protein expression.
Notes: (A) CM-H2 DCFDA-loaded endothelial cells were treated with H2O2 (100 μM), vehicle, or the AuNPs (2 ppm) for the indicated time intervals. Intracellular ROS production was measured by fluorometry (n=4). (B) The cells were treated with thapsigargin, vehicle (ddH2O), or the AuNPs for the indicated time intervals; the ER stress-related protein expression was determined by Western blotting (n=3).
Abbreviations: AuNPs, gold nanoparticles; ER, endoplasmic reticulum; ROS, reactive oxygen species.
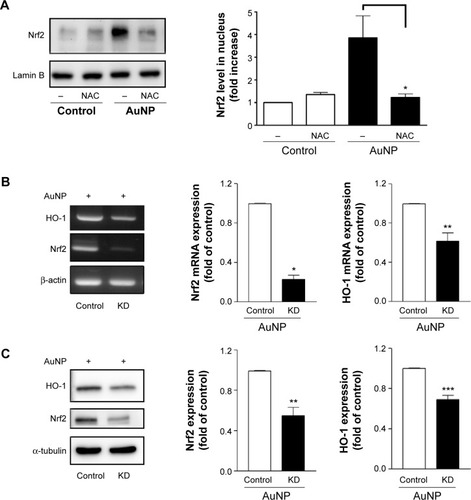
NAC diminishes Nrf2 translocation and Nrf2 knockdown reduces AuNP-induced HO-1 expression
We have shown that the AuNPs induced Nrf2 expression, translocation, and binding to the E2 enhancer region of HO-1 (, , and ). We next examined whether NAC can affect the AuNP-induced Nrf2 translocation. shows that NAC significantly inhibited Nrf2 translocation to the nucleus. Then, to verify the importance of Nrf2 in promoting HO-1 expression following AuNP treatment in the ECs, siRNA knockdown (KD) of Nrf2 was performed and its effect on the HO-1 mRNA and protein levels was determined. As shown in , siRNA KD of Nrf2 reduced the Nrf2 mRNA and protein levels by approximately 50~70%, and, concomitantly, the AuNP-induced HO-1 mRNA and protein expression was suppressed by approximately 40% and 30%, respectively.
Figure 7 Effects of NAC on Nrf2 translocation and Nrf2 KD on HO-1 expression.
Notes: (A) The cells were treated with the AuNPs (2 ppm) in the presence or absence of NAC (1 mM) for 1.5 hours. The nuclear fraction of the cell lysates was prepared and the Nrf2 level in this fraction was determined by Western blotting and densitometry. (B and C) Effect of Nrf2 KD on the AuNP-induced HO-1 mRNA and protein expressions. The cells were transfected with control or Nrf2 siRNAs for 72 hours, followed by the addition of vehicle or the AuNPs for 4 hours. Nrf2 and HO-1 expression were determined by (B) RT-PCR analysis and (C) Western blotting. *P<0.05, **P<0.01, and ***P<0.001 versus the control.
Abbreviations: AuNPs, gold nanoparticles; HO-1, heme oxygenase-1; KD, knockdown; NAC, N-acetylcysteine; Nrf2, nuclear factor erythroid 2-related factor 2; RT-PCR, reverse transcription polymerase chain reaction.
Discussion
HO-1 is the inducible isoform of the first and rate-limiting enzyme in the heme degradation pathway. HO-1 induction generally protects the cell against cytotoxicity from oxidative stress and apoptotic cell death. However, it was recently reported that HO-1 can also have immunomodulatory and anti-inflammatory properties.Citation6 In the present study, we showed that the AuNPs were able to induce HO-1 expression in human vascular ECs (). The AuNPs at 2 ppm were sufficient to promote HO-1 expression, and this induction could be observed at 4 hours of AuNP treatment. Based on the radius and atomic weight of the Au atom, the molarity of the 2 ppm AuNPs (average size: 3.38 nm) was estimated to be 1.08 nM, and it is possible that this concentration could be achieved in vivo.
