
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Background
Indoleamine 2,3-dioxygenase (IDO) is an enzyme associated with the regulation of immune responses. Cytokines such as IFNγ induce its expression in endothelial cells originating from immune-privileged sites. In this study, we investigate regulators of IDO in primary endothelial cells from a non-immune-privileged site and determine whether IDO expression affects immune cell behavior.
Methods
IDO expression was determined using real-time quantitative polymerase chain reaction and immunoblotting. IDO activity was estimated using an IDO enzyme assay. Primary cells were transfected using microporation, and T-cell migration was determined using a cell transmigration assay.
Results
IDO is expressed in human saphenous vein endothelial cells after stimulation with IFNγ but not after treatment with TNFα, IL-1β, IL-2, IL-4, IL-6, or IL-10. VEGFβ and heparin negatively regulate IFNγ-driven increases in IDO. Overexpression of IDO in endothelial cells does not affect transmigration of T-cells.
Conclusion
IDO is expressed in human saphenous vein endothelial cells after stimulation with IFNγ. Heparin and angiogenesis stimulators such as VEGFβ negatively regulate its expression.
Keywords:
Introduction
The endothelial cell plays a central role in the control of inflammatory processes and can be involved in graft rejection, atherosclerosis, and vasculitides.Citation1–Citation3 In addition to the effect of endothelial cells in the recruitment of leukocytes into inflammation sites,Citation4,Citation5 they can also present antigens to white blood cells influencing their activation and transmigration.Citation6–Citation9
Indoleamine 2,3-dioxygenase (IDO) is an intracellular heme-containing enzyme that catalyzes the oxidative cleavage of the indole ring of L-tryptophan to produce kynurenine metabolites.Citation10 Depletion of L-tryptophan from the local microenvironment together with the production of kynurenine can modulate inflammation responses.Citation11,Citation12 IDO activation has been associated with antimicrobial effects and the modulation of T-cell responses,Citation13 at least partly by enhancement of proliferation of regulatory T-cells.Citation14,Citation15 Increased expression of IDO is related to a favorable environment for tumor growth and graft survival,Citation16–Citation19 the pathophysiology of depression,Citation20,Citation21 and protection against pulmonary hypertension.Citation22 IDO activity may be affected not only at the transcriptional and translational level, but also by posttranslational modification and SOCS3-associated differential degradation.Citation23–Citation25
What is currently not known is whether immune-regulatory molecules such as IDO mediate graft rejection exhibited by endothelial cells. When human umbilical vein endothelial cells (HUVECs) were stimulated with IFNγ, IDO expression and activity were increased. In addition, it was shown that IDO activity inhibition enhanced the ability of HUVECs to stimulate allogeneic T-cell responses.Citation26 Treatment of cells originating from non-immune-privileged sites such as human saphenous vein endothelial cells (HSVECs) with IFNγ did not upregulate IDO.Citation26 In a non-vascular in vitro model of human corneal endothelial cells (HCECs) IFNγ-driven upregulation of IDO had no effect on T cell proliferation.Citation27 In this study, we used primary cells isolated from a non-immune-privileged source, the saphenous vein, to investigate factors that affect IDO regulation and determined whether forced expression of IDO in these cells affected immune cell migration.
Materials and methods
Isolation of primary endothelial cells from human saphenous veins
Primary endothelial cells from human saphenous veins were isolated using the method described elsewhere.Citation28 Briefly, both ends of the vein were sutured and the lumen was filled with 10 mL prewarmed 1 mg/mL collagenase II solution (Sigma-Aldrich Co, St Louis, MO, USA) and incubated at 37°C for 10 minutes. Then collagenase II solution was collected and vein was flushed with 10 mL trypsin–ethylenediaminetetraacetic acid and the effluent was collected again. Cells were pelleted and plated in a T25 flask. Endothelial cells were further purified with anti-CD105 microbeads using MS columns (Miltenyi Biotech, Auburn, CA, USA). Cell fractions showing high levels of staining for von Willebrand factor were used.
Cell culture and propagation
Cells were maintained in a subconfluent monolayer at 37°C in a humidified atmosphere containing 5% CO2. Endothelial medium was prepared by combining equal volumes of endothelial SFM (Thermo Fisher Scientific, Waltham, MA, USA) and endothelial EGM2 medium (Walkersville, MD, USA) and adding per 500 mL medium 5 mL of penicillin/streptomycin, 5 mL glutamine, and 5 mL of 5 mg/mL endothelial cell growth supplement purchased from BD Biosciences (San Jose, CA, USA). They were subcultured using trypsin, and screening for mycoplasma contamination was carried out on a regular basis. For experimental use, cells were treated with recombinant human IFNγ, recombinant human TNFα, recombinant human IL-2, recombinant human IL-4, recombinant human IL-6, recombinant human IL-10, and recombinant human VEGFβ, all purchased from Peprotech (Rocky Hill, NJ, USA).
