
Abstract
Introduction
Silica nanoparticles (SiNPs) are one of the most widely used inorganic nanomaterials, and exposure to SiNP has been demonstrated to induce pulmonary inflammation, primarily promoted by the NLRP3-mediated macrophage pyroptosis. However, mechanisms underlying the activation of NLRP3 signaling are complex, and whether cathepsin B (CTSB), an enzyme released by the ruptured lysosome, could trigger NLRP3 assembly is controversial.
Methods
To further characterize the role of CTSB in silica-induced pyroptosis, we conducted this study by establishing SiNP exposure models in vitro. The morphological features of SiNPs were exhibited by the SEM and TEM, and the effects of SiNPs’ internalization on macrophages were examined by the TEM and immunofluorescent staining. Moreover, Western blot was performed to detect the expression of proteins related to pyroptosis and CTSB after blocking the expression of NLRP3 and CTSB.
Results
We found that SiNPs internalization caused the rupture of macrophage membrane and promoted the aging of cells with increased intracellular vacuoles. Also, the expression of NLRP3, ASC, Caspase-1, GSDMD, Pro-IL-1β, IL-1β, and CTSB increased under the stimulation of SiNP, which could be suppressed by additional treatment with MCC950, an NLRP3-specific inhibitor. Besides, we found SiNP joint treatment with leupeptin, a CTSB inhibitor, could inhibit the expression of CTSB, but it had no effect on the expression of NLRP3, ASC, and Caspase-1, and the process of macrophage pyroptosis was also not affected.
Conclusion
SiNP exposure induces rupture of macrophages and the release of lysosomal CTSB, but CTSB fails to specifically act on the NLRP3 inflammasome to induce pyroptosis which is causally linked to lung inflammation and fibrosis.
Introduction
Over the past decades, nanotechnology has achieved rapid development in a broad range of fields like medicine, cosmetics, drug delivery, and imaging.Citation1,Citation2 It has dramatically increased the opportunities for nanoparticle exposure, and the health effects of nanoparticle exposure have also drawn considerable attention. Silica nanoparticle (SiNP), one of the most widely used nanomaterials, is commonly exposed in occupational conditions such as coal mining, metallurgy, and constructions, while long-term inhalation of SiNP can lead to pulmonary inflammation and fibrosis. Unfortunately, the pulmonary damages caused by SiNP exposure are irreversible and have no effective treatments at the current stage.Citation3,Citation4 Recent studies reveal that macrophage pyroptosis, a pro-inflammatory and pro-autolytic programmed cell death, plays a crucial role in the pathogenesis of silica-induced pulmonary inflammation,Citation5 indicating that in-depth investigation of the mechanism of pyroptosis may contribute to the identification of potential therapeutic targets of related diseases.
Macrophages are primary cells in the innate immune system and play essential roles in inflammatory response by releasing cytokines.Citation6 Studies have shown that macrophage pyroptosis is an indispensable segment of silicosis, an occupational disease induced by long-term silica exposure to large populations worldwide.Citation7 For the pathogenesis of silicosis, available evidence suggests that the inhaled SiNPs are first recognized by the pattern recognition receptors (PRRs) on the macrophage, then trigger nucleotide-binding oligomerization domain-like receptors (NOD-like receptors, NLRs), mainly NLRP3, and initiate the assembly of the NLRP3 inflammasome, composed of NLRP3, ASC, and Pro-Caspase-1. The activated NLRP3 inflammasome further proteolyzes the precursor of Caspase-1, while the cleaved Caspase-1 promotes the maturation and secretion of interleukin-18 (IL-18) and IL-1β and induces pyroptosis of macrophages.Citation8 Many studies have shown that pyroptosis is essential for the formulation of lesions and fibrosis of the kidney, liver, and bone, but less research has been done on the lungs.Citation9,Citation10 Although the previous study has confirmed macrophage pyroptosis initiation upon silica internalizationCitation11, how the internalized SiNP triggers NLRP3 inflammasome assembly and macrophage pyroptosis is still uncertain.
