
Abstract
Purpose
Eggshell membrane (ESM) has been shown to contain naturally occurring bioactive components, and biological activities such as reducing proinflammatory cytokines, liver fibrosis, and joint pain in osteoarthritis sufferers have also been reported for ESM matrix as a whole. Nuclear factor kappa-light-chain-enhancer of activated B-cells (NF-κB) is a signaling protein found in the cytoplasm of nearly all human and animal cell types and is a primary regulator of immune function. The studies reported herein were designed to investigate the possible role that NF-κB activity might play in the reported biological activities of ESM.
Methods
Three ESM hydrolyzates produced via fermentation, enzymatic, or chemical hydrolysis were evaluated in vitro in either human peripheral blood mononuclear cell or THP-1 (human leukemic monocyte) cell cultures for NF-κB activity following 4-hour exposure. The hydrolyzates were compared with untreated control cells or cells incubated with lipopolysaccharide or ascorbic acid. The source of ESM activity was also evaluated.
Results
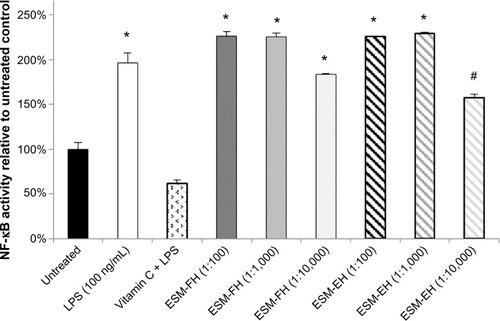
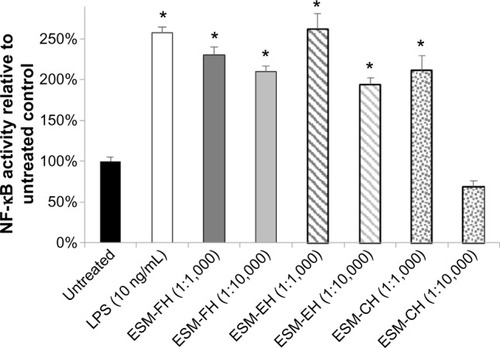
NF-κB levels were increased above levels found in untreated cells at all three dilutions (1:100, 1:1,000, and 1:10,000) for the fermentation hydrolyzate of ESM (ESM-FH) (P=0.021, P=0.020, P=0.009, respectively) in peripheral blood mononuclear cells. The enzymatic hydrolyzate of ESM (ESM-EH) also produced statistically significant levels of activated NF-κB at the 1:100 and 1:1,000 dilutions (P=0.004, P=0.006, respectively) but fell just shy of significance at the 1:10,000 dilution (P=0.073). Similarly, ESM-FH (P=0.021, P=0.002) and ESM-EH (P=0.007, P=0.007) activated NF-κB in THP-1 cells at 1:1,000 and 1:10,000 dilutions, respectively. The chemical hydrolyzate of ESM (ESM-CH) showed statistically significant levels of activation at the 1:1,000 dilution (P=0.005) but failed to differ from untreated cells at the 1:10,000 dilution (P=0.193) in THP-1 cells.
Conclusion
Results from our studies provide evidence that ESM hydrolyzates significantly activate NF-κB, and the source of this activity was investigated to confirm that it is inherent to ESM and not derived from bacterial contamination. Based on our findings, we propose a plausible hypothesis as to how increased NF-κB activity might translate into the in vivo efficacy that has been observed with ESM via an “oral tolerance” mechanism.
Introduction
Eggshell membrane (ESM), found between the calcified shell and the albumin in chicken eggs, is primarily composed of fibrous proteins such as collagen type ICitation1 that form the mesh-like structure of the bilayered material. ESMs have also been shown to contain other bioactive components, namely glycosaminoglycans (ie, dermatan sulfate,Citation2 chondroitin sulfate,Citation2 and hyaluronic acidCitation3). ESM is known to reduce the expression of various proinflammatory cytokines, including interleukin-1-beta (IL-1β) and tumor necrosis factor-alpha (TNF-α) both in vitroCitation4 and in vivo.Citation5 A proprietary form of ESM, commercially available as the branded product NEM®, has demonstrated safety and efficacy in multiple clinical trials in relieving joint pain and stiffness in humans with osteoarthritisCitation6–Citation8 and has been investigated for similar uses in various species of animals.Citation9,Citation10 ESM is also reported to have possible beneficial effects in liver fibrosis in rats through regulating gene expression.Citation11
Nuclear factor kappa-light-chain-enhancer of activated B-cells (NF-κB) is a protein complex found in the cytoplasm of nearly all human and animal cell types; the deoxyribonucleic acid (DNA)-binding activities of this family of transcription factors were first reported in 1986.Citation12 Once activated and translocated to the nucleus, NF-κB regulates the expression of multiple genes involved in the acute response to negative or harmful external cellular stimuli. Active NF-κB is a heterodimeric protein that activates DNA sequences known collectively as κB sites (see Citation13 for a detailed overview). There are two well-accepted paths to activation of NF-κB: the canonical or classical pathway and the noncanonical or alternative pathway. In either pathway (canonical or non-canonical), the inhibitor of κB kinase complex is activated when signaling molecules bind to cell surface receptors. Bacterial or viral antigens,Citation14,Citation15 various cytokines (eg, IL-1β, TNF-α),Citation16,Citation17 oxidative stress,Citation18,Citation19 ultraviolet irradiation,Citation20,Citation21 and free radicalsCitation22,Citation23 are just some of the stimuli known to trigger activation of one or the other of the NF-κB pathways, either through direct binding to cell surface receptors or by causing other signaling molecules to be formed, which then bind to the receptors.