IDO activity assay
IDO activity was determined in wells exhibiting cell confluency greater than 70% using a colorimetric assay based on detection of kynurenine as described previously.Citation29,Citation30 Briefly, L-kynurenine standards (0–200 μM) were prepared in endothelial medium. Kynurenine in control, test, and standard samples was precipitated by incubation with 30% trichloroacetic acid at 50°C for 30 minutes and centrifuged at 2,000× g for 10 minutes. Visualization of signal took place by adding equal volumes of Ehrlich reagent to the supernatant and incubation at 65°C for 15 minutes. Ehrlich reagent was prepared using 6.2 mL 1-propanol, 1.5 mL distilled water, 2.6 mL of 70% perchloric acid, and 1.5 g of 4-dimethylbenzinamide, all purchased from Sigma-Aldrich Co. Absorbance was measured at 492 nm in a colorimetric plate reader.
Cell homogenization and Western blotting
Control and treated cells were harvested from flasks, washed in phosphate-buffered saline, and lysed in SDS sample buffer (62.5 mM Tris-HCl, pH 6.8, 2% w/v SDS, 10% glycerol, 50 mM DTT, and 0.01% bromophenol blue) for 5 minutes on ice and sonicated for 10–15 seconds. Samples were then heated for 5 minutes at 90°C, micro-centrifuged briefly, and stored at −20°C. Antibodies against IDO and SOCS3 were purchased from Abcam (Cambridge, UK). Primary antibody of interest diluted in blocking buffer (1.5% milk in Tris-buffered saline/Tween 20, pH 7.6) containing 5% BSA was added to the membranes and left incubating overnight at 4°C. Membranes were then washed and incubated with secondary antibody. Signal was visualized on X-ray films using ECL Western blotting reagents (Amersham Biosciences, Little Chalfont, UK).
Real-time polymerase chain reaction
Cells were washed with phosphate-buffered saline, and mRNA was obtained using TRIzol. cDNA was obtained using the SuperScript First-Strand Synthesis System for RT-PCR. Detection of product was made using SYBR Green (Thermo Fisher Scientific). Relative quantification of mRNA transcription was performed by normalizing the Ct value of IDO to the Ct value of actin in control:
Then the ΔΔCt product was calculated by subtracting the dCt value of controls from the dCt value of treated cells:
Using the formula 2−ΔΔCt, the relative enhancement ratio of IDO transcription was estimated. The following primer sequences were used: HPRT forward: 5′-GCAGACTTTGCTTTCCTTGGTC-3′; HPRT reverse: 5′-CTGGCTTATATCCAACACTTCGTG-3′; IDO forward: 5′-GGTCATGGAGATGTCCGTTAA-3′; IDO reverse: 5′-ACCAATAGAGAGACCAGGAAGAA-3′.
Transfection of endothelial cells using microporation
Microporation apparatus, tips, buffers and the MP-1096 kit were purchased from Labtech International (Ringmer, UK). The desired number of cells were resuspended in 24 μL of microporation suspension buffer and kept at 4°C for the duration of the experiment. One microgram DNA was added and cells were transfected using microporation. The vectors used were pcDNA3.1 (empty vector), pSMART2G (empty vector), pcDNA3.1-EGFP, and pSMART2G-IDO. Voltage was set at 1,350 mV, pulse number at 1, and duration at 30 ms, and p10 tips were used. After microporation, cell aliquots were put in endothelial medium in 24-well plates at 37°C and left to incubate overnight. For experiments, cells were used in the first 3 days after transfection.
Transmigration assay
HSVECs were plated on a 24-well Transwell chamber at full confluence and left to incubate for 1 day. PBMCs were obtained from human donors or buffy coat using density centrifugation with Ficoll-Paque. T-cells were obtained from PBMCs using MACS bead separation, employing negative selection with biotin-conjugated antibodies against CD14, CD16, CD19, CD36, CD56, and CD235. Fractions were stained with CD3 fluorescein isothiocyanate to determine purity and those staining positive were used for experiments. T-cell fractions containing 3×105 cells were applied in 1:10 dilution on the top of the endothelial cells in the Transwell chambers and co-cultured for a further day. T-cells transmigrating through the endothelial monolayer were collected on the bottom chamber and their number determined using a hemocytometer.
