
Abstract
Introduction
Zinc is essential for the growth and differentiation of immune cells. Zinc insufficiency affects immune system function, thereby increasing infection susceptibility, autoimmunity, and allergies. Here, we aimed to determine the effects of zinc supplementation on T cell subpopulations, regulatory T (Tregs), T helper 1 (Th1), and T helper 17 (Th17) cells, in mixed lymphocyte cultures (MLC).
Methods
Allogeneic immune reactions were imitative using mixed lymphocyte cultures, followed by incubation with zinc to further monitor their effects. Cells were analyzed by flow cytometry. Production of Interferon-gamma (IFNγ), Interleukin-17 A (IL17A), and IL10 were analyzed by enzyme-linked immunosorbent assay. Th1 cell-specific Tbet, Th17 cell-specific RORC2, and Tregs-specific Foxp3 expression levels were determined by quantitative real-time PCR.
Results
Zinc supplementation at a physiological dose significantly increased CD4+ Foxp3+ Tregs and CD25+ Foxp3+ Tregs numbers and slightly decreased CD4+ RORC2+ and CD25+ RORC2+ Th17 cell numbers. A significant reduction in IFNγ production was observed in both restimulated T cells with autologous peripheral blood mononuclear cell (PBMC) and allogeneic PBMC compared to that in untreated T cells. Zinc significantly reduced IL17 expression, but the increase in IL10 expression was insignificant. In zinc-supplemented MLC, a non-significant decrease in Th1 or Th17 cell-specific transcription factors expression was observed, whereas there was a significant increase in Tregs-specific transcription factor expression.
Conclusion
Zinc can stabilize Tregs participating in adverse immune reactions or in an in vitro transplantation model.
Introduction
Nutrition critically affects the immune system function, and malnutrition has been reported to cause immunodeficiency worldwide.Citation1 For instance, zinc is essential for the development and differentiation of highly proliferating systems such as the immune system. Therefore, zinc deficiency influences the functioning of the immune system by increasing the person’s susceptibility to infectious diseases, autoimmunity, and allergies.Citation2,Citation3 Zinc (Zn) is critical for T-cell development and activation.Citation4,Citation5 Accordingly, in zinc-deficient patients, zinc level restitution recovers immune function and decreases the rate of infection in vivo.Citation6 In contrast, zinc administration induced concentration-dependent immunomodulatory effects in individuals with normal zinc levels. While immune function is boosted by a moderate concentration of zinc, it is also suppressed by high doses of zinc.Citation7,Citation8 According to these studies, zinc administration has a potential therapeutic role in graft-versus-host diseases (GVHD) and T cell-mediated autoimmunity.Citation5,Citation9
The T helper 17 (Th17) cells are a subtype of CD4 T lymphocytes and are responsible for extracellular pathogen clearance; however, they also cause several autoimmune diseases.Citation10,Citation11 Th17 cells are characterized by the expression of the retinoic acid receptor (RAR)-related orphan receptor (ROR)γt transcription factor, which is encoded by the Rorc gene.Citation12 Regarding immune tolerance and autoimmune diseases, Th17 cells have an opposing function to that of Treg cells. Th17 cells have been linked with several autoimmune and inflammatory diseases.Citation13 In addition, Th17 cells have been found to have a crucial role in the recruitment of immune cells to the site of inflammation and induce antigen-presenting cell maturation, which can contribute to acute and chronic allograft rejection.Citation14
The T helper 1 (Th1) cells are another CD4 subtype that has a fundamental role in cell-mediated immunity and graft rejection by producing the interferon-gamma (IFNγ) cytokines. The IFNγ is an inflammatory cytokine that can induce inflammation and stimulate phagocytes activation, macrophage function, and antigen presentation to T cells.Citation15
