Abstract
There is a need for a rapid, accurate serodiagnostic test useful for both women and men infected by Trichomonas vaginalis, which causes the number one sexually transmitted infection (STI). Women and men exposed to T. vaginalis make serum antibody to fructose-1,6-bisphosphate aldolase (ALD), α-enolase (ENO), and glyceraldehyde-3-phosphate dehydrogenase (GAP). We identified, by epitope mapping, the common and distinct epitopes of each protein detected by the sera of women patients with trichomonosis and by the sera of men highly seropositive to the immunogenic protein α-actinin (positive control sera). We analyzed the amino acid sequences to determine the extent of identity of the epitopes of each protein with other proteins in the databanks. This approach identified epitopes unique to T. vaginalis, indicating these peptide-epitopes as possible targets for a serodiagnostic test. Individual or combinations of 15-mer peptide epitopes with low to no identity with other proteins were reactive with positive control sera from both women and men but were unreactive with negative control sera. These analyses permitted the synthesis of a recombinant His6 fusion protein of 111 amino acids with an Mr of ~13.4 kDa, which consisted of 15-mer peptides of two distinct epitopes each for ALD, ENO, and GAP. This recombinant protein was purified by affinity chromatography. This composite protein was detected by enzyme-linked immunosorbent assay (ELISA), dot blots, and immunoblots, using positive control sera from women and men. These data indicate that it is possible to identify epitopes and that either singly, in combination, or as a composite protein represent targets for a point-of-care serodiagnostic test for T. vaginalis.
Introduction
The literature is replete with the negative health outcomes to women that result from trichomonosis, the sexually transmitted infection (STI) caused by Trichomonas vaginalis.Citation1 Although published reports frequently state that men clear this STI, there have been no rigorous clinical studies to support this contention. A remarkable feature of this STI among women and men is persistence,Citation1 where in men, the parasite has been detected in the prostate and where recently T. vaginalis DNA was detected in hyperplastic prostate tissue.Citation2,Citation3 Indeed, this hallmark for T. vaginalis may relate to the fact that in recent population studies, most women are asymptomaticCitation4,Citation5 despite high prevalence rates of infection in women.Citation4,Citation6 That the host–parasite interrelationship is exceedingly complex is now appreciated, as evidenced by the role of environmental factors regulating the expression of virulence attributesCitation7–Citation10 and the identification of preparatory factors for colonization and pathogenesis.Citation11–Citation19
Epidemiological studies indicating the relation between seropositivity and prostate cancerCitation20–Citation22 and the recent report showing T. vaginalis DNA at high rates in prostatic tissueCitation3 have made evident the need for a better point-of-care (POC) serodiagnostic test that is applicable to men with trichomonosis and that would be also useful for women patients with trichomonosis. There is currently a commercially available POC diagnostic test for trichomonosisCitation23–Citation25 (OSOM® Trichomonas Rapid Test, Sekisui Diagnostics, Lexington, MA, USA), but this test is useful only for women patients with trichomonosis. Nucleic acid amplification tests (NAATs) have proven useful and effective in the detection of T. vaginalis infections in both women and men.Citation26–Citation29 However, these tests require trained personnel, expensive equipment, and shipment of possibly labile samples to special facilities for processing. Thus, an inexpensive, rapid, sensitive, noninvasive POC serodiagnostic test for women and men may provide a platform for screening large at-risk cohorts. Such a test would be especially suitable for examining men if the connection between trichomonosis and prostate cancer is validated. Recently, the highly immunogenic trichomonad protein α-actininCitation30 (ACT) was analyzed and found to have potential as a serodiagnostic target.Citation31 However, we hypothesize that surface protein immunogens of T. vaginalis have equal if not more utility to form the basis of POC antibody-based serological tests for women and men.
We present data on the characterization of three metabolic enzyme immunogens, fructose-1,6-bisphosphate aldolase (ALD), α-enolase (ENO),Citation32 and glyceraldehyde-3-phosphate dehydrogenase (GAP).Citation19 The sera from women patients with trichomonosis and the sera from men, which are both highly seropositive for ACTCitation20–Citation22,Citation31 (the target protein for screening men for seropositivity), are immunoreactive with these proteins (positive control sera). Therefore, we identified the epitopes detected by positive control sera from women and men, and sequence identity analysis permitted identification of epitopes unique to T. vaginalis, based on minimal or absent identity to amino acid sequences of other proteins in databanks. Synthesis of 15-mer peptides containing the epitope sequences were screened individually or in combination and found to be immunoreactive with seropositive sera. Lastly, a plasmid was constructed that contained a synthesized DNA that encoded for a recombinant protein consisting of six sequential 15-mer peptides referred to as series of epitopes (SOE), within which were epitopes for ALD, ENO, and GAP. This recombinant SOE (rSOE) was detected by the positive control sera from women and men, indicating its potential as a composite target for the serodiagnosis of trichomonosis.