It is currently believed that lysosomal destruction dominates the assembly of NLRP3 inflammasome. Generally, crystals or particulate matters enter cells through endocytosis, resulting in swelling and dissolution of lysosomes. However, the dissolved and ruptured lysosomes are found to release Cathepsin B (CTSB), which may activate the assembly of the NLRP3 inflammasome. CTSB is a ubiquitous cysteine cathepsin mainly located in lysosomes and plays crucial roles in various physiological processes, such as inflammation, immune response, apoptosis, and extracellular matrix transformation.Citation11 Also, CTSB has been demonstrated to promote liver fibrosis, atherosclerosis, rheumatoid arthritis, and other diseases.Citation12,Citation13 For example, Tang finds that CTSB plays a pro-inflammatory role via activating the NLRP3 inflammasome, where CTSB aggravates inflammatory response in coxsackievirus B3-induced myocarditis.Citation14,Citation15 However, several studies have also shown that lysosomal disruption and CTSB are not always required for particle-induced assembly of the NLRP3 inflammasome. Hari et al report that the assembly of NLRP3 inflammasome can occur in the absence of CTSB.Citation16
To further investigate the role of CTSB in silica-induced macrophage pyroptosis, we established a SiNP exposure model using macrophages in vitro and examined the regulation of CTSB on SiNP-induced macrophage pyroptosis, and we showed data on the mechanism of silica-induced macrophage pyroptosis and clarified the potential role of CTSB in mediating silica-induced pulmonary inflammation and fibrosis.
Materials and Methods
Materials
ASC expressing mouse macrophage RAW264.7 (RAW-ASC) was purchased from InvivoGen (CA, USA). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) antibody, goat anti-rabbit IgG HRP-binding secondary antibody, and Dulbecco’s modified eagle’s high glucose medium (DMEM) were purchased from Servicebio (Wuhan, China). Silica nanoparticles (~30 nm, 99% of purity), adenosine triphosphate (ATP), penicillin and streptomycin, and lipopolysaccharide (LPS) were purchased from Solarbio (Beijing, China). Fetal bovine serum (FBS) was obtained from BI (Israel). MCC950 was obtained from Sigma-Aldrich (538120, MO, USA). Leupeptin and Lyso-Tracker-Red kits were purchased from Beyotime (103476-89-7, C1046, Beijing, China). Primary antibodies of ASC, Caspase-1, GSDMD, IL-1β, CTSB were purchased from ABclonal (A16672, A16792, A18281, A16288, A0697, Wuhan, China), and the NLRP3 antibody was obtained from Affinity Biosciences (DF7438, Cincinnati, OH, USA).
Transmission Electron Microscopy and Scanning Electron Microscopy
The morphology of the SiNP was characterized by transmission electron microscopy (TEM) and scanning electron microscope (SEM). After pretreatment with LPS for 6 h, RAW-ASC cells were exposed to SiNP (100 μg/mL) for 4 h. Then, the cells were fixed at 4℃ in phosphate-buffered saline (PBS) containing 2.5% paraformaldehyde. The samples were gradually dehydrated as follows: 30% ethanol for 15 minutes, 50% ethanol for 15 minutes, 70% ethanol for 15 minutes, 90% ethanol for 15 minutes, 90% acetone for 15 minutes, and four times as much as 100% acetone for 15 minutes. Subsequently, the samples were embedded in the resin and Polymerization at 37℃ for 12 hours. Ultrathin slices were cut on ultrathin slicers and fixed on copper grids. The grid was stained with uranyl acetate and lead citrate, air dried, and the cells were observed and photographed by a transmission electron microscope.