Regulatory T cells (Tregs) play an important role in immune suppression and self-tolerance. Tregs express CD4+ and CD25+ and exclusively express the forkhead/winged helix family transcriptional repressor Foxp3 (Forkhead-Box-Protein P3), which is necessary for Treg cell function and development.Citation13 In addition, Tregs are recognized from other subpopulations of CD4 cells by the expression of Foxp3.Citation16 Tregs have many subsets, including natural Tregs (nTregs), that occur throughout thymus development. Tregs are naturally released from the thymus into other tissues, representing 5–10% of CD4+ cells.Citation17 Another type of Tregs is induced Tregs (iTregs), which can be promoted in vivo or in vitro through T cell receptor stimulation.Citation18,Citation19 Importantly, iTregs secrete interleukin-10 (IL-10) and transforming growth factor β (TGF-β).Citation20,Citation21 The IL-10 and TGF-β are anti-inflammatory cytokines that act as immunosuppression agents which suppress the inflammation and T cell activations by reducing antigen presentation ability of dendritic cells, particularly in autoimmune diseases and rejection of transplantation.Citation21,Citation22
Previous research has shown that zinc critically modulates the immune response, including its enhancement, suppression, and regulatory effects.Citation23 Zinc deficiency leads to impaired T cell homeostasis, which disrupts immune tolerance and results in adverse immune reactions such as hypersensitivity, autoimmunity, and rejection of transplantation.Citation23–25 Several studies have revealed the impact of zinc on T cell subpopulations in allergies, autoimmune diseases, and transplantation. However, most of them have investigated the influence of zinc supplementation on Treg or Th1 or both cells in in-vivo and in-vitro models.Citation26–28 Research on how zinc supplementation affects Th17-cell function or all T-cell subtypes in the same environment in transplantation still needs to be conducted. Therefore, this study aimed to investigate the influence of zinc supplementation on Th1, Th17, and Treg cells in mixed lymphocyte cultures (MLC) as an in vitro model for allogeneic reactions and transplantation.
Materials and Methods
The Isolation of Human Peripheral Blood Mononuclear Cells and Making Mixed Lymphocyte Culture
Whole blood samples were obtained from fit youthful donors, for isolation of peripheral blood mononuclear cells (PBMC) using ficoll–hypaque density centrifugation (Biochrom, Berlin, Germany). These cells were obtained from interface, washed two times with PBS, and suspended in RPMI 1640 that contains 2 mM L-glutamine, FCS 10% (inactivated by heating at 56°C for 30 min), 100 U/mL streptomycin sulfate, and 100 U/mL potassium penicillin (all from Sigma-Aldrich, Darmstadt, Germany). A final concentration of 2×106 cells/mL was adjusted. To prepare two-way MLC, 2×106 PBMC/mL was obtained from two genetically different donors, then incubated with RPMI media or add-ed-on 50 μM zinc sulfate for 15 min (Sigma-Aldrich, Darmstadt, Germany), and next mixed at 1:1 ratio within pyrogen-free 24-well dishes for the specified time (VWR Scientific Products, Radnor, USA). Each incubation stage was performed in humidified 5% CO2 incubator at 37°C.Citation27
Preparing Alloantigen Specific T Cells
To prime T cells, PBMC could be used in a final concentration of 2×106 PBMC/mL. Priming B cells (BJAB cells) were fixed by the same volume of 3% paraformaldehyde (Sigma-Aldrich, Darmstadt, Germany) for three minutes. The B cells were then washed thrice with PBS (Sigma-Aldrich, Germany). The cell population of 2×106 PBMC/mL and fixed BJAB were incubated within or without 50 μM zinc sulfate for 15 min and then mixed within a pyrogen-free 24-well dish at 5:1 ratio for five days. Subsequently, the cell culture media were removed and the cells were adjusted to 5 × 105/mL in fresh culture media. Further, Staphylococcus aureus protein A (SPA) 2.5 μg/mL (Pharmacia Inc., New York, USA) was added to initiate the T cell expansion for two days, and 100 U/mL human IL2 was added for three days. A total of 2 × 106/mL expanded T cells (responders) were mixed at a ratio of 1:1 with PBMC either autologous or allogeneic (stimulators) for five days for T cell restimulation. All stages of incubation were performed at 37°C in a humidified 5% CO2 atmosphere.Citation23 Then, the supernatant was extracted and used for cytokine analysis.