Statistical analysis of results
Results are presented as means + standard error of the mean. Statistical significance was carried out using a t-test assuming one-tailed distribution using two-sample unequal variance. P<0.05 was considered as a statistically significant value. Microsoft Excel data processing functions were used for data processing and analysis.
Results
IFNγ is the main regulator of IDO expression in primary HSVECs
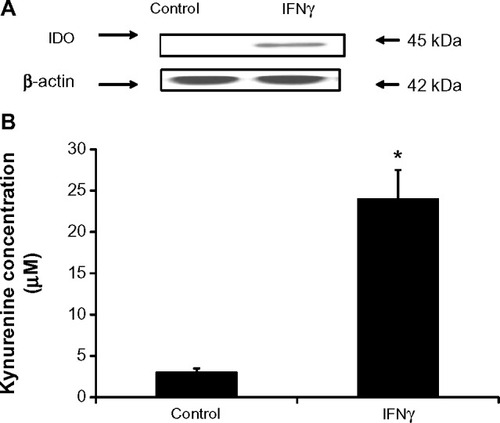
Differential regulation of IDO in endothelial cells stimulated with IFNγ was evident in other studies depending on the endothelial cell origin.Citation26,Citation27 To demonstrate whether IDO expression in primary HSVECs was increased after IFNγ stimulation, these primary cells were treated with concentrations of IFNγ (0–120 ng/mL) for 2 days. IDO expression was assessed using immunoblotting, and IDO activity was assessed using an IDO activity assay. Results show that treatment of HSVECs with IFNγ (80 ng/mL) resulted in increased IDO expression and IDO activity ( and ).
Figure 1 IFNγ upregulates IDO in primary human saphenous vein endothelial cells (HSVECs).
Notes: cells were treated with IFNγ (80 ng/mL) for 2 days. IDO expression levels were assessed using immunoblotting, and IDO activity was assessed using the L-kynurenine IDO activity assay. Results show that IDO protein levels (A) and IDO activity (B) are increased after HSVECs are stimulated with IFNγ for 2 days. Immunoblotting data (A) are presented as a representative blot of three experiments and kynurenine activity assay (B) is presented as means + standard error of the mean (n=4). *P<0.05.
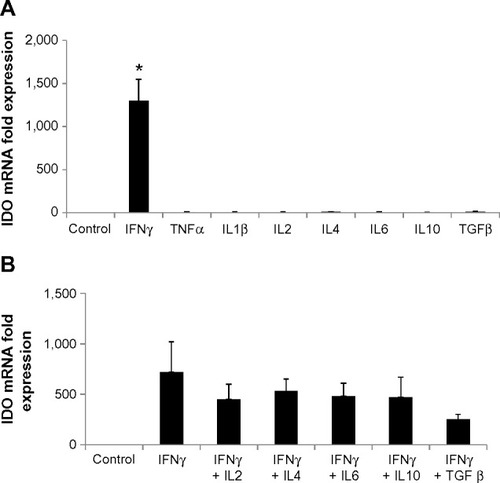
To investigate what other regulators drive expression of IDO, HSVECs were treated with IFNγ alone or in combination with cytokines in relevant concentrations such as TNFα (80 ng/mL), IL-1β (20 ng/mL), IL-2 (20 ng/mL), IL-4 (40 ng/mL), IL-6 (40 ng/mL), IL-10 (40 ng/mL), and TGFβ (100 ng/mL) for 2 days. IDO mRNA expression was assessed using real-time quantitative polymerase chain reaction. Results show that IDO mRNA was increased after treatment with IFNγ but not after treatment with the other cytokines (). In addition, co-treatment of HSVECs with IFNγ and these cytokines did not result in significant change in IFNγ-driven increases in IDO expression and activity, suggesting that IFNγ was the main regulator of IDO in HSVECs ( and ).
Figure 2 IFNγ is the main regulator of IDO in human saphenous vein endothelial cells (HSVECs).
Notes: Cells were treated with IFNγ (80 ng/mL), TNFα (80 ng/mL), ILIβ (20 ng/mL), IL2 (20 ng/mL), IL4 (40 ng/mL), IL6 (40 ng/mL), IL10 (40 ng/mL), and TGFβ (100 ng/mL) either alone or in combination for 2 days. IDO expression was determined using real-time quantitative polymerase chain reaction. Results show that only IFNγ significantly increases expression of IDO in HSVECs (A). Co-treatment of cells with IFNγ and the other cytokines did not result in changes in IFNγ-driven increases in IDO expression (B). Results are presented as the means of three experiments + standard error of the mean. *P<0.05.