Cell Culture and Intervention
RAW-ASC cells were grown in DMEM containing 10% fetal bovine serum and 1% penicillin-streptomycin and incubated in a 37℃ cell culture chamber containing 5% CO2. A total of 2 × 104 cells/well were seeded in a 96-well plate, the SiNP group was pretreated with LPS (1 μg/mL) for 6 h and then exposed to 100 μg/mL SiNP, the control group received sham interventions. To explore the role of CTSB in SiNP-induced pyroptosis, we treated the cells with MCC950 (10 μM) and leupeptin (1 μg/mL), specific inhibitors NLRP3 and CTSB. These inhibitors were added 1 h before LPS intervention.
Western Blotting
Corresponding interventions were performed by seeding 2 × 106 RAW-ASC cells/well in a 6-well plate, and macrophages were lysed using RIPA lysis buffer containing a protease inhibitor cocktail on the completion of the intervention. To remove insoluble proteins, we centrifuged the mixture at 12,000 g, 4℃ for 30 min. Then, the supernatant was collected, and the protein was quantified, among which 10 μg of protein was separated and transferred onto the PVDF membrane and sealed with 5% skim milk. The target proteins were incubated overnight at 4℃ with specific primary antibodies. After rinsing with TBST, the membrane was incubated with the secondary antibody for 1 h. An Electrochemiluminescence working solution was employed to react with the membrane, and the membranes were visualized using a Fluor Chem HD2Gel imaging system (ProteinSimple, San Jose, CA, USA). The expression of the proteins was standardized with GAPDH, and each sample was repeated 3 times.
Lyso-Tracker Red Staining
According to the manufacturer’s instructions, lysosome staining was performed using a Lyso-Tracker Red kit. In brief, 2 × 104 RAW-ASC cells/well were seeded in a 96-well plate, to which 50 nM of Lyso-Tracker Red were added, followed by incubation at 37 ℃ for 30 min. After rinsing with PBS 3 times, the cells were imaged under an ImageXpress®Micro Confocal system (Molecular Devices, CA, USA).
Immunofluorescence Staining
A total of 2 × 104 RAW-ASC cells were seeded in a 96-well plate. Upon the completion of the intervention, the cells were washed with PBS 3 times and fixed with 4% paraformaldehyde. Then, the cells were saturated with 0.3% Triton X-100 for 10 min and blocked with goat serum for 30 min at room temperature. After the serum was discarded, the cells were incubated with specific primary antibodies (GSDMD, NLRP3, Caspase-1, IL-1β, 1:200) overnight at 4 ℃, and the fluorescence-conjugated secondary antibodies were added, followed by incubation in the dark at room temperature for 1 h. Then, 10 μL of Hoechst was added to cells to label the nuclei in blue for 10 min. Finally, the cells were subjected to the ImageXpress®Micro Confocal system to visualize the expression of the proteins.
Statistical Analysis
All data in this study were collected in Excel 2019 and summarized, visualized, and analyzed using R v3.6.3 for windows. Differences between groups were compared using the two-tailed t-test, and the quantitative results with normal distribution were displayed in a bar chart with a star when the difference was statistically significant at the 0.05 level.
Results
Characterization and Macrophage Endocytosis of SiNP
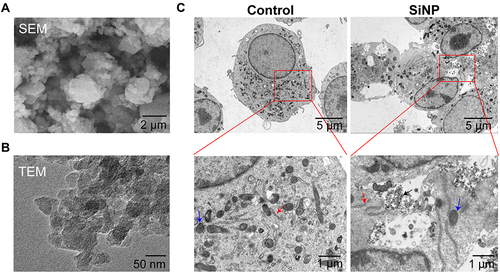
As shown in and , the morphological features and structure of SiNP were visualized using SEM and TEM, where we found the average diameter of SiNP was approximately 30 nm, ranging from 20 nm to 45 nm, and the particles were of high dispersion and were seldom aggregated. shows the ultrastructure of macrophages and the intracellular distribution of SiNP. Those macrophages without SiNP exposure were spherical, and many cells extended pseudopodia; the regular organelles such as nuclei, mitochondria and Golgi apparatus were determined. In contrast, many SiNPs along with the ruptured cell membrane, fuzzy mitochondria, vacuoles, and unevenly stained nuclei were determined in the silica-exposed macrophages.