Flow Cytometry Analysis of Foxp3 and RORC2
Anti-human CD4 (FITC), anti-human CD25 (FITC), and anti-human Foxp3 (PE) antibodies were obtained from BD Biosciences, Heidelberg, Germany. Also, Anti-human RORC2/RORγT-PE antibodies were obtained from R&D Systems, USA. The cells (1 × 106 cells) were washed twice with FACS buffer and stained for surface markers by incubation with the anti-human antibodies for 20 min in the dark at 4°C. For additional intracellular staining, cells were fixed and permeabilized using a fix/perm kit (BD Biosciences, Heidelberg, Germany) according to the manufacturer’s instructions. The FACS Calibur from (BD Biosciences, New York, USA) was used to detect fluorescence.
Cytokines Measurement
IFN-γ and IL10 levels were detected using ELISA kits (BD Biosciences New York, USA), and IL17A levels were measured using an ELISA kit (R&D Systems New York, USA) as directed by the manufacturer.
Quantitative Real-Time PCR
The mRNA of 4×106 cells was isolated after performing cell lysis by 1 mL TRIzol. After performing cell lysis by 1 mL Tri reagent (Thermo Fisher, Waltham, Massachusetts, USA), mRNA of 4×106 cells was isolated. The mRNA was further transcribed to cDNA using a cDNA Synthesis Kit (Quantabio, USA), as directed by the manufacturer’s protocol. Quantitative real-time PCR was performed using a Step-One Plus real-time PCR System with Power SYBR Green (Thermo Fisher Scientific, USA). The following oligonucleotide sequences were used: house-keeping gene PBGD, (forward) 5-′ACGATCCCGAGACTCTGCTTC-3′ and (reverse) 5′- GCACGGC-TACTGG-CACACT-3′; Th1 cell-specific Tbet, (forward) 5-′AGGGACGGCGGATGTTCCCA-3′ and (reverse) 5′-GC TGCCCTCGGCCTTTCCAC-3′; Th17 cell-specific RORC2, (forward) 5-CAGTCATGAGAACACAAATTGAAGTG-3′ and (reverse) 5′-CAGGTGATAACCCCGTAGTGGAT-3; and Tregs-specific Foxp3, (forward) 5-CACCTGGC TGGGAAAATGG-3′ and (reverse) 5′-GGAGCCCTTGTCGGATGAT-3′. Each PCR sample was diluted to a final volume of 20 μL using distilled H2O (6 μL), SYBR Select PCR Master Mix10 μL (Thermo Fisher Scientific, Waltham, Massachusetts, USA), 1 μL of both forward and reverse primers (4 μM), and 2 μL (50 ng/mL) or a negative control distilled water. PCR was done as duplicates with cycling parameters as follows: 95°C for 5 min, followed by 40 cycles of 94°C for 20s, and 60°C for 30s. For quantification, comparative cycle threshold method (ΔΔCT) was used to normalize the results to the PBGD housekeeping gene.Citation27
Statistical Analysis
The statistical significance was calculated by Student’s t-test using the GraphPad Prism software (version 5.01, USA).
Results
Effect of Zinc on Tregs and Th17 Cells in the Mixed Lymphocyte Cultures
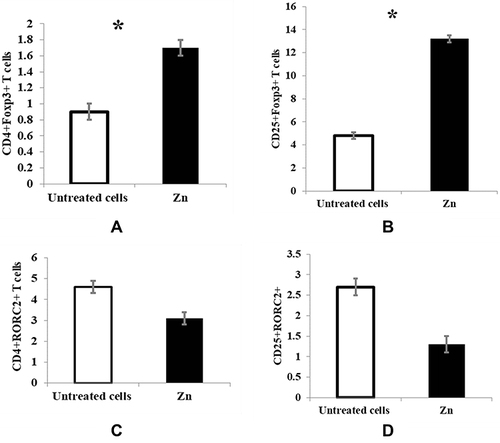
Regarding the effect of zinc on the Tregs population and Th17 cells, the cells were adjusted as 2×106 cells, next incubated with 50 μM zinc for 15 minutes preceding MLC generation and following five days of MLC incubation, Tregs and Th17 cells% were analyzed by flow cytometry. Zinc supplementation at a physiological dose (50 μM) significantly increased the number of CD4+ Foxp3+ Tregs and CD25+ Foxp3+ Tregs (p < 0.05) compared with untreated T cells ( and ). In contrast, zinc treatment insignificantly decreased the number of CD4+ RORC2+ and CD25+ RORC2+ Th17 cells ( and ).