VEGF, heparin, and nitric oxide are negative regulators of IDO in HSVECs
IFNγ has exhibited antiangiogenic effects in human cornea by downregulating expression of VEGFα.Citation31 To identify whether crosstalk of IFNγ and VEGF pathways exists in HSVECs, these cells were stimulated with IFNγ (80 ng/mL) alone or in combination with VEGFβ (40 ng/mL) for 1 day and 2 days. Expression of IDO was determined using immunoblotting with a monoclonal antibody against human IDO. IDO was not expressed in unstimulated cells or cells treated with VEGFβ only (data not shown). When HSVECs were treated with IFNγ, then IDO protein levels were increased, whereas co-treatment of HSVECs with IFNγ and VEGFβ decreased IDO levels relative to stimulation with IFNγ only (). SOCS3 has been shown to target IDO for proteasomal degradation.Citation25 To investigate the possibility that SOCS3 protein levels are regulated in a similar manner to IDO’s protein levels, its expression was investigated using immunoblotting. SOCS3 protein levels remained unchanged in cells treated with IFNγ only. However, when cells were co-treated with IFNγ and VEGFβ, SOCS3 protein levels increased after 24 hours and completely eliminated after 48 hours ().
Figure 3 Negative regulators of IDO expression in primary human saphenous vein endothelial cells (HSVECs).
Notes: Cells were treated with IFNγ (80 ng/mL) either alone or in combination with VEGFβ (40 ng/mL), heparin (10 mg/mL and 20 mg/mL), or nitric oxide donor DetaNO (10 μM and 50 μM). Expression of IDO and SOCS3 was determined using immunoblotting. Results show that IDO expression is reduced in cells treated with IFNγ and VEGFβ relative to cells treated with IFNγ only. Protein levels of SOCS3 also decreased after 48 hours of treatment of HSVECs with IFNγ and VEGFβ relative to cells treated with IFNγ only (A). Co-treatment of HSVECs with IFNγ and heparin also resulted in a reduction of IFNγ-driven increase in IDO activity (B). Co-treatment of HSVECs with IFNγ and DetaNO resulted in reduction of IFNγ-driven increases in IDO activity (C) but not in IDO expression levels (D). Immunoblotting data are presented as representative blots that were repeated at least three times. IDO activity results are presented as means + standard error of the mean of three independent experiments. *P<0.05.
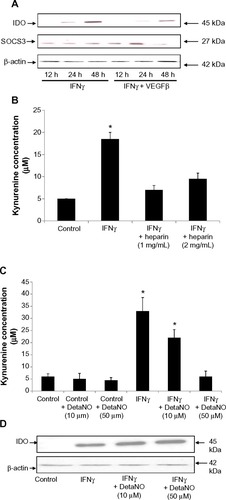
We have previously used heparin in the culture of endothelial cells and questioned whether it interferes with the induction of IDO.Citation26 For this purpose, we measured the IDO response to IFNγ in the presence and absence of heparin. HSVECs were stimulated with IFNγ (80 ng/mL) with or without cell culture-relevant concentrations of heparin (1 mg/mL and 2 mg/mL). IDO activity was assessed using an IDO activity assay. This concentration of heparin was chosen because it is detected in endothelial cell media. Results show that heparin abrogated IFNγ-driven increases in IDO activity ().
Differential regulation of IDO activity by nitric oxide (NO) has been shown in monocyte-derived macrophages but not in microglial cells.Citation32 To investigate whether NO has a regulatory effect on IDO activity in HSVECs, cells were stimulated with IFNγ (80 ng/mL) with or without the NO donor DetaNO (10 μM and 50 μM). IDO expression levels were assessed using immunoblotting and IDO activity was assessed using the IDO activity assay. Results show that NO decreased IFNγ-driven increases in IDO activity. However, IDO protein levels did not follow the same regulation pattern ().