Figure 1 Characterization of silica nanoparticles (SiNPs) in vitro. (A and B) Images of scanning electron microscopy (SEM) and transmission electron microscopy (TEM) showing SiNP in the forms of powder and solution. (C) TEM images showing the phagocytosis of SiNPs by RAW-ASC cells. RAW-ASC cells were pretreated with LPS for 6 h and then exposed to SiNP for 4 h. The control group received sham interventions. Black Arrow: SiNP; Red arrow: Golgi apparatus; Blue Arrow: Mitochondria.
Phagocytosis of SiNP Leads to Macrophage Pyroptosis
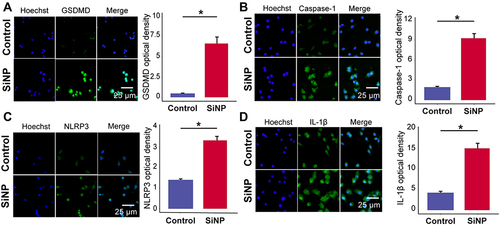
The internalization of xenobiotics triggers the inflammatory response of macrophages,Citation17 thereby we detected the effect of SiNP exposure on the initiation of macrophage pyroptosis, a pro-inflammatory and autolytic programmed cell death. As shown in , the expression of pyroptotic biomarkers such as NLRP3, GSDMD, Caspase-1, and IL-1β was detected by immunofluorescence. Compared with the control group, the immunofluorescence intensity increased after silica exposure, suggesting that macrophages exposed to SiNP undergone pyroptosis.
Figure 2 SiNP induces pyroptosis in RAW-ASC cells. RAW-ASC cells were pretreated with LPS for 6 h and then exposed to SiNP for 4 h. (A–D) Immunofluorescence images showing the expression of GSDMD, Caspase-1, NLRP3, IL-1β in RAW-ASC cells. Quantitative analysis and comparison of fluorescence intensity of images between groups. N=3. *P < 0.05.
Macrophage Pyroptosis Induced by SiNP Exposure Depends on NLRP3
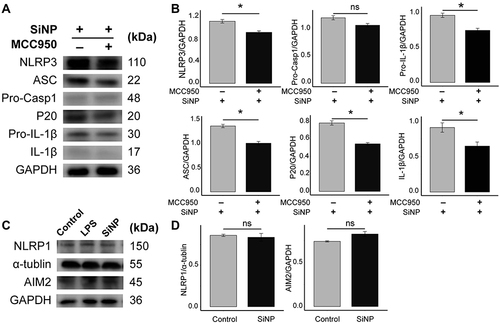
Accordingly, either NLRP3, AIM2, or NLRC4 inflammasome activation led to pyroptosis.Citation18 Here, we explored in which manner SiNP-induced pyroptosis is dependent and showed the results in (supported by the raw data deposited in Figures S1–S11). Utilizing MCC950, we blocked the activity of NLRP3. Compared with macrophages merely exposed to SiNPs, those cells that received joint treatment of SiNP and MCC950 showed attenuated NLRP3, ASC, cleaved Caspase-1 subunit P20, Pro-IL-1β, and IL-1β, while MCC950 had no effect on Pro-Caspase-1. Additionally, the alteration of AIM2 and NLRP1 was not statistically significant as compared to macrophages without SiNP exposure. These data suggest that NLRP3 plays a crucial role in mediating SiNP-induced macrophage pyroptosis.
Figure 3 SiNP exposure induces NLRP3-dependent pyroptosis in RAW-ASC cells. RAW-ASC cells were pretreated with MCC950 for 1 h, then LPS for 6 h and SiNP for 4 h. (A) Expression of NLRP3, ASC, Pro-Caspase-1 (Pro-Casp1), P20, Pro-IL-1β, IL-1β in RAW-ASC cells. (B) Quantitative analysis and comparison of proteins related to pyroptosis in A. The expression of these proteins was quantified by normalizing to GAPDH. (C) Expression of NLRP1 and AIM2 in RAW-ASC cells. (D) Quantitative analysis and comparison of proteins related to pyroptosis in C. The expression of NLRP1 and AIM2 was quantified by normalizing to α-tubulin and GAPDH. N=3. *P < 0.05.