Figure 1 Effect of zinc on Tregs and Th17 cells in MLC. The cells were adjusted as 2×106 cells, next incubated with 50 μM zinc for 15 minutes preceding MLC generation, and following five days of MLC incubation, Tregs and Th17 cells% were analyzed by flow cytometry. Untreated cells (white bars) and cells incubated with 50 μM zinc (black bars) are represented in the graphs. (A) CD4+Foxp3+ T cells, p = 0.031 (B) CD25+Foxp3+ T cells, (p = 0.043) (C) CD4+RORC2+ T cells, (p = 0.083) (D) CD25+RORC2+ T cells, (p = 0.097). Results are presented as mean values ± SEM. *Significance (p < 0.05).
Effect of Zinc Supplementation on Cytokines
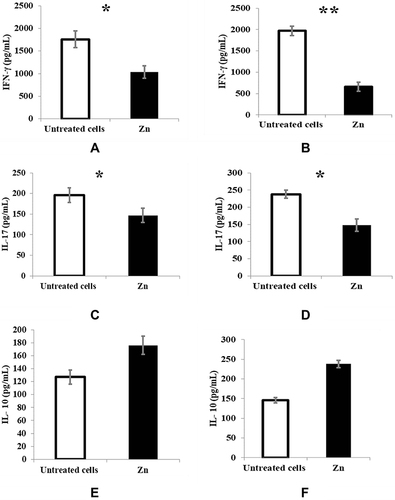
Regarding cytokine measurement by ELISA, zinc supplementation significantly reduced IFNγ production in both restimulated T cells with autologous PBMC (p < 0.05) () and allogeneic PBMC (p < 0.01) () compared with untreated T cells. Zinc also significantly reduced IL17 (p < 0.05) ( and ), but the IL10 increase was insignificant ( and ).
Figure 2 Effect of zinc supplementation on IFNγ, IL17 & IL10 production as measured by ELISA. Untreated restimulated T cells (white bars) and treated cells with 50μm zinc (black bars) are represented in the graphs. For each cytokine, (A, C, E) restimulated T cells with autologous PBMC & (B, D, F) Restimulated T cells with allogeneic PBMC. (A) p = 0.044. (B) p = 0.001. (C) p = 0.023. (D) p = 0.041. (E) p = 0.073. (F) p = 0.084. Results are presented as mean values ± SEM. *Significance (p < 0.05), **significance (p < 0.01).
The mRNA Expression of Tbet for Th1, RORC2 for Th17 and Foxp3+ for Treg Cells
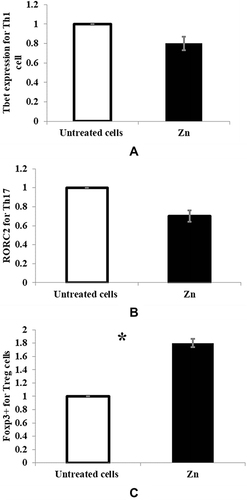
Analysis of Th1 cell-specific Tbet expression, Th17 cell-specific RORC2 expression, and Tregs-specific Foxp3 expression by real-time PCR showed an insignificant decrease in Th1 or Th17 cell-specific transcription factor mRNA expression in zinc-supplemented MLC ( and ), while a significant increase in Tregs-specific transcription factor mRNA expression (p < 0.05) was observed in zinc-supplemented MLC (). Untreated control MLC was used to normalize the results.
Figure 3 The mRNA expression of (A) Tbet for Th1 cell, p = 0.068 (B) RORC2 for Th17 cell, p = 0.075 and (C) Foxp3+ for Tregs, p = 0.018 were measured by real-time PCR. Untreated MLC (white bars) and treated cells with 50μm zinc (black bars) are represented in the graphs. *Significance of p < 0.05.