Materials and methods
SPOTs membrane synthesis for ALD, ENO, and GAP proteins
Oligopeptides (Custom Peptide Arrays) that have been immobilized on membranes (SPOTs system; Sigma-Aldrich Corp, St Louis, MO, USA) are routinely used for epitope identification, and this technology was used recently on a trichomonad immunogenic protein.Citation31 Thereby, oligopeptides derived from the sequences of T. vaginalis ALD (GenBank® accession number AAW78351), ENOCitation31 (GenBank accession number AAK73099), and GAPCitation19 (GenBank accession number AAA30325) were synthesized on activated membranes. ALD is comprised of 328 amino acids (36.3 kDa), ENO is comprised of 472 amino acids (51.3 kDa), and GAP is comprised of 350 amino acids (38.1 kDa). Five to 10 nmol of each peptide was covalently bound to a Whatman 50 cellulose support (Whatman International Ltd, Maidstone, UK) by the C-terminus, using Fmoc-L amino acid chemistry, and had an acetylated N-terminus. The oligopeptides, which spanned the entire sequence of the protein, were 11-mer amino acids in length and had a sequential overlap of eight amino acids.
Probing the ALD, ENO, and GAP SPOTs membrane with sera from women and men and monoclonal antibodies (MAbs)
During the course of our research on T. vaginalis, we have examined, for reactivity to the trichomonad immunogenic protein ACT, hundreds of sera samples of women patients with trichomonosis as well as thousands of sera samples of men, in studies determining the relation between serostatus and prostate cancer.Citation19–Citation22,Citation31–Citation36 Unreactive sera (negative control sera) and highly reactive sera (positive control sera) with ACT were recently used to identify the ACT epitopes.Citation31 Importantly, individual ACT-seropositive sera of women and men gave similar reactivity to trichomonad proteins and ACT, and this permitted us to pool sera, to have sufficient amounts for conducting epitope mapping experiments, as outlined below.
Each ALD, ENO, and GAP membrane was washed with a small volume of 100% MeOH for 5 minutes to avoid precipitation of hydrophobic peptides during the following procedure. After a 10-minute wash performed three times (3×) in 25 mL of tris(hydroxymethyl)aminomethane (Tris)-buffered saline (TBS; 50 mM Tris-HCl, pH 8.0, 137 mM NaCl, and 2.7 mM KCl), the SPOTs membrane was incubated in blocking buffer (TBS-5% bovine serum albumin [BSA]) for 2 hours. All reactions with the SPOTs membrane were conducted at room temperature (RT), with gentle agitation on a shaker platform. The membrane was first incubated with a 1:10 dilution of negative control sera from women or men. No spots were detected for each protein membrane. Next, the membranes were incubated similarly with a hybridoma supernatant containing monoclonal antibody (MAb) B44Citation32 and MAb B43Citation19 reactive with epitopes to each of the ENO and GAP proteins, respectively, in addition to MAbs newly generated to the proteins (–). This was done to determine whether the reactions of these MAbs were identical to those seen using positive control sera from women and men. Finally, the membranes were incubated with positive control sera from women or men. All incubations with sera and MAbs were for 3 hours prior to washing and processing (described below). Importantly, the sera from women and men were categorized as either unreactive (negative control sera) or highly reactive (positive control sera) with ACT, a highly immunogenic trichomonad protein,Citation30,Citation31 as before.Citation20–Citation22
Table 1 EpitopesTable Footnotea,Table Footnoteb of Trichomonas vaginalis fructose-1,6-bisphosphate aldolase are reactive with MAbs and human sera
Table 3 EpitopesTable Footnotea of Trichomonas vaginalis glyceraldehyde-3-phosphate dehydrogenase are reactive with MAbs and human sera
After washing the SPOTs membranes 3× for 5 minutes in TBS, the membrane was processed for color development and SPOTs analysis, as detailed recently.Citation31 Images of the membranes were obtained with an Apple iPhone 4 (Apple Inc, Cupertino, CA, USA) and for assessment of the extent of reactivity and determination of the epitope amino acid sequences (–). The epitope sequences were compared with other proteins by using the protein-protein basic local alignment search tool (BLAST [http://www.ncbi.nlm.nih.gov/BLAST]). Amino acid sequence alignments of the proteins were performed with CLC Protein Workbench, Version 6.5, (Aarhus, Denmark). Hydrophobicity plotsCitation37 and antigenicity plotsCitation38 were constructed using Lasergene® MegAlign® (DNASTAR Inc, Madison, WI, USA). Finally, the membranes were regenerated through a series of washings, also as previously described.Citation31
Synthesis of individual 15-mer epitopes
Peptide epitopes from each protein were selected, based on low percent identity of the amino acid protein sequence and solubility in aqueous solutions, for handling of the oligopeptides. The 15-mer amino acid sequences () were sent to Sigma-Aldrich Corp and synthesized using their PEPscreen® Custom Peptide service. Each individual 15-mer peptide was acetylated at the amino terminus and was amidylated at the carboxy terminus. The 15-mer peptide was screened by mass spectrometry to determine the yield and purity of each product. Peptide epitopes were received with a pass/fail designation and the amount of peptide was provided. Three ACT 15-mer peptides, as described recently,Citation31 were used as positive controls in dot blots (below). These peptides were designated ACT1, ACT2, and ACT3 and corresponded to the amino acid sequences AQPLYDEAIAFKEEV, FKDTFKYFDKDKSNS, and SVNRHHSQLITYIKH, respectively. The underlined amino acids represent the epitopes, as characterized before.Citation31
Table 4 List of synthetic 15-mer peptides of representative epitopes of ALD, ENO, GAP, and ACT that are reactive with positive control women and men sera
Dot-blot detection of individual and combined 15-mer epitopes with women and men sera
Approximately 1 µg of individual and/or a combination of synthetic peptides were dot-blotted onto a nitrocellulose membrane (Bio-Rad Laboratories, Hercules, CA) diluted and allowed to air dry for 30 minutes at 37°C. These dot blots were fit into individual wells of a 96-well microtiter enzyme-linked immunosorbent assay (ELISA) plate. Then, 100 µL of 2% ELISA-grade BSA (Sigma-Aldrich Corp) in phosphate-buffered saline (PBS) (eBSA-PBS), pH 7.4, was added and incubated for 2 hours at RT, after which 5 µL of a 1:1 dilution (v/v) of sera (from T. vaginalisc –negative-control and –positive-control women or men) in PBS, pH 7.4, was added and incubated for 30 minutes at RT. The remainder of the procedure was as detailed recently.Citation31 Densitometric scans were produced using the ImageJ software (rsbweb.nih.gov/ij).