Regarding the action mechanism of the AuNPs in promoting HO-1 expression in the ECs, we suggest that the Nrf2- and Bach1-mediated transcriptional pathways are major targets. A previous study has shown that Bach1 is a specific repressor of HO-1 and can be inactivated by arsenite, a potent inducer of oxidative stress.Citation30 However, an antioxidant (tert-butylhydroquinone) can also induce Bach1 phosphorylation at tyrosine486 and lead to rapid nuclear export of Bach1 that allows Nrf2 to bind to the ARE and activate defensive gene expression.Citation15 In the present study, there are several lines of evidence supporting the hypothesis that AuNPs target the Nrf2/Bach1 signaling pathways. First, the AuNPs increased the Nrf2 protein level (). Second, the AuNPs induced Nrf2 translocation to the nucleus and Bach1 export from the nucleus (). Third, the AuNP-mediated HO-1 expression could be attenuated by LMB, and the AuNPs induced Bach1 tyrosine phosphorylation (). Fourth, the AuNP treatment augmented Nrf2 binding to the E2 enhancer region of the HO-1 promoter (). Fifth, siRNA KD of Nrf2 reduced the cellular Nrf2 level and compromised the AuNP-induced HO-1 mRNA and protein expression (). Several natural and synthetic compounds are reported to be able to induce Nrf2.Citation12 Some of them are plant-derived compounds with chemopreventive properties, such as sulforaphane (broccoli),Citation31 curcumin (turmeric),Citation32,Citation33 and resveratrol (grapes).Citation34,Citation35 Others include trivalent arsenite and heavy metals.
Nrf2 has been shown to associate with Keap1, which is sensitive to redox reactions. Generally, in the absence of oxidative stress, Nrf2 is sequestered in the cytosol by the Keap1 homodimer, which acts as a substrate adaptor for the ubiquitination of Nrf2 in a cullin-3 (Cul3)-dependent manner.Citation36 It is proposed that subsets of the cysteine residues in Keap1 are modified in cells in response to chemical/oxidative stimuli. These modifications potentially result in the release of Nrf2 from Keap1, disturbing the transfer of ubiquitin. As a result, Nrf2 is no longer targeted for degradation, and the newly synthesized, free Nrf2 accumulates in the cytosol.Citation37 Interestingly, it has also been reported that DNA binding sites (ARE/xenobiotic response element) within the promoter region of Nrf2 are present, suggesting the ability of Nrf2 to regulate its own transcription, ie, autoregulation.Citation38 In this study, the AuNPs were able to increase Nrf2 protein, but not mRNA, expression in the ECs, implying that the AuNP-mediated induction resulted from posttranslational modifications of Nrf2 protein or Keap1. Because the AuNP-induced HO-1 expression and Nrf2 translocation could be abolished by the ROS scavenger/disulfide linkage reductant NAC ( and ), we hypothesized the involvement of a redox reaction/oxidative stress on Keap1 in the ECs in response to the AuNP treatment. However, the AuNPs did not cause intracellular ROS production or ER stress in the ECs (). Therefore, we speculate that NAC may not act as a ROS scavenger to inhibit the AuNP-induced HO-1 expression. It has been reported that NAC possesses a reducing property through its thiol-disulfide exchange activityCitation39 and can exert its action by directly reacting with some small molecules such as GSH and thiol-containing components, which then have the ability to interact with other thiol-containing proteins.Citation40,Citation41 In fact, NAC is also known for breaking or reducing disulfide linkages (-S-S-) and is largely used in the clinic as an antitussive/mucolytic agent.Citation27 Moreover, NAC is an antioxidant and possesses the ability to enhance cellular GSH synthesis.Citation27 From this viewpoint, we suggest that AuNPs can affect the cysteinyl thiol-group status of Keap1 because the oxidation, disulfide linkage formation, S-alkylation, and modification of the thiol-containing cysteines have been reported to change the conformation of Keap1, leading to Nrf2 dissociation from Keap1.Citation42 This may be the reason for the diminished AuNP-induced Nrf2 activation and/or HO-1 expression only observed in the presence of NAC, GSH, and DTT (thiol-containing antioxidants) rather than vit C and Trolox (non-thiol-containing antioxidants) ( and ). Some polyenes, such as chlorophylls and porphyrins, are Nrf2 inducers, and the polyphenols curcumin and quercetin have antioxidant activity and can also be HO-1 inducers.Citation32,Citation43 However, AuNPs do not possess these polyenic structures. Therefore, it remains to be investigated how AuNPs directly or indirectly affect Keap1 status and/or Nrf2 activation.