Discussion
Apart from protecting the body from various pathogens, paradoxically, immune cells also contribute to different health issues, such as autoimmune diseases and graft rejection; therefore, the involvement of these cells and their interactions should be of prime interest. For instance, Tregs and Th17 are essential antagonists for a well-balanced immune reaction; the former establishes immune tolerance, while the latter triggers a pro-inflammatory immune reaction.Citation28 Various factors can modulate immune cell differentiation and function, including infection, smoking, and diet.Citation1
Zinc is a micronutrient that affects the integrity of the immune system. T-cell stimulation and development are zinc-dependent process, and Zn deficiency leads to thymus atrophy with lymphopenia.Citation5 As zinc is critical for T cell function, this study aimed to determine the influence of zinc supplementation on T cell subpopulations: Tregs, Th1, and Th17 cells in mixed lymphocyte cultures in an in vitro transplantation model.
The effect of zinc on the population of Tregs and Th17 cells was studied using flow cytometry, and our findings are based on the fact that RORC2+ and Foxp3+ T cells are two non-dependent and mutually exclusive cell subpopulations. It was found that zinc supplementation at a physiological dose significantly increased the number of CD4+ Foxp3+ Tregs and CD25+ Foxp3+ Tregs (p < 0.05). In contrast, zinc treatment decreased the number of CD4+ RORC2+ and CD25+ RORC2+ Th17 cells, but this decrease was insignificant. Some previous studies have obtained similar results; they revealed that physiological doses of zinc enhanced the number of Tregs without suppressing the immune system and slightly affecting the Th17 number. Also, it has been found that zinc can induce IFNγ expression, while IL2 and IL10 did not affect it.Citation23,Citation29,Citation30
Graft versus host disease is a major challenge, and cytokine levels, such as IFN-γ, are sensitive indicators of graft rejection in MLC, hence correlating with the severity of acute GVHD.Citation31–33 For that reason, zinc can suppress allogeneic immune responses because of its ability to stabilize induced Tregs.Citation24 In our study, pro-inflammatory cytokine IFNγ production was significantly reduced in both zinc-treated restimulated T cells with autologous PBMC (p < 0.05) and allogeneic PBMC (p < 0.01) compared with untreated T cells. Faber et al confirmed that zinc reduces IFNγ in allogeneic reactions.Citation34 Other researchers found beneficial effects of zinc supplementation in transplantation, as zinc in a dose-dependent way reduced allograft rejection in allogeneic rodent cardiac transplantation modelsCitation35,Citation36 and maintained functional grafts in intraportal-islets in rat recipients,Citation37 suggesting either reduced production of pro-inflammatory cytokines or inhibited allograft-cell apoptosis as a possible reason. Zinc can inhibit Th1 cell proliferation and, consequently, inhibit pro-inflammatory IFNγ expression in Th1-motivated allogeneic immune reactions. Another explanation for the inhibition of IFNγ expression is a novel molecular mechanism by which zinc can induce KLF-10 (Krüppel-like factor-10) and stabilize Foxp3 transcription factors that positively correlate with iTregs stabilization and negatively correlates with IRF-1 (interferon regulatory factor-1), consequently inhibiting IFNγ production.Citation19 Subsequently, zinc not only stabilizes iTregs in MLC but also diminishes IFNγ pro-inflammatory cytokine production and consequently suppresses allogeneic immune reactions.