Recombinant SOEprotein expression and purification
The SOE encoding for six 15-mer amino acid sequences, two each of which contained epitopes for ALD, ENO, and GAP, consisted of 111 amino acids for an Mr of 13.35 kDa. These epitopes were chosen because of minimal or no amino acid sequence identity to other proteins in databanks. The DNA encoding for the SOE with a His6 tag at the carboxy terminus was cloned into a pET23b expression plasmid construct containing an ampicillin (amp) resistance locus obtained from Gen-way Biotech, Inc (San Diego, CA, USA) that was transformed into Escherichia coli Bl21DE3. Recombinant E. coli (rE. coli) was stored as glycerol stocks at −70°C until used, which were thawed and streaked onto Luria Broth (LB) agar plates (10 g/L tryptone, 5 g/L yeast extract, 10 g/L NaCl, and 15 g/L agar) containing 25 µg amp. Isolated colonies were inoculated into 200 mL fresh LB containing amp and incubated in a shaker incubator at 37°C and 220 rpm. After overnight (o/n) growth, rE. coli were inoculated into fresh LB medium with amp and incubated for 3 hours at 37°C at 220 rpm, prior to the addition of 1 mM isopropylthiogalactoside (IPTG) (Sigma-Aldrich Corp, St. Louis, MO, USA) and incubation for an additional 3 hours. The rE. coli were centrifuged using a Sovall SLA-1500 rotor (Thermo Fisher Scientific Inc, Waltham, MA, USA) at 8,000 rpm and 4°C for 15 minutes. The supernatant was decanted, and the pellet was stored at −80°C until used.
At different time intervals prior to and after the IPTG addition, 1 mL of rE. coli was microfuged at 10,000 rpm for 15 minutes, and pellets were prepared for sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) (34, 35), for analysis of recombinant SOE::His6 fusion protein (12.2 kDa) expression after IPTG addition. Immunoblot analysis after SDS-PAGE confirmed the synthesis of SOE::His6, using as probe-positive control, the sera of women and men who were seropositive to ACT, as defined above.
For purification of recombinant SOE::His6 (rSOE), pelleted rE. coli were thawed for 15 minutes on ice and suspended in 10 to 20 mL lysis buffer (50 mM Tris-HCl, pH 8.0, 300 mM NaCl, 10 mM β-mercaptoethanol (β-ME), and 0.1% Triton-X100). Lysates were sonicated 3× at RT for 30 seconds, with samples placed on ice at 30-second intervals. Sonicates were centrifuged using a Sorvall SS-34 rotor (Thermo Fisher Scientific Inc) at 8,000 rpm and 4°C for 20 minutes to remove insoluble debris. Clarified supernatant was applied to a nickel-nitrilotriacetic acid (Ni2+-NTA) Superflow affinity column that binds the His6 tag on the protein, according to the manufacturer’s instructions (Qiagen Inc, Venlo, The Netherlands). Pooled, purified SOE protein preparations were concentrated and dialyzed against PBS using a stirred Amicon™ Ultrafiltration Cell according to the manufacturer’s instructions (EMD Millipore, Billerica, MA, USA). The protein concentration was determined by the Lowry protein assayCitation39, with BSA as a standard. Purification of the rSOE::His6 protein was confirmed by SDS-PAGE and immunoblot, as above. After SDS-PAGE, acrylamide gels were stained with Coomassie brilliant blue for visualization of the purified fusion protein used for immunoblotting. Positive efficient blotting of the recombinant protein was determined by staining the nitrocellulose membrane with Ponceau S solution. Both Coomassie brilliant blue and Ponceau S stains were from Sigma-Aldrich.