A previous study has shown that AuNPs from the same source as this study, but with larger sizes, elicit cytotoxicity in human CML K562 cells. Clusters of the AuNPs are detected in the plasma membrane or are endocytosed by cells, as determined by TEM.Citation22 Several toxicological reports show that many AuNPs have cytotoxicity in vitro and in vivo.Citation44–Citation46 Furthermore, some AuNPs and silver nanoparticles are able to induce HO-1 expression by causing cytotoxicity and oxidative stress.Citation19,Citation20,Citation47 However, the AuNPs in this study caused Nrf2 activation and HO-1 expression in a short incubation. The TEM analysis revealed that some AuNPs were located in the cytoplasm at 2 hours of treatment (data not shown), suggesting that the uptake of AuNPs by ECs can be an early event. Nevertheless, they did not cause cytotoxicity in the ECs, even with treatments up to 16 hours (unpublished data). Therefore, the induction of HO-1 by AuNPs does not appear to promote cell death. A report has shown that AuNPs do not increase cytotoxicity in the ECs, but increase their susceptibility to cell death.Citation48 In this regard, the long-term effects of AuNPs on ECs need to be further investigated.
Increased levels and activity of HO-1 have been reported to ameliorate injuries to several tissues induced by a number of diseases, including inflammatory diseases, diabetes, heart disease, hypertension, and others.Citation6 For example, HO-1 has been reported to inhibit the expression of proinflammatory CAM genes associated with EC activation.Citation11 Our preliminary result showed that HO-1 induction was partially involved in regulating CAM expression in the ECs, suggesting HO-1 induction and anti-EC inflammation.
Conclusion
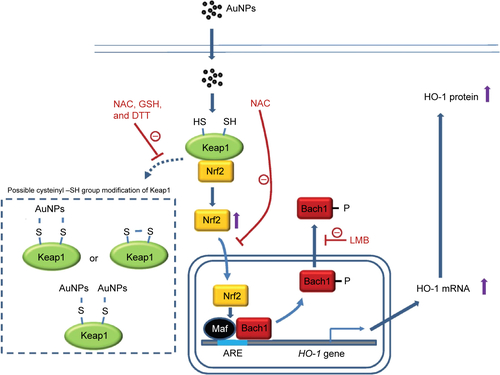
We provide here the first evidence that AuNPs can induce HO-1 expression in human vascular ECs by enhancing Nrf2 translocation and Bach1 tyrosine phosphorylation and export, leading to Nrf2 binding to the ARE promoter region of the HO-1 gene to drive HO-1 mRNA and protein expression. The induction does not appear to be mediated by inducing ROS and ER stress, but can be blocked by a ROS scavenger (NAC) and an inhibitor of nucleocytoplasmic export (LMB). The possible mechanism for the induction is summarized in . Therefore, this study elucidates the mechanism of AuNP-induced HO-1 expression and may also explain the anti-inflammatory activity of AuNPs toward vascular ECs.
Acknowledgments
This work was supported by research grants (MOST 101-2632-B-030-001-MY3 and 101CGH-FJU-08) from the National Science Council and Cathay General Hospital (Taipei, Taiwan), respectively.
Supplementary material
Figure S1 The proposed mechanism of action of the AuNPs in causing HO-1 expression in human vascular endothelial cells.
Abbreviations: ARE, antioxidant-response element; AuNPs, gold nanoparticles; DTT, dithiothreitol; GSH, glutathione; LMB, leptomycin B; NAC, N-acetylcysteine.
Disclosure
The authors report no conflicts of interest in this work.