IL17 is another pro-inflammatory cytokine that is produced by Th 17 cells. It plays a role in the clearance of different pathogens, rejection of solid organ allografts, and several autoimmune and inflammatory disorders.Citation38 In our study, we found that zinc also significantly reduces IL17 (p < 0.05) in both, restimulated T cells with autologous PBMC and allogeneic PBMC compared with untreated T cells. Similarly, zinc was found to interfere with Th17 cell activation and, consequently, IL17 expression in vitro.Citation39 Significantly, zinc suppresses IL17 due to its inhibitory effect on the IL6/STAT3 (signal transducer and activator of transcription) signaling pathway, which is an important step in the Th17 cell development.Citation40 Moreover, a study found that zinc deficiency can promote Th17 cell polarization and Treg cell dysfunction, which makes zinc a promising therapeutic approach to reducing autoimmune diseases associated with Th17 cells.Citation41
Tregs can also produce immunosuppressive cytokines such as IL10, which play a role in inflammation resolution, synthesis of Th1 inflammatory cytokines, and inhibition of macrophages and dendritic cells.Citation42 In this study, it was found that zinc slightly increases IL10 in both restimulated T cells with autologous PBMC and allogeneic PBMC compared with untreated T cells. In agreement with this finding, Rosenkranz et al reported similar result.Citation23 Increasing IL10 level is beneficial as it negatively affects GVHD development and protects against acute constant rejection.Citation20,Citation43 At the molecular level, we observed a non-significant decrease in Th1 or Th17 cells-specific transcription factors mRNA expression (Tbet and RORC2, respectively), while a significant increase in Tregs (Foxp3)-specific transcription factor mRNA expression (p < 0.05) was observed in zinc-supplemented MLC. Parallel results were observed by Maywald et al, who concluded that zinc addition selectively has a positive effect on the Tregs population in MLC through stimulating FoxP3 activity and damping the suppressed impact of Interferon Regulatory Factor 1 (IRF-1) on FoxP3.Citation24 Additionally, other researchers observed that zinc significantly suppressed pre-activated human Th1 and Th17 cell proliferation and cytokine (IFNγ and IL17) production in vitro.Citation15 Comparable research examined the suppressed impact of zinc on Th17, they found that zinc can inhibit IL-6 and IL-1β-mediated signaling pathways which negatively impacts Th17 differentiation.Citation25
Conclusions
In conclusion, zinc could induce and stabilize Tregs, which play a basic role in adverse immunological reactions or in vitro transplantation models. Therefore, zinc might be considered a promising candidate for clinical treatment of T cell-induced disorders, such as allergy, autoimmune disease, and transplantation. However, this finding should be confirmed in in vivo studies.
Data Sharing Statement
The original contributions of this study are included in this article. Further inquiries can be directed to the corresponding authors.
Ethical Approval
Ethical approval was obtained from the Scientific Research Ethics Committee of the Faculty of Applied Medical Sciences, Taibah University (approval number 2023/144/101 MLT). All procedures were carried out in accordance with the principles of the Helsinki Declaration. Written informed consent was obtained from all patients.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no conflicts of interest in this work.
Additional information
Funding
References
- Calder PC. Feeding the immune system. Proce Nutrition Soc. 2013;72(3):299–309. doi:10.1017/s0029665113001286
- Ibs KH, Rink L. Zinc-altered immune function. J Nutr. 2003;133(5):1452S–1456S. doi:10.1093/jn/133.5.1452s
- Walker CF, Black RE. Zinc and the risk for infectious disease. Annu Rev Nutr. 2004;24(1):255–275. doi:10.1146/annurev.nutr.23.011702.073054