ELISA using rSOE
ELISA was performed by immobilizing purified rSOE protein onto 96-well, flat-bottom Nunc polystyrene plates (Thermo Fisher Scientific, Rochester, NY), as detailed recently.Citation31 Briefly, each well was coated with 100 µL containing 1 µg of rSOE diluted in sodium carbonate (28.6 mM):sodium bicarbonate (71.4 mM) buffer, pH 9.6, prepared in PBS, and the plates were incubated o/n at RT with gentle agitation. Each plate was then washed 3× with PBS-1% Tween 20 (PBS-T). On the third wash, the plates were incubated in PBS-T for 5 minutes at RT with gentle agitation, prior to removing the PBS-T. The plates were then incubated upside down o/n at RT on paper towels before being covered with plastic wrap and stored at 4°C until used. For testing, plates were washed twice with PBS-T. On the second wash, the plates were incubated in PBS-T at RT for 5 minutes with gentle agitation and slap dried. Each well was then blocked with 200 µL of eBSA for 2 hours at 37°C. Plates were then washed twice with PBS-T. On the second wash, the plates were incubated in PBS-T at RT for 5 minutes with gentle agitation, followed by removing the PBS-T. Next, 100 µL of a 1:25 dilution of the women and men sera in eBSA-PBS was added to each well in duplicate and incubated at RT for 5 minutes with gentle agitation before incubation for 4 hours at 37°C. The plates were washed 3× with PBS-T. On the third wash, the plates were incubated in PBS-T for 5 minutes at RT with gentle agitation. After removal of the PBS-T, 100 µL of secondary horseradish peroxidase-conjugated goat-anti-human immunoglobulin (Ig)G (Fc-specific) immunoglobulin IgG (Fc-specific) (Bio-Rad Laboratories, Hercules, CA), diluted 1:1,500 in eBSA-PBS, was added to each well and incubated at RT with gentle agitation for 5 minutes, before incubation for 1 hour at 37°C. The plates were washed 3× with PBS-T, as above, prior to addition of 100 µL of color development solution (phosphate-citrate buffer with sodium perborate and 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt) prepared according to the manufacturer’s (Sigma-Aldrich Corp) instructions and incubated at RT with gentle agitation for 15 minutes. Absorbance values at 405 nm (A405 mm) were obtained using a Bio-Tek Synergy HT plate reader (Bio-Tek Instruments, Inc, Winooski, VT, USA).
Results
Epitope characterization of ALD, ENO, and GAP by reactive sera of women and men
– list the epitopes detected for each of the three proteins. There were 13, 18, and 19 epitopes detected for ALD, ENO, and GAP, respectively. Available MAbs also detected unique epitopes for each protein. Sequence identity comparisons with other microbial and fungal pathogens as well as human homologues revealed that some epitopes had >50% identity at the amino acid–protein sequence level, as indicated. It is noteworthy that some epitopes had identity to other pathogens causing STIs and to human homologues. Importantly, positive control sera from both women and men recognized the same epitopes. Interestingly but perhaps not surprisingly, MAbs to ALD and ENO, but not GAP, detected epitopes not recognized by the positive control sera from the women and men. These data suggest that it is possible to identify epitopes unique to ALD, ENO, and GAP and with little to no identity to other proteins. Of interest is the fact that inclusion of other neighboring amino acids for each of the epitopes significantly decreased the percent identity to each of the homologues examined. Finally, only ALD had almost threefold more epitopes recognized by the women sera (eight epitopes) versus the men sera (three epitopes). Both ENO and GAP had approximately the same number of epitopes each. Lastly and interestingly, women and men detected both identical and distinct linear epitopes throughout the protein.
Sequence alignment of the T. vaginalis ALD proteins with proteins of other representative organisms and Homo sapiens
We have previously presented amino acid sequence alignments for ENO and GAP.Citation19,Citation32 In we now show the amino acid sequence comparison of ALD with the homo-logues indicated in . The boxed amino acids contained in the T. vaginalis sequence are the epitopes presented in . The order from top to bottom of the sequences represents the highest to lowest percent identities.
Figure 1 Sequence analyses of Trichomonas vaginalis fructose-1,6-bisphosphate aldolase.
Abbreviations: ALD, aldolase; Ca, Candida albicans; Ec, Escherichia coli; Hs, Homo sapiens; M, men; MAbs, monoclonal antibodies; Ng, Neisseria gonorrhoeae; Sa, Staphylococcus aureus; Sc, Saccharomyces cerevisiae; Spn, S. pneumoniae; Spy, Streptococcus pyogenes; Tp, Treponema pallidum; W, women; ALD12 and ALD64, designate MAbs to aldolase as per .

Hydrophobicity and antigenicity profiles of the ALD, ENO, and GAP sequences
presents analyses of hydrophobicityCitation37 and antigenicityCitation38 alignments relative to the epitopes along the protein (top of each profile). Of interest is that with few exceptions, the epitopes represent hydrophilic regions contained within the protein (values <0), perhaps consistent with the presentation of amino acids for antibody synthesis and recognition. The hydrophilic domains correspond to increased antigenicity (values >0), as shown in the bottom profiles. Based on these features of the epitopes, we then selected seven epitopes for ALD, eight for ENO, and six for GAP for synthesis of 15-mer peptides encoding the epitopes, as shown in , and the individual amino acid sequence encoding the epitope within the 15-mer peptide is shown with underlining.