NF-κB was initially investigated for its critical role in regulating the immune response to infection. However, in the decades since its discovery, the dysregulation of NF-κB has been associated with numerous classical inflammatory diseases such as sepsis, asthma, rheumatoid arthritis, and inflammatory bowel disease (see Citation24–Citation27 for a review). Interestingly, a number of diseases that are not obviously inflammatory in nature are also associated with NF-κB dysregulation, including atherosclerosis,Citation28,Citation29 Alzheimer’s disease,Citation30,Citation31 multiple sclerosis,Citation32,Citation33 diabetes,Citation34,Citation35 and various cancers.Citation36–Citation38
Because of its fundamental importance in immune system function and inflammatory diseases, the studies reported herein were designed to investigate the possible role that NF-κB activity might play in the reported biological activities of ESM.
Materials and methods
Reagents
All chemicals and reagents were used as received unless otherwise indicated. High-performance liquid chromatography (HPLC) grade water, sodium bicarbonate, sodium carbonate monohydrate, sodium hydroxide, and filter paper (P4 grade, medium-fine porosity) were obtained from Fisher Scientific (Pittsburgh, PA, USA). Purified alkaline protease from Bacillus licheniformis was obtained from Bio-Cat, Inc. (Troy, VA, USA). ESM (from Gallus gallus) used in the preparation of ESM hydrolyzates was obtained from ESM Technologies, LLC (Carthage, MO, USA). Phosphate-buffered saline (PBS, pH 7.4), Roswell Park Memorial Institute-1640 culture medium, fetal bovine serum (FBS), L-glutamine 200 mM, penicillin–streptomycin 100X solution, glacial acetic acid, and methanol were obtained from Sigma-Aldrich Co (St Louis, MO, USA). TransAM® NF-κB p65 enzyme-linked immunosorbent assay (ELISA) kits were obtained from Active Motif (Carlsbad, CA, USA). Bradford method protein assay kits were obtained from Bio-Rad Laboratories Inc. (Hercules, CA, USA).
Preparation of ESM hydrolyzates
Fermentation hydrolyzate
ESM powder (2.8 g) was suspended in 50 mL of purified water (HPLC grade) in a 100 mL round-bottomed flask, followed by the addition of 1.4 g of sucrose and 230 μL of a proprietary inoculum of bacteria and yeasts. The slurry was allowed to ferment at room temperature (~25°C) for 6 hours with moderate stirring (~500 rpm). The slurry was vacuum filtered to remove unreacted solids. The resulting clear, light brown solution contains 5%–7% dissolved solids and was divided into ~1 mL aliquots, which were stored frozen (−20°C) until later use in the bioassays. This preparation is referred to as fermentation hydrolyzate of ESM (ESM-FH).
Enzymatic hydrolyzate
ESM powder (1.0 g) was suspended in a buffer mixture (pH 9.3) comprising 15 mL of 0.1 M sodium bicarbonate and 5 mL of 0.1 M sodium carbonate in a 50 mL round-bottomed flask. Alkaline protease powder (20% w/w to ESM) was added, and the slurry was allowed to stir (∼500 rpm) at 60°C for 5 days. The slurry was vacuum filtered to remove unreacted solids. The resulting clear, light brown solution contains 5%–7% dissolved solids and was divided into ∼1 mL aliquots, which were stored frozen (−20°C) until later use in the bioassays. This preparation is referred to as enzymatic hydrolyzate of ESM (ESM-EH).
Chemical hydrolyzate
The hydrolysis was performed according to the method of Strohbehn et al.Citation39 Briefly, ESM powder (1.0 g) was suspended in a 5% (w/v) sodium hydroxide solution in a 25 mL round-bottomed flask. The slurry was allowed to stir (∼500 rpm) at 50°C for 4 hours. The mixture was allowed to cool and was vacuum filtered to remove any eggshell. The pH of the filtrate was adjusted to ∼7 with a 5% (w/v) acetic acid solution while keeping the mixture temperature at ≤4°C. The solution was desalted using a dialyzing membrane with two to three equal volumes of water. The resulting clear, pale yellow solution was adjusted to contain 5%–7% dissolved solids and was divided into ∼1 mL aliquots, which were stored frozen (−20°C) until later use in the bioassays. This preparation is referred to as chemical hydrolyzate of ESM (ESM-CH).
Cell source and culture conditions
Human peripheral blood mononuclear cells (PBMCs) were obtained from Astarte Biologics, LLC (Bothell, WA, USA). PBMCs were cultured at 3.2×106 cells/well in culture medium (RPMI-1640 medium supplemented with 10% FBS, L-glutamine [2 mM], penicillin [100 U/mL], and streptomycin [100 mg/mL]). The human leukemic monocyte cell line THP-1 was obtained from American Type Culture Collection (Manassas, VA, USA) and was cultured at 2.0×105 cells/well in culture medium (RPMI-1640 medium supplemented with 10% FBS, 2-mercaptoethanol [0.05 mM], and 1% penicillin–streptomycin). Both PBMC and THP-1 cell cultures were utilized for the subsequent determination of NF-κB activity.