Increased expression of IDO in HSVECs does not affect T-cell transmigration properties
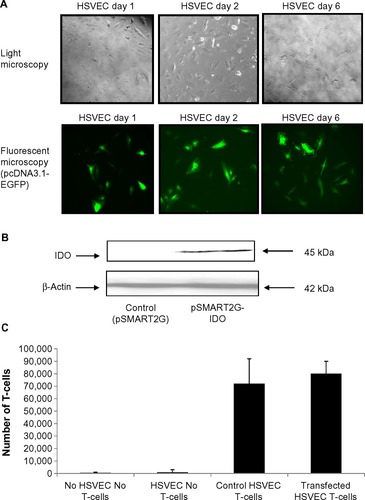
To determine the effect of IDO overexpressed in endothelial cells independently of IFNγ stimulation, primary HSVECs were transiently transfected with EGFP using microporation. EGFP fluorescence was detected up to 6 days using fluorescence microscopy (). Then, HSVECs were transiently transfected with an IDO plasmid. Overexpression of IDO was confirmed using immunoblotting (). IDO-overexpressing HSVECs were plated on Transwell chambers and left to incubate for 1 day. T-cells were then added in the Transwell chambers and coincubated with HSVECs for 24 hours further. The number of cells detected in the lower compartment of the Transwell chamber was counted using a hemocytometer. Results show that IDO overexpression in HSVECs did not affect the number of T-cells transmigrating through the endothelial cell monolayer barrier ().
Figure 4 IDO does not affect T-cell transmigration properties.
Notes: Human saphenous vein endothelial cells (HSVECs) were transfected with pcDNA3.1-EGFP and signal was detected up to 6 days after transfection. HSVECs were then transiently transfected with pSMART2G-IDO. Transfected cells were plated in Transwell chambers and left to incubate for 1 day. T-cells were then added and incubation was continued for a further day. T-cells transmigrated through the endothelial monolayer were then counted using a hemocytometer. Results show that IDO does not affect the number of T-cells transmigrating through the endothelial cell barrier. Results are presented as an average of one experiment that was repeated three times with similar results.
Discussion
IDO is an enzyme that metabolizes tryptophan and is known to be central in the regulation of immune responses. IDO can be expressed in endothelial cells and alters T-cell responses to the endothelium. The aim of this study was to identify regulators of IDO expression in HSVECs and to determine whether IDO expression in these primary cells pooled together from different donors affects inflammation-related processes.
HSVECs were treated with proinflammatory and anti-inflammatory cytokines. IDO transcription and protein levels and activity were then assessed. We were not able to detect IDO expression in endothelial cells cultured under normal conditions. Stimulation of HSVECs with different concentrations of TNFα, IL-1β, IL-2, IL-4, IL-6, and IL-10 either alone or in combinations had a negligible effect on IDO expression that did not result in a significant functional effect as determined by production of kynurenines (data not shown). However, stimulation of cells with IFNγ resulted in an increase in IDO expression and activity. The high concentrations of IFNγ used to achieve upregulation of IDO in HSVECs did not result in detrimental effects to the cells. Co-stimulation of IFNγ-treated endothelial cells with cytokines such as TNFα, IL-1β, IL-2, IL-4, IL-6, and IL-10 did not affect IDO expression and activity relative to cells treated with IFNγ.
Conflicting reports on the ability of HSVECs to increase IDO expression after treatment with IFNγ have been reported, and we have described that IFNγ is unable to upregulate IDO activity in HSVECs while confirming upregulation in HUVECs.Citation26 However, those previous experiments were performed with heparin as an additive to endothelial cell media to enhance cell proliferation and survival. To investigate whether heparin affects IFNγ-driven increases in IDO activity in HSVECs we replaced heparin with endothelial cell growth supplement factors, golden grade fetal calf serum and used specialized endothelial cell medium. By optimizing these cell culture conditions no significant loss of endothelial cell viability was observed. In addition HSVECs confluency was greater than 70% in experimental conditions. Using these experimental conditions we have shown that heparin, in concentrations routinely used in cell culture and experimental media, counteracts the ability of IFNγ to upregulate IDO in HSVECs. Heparin interferes with IFNγ activity by binding to the cytokine and reducing its ability to bind and activate the IFNγ receptor.Citation33,Citation34 Heparin counteracts IFNγ-induced increases of iNOS in rat endothelial cells,Citation35 apoptosis of primary trophoblasts,Citation36 and fractalkine expression in human endothelial cells.Citation37 These results suggest that endothelial cells located in non-immune-privileged tissues can exhibit induction of IDO activity after IFNγ stimulation in a similar manner to the endothelial cells associated with immune-privileged tissue.