Figure 2 Hydrophobicity and antigenicity profiles of Trichomonas vaginalis α-enolase, aldolase, and glyceraldehyde-3-phosphate dehydrogenase proteins.
Abbreviations: A and ALD, fructose-1,6-bisphosphate aldolase; E and ENO, α-enolase; G and GAP, glyceraldehyde-3-phosphate dehydrogenase; M, men; MAb, monoclonal antibody; W, women; ALD13, ALD55, ALD11, ALD25, and B44, designate MAbs to alpha-enolase, ALD12/64 designates a MAb to aldolase, and ALD32, ALD30, and B43 designate MAbs to glyceraldehyde-3-phosphate dehydrogenase.
Reaction of 15-mer peptides with epitopes with women and men sera
presents results from representative dot-blot reactions, using the positive control sera from women and men, of 15-mer peptides () for ALD (), ENO (), and GAP (). Peptides derived from ACT () were used as a positive control.Citation31 Reactivity was detected for each 15-mer peptide, albeit at different levels of spot intensities. No peptides were detectable using the negative control sera for both women and men that was determined to be unreactive with the full-length proteins. These data suggest that some peptide epitopes have potential as serodiagnostic targets.
Figure 3 Representative experiment showing dot blots of individual 15-mer peptide epitopes.
Abbreviations: ACT, α-actinin; ALD, fructose-1,6-bisphosphate aldolase; ENO, α-enolase; GAP, glyceraldehyde-3-phosphate dehydrogenase.
An important finding is that while epitope mapping revealed epitopes unique to women and men (–), in fact, individual 15-mer peptides with epitopes were seen by the positive control sera from both women and men. For example, ALD1 (epitope A-W1, ) and ALD5 (epitope A-W6) were initially identified with the positive control women sera, but these epitopes were also positive as 15-mer peptides when tested with men sera. Likewise, ENO7 (epitope E-W7) and GAP3 (G-W2) were detected by men sera, and GAP (epitope G-M6) was seen by women sera. These results indicate that serum reactivity of epitopes requires examination under different experimental conditions.
We next wanted to perform dot blots using random combinations of the 15-mer peptides (), to determine whether an increased extent of reactivity was seen for women and men sera. We further wanted combinations of peptides that might give equal reactivity for both sera. shows, in duplicate, the intensity of signal for each combination with the positive control women and men sera (labeled 4+/5+), while no detection of the peptide cocktails was evident with negative control sera (labeled -0-). Densitometric scans (bars 1–7) revealed overall better extent of reactions with the combination of peptides compared with the individual peptides for both women and men sera (data not shown). The combined peptides ACT2 and ACT3 (bars 8) served as positive controls with positive control sera from men and women reactive with ACT,Citation31 and the pooled peptides GAP1, GAP6, and ENO3 (bars 9) as well as GAP5, ENO5, and ALD6 (bars 10) showed no reactivity with the negative control sera of both women and men.
Figure 4 Representative experiments in duplicate dot blots of combinations of 15-mer peptide epitopes (A) and densitometric scans of reactive dot blots (B).
Abbreviations: ACT, α-actinin; ALD, fructose-1,6-bisphosphate aldolase; con, control; ELISA, enzyme-linked immunosorbent assay; ENO, α-enolase; GAP, glyceraldehyde-3-phosphate dehydrogenase glyceraldehyde; neg, negative.
Expression, purification, and immunodetection of recombinant SOE
We hypothesized that a novel protein, comprised of a series of 15-mer peptides immunoreactive with positive control sera from women and men, could be synthesized for analysis as a serodiagnostic target. We, therefore, constructed a plasmid encoding for a His6 fusion recombinant protein of 111 amino acids (Mr 13.35 kDa), within which were the epitopes for GAP2, GAP3, ENO6, ENO7, ALD4, and ALD5 (). Underlined amino acids indicate the 15-mer peptides (). These 15-mer amino acids were chosen because of little or no identity to other proteins in databanks (–). Recombinant E. coli expressing the SOE (lane 1) was synthesized and found to be soluble in lysates (lane 2) (Materials and methods), which permitted purification in Ni2+-NTA affinity chromatography, as shown in (lanes 6–11). The relative mobility of the SOE compared with molecular weight standards showed an Mr slightly greater than the calculated 13.35 kDa.
Figure 5 The 111 amino acid sequence and expression and purification of the rSOE encoding epitopes of GAP, ENO, and ALD.
Abbreviations: ALD, fructose-1,6-bisphosphate aldolase; ENO, α-enolase; GAP, glyceraldehyde-3-phosphate dehydrogenase; His6, hexahistidine; MAb, monoclonal antibody; mw, molecular weight; Ni2+, nickel; NTA, nitrilotriacetic acid; rSOE, recombinant protein of sequences of epitopes; SDS-PAGE, sodium dodecyl sulfate polyacrylamide gel electrophoresis; SOE, sequences of epitopes.