Evaluation of the in vitro NF-κB activity of ESM hydrolyzates
The in vitro NF-κB activity of ESM hydrolyzates was initially evaluated in human PBMCs utilizing the facilities and services of NIS Labs (Klamath Falls, OR, USA). PBMCs were plated at a density of 3.2×106 cells/well in culture medium in 12-well culture plates (growth area 3.8 cm2/well). The primary cells were plated in duplicate in two independent experiments (n=2 per condition). Aliquots of ESM hydrolyzates (ESM-FH or ESM-EH) were allowed to thaw and were diluted 1:1 with PBS prior to filtering through a 0.22 micron cellulose acetate disc to sterilize them. An additional two tenfold dilutions of the ESM hydrolyzates were prepared with culture medium. Each dilution (60 μL) was added to a culture well (3 mL total volume), resulting in final ESM hydrolyzate dilutions of 1:100, 1:1,000, and 1:10,000, and the cells were incubated at 37°C, 5% CO2 for 4 hours. Not knowing whether ESM hydrolyzates would be activating or inhibitory, a known activator of NF-κB, lipopolysaccharide (LPS; 100 ng/mL), was used as a positive control, and a known inhibitor of NF-κB, vitamin C (ascorbic acid; 20 mM),Citation40 was used as a comparator of the inhibitory effect on LPS-treated cells. Activity was also compared with untreated cells. Cells were pelleted by centrifuging at 150 x g (1,200 rpm) for 3 minutes and the supernatant was discarded. Cell lysates were prepared from the incubated cultures via bead milling. The whole cell lysates were assayed for NF-κB activity via a commercially available ELISA kit according to the manufacturer’s instructions. Samples were assayed in duplicate and were normalized for total protein content via the Bradford method according to the manufacturer’s instructions.
The determination of the in vitro NF-κB activity of ESM hydrolyzates in THP-1 cells was performed utilizing the facilities and services of the Jordan Valley Innovation Center within the Center for Biomedical and Life Sciences at Missouri State University (JVIC-MSU; Springfield, MO, USA). THP-1 cells were incubated in suspension for 3–5 days and were subsequently plated at a cell density of 2.0×105 cells/well in culture medium in 24-well culture plates (growth area 1.9 cm2/well). The cells were plated using a volume of 350 μL per well in duplicate in three independent experiments (n=3 per condition). Aliquots of ESM hydrolyzates (ESM-FH, ESM-EH, or ESM-CH) were allowed to thaw prior to filtering through a 0.2 micron polyethersulfone disc to sterilize them. The ESM hydrolyzates were then diluted (1:1,000; 1:10,000) with culture medium. Each dilution was added to a culture well (3 mL total volume), and the cells were incubated at 37°C, 5% CO2 for 4 hours. Because ESM hydrolyzates were found to be activating in PBMCs, only LPS (10 ng/mL) was used as a positive control. Activity was again compared with untreated cells. Cells were pelleted by centrifuging at 150 x g (1,200 rpm) for 3 minutes and the supernatant was discarded. Cell lysates were prepared from the incubated cultures by adding 45 μL of complete lysis buffer (containing dithiothreitol and protease inhibitor). The whole cell lysates were assayed for NF-κB activity via a commercially available ELISA kit according to the manufacturer’s instructions. Samples were assayed in duplicate and were normalized for total protein content via the Bradford method according to the manufacturer’s instructions.
Evaluation of the ESM source of NF-κB activation
An initial evaluation of the source of NF-κB activity was performed utilizing the facilities of NIS Labs using respective appropriate controls. PBMCs (prepared as described previously) were exposed to aliquots of ESM hydrolyzate or control solutions for 4 hours prior to NF-κB activity determination. ESM-FH NF-κB activity was compared with untreated cells and cells treated with the fermentation broth (minus ESM). Similarly, the ESM-EH was compared with untreated cells and cells treated with the enzyme digest solution alone. A further, more discerning, evaluation of the source of in vitro NF-κB activity of ESM hydrolyzates in THP-1 cells was performed utilizing the facilities and services of JVIC-MSU. THP-1 cells were prepared as described previously. Aliquots of ESM hydrolyzates (ESM-FH, ESM-EH, or ESM-CH) were allowed to thaw prior to filtering through a 0.2 micron polyethersulfone disc to sterilize them. The ESM hydrolyzates were then diluted (1:1,000; 1:10,000) with culture medium. Each dilution was added to a culture well as well as separate wells for hydrolyzate + polymyxin B (10 mg/mL) and hydrolyzate + lipoprotein lipase (10 mg/mL). The cells were incubated at 37°C, 5% CO2 for 4 hours. LPS (10 ng/mL) was again used as a positive control. Activity was compared with untreated cells, cells treated with polymyxin B only, cells treated with LPS + polymyxin B, and cells treated with lipoprotein lipase only. Cell cultures were processed and whole cell lysates were analyzed as described previously.