SiNP-Induced Macrophage Pyroptosis Accompanies by Lysosome Rupture and Release of CTSB
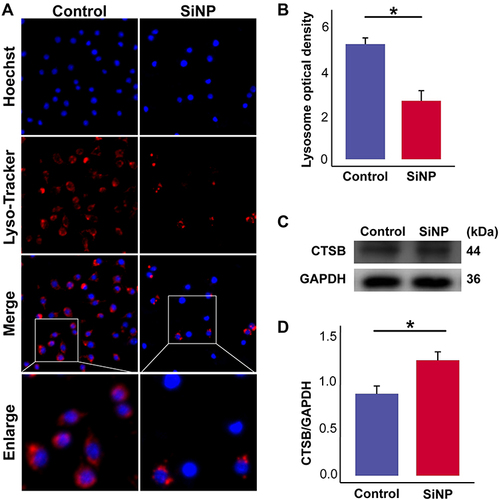
Given the essential role of the lysosome in digesting most phagocyted xenobiotics, we checked the effects of SiNP exposure on CTSB, an enzyme that primarily exists in the lysosome. Compared with the control group, and showed a significant reduction in the number of lysosomes after SiNP exposure. Consistently, the expression of CTSB as shown in and was upregulated after SiNP exposure. was supported by the raw data deposited in Figures S12 and S13. These data suggest that SiNP exposure can cause lysosomal rupture and promote the release of CTSB.
Figure 4 SiNP induces lysosome rupture and release of CTSB. RAW-ASC cells were pretreated with LPS for 6 h and then exposed to SiNP for 4 h. (A) Lysosomes were visualized by the Lyso-Tracker Red method. (B) Quantitative analysis and comparison of fluorescence intensity of lysosomes between groups. (C) Expression of CTSB in RAW-ASC cells. (D) Quantitative analysis and comparison of CTSB, which was quantified by normalizing to GAPDH. N=3. *P < 0.05.
CTSB Has No Effect on NLRP3-Dependent Macrophage Pyroptosis
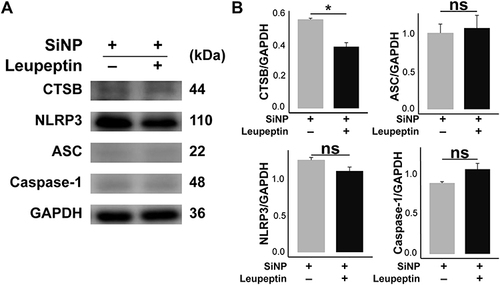
To explore the mediation of CTSB between SiNP recognition and NLRP3 inflammasome assembly, we treated the SiNP-exposed macrophages with leupeptin, a CTSB-specific inhibitor, and detected the expression of CTSB and proteins related to NLRP3-dependent macrophage pyroptosis. As shown in (supported by the raw data deposited in Figures S14–S18), the expression of CTSB was effectively suppressed after leupeptin intervention, but the changes of NLRP3, ASC, and Caspase-1 had no statistical significance as compared to macrophages merely treated with SiNP. These results demonstrate that CTSB is not a must for SiNP-induced macrophage pyroptosis.
Figure 5 Effect of CTSB on SiNP-induced macrophage pyroptosis. RAW-ASC cells were pretreated with leupeptin for 1 h, then LPS for 6 h and SiNP for 4 h. (A) The expression of CTSB, NLRP3, ASC, Caspase-1 in RAW-ASC cells. (B) Quantitative analysis and comparison of proteins in A. The expression of these proteins was quantified by normalizing to GAPDH. N=3. *P < 0.05.