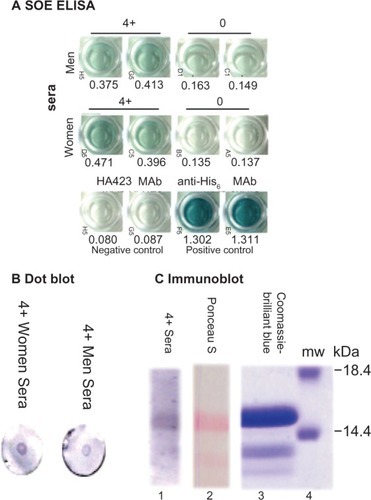
Lastly, we performed assays to assess whether this novel recombinant protein was detectable with positive control sera from women and men, as above. shows representative reactions by ELISA, of rSOE immobilized onto individual wells of 96-well microtiter plates. Negative control sera of women and men had little to no reactivity, as evidenced by low A405 nm with the rSOE in comparison with the strong signals (high A405 nm) obtained with the positive control sera from women and men. A very strong reaction was seen with rSOE using the positive control MAb to hexahistidine in the fusion recombinant protein. As expected, the MAb HA4232Citation31 that was reactive with the ACT used as a negative control gave no reactivity with the rSOE. Further, demonstrates that the positive control sera from women and men detected the rSOE by dot blots () and by immunoblot after SDS-PAGE and immunoblotting onto nitrocellulose (). The Coomassie-brilliant blue stained gel is a duplicate from the SDS-PAGE used for immunoblotting of the rSOE, and the Ponceau S-stained nitrocellulose is a duplicate included to show the transfer of the rSOE protein onto nitrocellulose.
Figure 6 Immunodetection of the rSOE by ELISA (A), dot blot (B), and immunoblotting after SDS-PAGE (C).
Abbreviations: ELISA, enzyme-linked immunosorbent assay; His6, hexahistidine; MAb, monoclonal antibody; mw, molecular weight; rSOE, recombinant protein of sequences of epitopes; SDS-PAGE, sodium dodecyl sulfate polyacrylamide gel electrophoresis.
Discussion
In this study, we have successfully identified epitopes of T. vaginalis ALD, ENO,Citation31 and GAP,Citation19 reactive with positive control sera from women and men, as serodiagnostic targets. Indeed, ACT is a target for a serodiagnostic test that would be worthwhile for both women and men exposed to this STI compared with the currently used POC OSOM® test sold by Sekisui Diagnostics, Inc, which works only for women patients with trichomonosis.Citation24,Citation25 The utility of this serum antibody target prompted us to examine for other immunogenic proteins, to which women patients and ACT-seropositive men make serum antibody and which might serve in future serodiagnostics. We, therefore, felt it important to perform a stepwise approach that would identify epitopes of immuno-genic proteins studied previously, such as ENOCitation32 and GAP.Citation19 More recently, we determined that another immunogenic protein is ALD (Alderete, unpublished data, 2013). In this study, we showed that 15-mer peptides containing the epitopes were readily detected by the positive control sera of women and men but not the negative control sera. These findings reinforce the notion that epitopes with little sequence identity to other known proteins, especially those of microbial pathogens and humans, can be identified to serve as serodiagnostic targets.
Interestingly, other microbial pathogens, including sexually transmitted disease agents, have high amino acid sequence identity to some of the epitopes detected by the positive control sera from women and men reactive to T. vaginalis (–). Although we have no evidence that the sera of patients infected by these other microbial pathogens used for comparison of sequence identity are cross-reactive with the epitopes identified in our study, we feel that this method and analysis would select for protein sequences of T. vaginalis that would be specific targets for the serodiagnosis of this STI. The fact that increasing the number of amino acids surrounding the epitopes significantly decreased the percent sequence identity further affirms the diminished possibility of any immuno-cross-reactivity. Nonetheless, it is conceivable that in fact, during infections by these other pathogens, patients may have antibody to similar sequences of their respective proteins.
An interesting finding is that the epitopes detected by the MAbs were distinct from those seen by the positive control sera from women and men, suggesting different presentations during immunization compared with natural infections. Further, women and men did not react with the identical epitopes, and this may be the result of the uniqueness of host responses, due to the distinct anatomy of the urogenital regions and immune surveillance. Although speculative, this may indicate that future serodiagnostics will need to take into account those epitopes detected by both women and men.
It is noteworthy that the trichomonad ENO has three epitopes () and GAP has six epitopes () with >50% sequence identity to the human homologue. Recently, it has been shown that patients with Streptococcus pyogenes infections had serum antibody to streptococcal ENO, and this antibody was cross-reactive with human tissues, such as the liver.Citation40,Citation41 While the adverse consequences of this remain undefined, such host responses may lead to autoimmune manifestations. Indeed, if infections result in tissue damage, this may be the result of such immuno-cross-reactivity and not just microbial toxins. Given this possibility, this requires further consideration.
Table 2 EpitopesTable Footnotea of Trichomonas vaginalis α-enolase are reactive with MAbs and human sera
As a direct result of epitope identification and characterization, we reasoned that it might be possible to synthesize a recombinant hybrid protein (rSOE) comprised of epitopes from the three immunogenic proteins of ALD, ENO, and GAP (). Indeed, this rSOE was readily purified and served as an immunotarget detected by various assays (). We feel this may be a novel way to produce such an amalgam of epitopes unique to a particular pathogen, such as T. vaginalis, and recognized by both women and men, regardless of the distinct natures of infection and protein presentations during immune surveillance. Such an rSOE may lead to novel platforms for rapid and accurate POC serodiagnostics for infectious diseases.