Statistical analysis
Statistical analyses were done either as a two-tailed, independent group Student’s t-test or a Kruskal–Wallis test for multiple comparisons. Statistical significance was accepted at a P-value of P<0.05. SYSTAT software (version 13) was used for all statistical analyses.Citation41
Results
Evaluation of the in vitro NF-κB activity of ESM hydrolyzates
There were statistically significant levels of NF-κB activation versus untreated cells at all three dilutions (1:100, 1:1,000, and 1:10,000) for the ESM-FH (P=0.021, P=0.020, P=0.009, respectively) in PBMCs. The ESM-EH also showed statistically significant levels of activation at the 1:100 and 1:1,000 dilutions (P=0.004, P=0.006, respectively) but fell just shy of significance at the 1:10,000 dilution (P=0.073). Both the fermentation and enzymatic hydrolyzates of ESM at the 1:100 and 1:1,000 dilutions activated NF-κB in human PBMCs comparably with LPS at 100 ng/mL ().
Figure 1 NF-κB activity of fermentation and enzymatic hydrolyzates of ESM in human PBMCs. Human PBMCs were incubated for 4 hours with either medium (untreated control), LPS, vitamin C + LPS, or each dilution of ESM hydrolyzate. Whole cell lysates were then analyzed for NF-κB activity after normalizing for protein content. Bars represent mean ± standard error of the mean; values are presented relative to untreated control.
Notes: *P<0.05 versus untreated control; #P<0.10 versus untreated control.
Abbreviations: NF-κB, nuclear factor kappa-light-chain-enhancer of activated B-cells; ESM, egg shell membrane; PBMCs, peripheral blood mononuclear cells; LPS, lipopolysaccharide; ESM-FH, ESM fermentation hydrolyzate; ESM-EH, ESM enzymatic hydrolyzate.
Similarly, there were statistically significant levels of NF-κB activation versus untreated cells at both dilutions (1:1,000 and 1:10,000) for the fermentation and enzymatic hydrolyzates of ESM (ESM-FH: P=0.021, P=0.002; ESM-EH: P=0.007, P=0.007, respectively) in THP-1 cells. The ESM-CH also caused statistically significant levels of activation at the 1:1,000 dilution (P=0.005) but failed to differ from untreated cells at the 1:10,000 dilution (P=0.193). Both the fermentation and enzymatic hydrolyzates of ESM at the 1:1,000 dilution activated NF-κB in human THP-1 cells comparably with LPS at 10 ng/mL ().
Figure 2 NF-κB activity of fermentation, enzymatic, and chemical hydrolyzates of ESM in the human THP-1 monocyte cell line. Human THP-1 monocytes were incubated for 4 hours with either medium (untreated control), LPS, or each dilution of ESM hydrolyzate. Whole cell lysates were then analyzed for NF-κB activity after normalizing for protein content. Bars represent mean ± standard error of the mean; values are presented relative to untreated control.
Note: *P<0.05 versus untreated control.
Abbreviations: NF-κB, nuclear factor kappa-light-chain-enhancer of activated B-cells; ESM, egg shell membrane; LPS, lipopolysaccharide; ESM-FH, ESM fermentation hydrolyzate; ESM-EH, ESM enzymatic hydrolyzate; ESM-CH, ESM chemical hydrolyzate.
Evaluation of the ESM source of NF-κB activation
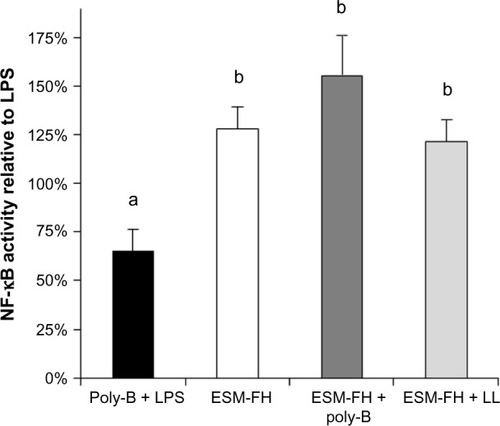
No increased NF-κB activity was observed in the cells treated with fermentation broth alone; however, ESM-FH activity was similar to what was observed previously (data not shown). Similarly, no increased NF-κB activity was observed with enzyme digest alone; however, ESM-EH activity was similar to what was observed previously (data not shown). Polymyxin B (10 mg/mL) adequately inhibited activation by LPS at 10 ng/mL () but showed a small degree of NF-κB activation itself (although not statistically significant). Lipoprotein lipase had no significant impact on NF-κB activity when used alone. These effects were confirmed in the separate bioassays of all three hydrolyzates (data not shown). There was no statistically significant difference between the ESM hydrolyzate alone samples and the ESM hydrolyzate + polymyxin B samples for any of the hydrolyzates tested (1:10,000 dilution) (ESM-FH, P=0.262; ESM-EH, P=0.727; ESM-CH, P=0.240). Similarly, there was no statistically significant difference between the ESM hydrolyzate alone samples and the ESM hydrolyzate + lipoprotein lipase samples for any of the hydrolyzates tested (1:10,000 dilution) (ESM-FH, P=0.539; ESM-EH, P=0.855; ESM-CH, P=0.281). Data for ESM-FH are presented in . Similar results were observed for ESM-EH and ESM-CH (data not shown).