- Plum LM, Rink L, Haase H. The essential toxin: impact of zinc on human health. Int J Environ Res Public Health. 2010;7(4):1342–1365. doi:10.3390/ijerph7041342
- Haase H, Rink L. Functional significance of zinc-related signaling pathways in immune cells. Annu Rev Nutr. 2009;29(1):133–152. doi:10.1146/annurev-nutr-080508-141119
- van der Laan M, Büttgenbach A, Wolf J, Rink L, Wessels I. The role of zinc in GM-CSF-induced signaling in human polymorphonuclear leukocytes. Mol Nutr Food Res. 2022;66(14):2101106. doi:10.1002/mnfr.202101106
- Peng‐Winkler Y, Wessels I, Rink L, Fischer HJ. Zinc levels affect the metabolic switch of T cells by modulating glucose uptake and insulin receptor signaling. Mol Nutr Food Res. 2022;66(9):2100944. doi:10.1002/mnfr.202100944
- Schmitt AK, Puppa MA, Wessels I, Rink L. Vitamin D3 and zinc synergistically induce regulatory T cells and suppress interferon-γ production in mixed lymphocyte culture. J Nutr Biochem. 2022;102:108942. doi:10.1016/j.jnutbio.2022.108942
- Wessels I, Fischer HJ, Rink L. Dietary and physiological effects of zinc on the immune system. Annu Rev Nutr. 2021;41(1):133–175. doi:10.1146/annurev-nutr-122019-120635
- Cousins RJ, Liuzzi JP, Lichten LA. Mammalian zinc transport, trafficking, and signals. J Biol Chem. 2006;281(34):24085–24089. doi:10.1074/jbc.r600011200
- Zheng SG, Wang JH, Stohl W, Kim KS, Gray JD, Horwitz DA. TGF-β requires CTLA-4 early after T cell activation to induce FoxP3 and generate adaptive CD4+CD25+ regulatory cells. J Immunol. 2006;176(6):3321–3329. doi:10.4049/jimmunol.176.6.3321
- Capone A, Volpe E. Transcriptional regulators of T helper 17 cell differentiation in health and autoimmune diseases. Front Immunol. 2020;11:348. doi:10.3389/fimmu.2020.00348
- Kleinewietfeld M, Manzel A, Titze J, et al. Sodium chloride drives autoimmune disease by the induction of pathogenic TH17 cells. Nature. 2013;496(7446):518–522. doi:10.1038/nature11868
- Abadja F, Sarraj B, Ansari MJ. Significance of T helper 17 immunity in transplantation. Curr Opin Organ Transplant. 2012;17(1):8–14. doi:10.1097/MOT.0b013e32834ef4e4
- Guttek K, Wagenbrett L, Reinhold A, Grüngreiff K, Reinhold D. Zinc aspartate suppresses proliferation and Th1/Th2/Th17 cytokine production of pre-activated human T cells in vitro. J Trace Elements Med Biol. 2018;49:86–90. doi:10.1016/j.jtemb.2018.05.003
- Schmidt A, Oberle N, Krammer PH. Molecular mechanisms of treg-mediated T cell suppression. Front Immunol. 2012;3:51. doi:10.3389/fimmu.2012.00051
- Workman CJ, Szymczak-Workman AL, Collison LW, Pillai MR, Vignali DAA. The development and function of regulatory T cells. Cell Mol Life Sci. 2009;66(16):2603–2622. doi:10.1007/s00018-009-0026-2
- Szymczak-Workman AL, Workman CJ, Vignali DAA. Cutting edge: regulatory T cells do not require stimulation through their TCR to suppress. J Immunol. 2009;182(9):5188–5192. doi:10.4049/jimmunol.0803123
- Szymczak-Workman AL, Delgoffe GM, Green DR, Vignali DAA. Cutting edge: regulatory T cells do not mediate suppression via programmed cell death pathways. J Immunol. 2011;187(9):4416–4420. doi:10.4049/jimmunol.1100548
- Zheng HX, Burckart GJ, McCurry K, et al. Interleukin-10 production genotype protects against acute persistent rejection after lung transplantation. J Heart Lung Trans. 2004;23(5):541–546. doi:10.1016/s1053-2498(03)00303-6
- Li XC, Turka LA. An update on regulatory T cells in transplant tolerance and rejection. Nat Rev Nephrol. 2010;6(10):577–583. doi:10.1038/nrneph.2010.101
- Gan X, Gu J, Ju Z, Lu L. Diverse Roles of Immune Cells in Transplant Rejection and Immune Tolerance. Engineering. 2022;10:44–56. doi:10.1016/j.eng.2021.03.029
- Rosenkranz E, Metz CHD, Maywald M, et al. Zinc supplementation induces regulatory T cells by inhibition of Sirt-1 deacetylase in mixed lymphocyte cultures. Mol Nutr Food Res. 2015;60(3):661–671. doi:10.1002/mnfr.201500524