An important finding is that epitope mapping initially detected epitopes unique to positive control sera from women versus men (−). However, when tested individually, 15-mer peptides with epitopes were, in fact, seen by sera of both women and men (). For example, ALD1 (epitope A-W1) and ALD5 (epitope A-W6) were initially identified with positive control women sera (), but these epitopes were also positive as 15-mer peptides when tested with men sera (). Likewise, ENO7 (epitope E-W7) and GAP3 (G-W2) were detected by men sera, and GAP5 (epitope G-M6) was seen by women sera. These results indicate that serum reactivity of epitopes requires examination under different experimental conditions. Further, these findings inform that 11-mer cellulose-immobilized linear peptides for SPOTs analysis (–) present epitopes in a way that is different from 15-mer peptides immobilized on alternative surfaces, like nitrocellulose or microtiter wells. Likewise, serum antibody appears to distinguish the epitopes depending on the immobilizing surface, which may permit certain conformations accessible to antibody.
In summary, both women and men exposed to T. vaginalis make serum antibody to numerous trichomonad proteins.Citation19,Citation30–Citation36 As in an earlier recent study for the immunogenic trichomonad protein ACT,Citation31 we have identified epitopes of ALD, ENO, and GAP that are unique to this organism and showed their usefulness as serodiagnostic targets. We have also established the ability to synthesize a novel hybrid recombinant protein composed of numerous epitopes of ALD, ENO, and GAP proteins (called SOE) that forms a platform for a future serodiagnostic test. We have shown that such a hybrid rSOE can be the basis of a test for both women and men.
Acknowledgments
We thank Laurence N Kolonel, MD, PhD of the University of Hawaii Cancer Center and Siobhan Sutcliffe, PhD of Washington University at St Louis for their generous contribution of additional male sera that was highly seropositive for ACT-P2 and that permitted us to screen for epitopes of ACT. We also want to thank Patrick Joyce, Adan Medina, and Clare Tang, undergraduate student members of the laboratory, for their assistance during the course of these experiments and for their thoughtful discussions. This work was supported in part by the Washington Research Foundation.
Disclosure
The authors report no conflict of interest in this work.
References
- HobbsMMSeñaACSwygardHSchwebkeJRTrichomonas vagi-nalis and trichomoniasisHolmesKKSparlingPFStammWEPiotPWasserheitJNCoreyLCohenMSWattsDHSexually Transmitted Diseases4th edNew York, NYMcGraw Hill Medical2008771794
- GardnerWAJrCulbersonDEBennettBDTrichomonas vaginalis in the prostate glandArch Pathol Lab Med198611054304322421689
- MittereggerDAberleSWMakristathisAHigh detection rate of Trichomonas vaginalis in benign hyperplastic prostatic tissueMed Microbiol Immunol2012201111311621660495
- BachmannLHHobbsMMSeñaACTrichomonas vaginalis genital infections: progress and challengesClin Infect Dis201153Suppl 3S160S17222080269
- HuppertJSTrichomoniasis in teens: an updateCurr Opin Obstet Gynecol200921537137819491679
- KrashinJWKoumansEHBradshaw-SydnorACTrichomonas vaginalis prevalence, incidence, risk factors and antibiotic-resistance in an adolescent populationSex Transm Dis201037744044420351623
- LehkerMWAldereteJFBiology of trichomonosisCurr Opin Infect Dis2000131374511964771
- Figueroa-AnguloEERendón-GandarillaFJPuente-RiveraJThe effects of environmental factors on the virulence of Trichomonas vaginalisMicrobes Infect201214151411142723022315
- HuangKYHuangPJKuFMLinRAldereteJFTangPComparative transcriptomic and proteomic analyses of Trichomonas vaginalis following adherence to fibronectinInfect Immun201280113900391122927047
- HorváthováLŠafaríkováLBaslerMTranscriptomic identifi-cation of iron-regulated and iron-independent gene copies within the heavily duplicated Trichomonas vaginalis genomeGenome Biol Evol20124101017102922975721
- MuñozCPérezMOrregoPRA protein phosphatase 1 gamma (PP1γ) of the human protozoan parasite Trichomonas vaginalis is involved in proliferation and cell attachment to the host cellInt J Parasitol201242871572722713760
- Rendón-GandarillaFJRamón-Luing LdeLOrtega-LópezJRosa de AndradeIBenchimolMArroyoRThe TvLEGU-1, a legumain-like cysteine proteinase, plays a key role in Trichomonas vaginalis cytoadherenceBiomed Res Int2013201356197923509742
- Casta e Silva FilhoFde SouzaWLopesJDPresence of laminin-binding proteins in trichomonads and their role in adhesionProc Natl Acad Sci U S A19888521804280462973059
- ArroyoRGonzález-RoblesAMartínez-PalomoAAldereteJFSignalling of Trichomonas vaginalis for amoeboid transformation and adhesion synthesis follows cytoadherenceMol Microbiol1993722993098446032