Figure 3 Effects of poly-B and LL on in vitro NF-κB activity in untreated and LPS-treated THP-1 monocytes. Human THP-1 monocytes were incubated for 4 hours with either medium (untreated control), LPS, poly-B, poly-B + LPS, or LL. Whole cell lysates were then analyzed for NF-κB activity after normalizing for protein content. Bars represent mean ± standard error of the mean; values are presented relative to LPS.
Note: Columns with differing letters are statistically (P<0.05) different.
Abbreviations: poly-B, polymyxin B; LL, lipoprotein lipase; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B-cells; LPS, lipopolysaccharide.
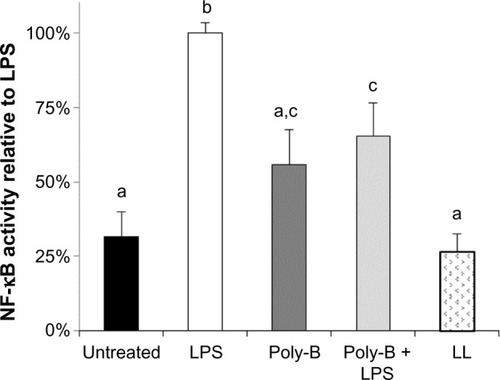
Figure 4 NF-κB activity of a fermentation hydrolyzate of ESM in THP-1 monocytes with and without poly-B or LL. Human THP-1 monocytes were incubated for 4 hours with either poly-B + LPS, ESM-FH alone (1:10,000 dilution), ESM-FH + poly-B, or ESM-FH + LL. Whole cell lysates were then analyzed for NF-κB activity after normalizing for protein content. Bars represent mean ± standard error of the mean; values are presented relative to LPS.
Note: Columns with differing letters are statistically (P<0.05) different.
Abbreviations: NF-κB, nuclear factor kappa-light-chain-enhancer of activated B-cells; ESM, egg shell membrane; poly-B, polymyxin B; LL, lipoprotein lipase; LPS, lipopolysaccharide; ESM-FH, ESM fermentation hydrolyzate.
Discussion
NF-κB is of fundamental importance to the functioning of innate immunity. Its dysregulation is implicated in the pathology of a considerable variety of human diseases. The goal of our studies was to investigate the potential role NF-κB might play in the beneficial biological activities reported previously for ESM. ESM hydrolyzates were chosen for our studies because of 1) the simplicity of their addition to cell cultures, 2) the variety of preparation methods available (eg, fermentation, enzymatic, or chemical), and 3) the potential to generate bioactive substances possessing differing activity profiles. Human PBMCs obtained from healthy volunteers were initially chosen for these studies owing to the wide variety of mononuclear cells present in this source (eg, T-lymphocytes, B-lymphocytes, natural killer cells, monocytes, and dendritic cells).Citation42,Citation43 Readily available THP-1 monocytes were subsequently used because of their antigen presenting characteristics that are similar to those of dendritic cells found in the gut, which are not readily available.
At the outset, it was unknown whether ESM hydrolyzates would stimulate or inhibit active levels of NF-κB. Previous workCitation4 in which an extract of ESM was shown to reduce the expression of proinflammatory cytokines in mitogen-challenged human PBMCs provided preliminary data that the hydrolyzates’ effect on NF-κB would be inhibitory. The data reported here show that ESM hydrolyzates significantly activate NF-κB in unstimulated cells. The ESM-FH and the ESM-EH showed quite similar activities in both PBMCs and THP-1 cell cultures. The ESM-CH, however, was about tenfold less active than the other hydrolyzates in THP-1 monocytes. As seen in , the 1:1,000 dilution of ESM-CH had approximately the same activity as the 1:10,000 dilutions of ESM-FH and ESM-EH. This result is logically consistent with the differing specificities of the hydrolysis conditions. That is, both microorganisms and purified enzymes will be fairly specific in their sites of hydrolysis within the ESM protein structure, whereas chemical hydrolysis is quite indiscriminate. It is likely that the NF-κB-activating soluble peptides that are formed initially during hydrolysis are being further degraded by the harsh chemical conditions, resulting in lower activity in cell culture.
Because ESM hydrolyzates induced increased levels of NF-κB, we felt it necessary to broadly explore the possible sources of this activity. Our immediate concern was to determine whether the activity was inherent in ESM or if it was due to materials used to produce the hydrolyzates (ie, fermentation inoculum or enzyme) or to some other type of contaminant. As neither the fermentation inoculum nor the enzyme produced elevated NF-κB activity, it was necessary to consider alternative sources for the observed activity. Degradation products of extracellular matrix components, such as fibronectin, collagen, proteoglycans, and hyaluronan, are believed to play a significant role in the pathogenesis of both osteo- and rheumatoid arthritis, likely through autoimmune mechanisms,Citation44–Citation46 and at least some of these components do so via activation of NF-κB.Citation47,Citation48 Hyaluronan,Citation3 proteoglycans,Citation2 and collagensCitation1 all naturally occur in ESM and may be responsible for at least part of the activity reported here. However, they occur at relatively low levels, up to 2% each hyaluronan and proteoglycans and up to 5% collagen. Therefore, we speculate that other sources may provide a more likely explanation for the observed stimulatory activity.