Discussion
As an emerging risk factor for various diseases, SiNP is increasingly demonstrated to play crucial roles in immune response, oxidative stress, and a series of programmed cell death.Citation19,Citation20 To clarify the mechanism underlying silica-induced pulmonary inflammation, we establish silica exposure models in vitro from the perspective of macrophages and show the crucial role of cell pyroptosis in the pathogenesis of silica-induced physical disorders. Notably, we exhibit further evidence for the role of CTSB, a lysosomal enzyme still in controversy to induce NLRP3 inflammasome assembly. We demonstrate that silica exposure causes inflammation in a macrophage, while the inhibition of NLRP3 can suppress the activation of Caspase-1 and the release of inflammatory cytokines during SiNP-induced pyroptosis. Moreover, we found increased expression of CTSB, but suppressing CTSB using leupeptin had no effect on pyroptosis. These findings show further evidence of CTSB in mediating SiNP-induced macrophage pyroptosis, which may contribute to a better understanding of the pathogenesis of silica-induced pulmonary inflammation and fibrosis.
With the extensive application of SiNP, the threat of SiNP exposure to human health is also increasing, among which the pulmonary physical and functional damage has become an urgent issue concerned by current studies.Citation21 Macrophage is one of the primary components of the innate immune system and is extremely sensitive to SiNP stimulation.Citation22 Studies have demonstrated that SiNP can act as an irritant to stimulate the assembly of NLRP3 inflammasome in macrophages and promote the secretion of IL-1β under the mediation of Caspase-1, thereby participating in the occurrence of related inflammatory diseases.Citation23 Also, NLRP3 inflammasome is involved and accounts for a large proportion of the whole development of inflammatory responses.Citation24 Therefore, studying the upstream mechanism of NLRP3 inflammasome assembly may provide a new target for the treatment of pulmonary inflammation.
SiNP-induced pyroptosis is a newly defined pro-inflammatory and autolytic programmed cell death mediated by NLRP3 inflammasome.Citation25 However, other inflammasomes have also been reported to trigger pyroptosis.Citation26 Therefore, we use MCC950, a potent and selective inhibitor of NLRP3, to explore the role of NLRP3 in mediating SiNP-induced macrophage pyroptosis. We find that the lysosomes are swollen, deformed, and ruptured under the stimulation of SiNP, causing the release of CTSB and promoting the assembly of the NLRP3 inflammasome, and finally resulting in the secretion of inflammatory cytokines, which are consistent with the findings of Bai and Wang.Citation27,Citation28 Although lysosomal destruction has been reported to activate NLRP3 signaling, literature remains elusive concerning the importance of CTSB in mediating NLRP3 inflammasome assembly in macrophages.Citation29 Halle et al and other two scholars find that CTSB is not essential for NLRP3 inflammasome assembly following ATP or nigericin stimulation of macrophages.Citation16,Citation30,Citation31 Our results show that, although SiNP exposure can upregulate NLRP3, the upregulation of NLRP3 is unchanged after the cells being treated with leupeptin, suggesting that the release of CTSB is not specific to NLRP3 inflammasome assembly. Broadly, CA-074Me is commonly used as a pan-cathepsin inhibitor, and it could inhibit not only CTSB but also other cathepsins.Citation32 Direct evidence has not been reached on the dependency of CTSB upon CA-074Me-medicated NLRP3 inflammasome assembly.
To sum up, we have successfully defined the biological toxicity of SiNP in vitro, whose exposure leads to the rupture of the macrophage membrane and the release of lysosomal CTSB, but CTSB cannot specifically act on the NLRP3 inflammasome to induce pyroptosis that is causally associated with pulmonary inflammation and fibrosis.
Disclosure
The authors report no conflicts of interest in this work.
Acknowledgments
This work was financially supported by the National Natural Science Foundation of China (82003405), and the Natural Science Foundation of Shandong (ZR2020QH290). The funders had no role in study design, data collection, and interpretation, or the decision to submit the work for publication.