References
- CinesDBPollakESBuckCAEndothelial cells in physiology and in the pathophysiology of vascular disordersBlood19989110352735619572988
- ZhangHParkYWuJRole of TNF-alpha in vascular dysfunctionClin Sci (Lond)2009116321923019118493
- SorescuDSzöcsKGriendlingKKNAD(P)H oxidases and their relevance to atherosclerosisTrends Cardiovasc Med2001113–412413111686001
- NitureSKKhatriRJaiswalAKRegulation of Nrf2-an updateFree Radic Biol Med201466364423434765
- JaiswalAKNrf2 signaling in coordinated activation of antioxidant gene expressionFree Radic Biol Med200436101199120715110384
- AbrahamNGKappasAPharmacological and clinical aspects of heme oxygenasePharmacol Rev20086017912718323402
- CruseIMainesMDEvidence suggesting that the two forms of heme oxygenase are products of different genesJ Biol Chem19882637334833533343248
- MainesMDTrakshelGMKuttyRKCharacterization of two constitutive forms of rat liver microsomal heme oxygenase. Only one molecular species of the enzyme is inducibleJ Biol Chem198626114114193079757
- TrueALOliveMBoehmMHeme oxygenase-1 deficiency accelerates formation of arterial thrombosis through oxidative damage to the endothelium, which is rescued by inhaled carbon monoxideCirc Res2007101989390117885218
- LiuXSpolaricsZMethemoglobin is a potent activator of endothelial cells by stimulating IL-6 and IL-8 production and E-selectin membrane expressionAm J Physiol Cell Physiol20032855C1036C104612839837
- SoaresMPSeldonMPGregoireIPHeme oxygenase-1 modulates the expression of adhesion molecules associated with endothelial cell activationJ Immunol200417263553356315004156
- PaineAEiz-VesperBBlasczykRImmenschuhSSignaling to heme oxygenase-1 and its anti-inflammatory therapeutic potentialBiochem Pharmacol201080121895190320643109
- ItohKWakabayashiNKatohYKeap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domainGenes Dev199913176869887101
- KasparJWNitureSKJaiswalAKNrf2:INrf2 (Keap1) signaling in oxidative stressFree Radic Biol Med20094791304130919666107
- KasparJWJaiswalAKAntioxidant-induced phosphorylation of tyrosine 486 leads to rapid nuclear export of Bach1 that allows Nrf2 to bind to the antioxidant response element and activate defensive gene expressionJ Biol Chem2010285115316219897490
- MukherjeePBhattacharyaRWangPAntiangiogenic properties of gold nanoparticlesClin Cancer Res20051193530353415867256
- KempMMKumarAMousaSGold and silver nanoparticles conjugated with heparin derivative possess anti-angiogenesis propertiesNanotechnology2009204545510419822927
- TsaiCYShiauALChenSYAmelioration of collagen-induced arthritis in rats by nanogoldArthritis Rheum200756254455417265489
- KangSJRyooIGLeeYJKwakMKRole of the Nrf2-heme oxygenase-1 pathway in silver nanoparticle-mediated cytotoxicityToxicol Appl Pharmacol20122581899822036727
- AueviriyavitSPhummiratchDManiratanachoteRMechanistic study on the biological effects of silver and gold nanoparticles in Caco-2 cells – induction of the Nrf2/HO-1 pathway by high concentrations of silver nanoparticlesToxicol Lett20142241738324126012
- YenHJHsuSHTsaiCLCytotoxicity and immunological response of gold and silver nanoparticles of different sizesSmall20095131553156119326357
- TsaiYYHuangYHChaoYLIdentification of the nanogold particle-induced endoplasmic reticulum stress by omic techniques and systems biology analysisACS Nano20115129354936922107733
- WuWBHungDKChangFWOngETChenBHAnti-inflammatory and anti-angiogenic effects of flavonoids isolated from Lycium barbarum Linnaeus on human umbilical vein endothelial cellsFood Funct20123101068108122751795
- LoHMLaiTHLiCHWuWBTNF-α induces CXCL1 chemokine expression and release in human vascular endothelial cells in vitro via two distinct signaling pathwaysActa Pharmacol Sin201435333935024487964
- MaruyamaAMimuraJHaradaNItohKNrf2 activation is associated with Z-DNA formation in the human HO-1 promoterNucleic Acids Res201341105223523423571756
- MorrisonDCJacobsDMBinding of polymyxin B to the lipid A portion of bacterial lipopolysaccharidesImmunochemistry19761310813818187544