- Maywald M, Wessels I, Zinc Signals RL. Immunity. Int J Mol Sci. 2017;18(10):2222. doi:10.3390/ijms18102222
- Kim B, Lee WW. Regulatory role of zinc in immune cell signaling. Mol Cells. 2021;44(5):335–341. doi:10.14348/molcells.2021.0061
- Atif M, Conti F, Gorochov G, Oo YH, Miyara M. Regulatory T cells in solid organ transplantation. Clin Transl Immunol. 2020;9(2). doi:10.1002/cti2.1099
- Maywald M, Rink L. Zinc supplementation induces CD4+CD25+Foxp3+ antigen-specific regulatory T cells and suppresses IFN-γ production by upregulation of Foxp3 and KLF-10 and downregulation of IRF-1. Eur J Nutr. 2016;56(5):1859–1869. doi:10.1007/s00394-016-1228-7
- Kleinewietfeld M, Hafler DA. The plasticity of human Treg and Th17 cells and its role in autoimmunity. Semin Immunol. 2013;25(4):305–312. doi:10.1016/j.smim.2013.10.009
- Dünkelberg S, Maywald M, Schmitt AK, Schwerdtle T, Meyer S, Rink L. The interaction of sodium and zinc in the priming of T cell subpopulations regarding Th17 and Treg cells. Mol Nutr Food Res. 2020;64(2):1900245. doi:10.1002/mnfr.201900245
- Wu C, Yosef N, Thalhamer T, et al. Induction of pathogenic TH17 cells by inducible salt-sensing kinase SGK1. Nature. 2013;496(7446):513–517. doi:10.1038/nature11984
- Dickinson A. Predicting graft-versus-host disease in allogeneic bone marrow transplant recipients. Transpl Immunol. 1994;2(2):154–155. doi:10.1016/0966-3274(94)90051-5
- Danzer SG, Rink L. Reverse transcription polymerase chain reaction for cytokines in the mixed lymphocyte culture. Is there a role in renal transplantation? Nephrol Dialysis Trans. 1996;11(7):1246–1247. doi:10.1093/ndt/11.7.1246
- van der Meer A, Wissink WM, Schattenberg AVMB, Joosten I. Interferon-gamma-based mixed lymphocyte culture as a selection tool for allogeneic bone marrow donors other than identical siblings. Br J Haematol. 1999;105(2):340–348. doi:10.1111/j.1365-2141.1999.01342.x
- Faber C, Gabriel P, Ibs KH, Rink L. Zinc in pharmacological doses suppresses allogeneic reaction without affecting the antigenic response. Bone Marrow Transplant. 2004;33(12):1241–1246. doi:10.1038/sj.bmt.1704509
- Kown MH, van der Steenhoven T, Blankenberg FG, et al. Zinc-mediated reduction of apoptosis in cardiac allografts. Circulation. 2000;102(Supplement3):III-228-III–232. doi:10.1161/01.cir.102.suppl_3.iii-228
- Kown MH, van der Steenhoven TJ, Jahncke CL, et al. Zinc chloride–mediated reduction of apoptosis as an adjunct immunosuppressive modality in cardiac transplantation. J Heart Lung Trans. 2002;21(3):360–365. doi:10.1016/s1053-2498(01)00384-9
- Okamoto T, Kuroki T, Adachi T, et al. Effect of Zinc on early graft failure following intraportal islet transplantation in rat recipients. Ann Transplant. 2011;16(3):114–120. doi:10.12659/aot.882003
- Crome SQ, Wang AY, Levings MK. Translational mini-review series on Th17 cells: function and regulation of human T helper 17 cells in health and disease. Clin Exp Immunol. 2010;159(2):109–119. doi:10.1111/j.1365-2249.2009.04037.x
- Reda R, Abbas AA, Mohammed M, et al. The Interplay between Zinc, Vitamin D and, IL-17 in Patients with Chronic Hepatitis C Liver Disease. J Immunol Res. 2015;2015:846348. doi:10.1155/2015/846348
- Overbeck S, Rink L, Haase H. Modulating the immune response by oral zinc supplementation: a single approach for multiple diseases. Arch Immunol Ther Exp (Warsz). 2008;56(1):15–30. doi:10.1007/s00005-008-0003-8
- Kulik L, Maywald M, Kloubert V, Wessels I, Rink L. Zinc deficiency drives Th17 polarization and promotes loss of Treg cell function. J Nutr Biochem. 2019;63:11–18. doi:10.1016/j.jnutbio.2018.09.011
- Foster M, Samman S. Zinc and regulation of inflammatory cytokines: implications for cardiometabolic disease. Nutrients. 2012;4(7):676–694. doi:10.3390/nu4070676
- Xiaoping J, Wang J, Xu B, Cao Y, Lü S. Interleukin-10 and graft-versus-host disease. N Eng J Med. 2004;350(13):1361–1362. doi:10.1056/nejmc033110