References
- Pan J, Wan D, Gong J. PEGylated liposome coated QDs/mesoporous silica core-shell nanoparticles for molecular imaging. Chem Commun. 2011;47(12):3442–3444. doi:10.1039/c0cc05520d
- Yetisen AK, Qu H, Manbachi A, et al. Nanotechnology in textiles. ACS Nano. 2016;10(3):3042–3068. doi:10.1021/acsnano.5b08176
- Frohlich E, Roblegg E. Oral uptake of nanoparticles: human relevance and the role of in vitro systems. Arch Toxicol. 2016;90(10):2297–2314. doi:10.1007/s00204-016-1765-0
- Pietroiusti A, Bergamaschi E, Campagna M, et al. The unrecognized occupational relevance of the interaction between engineered nanomaterials and the gastro-intestinal tract: a consensus paper from a multidisciplinary working group. Part Fibre Toxicol. 2017;14(1):47. doi:10.1186/s12989-017-0226-0
- Yin H, Fang L, Wang L, et al. Acute silica exposure triggers pulmonary inflammation through macrophage pyroptosis: an experimental simulation. Front Immunol. 2022;13:874459. doi:10.3389/fimmu.2022.87445
- Su DL, Lu ZM, Shen MN, Li X, Sun LY. Roles of pro- and anti-inflammatory cytokines in the pathogenesis of SLE. J Biomed Biotechnol. 2012;2012:347141. doi:10.1155/2012/347141
- Tan S, Chen S. The mechanism and effect of autophagy, apoptosis, and pyroptosis on the progression of silicosis. Int J Mol Sci. 2021;22(15):8110. doi:10.3390/ijms22158110
- Ishaq F, Sereesongsaeng N, Johnston MC, Scott CJ, Burden RE. Lysosomal cysteine proteases are mediators of cell death in macrophages following exposure to amorphous silica nanoparticles. Chem Biol Interact. 2022;356:109882. doi:10.1016/j.cbi.2022.109882
- Hou W, Wei X, Liang J, et al. HMGB1-induced hepatocyte pyroptosis expanding inflammatory responses contributes to the pathogenesis of Acute-on-Chronic Liver Failure (ACLF). J Inflamm Res. 2021;14:7295–7313. doi:10.2147/JIR.S336626
- Hu Y, Shi Y, Chen H, et al. Blockade of autophagy prevents the progression of hyperuricemic nephropathy through inhibiting NLRP3 inflammasome-mediated pyroptosis. Front Immunol. 2022;13:858494. doi:10.3389/fimmu.2022.858494
- Wu QQ, Xu M, Yuan Y, et al. Cathepsin B deficiency attenuates cardiac remodeling in response to pressure overload via TNF-alpha/ASK1/JNK pathway. Am J Physiol Heart Circ Physiol. 2015;308(9):H1143–1154. doi:10.1152/ajpheart.00601.2014
- Bao W, Fan Q, Luo X, et al. Silencing of Cathepsin B suppresses the proliferation and invasion of endometrial cancer. Oncol Rep. 2013;30(2):723–730. doi:10.3892/or.2013.2496
- Tong B, Wan B, Wei Z, et al. Role of cathepsin B in regulating migration and invasion of fibroblast-like synoviocytes into inflamed tissue from patients with rheumatoid arthritis. Clin Exp Immunol. 2014;177(3):586–597. doi:10.1111/cei.12357
- Tang Y, Cao G, Min X, et al. Cathepsin B inhibition ameliorates the non-alcoholic steatohepatitis through suppressing caspase-1 activation. J Physiol Biochem. 2018;74(4):503–510. doi:10.1007/s13105-018-0644-y
- Wang Y, Jia L, Shen J, et al. Cathepsin B aggravates coxsackievirus B3-induced myocarditis through activating the inflammasome and promoting pyroptosis. PLoS Pathog. 2018;14(1):e1006872. doi:10.1371/journal.ppat.1006872