VEGFα has been associated with IDO expression upregulation in prostaglandin E2- and TNFα-treated dendritic cells.Citation38 In addition, IFNγ has exhibited antiangiogenic effects in human cornea by downregulating expression of VEGFα.Citation31 In this study, we show for the first time that angiogenesis stimulators such as VEGFβ have a negative effect on IFNγ-driven increases in IDO expression and also regulate SOCS3 protein levels. Orabona et al and Pallota et al have shown that SOCS3 mediates proteasome degradation of IDO in dendritic cells.Citation25,Citation39 It is tempting to hypothesize that the same mechanism takes place in primary endothelial cells co-treated with VEGFβ and IFNγ. However, further research is needed to prove a direct link between VEGFβ and SOCS3-mediated proteasome degradation of IDO. NO has been shown to affect IDO activity in monocyte-derived macrophages at the posttranslational levels but not in other biological models such as microglial cells.Citation32 In our study, we confirm that NO has a negative effect on IDO activity at the posttranslational level. IDO catabolizes tryptophan by inserting oxygen to generate N-formyl-kynurenine. NO may inhibit this function by displacing bound tryptophan and oxygen after binding to IDO’s active heme site.Citation40
The recruitment of inflammatory leukocytes from blood vessels is controlled by the expression of adhesion molecules on the endothelium as well as the secretion and presentation of chemokines.Citation26 To determine whether IDO expression in HSVECs affected T-cell transmigration, IDO was overexpressed in these cells using microporation. This method resulted in transfection efficiency of approximately 50% that is stable for at least 6 days. Transfected HSVECs were allowed to attach onto Transwell chambers and form a monolayer. The number of T-cells transmigrating through the endothelial monolayer transfected with IDO versus the number of T-cells transmigrating through the endothelial cells expressing basal levels of IDO was then determined. Using this technique, we were not able to demonstrate a significant change in T-cell transmigration. It is possible that the number of non-transfected cells in the monolayer was high enough to allow passage of T-cells. However, controls using IFNγ-treated HSVECs did not result in a significant change in the number of transmigrated cells.
IDO regulates immune responses which otherwise could cause tissue damage. The endothelial cell is one of the first contacts of an activated lymphocyte, hence whether it has the mechanistic capability to increase IDO expression and regulate the immune response is highly significant. Max et al have demonstrated that IFNγ-induced IDO expression in Ewing sarcoma cells decreased T-cell proliferation in co-culture assays 3 days after treatment, but no effect on T-cell proliferation was evident 1 day after treatment.Citation41 In contrast, Lahdou et al have shown that upregulation of IDO in corneal endothelial cells does not affect T-cell proliferation.Citation27 In addition, they demonstrated that extraordinarily high levels of L-kynurenine close to 400 μM are necessary for a suppressive effect on T-cell proliferation to be detected. In our study, we achieved relatively low levels of L-kynurenine concentration up to 10 μM (transfected cells) (data not shown) and 40 μM (IFNγ-treated cells) when assessed in 24-well plates. However, the levels of IFNγ used in our study were higher than the physiological levels of IFNγ found in blood serum of patients treated for infections, although it cannot be excluded that higher levels of IFNγ accumulate in the target tissue’s microenvironment. So we believe that, with our chosen experimental procedures, we do not underestimate the IFNγ concentration and IDO expression the in vivo endothelial cells may be subjected to. In addition, it is important to distinguish between effects on cell proliferation and direct effects on transmigration properties of the cells. It is possible that the duration of lymphocyte exposure and the concentration of kynurenines the lymphocytes are subjected to, in the microenvironment of IDO-overexpressing cells, may dictate T-cell response. It is of interest to establish whether IDO may have additional roles in endothelium. Park et al have suggested that the role of IDO is different between target tissues and infiltrated immune cells while demonstrating that low endothelial IDO levels correlate with lower non-relapse mortality in patients with acute graft-versus-host disease after allogeneic hematopoietic stem cell transplantation.Citation42 Wang et al have shown in mouse models of systemic inflammation that IDO expressed in vascular endothelial cells results in the production of kynurenines which regulate vascular tone and blood pressure regulation.Citation43
The data presented in this study build on previous published observations and altogether provide an insight into the mechanism by which endothelial cells regulate immune responses and the role of endothelial IDO. Our results show that HSVECs have the capability to upregulate IDO after stimulation with IFNγ but, at relatively low kynurenine concentrations, do not directly affect T-cell transmigration properties. We also show for the first time that angiogenesis stimulators negatively regulate the IFNγ-driven increases in IDO expression. Given the interest that exists in IDO and graft survival, it is of interest to investigate whether angiogenesis regulators can promote or inhibit graft survival in vivo and the possibility that this might be an IDO-dependent effect.
Acknowledgments
This study was funded by the British Heart Foundation (BHF).
Supplementary materials
Figure S1 IDO dose–response curve.
Note: Results are presented as means + standard deviation of one experiment.