Bacterial cellular components and DNA have been found in the joints of patients suffering from a variety of arthritides,Citation49 and the innate immune response to these contaminants is believed to play a possible role in the pathogenesis of these diseases.Citation50,Citation51 Additionally, Pugh et alCitation52 found that the majority of in vitro macrophage activation exhibited by extracts of a number of common “immune enhancing” botanicals (Echinacea, ginseng, and alfalfa) was not inherent in the botanical compounds themselves but was due to contamination with bacterial cellular components such as bacterial lipoproteins and LPS. Being a natural product, it was logical that ESM would be contaminated with various microbial species. Although the ESM was heat treated to substantially reduce the living bioburden prior to hydrolysis, the cellular components from the formerly living microorganisms would remain.
Intact microorganisms should have been removed during filtration of the hydrolyzates through 0.2 micron filter membranes prior to the initial assays; however, cellular fragments from these microorganisms could not be ruled out as the source of activation. Therefore, we investigated the possible contribution of LPS and/or bacterial lipoprotein to the observed activation of NF-κB by ESM hydrolyzates. LPS activity is abrogated by the long-known antibiotic polymyxin B,Citation53 and the enzyme lipoprotein lipase deactivates bacterial lipoprotein through cleavage of the lipid–protein bond.Citation52 All three hydrolyzates (ESM-FH, ESM-EH, and ESM-CH) were evaluated alone and in combination with either polymyxin B or lipoprotein lipase. If a significant proportion of the NF-κB activity of the hydrolyzates was derived from either of these bacterial cellular components, the polymyxin B-treated and lipoprotein lipase-treated cells would have exhibited substantially decreased activity compared with the hydrolyzates alone. However, no differences were observed between the ESM hydrolyzates alone and either the ESM hydrolyzates + polymyxin B or ESM hydrolyzates + lipoprotein lipase. With no evidence that the NF-κB activation by ESM hydrolyzates is derived from bacterial cellular components, it would seem that this activity is attributable to the inherent bioactive components present in ESM. Further work will be needed to elucidate whether this activity draws from known stimulatory components of ESM (hyaluronan, proteoglycans, collagens) or from an as yet unidentified peptide or peptides produced from the various hydrolysis conditions.
Since the vast majority of NF-κB dysregulation encountered in disease pathology is that of elevated or inappropriately activated NF-κB,Citation24–Citation27 it would seem counterintuitive that ESM hydrolyzates activating NF-κB would contribute to ESM’s observed in vivo efficacy. Here we raise the possibility that this efficacy is achieved through an “oral tolerance” mechanism. Oral tolerance refers to the phenomenon of a reduced peripheral immune response (tolerance) that results from the repeated exposure of the mucosal immune system in the gut to ingested protein antigens. Oral tolerance to immunogenic peptides that are repeatedly ingested is believed to result from immune surveillance within the gut-associated lymphoid tissue as a way for the body to prevent an inappropriate or unnecessary immune response to proteins normally consumed in the diet. This tolerogenic behavior is thought to occur through the presentation of these peptides by antigen-presenting cells in the gut-associated lymphoid tissue to circulating T-cells, causing them to transform into regulatory T-cells (Treg cells), which results in anergy and even active suppression by these cells when they return to circulation.Citation54,Citation55 Oral tolerance has been shown to be effective in a variety of autoimmune diseases, including arthritis, diabetes, colitis, and multiple sclerosis (see Citation56 and Citation57 for a review). There has been particular attention paid to autoimmune arthritides in this area of research in which cartilage autoantigens (particularly type II collagen) produce antigen-specific suppression of the immune response. In addition, “bystander suppression”, wherein Treg cells produce a higher proportion of general anti-inflammatory cytokines (eg, IL-4, IL-10, and transforming growth factor-beta) suppressing the local autoimmune reaction independently of the antigen type, is also an active area of arthritis research.Citation58,Citation59 Further work will be needed to investigate whether ESM might function via 1) antigen-specific immune suppression from its extracellular matrix content (ie, collagen, proteoglycans, and hyaluronan), 2) bystander suppression, 3) some unrelated mechanism, or 4) some combination of these.
Conclusion
A great deal of research has been conducted relating to NF-κB since its discovery, particularly in the last couple of decades, and with so many people suffering from diseases involving its dysregulation, this trend is sure to increase. We presented here results that ESM hydrolyzates produced via three different hydrolysis conditions (ie, fermentation, enzymatic, and chemical) activate NF-κB in both human primary cells and a human leukemic cell line. The source of this activity was also investigated to confirm that it is inherent to ESM and not derived from hydrolysis conditions or from bacterial contamination. We also proposed a plausible hypothesis, via an oral tolerance mechanism, as to how this NF-κB activity might translate into the in vivo efficacy that has been previously observed with ESM. Further work is needed to confirm this proposed mechanism of action and to possibly identify the component(s) of ESM that is/are the source of the reported activity.
Acknowledgments
The sponsor of the studies was ESM Technologies, LLC.