- GarciaAFChangTHBenchimolMKlumppDJLehkerMWAldereteJFIron and contact with host cells induce expression of adhesins on surface of Trichomonas vaginalisMol Microbiol20034751207122412603729
- GarciaAFBenchimolMAldereteJF2005. Trichomonas vaginalis polyamine metabolism is linked to host cell adherence and cytotoxicityInfect Immun20057352602261015845462
- KucknoorASMundodiVAldereteJFAdherence to human vaginal epithelial cells signals for increased expression of Trichomonas vaginalis genesInfect Immun200573106472647816177319
- GarciaAFAldereteJCharacterization of the Trichomonas vaginalis surface-associated AP65 and binding domain interacting with trichomonads and host cellsBMC Microbiol2007711618158858
- LamaAKucknoorAMundodiVAldereteJFGlyceraldehyde-3-phosphate dehydrogenase is a surface-associated, fibronectin-binding protein of Trichomonas vaginalisInfect Immun20097772703271119380472
- SutcliffeSGiovannucciEAldereteJFPlasma antibodies against Trichomonas vaginalis and subsequent risk of prostate cancerCancer Epidemiol Biomarkers Prev200615593994516702374
- StarkJRJudsonGAldereteJFProspective study of Trichomonas vaginalis infection and prostate cancer incidence and mortality: Physicians’ Health StudyJ Natl Cancer Inst2009101201406141119741211
- SutcliffeSAldereteJFTillCTrichomonosis and subsequent risk of prostate cancer in the Prostate Cancer Prevention TrialInt J Cancer200912492082208719117055
- JonesHELippmanSACaiaffa-FilhoHHYoungTvan de WijgertJHPerformance of a rapid self-test for detection of Trichomonas vaginalis in South Africa and BrazilJ Clin Microbiol20135131037103923325818
- PillayALewisJBallardRCEvaluation of Xenostrip-Tv, a rapid diagnostic test for Trichomonas vaginalis infectionJ Clin Microbiol20044283853385615297548
- HegazyMMEl-TantawyNLSolimanMMEl-SadeekESEl-NagarHSPerformance of rapid immunochromatographic assay in the diagnosis of Trichomoniasis vaginalisDiagn Microbiol Infect Dis2012741495322727836
- LeeJJMoonHSLeeTYHwangHSAhnMHRyuJSPCR for diagnosis of male Trichomonas vaginalis infection with chronic prostatitis and urethritisKorean J Parasitol201250215715922711929
- GinocchioCCChapinKSmithJSPrevalence of Trichomonas vaginalis and coinfection with Chlamydia trachomatis and Neisseria gonorrhoeae in the United States as determined by the Aptima Trichomonas vaginalis nucleic acid amplification assayJ Clin Microbiol20125082601260822622447
- MunsonEMillerCNapieralaMAssessment of screening practices in a subacute clinical setting following introduction of Trichomonas vaginalis nucleic acid amplification testingWMJ2012111523323623189457
- MunsonKLNapieralaMMunsonEScreening of male patients for Trichomonas vaginalis with transcription-mediated amplification in a community with a high prevalence of sexually transmitted infectionsJ Clin Microbiol201351110110423100348
- AddisMFRappelliPPinto De AndradeAMIdentification of Trichomonas vaginalis alpha-actinin as the most common immunogen recognized by sera of women exposed to the parasiteJ Infect Dis199918051727173010515842
- NeaceCJAldereteJFEpitopes of the highly immunogenic Trichomonas vaginalis α-actinin are serodiagnostic targets for both women and menJ Clin Microbiol20135182483249023616456
- MundodiVKucknoorASAldereteJFImmunogenic and plasminogen-binding surface-associated alpha-enolase of Trichomonas vaginalisInfect Immun200876252353118070902
- LehkerMWAldereteJFIron regulates growth of Trichomonas vaginalis and the expression of immunogenic trichomonad proteinsMol Microbiol1992611231321310792
- AldereteJFNewtonEDennisCNealeKAThe vagina of women infected with Trichomonas vaginalis has numerous proteinases and antibody to trichomonad proteinasesGenitourin Med19916764694741774051
- AldereteJFNewtonEDennisCEngbringJNealeKAVaginal antibody of patients with trichomoniasis is to a prominent surface immunogen of Trichomonas vaginalisGenitourin Med19916732202252071124
- AldereteJFNewtonEDennisCNealeKAAntibody in sera of patients infected with Trichomonas vaginalis is to trichomonad proteinasesGenitourin Med19916743313341916796
- KyteJDoolittleRFA simple method for displaying the hydropathic character of a proteinJ Mol Biol198215711051327108955
- HoppTPWoodsKRPrediction of protein antigenic determinants from amino acid sequencesProc Natl Acad Sci U S A1981786382438286167991
- LowryOHRosebroughNJFarrALRandallRJProtein measurement with the Folin phenol reagentJ Biol Chem1951193126527514907713
- SunbladVBussmannLChiauzziVAPancholiVCharreauEHAlpha-enolase: a novel autoantigen in patients with premature ovarian failureClin Endocrinol (Oxf)200665674575117121525
- FontánPAPancholiVNociariMMFischettiVAAntibodies to streptococcal surface enolase react with human alpha-enolase: implications in poststreptococcal sequelaeJ Infect Dis200018261712172111069244