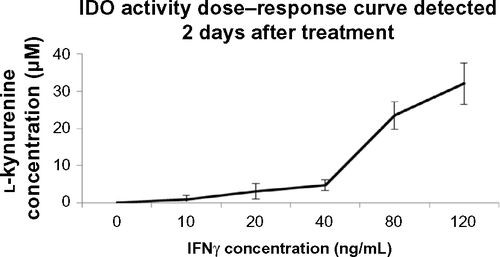
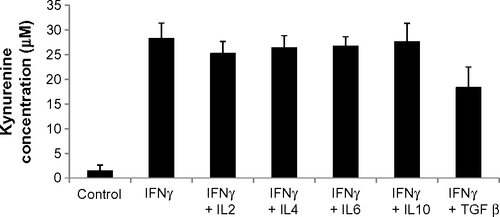
Figure S2 IDO activity after treatment of human saphenous vein endothelial cells with cytokines for 2 days.
Note: Results are presented as means + standard deviation of one experiment.
Disclosure
The authors report no conflicts of interest in this work.
References
- ManduteanuISimionescuMInflammation in atherosclerosis: a cause or a result of vascular disorders?J Cell Mol Med20121691978199022348535
- BiasucciLMCardilloMTBiomarkers of inflammation and endothelial function: the holy grail of experimental and clinical medicine?Vascul Pharmacol2012561–2262822198643
- SisBEndothelial molecules decipher the mechanisms and functional pathways in antibody-mediated rejectionHum Immunol201273121218122522863448
- DanielAEvan BuulJDEndothelial junction regulation: a prerequisite for leukocytes crossing the vessel wallJ Innate Immun20135432433523571667
- HeemskerkNvan RijsselJvan BuulJDRho-GTPase signaling in leukocyte extravasation: an endothelial point of viewCell Adh Migr201482677524621576
- CharreauBMolecular regulation of endothelial cell activation: novel mechanisms and emerging targetsCurr Opin Organ Transplant201116220721321415823
- MaiJVirtueAShenJWangHYangXFAn evolving new paradigm: endothelial cells – conditional innate immune cellsJ Hematol Oncol201366123965413
- MullerWAGetting leukocytes to the site of inflammationVet Pathol201350172223345459
- Marelli-BergFMFrascaLWengLLombardiGLechlerRIAntigen recognition influences transendothelial migration of CD4+ T cellsJ Immunol199916226967039916688
- FallarinoFGrohmannUPuccettiPIndoleamine 2,3-dioxygenase: from catalyst to signaling functionEur J Immunol20124281932193722865044
- MunnDHZhouMAttwoodJTPrevention of allogeneic fetal rejection by tryptophan catabolismScience19982815380119111939712583
- MellorALSivakumarJChandlerPPrevention of T cell-driven complement activation and inflammation by tryptophan catabolism during pregnancyNat Immunol200121646811135580
- MurakamiYHoshiMImamuraYAriokaYYamamotoYSaitoKRemarkable role of indoleamine 2,3-dioxygenase and tryptophan metabolites in infectious diseases: potential role in macrophage- mediated inflammatory diseasesMediators Inflamm2013201339198423476103
- SchmettererKGNeunkirchnerAPicklWFNaturally occurring regulatory T cells: markers, mechanisms, and manipulationFASEB J20122662253227622362896
- SantAJChavesFALeddonSATungJThe control of the specificity of CD4 T cell responses: thresholds, breakpoints, and ceilingsFront Immunol2013434024167504
- MunnDHBlocking IDO activity to enhance anti-tumor immunityFront Biosci (Elite Ed)2012473474522201909
- DürrSKindlerVImplication of indolamine 2,3 dioxygenase in the tolerance toward fetuses, tumors, and allograftsJ Leukoc Biol201393568168723325926
- PlattenMWickWVan den EyndeBJTryptophan catabolism in cancer: beyond IDO and tryptophan depletionCancer Res201272215435544023090118
- JohnsonTSMunnDHHost indoleamine 2,3-dioxygenase: contribution to systemic acquired tumor toleranceImmunol Invest2012416–776579723017145
- Catena-Dell’OssoMRotellaFDell’OssoAFagioliniAMarazzitiDInflammation, serotonin and major depressionCurr Drug Targets201314557157723531160
- MándiYVécseiLThe kynurenine system and immunoregulationJ Neural Transm2012119219720921744051
- XiaoYChristouHLiuLEndothelial indoleamine 2,3-dioxygenase protects against development of pulmonary hypertensionAm J Respir Crit Care Med2013188448249123822766
- FujigakiSSaitoKTakemuraML-tryptophan-L-kynurenine pathway metabolism accelerated by Toxoplasma gondii infection is abolished in gamma interferon-gene-deficient mice: cross-regulation between inducible nitric oxide synthase and indoleamine-2,3-dioxygenaseInfect Immun200270277978611796611