- Hari A, Zhang Y, Tu Z, et al. Activation of NLRP3 inflammasome by crystalline structures via cell surface contact. Sci Rep. 2014;4:7281. doi:10.1038/srep07281
- Chen J, Ding J, Wu Y, et al. Chromium oxide nanoparticle impaired osteogenesis and cellular response to mechanical stimulus. Int J Nanomedicine. 2021;16:6157–6170. doi:10.2147/IJN.S317430
- Yu P, Zhang X, Liu N, Tang L, Peng C, Chen X. Pyroptosis: mechanisms and diseases. Signal Transduct Target Ther. 2021;6(1):128. doi:10.1038/s41392-021-00507-5
- Lo Giudice C, Yang J, Poncin MA, et al. Nanophysical mapping of inflammasome activation by nanoparticles via specific cell surface recognition events. ACS Nano. 2021;16(1):306–316. doi:10.1021/acsnano.1c06301
- Kamikubo Y, Yamana T, Hashimoto Y, Sakurai T. Induction of oxidative stress and cell death in neural cells by silica nanoparticles. ACS Chem Neurosci. 2019;10(1):304–312. doi:10.1021/acschemneuro.8b00248
- Zhou F, Liao F, Chen L, Liu Y, Wang W, Feng S. The size-dependent genotoxicity and oxidative stress of silica nanoparticles on endothelial cells. Environ Sci Pollut Res Int. 2019;26(2):1911–1920. doi:10.1007/s11356-018-3695-2
- Costantini LM, Gilberti RM, Knecht DA. The phagocytosis and toxicity of amorphous silica. PLoS One. 2011;6(2):e14647. doi:10.1371/journal.pone.0014647
- Jia H, Liu Y, Guo D, He W, Zhao L, Xia S. PM2.5-induced pulmonary inflammation via activating of the NLRP3/caspase-1 signaling pathway. Environ Toxicol. 2021;36(3):298–307. doi:10.1002/tox.23035
- Ozaki E, Campbell M, Doyle SL. Targeting the NLRP3 inflammasome in chronic inflammatory diseases: current perspectives. J Inflamm Res. 2015;8:15–27. doi:10.2147/JIR.S51250
- Faria SS, Costantini S, De lima VCC, et al. NLRP3 inflammasome-mediated cytokine production and pyroptosis cell death in breast cancer. J Biomed Sci. 2021;28(1):26. doi:10.1186/s12929-021-00724-8
- Lu F, Lan Z, Xin Z, et al. Emerging insights into molecular mechanisms underlying pyroptosis and functions of inflammasomes in diseases. J Cell Physiol. 2020;235(4):3207–3221. doi:10.1002/jcp.29268
- Bai H, Yang B, Yu W, Xiao Y, Yu D, Zhang Q. Cathepsin B links oxidative stress to the activation of NLRP3 inflammasome. Exp Cell Res. 2018;362(1):180–187. doi:10.1016/j.yexcr.2017.11.015
- Wang J, Wang L, Zhang X, et al. Cathepsin B aggravates acute pancreatitis by activating the NLRP3 inflammasome and promoting the caspase-1-induced pyroptosis. Int Immunopharmacol. 2021;94:107496. doi:10.1016/j.intimp.2021.107496
- Campden RI, Zhang Y. The role of lysosomal cysteine cathepsins in NLRP3 inflammasome activation. Arch Biochem Biophys. 2019;670:32–42. doi:10.1016/j.abb.2019.02.015
- Bauer C, Duewell P, Mayer C, et al. Colitis induced in mice with dextran sulfate sodium (DSS) is mediated by the NLRP3 inflammasome. Gut. 2010;59(9):1192–1199. doi:10.1136/gut.2009.197822
- Halle A, Hornung V, Petzold GC, et al. The NALP3 inflammasome is involved in the innate immune response to amyloid-beta. Nat Immunol. 2008;9(8):857–865. doi:10.1038/ni.1636
- Montaser M, Lalmanach G, Mach L. CA-074, but not its methyl ester CA-074Me, is a selective inhibitor of cathepsin B within living cells. Biol Chem. 2002;383(7–8):1305–1308. doi:10.1515/BC.2002.147