Disclosure
KJR and FDL are employed by the sponsor of the studies. PLD and AO have no competing interests.
References
- WongMHendrixMJCvon der MarkKLittleCSternRCollagen in the egg shell membranes of the henDev Biol1984104128366203793
- BakerJRBalchDAA study of the organic material of hen’s-egg shellBiochem J19628235236113864145
- LongFDAdamsRGDeVoreDPinventorsPreparation of hyaluronic acid from eggshell membrane. United States patent US 69465519202005
- BensonKFRuffKJJensenGSEffects of natural eggshell membrane (NEM) on cytokine production in cultures of peripheral blood mononuclear cells: increased suppression of tumor necrosis factor-α levels after in vitro digestionJ Med Food201215436036822168811
- RuffKJDeVoreDPReduction of pro-inflammatory cytokines in rats following 7-day oral supplementation with a proprietary eggshell membrane-derived productMod Res Inflamm2014311925
- RuffKJDeVoreDPLeuMDRobinsonMAEggshell membrane: a possible new natural therapeutic for joint and connective tissue disorders. Results from two open-label human clinical studiesClin Interv Aging2009423524019554094
- RuffKJWinklerAJacksonRWDeVoreDPRitzBWEggshell membrane in the treatment of pain and stiffness from osteoarthritis of the knee: a randomized, multicenter, double-blind, placebo-controlled clinical studyClin Rheumatol20092890791419340512
- DaneschUSeyboldMRittinghausenRTreibelWBitterlichNNEM® brand eggshell membrane effective in the treatment of pain associated with knee and hip osteoarthritis: results from a six-center, open-label German clinical studyJ Arthritis201433136
- BauerKLDierenfeldESHartupBKEvaluation of a nutraceutical joint supplement in cranesProc North Am Crane Workshop2014122732
- DierenfeldESBaumDHampeLJensenJAtwellCWedekindKEvaluation of a nutraceutical joint supplement in camelsAmer Hol Vet Med Assoc J2014395966
- JiaHSaitoKAwWTranscriptional profiling in rats and an ex vivo analysis implicate novel beneficial function of egg shell membrane in liver fibrosisJ Funct Foods2013516111619
- SenRBaltimoreDMultiple nuclear factors interact with the immunoglobulin enhancer sequencesCell1986467057163091258
- GilmoreTDIntroduction to NF-κB: players, pathways, perspectivesOncogene2006256680668417072321
- AkiraSUematsuSTakeuchiOPathogen recognition and innate immunityCell200612478380116497588
- HaydenMWestAGhoshSNF-kB and the immune responseOncogene2006256758678017072327
- BegAFincoTNantermetPBaldwinATumor necrosis factor and interleukin-1 lead to phosphorylation and loss of IKBa: a mechanism for NF-κB activationMol Cell Biol1993136330133108497253
- FlodstriimMWelshNEizirikDCytokines activate the nuclear factor kB (NF-κB) and induce nitric oxide production in human pancreatic isletsFEBS Letters1996385468641463
- SchreckRAlbermannKBaeuerlePNuclear factor kB: an oxidative stress-responsive transcription factor of eukaryotic cells (a review)Free Rad Res Comms1992174221237
- ScandaliosJOxidative stress: molecular perception and transduction of signals triggering antioxidant gene defensesBraz J Med Biol Res200538995101416007271
- CooperSBowdenGUltraviolet B regulation of transcription factor families: roles of nuclear factor-kappa B (NF-κB) and activator protein-1 (AP-1) in UVB-induced skin carcinogenesisCurr Cancer Drug Targets2007732533417979627
- ReelfsOTyrrellRPourzandCUltraviolet A radiation-induced immediate iron release is a key modulator of the activation of NF-κB in human skin fibroblastsJ Invest Dermatol20041221440144715175035
- KonoHRusynIYinMNADPH oxidase–derived free radicals are key oxidants in alcohol-induced liver diseaseJ Clin Invest200010686787211018074
- SheHXiongSLinMZandiEGiuliviCTsukamotoHIron activates NF-κB in Kupffer cellsAm J Physiol Gastrointest Liver Physiol2002283G719G72612181188
- YamamotoYGaynorRRole of the NF-κB pathway in the pathogenesis of human disease statesCurr Mol Med2001128729611899077
- PrajapatiBSinghalMYashwantSharmaGGuptaVRole of NFκB in various immunological and inflammatory disordersInt J Toxicol Pharmacol Res2010213539
- CourtoisGGilmoreTMutations in the NF-κB signaling pathway: implications for human diseaseOncogene2006256831684317072331
- TakPFiresteinGNF-κB: a key role in inflammatory diseasesJ Clin Invest2001107171111134171
- BrandKPageSRoglerGactivated transcription factor nuclear factor-kappa B is present in the atherosclerotic lesionJ Clin Invest1996977171517228601637
- HajraLEvansAChenMHydukSCollinsTCybulskyMThe NF-kappa B signal transduction pathway in aortic endothelial cells is primed for activation in regions predisposed to atherosclerotic lesion formationProc Natl Acad Sci U S A200097169052905710922059
- AkamaKAlbaneseCPestellRVan EldikLAmyloid β-peptide stimulates nitric oxide production in astrocytes through an NFκB-dependent mechanismProc Natl Acad Sci U S A19989510579558009576964