- OhGSPaeHOChoiBM3-Hydroxyanthranilic acid, one of metabolites of tryptophan via indoleamine 2,3-dioxygenase pathway, suppresses inducible nitric oxide synthase expression by enhancing heme oxygenase-1 expressionBiochem Biophys Res Commun200432041156116215249210
- OrabonaCPallottaMTVolpiCSOCS3 drives proteasomal degradation of indoleamine 2,3-dioxygenase (IDO) and antagonizes IDO-dependent tolerogenesisProc Natl Acad Sci U S A200810552208282083319088199
- BeutelspacherSCTanPHMcClureMOLarkinDFLechlerRIGeorgeAJExpression of indoleamine 2,3-dioxygenase (IDO) by endothelial cells: implications for the control of alloresponsesAm J Transplant2006661320133016686756
- LahdouIEnglerCMehrleSRole of human corneal endothelial cells in T-cell-mediated alloimmune attack in vitroInvest Ophthalmol Vis Sci20145531213122124370834
- MouratidisPXGeorgeAJTransfection of indoleamine 2,3 dioxygenase in primary endothelial cellsMethods Mol Biol201061614915920379874
- TakikawaOKuroiwaTYamazakiFKidoRMechanism of interferon-gamma action. Characterization of indoleamine 2,3-dioxygenase in cultured human cells induced by interferon-gamma and evaluation of the enzyme-mediated tryptophan degradation in its anticellular activityJ Biol Chem19882634204120483123485
- AlegreELópezASGonzálezATryptophan metabolites interfere with the Ehrlich reaction used for the measurement of kynurenineAnal Biochem2005339118818915766729
- KommineniVKNagineniCNWilliamADetrickBHooksJJIFN-gamma acts as anti-angiogenic cytokine in the human cornea by regulating the expression of VEGF-A and sVEGF-R1Biochem Biophys Res Commun2008374347948418639520
- Alberati-GianiDMalherbePRicciardi-CastagnoliPKöhlerCDenis-DoniniSCesuraAMDifferential regulation of indoleamine 2,3-dioxygenase expression by nitric oxide and inflammatory mediators in IFN-gamma-activated murine macrophages and microglial cellsJ Immunol199715914194269200481
- SarrazinSBonnafféDLubineauALortat-JacobHHeparan sulfate mimicry: a synthetic glycoconjugate that recognizes the heparin binding domain of interferon-gamma inhibits the cytokine activityJ Biol Chem200528045375583756416155294
- FluhrHSpratteJHeidrichSEhrhardtJSteinmüllerFZygmuntMHeparin inhibits interferon-γ signaling in human endometrial stromal cells by interference with the cellular binding of interferon-γFertil Steril20119541272127720542267
- BonmannEJüttlerEKrestelHESprangerMHeparin inhibits induction of nitric oxide synthase by cytokines in rat brain microvascular endothelial cellsNeurosci Lett1998253295989774158
- HillsFAAbrahamsVMGonzález-TimónBHeparin prevents programmed cell death in human trophoblastMol Hum Reprod200612423724316556679
- HatakeyamaMImaizumiTTamoWHeparin inhibits IFN-gamma-induced fractalkine/CX3CL1 expression in human endothelial cellsInflammation200428171315072225
- MartiLCPavonLSeverinoPSibovTGuilhenDMoreira-FilhoCAVascular endothelial growth factor-A enhances indoleamine 2,3-dioxy-genase expression by dendritic cells and subsequently impacts lymphocyte proliferationMem Inst Oswaldo Cruz20131091707924141959
- PallottaMTOrabonaCVolpiCGrohmannUPuccettiPFallarinoFProteasomal degradation of indoleamine 2,3-dioxygenase in CD8 den-dritic cells is mediated by suppressor of cytokine signaling 3 (SOCS3)Int J Tryptophan Res20103919722084591
- ThomasSRTerentisACCaiHPost-translational regulation of human indoleamine 2,3-dioxygenase activity by nitric oxideJ Biol Chem200728233237782378717535808
- MaxDKühnölCDBurdachSNiuLStaegeMSFöllJLIndoleamine-2,3-dioxygenase in an immunotherapy model for Ewing sarcomaAnticancer Res2014346431644125368243
- ParkGChoiaYJLeeSEA paradoxical pattern of indoleamine 2,3-dioxygenase expression in the colon tissues of patients with acute graft-versus-host diseaseExp Hematol20144273474024732701
- WangYLiuHMcKenzieGKKynurenine is an endothelium-derived relaxing factor produced during inflammationNat Med201016327928520190767