- SamuniYGoldsteinSDeanOMBerkMThe chemistry and biological activities of N-acetylcysteineBiochim Biophys Acta2013183084117412923618697
- ShanYLambrechtRWDonohueSEBonkovskyHLRole of Bach1 and Nrf2 in up-regulation of the heme oxygenase-1 gene by cobalt protoporphyrinFASEB J200620142651265317065227
- WolffBSanglierJJWangYLeptomycin B is an inhibitor of nuclear export: inhibition of nucleocytoplasmic translocation of the human immunodeficiency virus type 1 (HIV-1) Rev protein and Rev-dependent mRNAChem Biol1997421391479190288
- ReichardJFMotzGTPugaAHeme oxygenase-1 induction by NRF2 requires inactivation of the transcriptional repressor BACH1Nucleic Acids Res200735217074708617942419
- KenslerTWCurpheyTJMaxiutenkoYRoebuckBDChemoprotection by organosulfur inducers of phase 2 enzymes: dithiolethiones and dithiinsDrug Metabol Drug Interact2000171–432211201301
- BalogunEHoqueMGongPCurcumin activates the haem oxygenase-1 gene via regulation of Nrf2 and the antioxidant-responsive elementBiochem J2003371Pt 388789512570874
- GargRGuptaSMaruGBDietary curcumin modulates transcriptional regulators of phase I and phase II enzymes in benzo[a]pyrene-treated mice: mechanism of its anti-initiating actionCarcinogenesis20082951022103218321868
- KodeARajendrasozhanSCaitoSYangSRMegsonILRahmanIResveratrol induces glutathione synthesis by activation of Nrf2 and protects against cigarette smoke-mediated oxidative stress in human lung epithelial cellsAm J Physiol Lung Cell Mol Physiol20082943L478L48818162601
- ChenCYJangJHLiMHSurhYJResveratrol upregulates heme oxygenase-1 expression via activation of NF-E2-related factor 2 in PC12 cellsBiochem Biophys Res Commun20053314993100015882976
- KobayashiAKangMIOkawaHOxidative stress sensor Keap1 functions as an adaptor for Cul3-based E3 ligase to regulate proteasomal degradation of Nrf2Mol Cell Biol200424167130713915282312
- BryanHKOlayanjuAGoldringCEParkBKThe Nrf2 cell defence pathway: Keap1-dependent and -independent mechanisms of regulationBiochem Pharmacol201385670571723219527
- KwakMKItohKYamamotoMKenslerTWEnhanced expression of the transcription factor Nrf2 by cancer chemopreventive agents: role of antioxidant response element-like sequences in the nrf2 promoterMol Cell Biol20022292883289211940647
- ZafarullahMLiWQSylvesterJAhmadMMolecular mechanisms of N-acetylcysteine actionsCell Mol Life Sci200360162012613655
- SunSYN-acetylcysteine, reactive oxygen species and beyondCancer Biol Ther20109210911019949311
- YuePZhouZKhuriFRSunSYDepletion of intracellular glutathione contributes to JNK-mediated death receptor 5 upregulation and apoptosis induction by the novel synthetic triterpenoid methyl-2-cyano-3, 12-dioxooleana-1, 9-dien-28-oate (CDDO-Me)Cancer Biol Ther20065549249716582599
- Dinkova-KostovaATHoltzclawWDColeRNDirect evidence that sulfhydryl groups of Keap1 are the sensors regulating induction of phase 2 enzymes that protect against carcinogens and oxidantsProc Natl Acad Sci U S A20029918119081191312193649
- ShenYWardNCHodgsonJMDietary quercetin attenuates oxidant-induced endothelial dysfunction and atherosclerosis in apolipoprotein E knockout mice fed a high-fat diet: a critical role for heme oxygenase-1Free Radic Biol Med20136590891524017971
- SasidharanAMonteiro-RiviereNABiomedical applications of gold nanomaterials: opportunities and challengesWiley Interdiscip Rev Nanomed Nanobiotechnol Epub2015324
- ZhangXDWuHYWuDToxicologic effects of gold nanoparticles in vivo by different administration routesInt J Nanomedicine2010577178121042423
- CoulterJAJainSButterworthKTCell type-dependent uptake, localization, and cytotoxicity of 1.9 nm gold nanoparticlesInt J Nanomedicine201272673268522701316
- FanZYangXLiYDeciphering an underlying mechanism of differential cellular effects of nanoparticles: an example of Bach-1 dependent induction of HO-1 expression by gold nanorodBiointerphases201271–41022589053
- LeitePEPereiraMRSantosCACamposAPEstevesTMGranjeiroJMGold nanoparticles do not induce myotube cytotoxicity but increase the susceptibility to cell deathToxicol In Vitro201529581982725790728