- AkamaKVan EldikLβ-amyloid stimulation of inducible nitric-oxide synthase in astrocytes is interleukin-1β- and tumor necrosis factor-α (TNFα)-dependent, and involves a TNFα receptor-associated factor- and NFκB-inducing kinase-dependent signaling mechanismJ Biol Chem2000275117918792410713108
- MiterskiBBohringerSKleinWInhibitors in the NFkappaB cascade comprise prime candidate genes predisposing to multiple sclerosis, especially in selected combinationsGenes Immun2002321121912058256
- YanJGreerJNF-kappa B, a potential therapeutic target for the treatment of multiple sclerosisCNS Neurolog Disord Drug Targets200876536557
- PatelSSantaniDRole of NF-κB in the pathogenesis of diabetes and its associated complicationsPharmacol Rep20096159560319815941
- AndreasenAKellyMBergRMøllerKPedersenBType 2 diabetes is associated with altered NF-κB DNA binding activity, JNK phosphorylation, and AMPK phosphorylation in skeletal muscle after LPSPLoS One201169e2399921931634
- BasseresDBaldwinANuclear factor-κB and inhibitor of κB kinase pathways in oncogenic initiation and progressionOncogene2006256817383017072330
- KordesUKrappmannDHeissmeyerVLudwigWScheidereitCTranscription factor NF-kappaB is constitutively activated in acute lymphoblastic leukemia cellsLeukemia20001439940210720133
- BauerleKTSchweppeREHaugenBRInhibition of nuclear factor-kappa B differentially affects thyroid cancer cell growth, apoptosis, and invasionMol Cancer2010911720492683
- StrohbehnREEtzelLRFigginsJinventorsSolubilized protein composition obtained from avian eggshell membrane. United States patent US 8173174582012
- BowieAGO’NeillLAJVitamin C inhibits NF-κB activation by TNF via the activation of p38 mitogen-activated protein kinaseJ Immunol2000165127180718811120850
- Systat Software, Inc Available at: http://www.systat.comAccessed January 6, 2015
- ReichertTDeBruyereMDeneysVLymphocyte subset reference ranges in adult caucasiansClin Immunol Immunopathol1991601902081712687
- Jentsch-UllrichKKoenigsmannMMohrenMFrankeALymphocyte subsets’ reference ranges in an age- and gender-balanced population of 100 healthy adults: a monocentric German studyClin Immunol2005116219219715993366
- HomandbergGADavisGManigliaCShrikhandeACartilage chondrolysis by fibronectin fragments causes cleavage of aggrecan at the same site as found in osteoarthritic cartilageOsteoarthritis Cartilage1997564504539536293
- YoshidaMTsujiMKurosakaDAutoimmunity to citrullinated type II collagen in rheumatoid arthritisMod Rheumatol200616527628117039307
- GyörgyBTóthfalusiLNagyGNatural autoantibodies reactive with glycosaminoglycans in rheumatoid arthritisArthritis Res Ther200810R11018789149
- NoblePWMcKeeCMCowmanMShinHSHyaluronan fragments activate an NF-kappa B/I-kappa B alpha autoregulatory loop in murine macrophagesJ Exp Med19961835237323788642348
- SaklatvalaJInflammatory signaling in cartilage: MAPK and NF-κB pathways in chondrocytes and the use of inhibitors for research into pathogenesis and therapy of osteoarthritisCurr Drug Targets2007830531317305508
- Van Der HeijdenIMWilbrinkBTchetverikovIPresence of bacterial DNA and bacterial peptidoglycans in joints of patients with rheumatoid arthritis and other arthritidesArthritis Rheum200043359359810728753
- ScanzelloCRPlaasACrowMKInnate immune system activation in osteoarthritis: is osteoarthritis a chronic wound?Curr Opin Rheumatol200820556557218698179
- KlinmanDDoes activation of the innate immune system contribute to the development of rheumatoid arthritis?Arthritis Rheum200348359059312632408
- PughNDTamtaHBalachandranPThe majority of in vitro macrophage activation exhibited by extracts of some immune enhancing botanicals is due to bacterial lipoproteins and lipopolysaccharidesInt Immunopharmacol2008871023103218486914
- MorrisonDCJacobsDMInhibition of lipopolysaccharide-initiated activation of serum complement by polymyxin BInfect Immun1976131298301175019
- FriedmanAWeinerHLInduction of anergy or active suppression following oral tolerance is determined by antigen dosageProc Natl Acad Sci U S A19949114668866928022835
- FariaAMCWeinerHLOral toleranceImmunol Rev2005206123225916048553
- GarsidePMowatAMKhorutsAOral tolerance in diseaseGut19994411371429862840
- FariaAMCWeinerHLOral tolerance: therapeutic implications for autoimmune diseasesClin Dev Immunol2006132–414315717162357
- StancikovaMStancikRGubzovaZRovenskyJCollagen in the treatment of rheumatic diseases: oral toleranceBratisl Med J199910010567571
- BroereFWietenLKoerkampEIKOral or nasal antigen induces regulatory T cells that suppress arthritis and proliferation of arthritogenic T cells in joint draining lymph nodesJ Immunol2008181289990618606641