
Abstract
Lung cancer is a leading cause of cancer death, and late diagnosis is one of the most important reasons for the high mortality rate. microRNAs (miRNAs) are key players in gene regulation and therefore in tumorigenesis. As far as lung carcinogenesis is concerned, miRNAs open novel fields in biomarker research, in diagnosis, and in therapy. In this review we focus on miR-21 in lung cancer and especially on how miR-21 is involved 1) as a biomarker in response or resistance to therapy or 2) as a therapeutic target.
Introduction
Lung cancer remains the number one cause of cancer-related deaths in both men and women, accounting for more than 1.4 million deaths per year.Citation1 It is classified into two main histological subtypes, non-small-cell lung cancer (NSCLC), which accounts for 85% of cases, and small-cell lung cancer (SCLC).Citation2 Although significant improvements have been made toward the early detection of the disease and early diagnosis and treatment, the 5-year overall survival (OS) rate for patients with NSCLC has not improved markedly.Citation3 High mortality is mainly due to late disease presentation at an advanced stage, due to tumor heterogeneity, limited understanding of lung cancer biology, and largely, due to the development of resistance to therapies. Thus, the identification of novel treatment strategies is critical and essential for lung cancer management. Moreover, there is an urgent need to find accurate biomarkers which could detect lung cancer with high sensitivity and specificity in the early state of disease.
microRNAs (miRNAs) are endogenous small noncoding RNAs (17–22 nucleotides), which seem to play a crucial role in tumorigenesis by regulating gene expression at the posttranscriptional level.Citation4,Citation5 The first miRNA was discovered in 1993 by Lee et al,Citation6 and the term microRNA was first presented in 2001.Citation7 Nowadays, more than 2,000 miRNAs have been identified in the human genome, and the vast majority of them are deregulated in cancer cells.Citation4,Citation8 The fact that each miRNA has the ability to target multiple genes within a pathwayCitation9 makes miRNAs one of the most abundant classes of regulatory genes in humans, regulating up to 30% of human protein coding genes.Citation10,Citation11 Due to their important role in many cancer types, miRNAs comprise some of the most attractive targets for therapeutic intervention in cancer today.
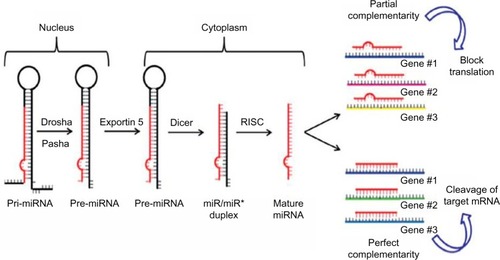
The biogenesis of miRNAs is a multistep process beginning in the nucleus where miRNAs are initially transcribed by RNA polymerase II into a precursor primary miRNA.Citation12 Then the primary miRNA is processed in the nucleus by the RNase III enzyme Drosha and the protein Pasha/DGCR8 into pre-miRNAs (approximately 70-nucleotide).Citation13 Subsequently, this pre-miRNA is transported out of the nucleus into the cytoplasm via Exportin-5,Citation14 where Dicer1 cleaves the hairpin loop of pre-miRNA.Citation15 The functional strand of the mature miRNA is then loaded together with Argonaute (Ago2) proteinsCitation16 into the RNA-induced silencing complex (RISC)Citation17 and finally, the mature single-stranded miRNA is loaded into the RISC, which mediates the degradation or translation inhibition of target mRNA by binding to its seed sequence in the target mRNA’s 3′-UTR ().Citation18,Citation19 miRNAs can be secreted extracellularly bound to lipoproteinsCitation20 or secreted in cell-derived extracellular vesicles such as exosomes as a method of cell-to-cell communication.Citation21 Recently, Melo et alCitation22 showed that breast cancer-associated exosomes contain miRNAs associated with the RISC-loading complex and that these exosome miRNAs display cell-independent capacity to process pre-miRNAs into mature miRNAs.
Figure 1 Biogenesis of miRNAs.
Abbreviations: miRNAs, microRNA; RISC, RNA-induced silencing complex.
miRNAs may act not only within cells, but also at other sites within the body via their circulation in peripheral blood and other body fluids.Citation23 Several studies have shown that serum and plasma miRNAs (called circulating miRNAs) present great promise as novel noninvasive biomarkers for the early diagnosis of various cancers due to their ease of access and long-term stability.Citation24–Citation26 However, as the accurate quantification of circulating miRNAs was more challenging than expected recently, Ferracin et alCitation27 used droplet digital PCR technology in order to assess the absolute levels of nine of the most abundant miRNAs (miR-320a, miR-21-5p, miR-378a-3p, miR-181a-5p, miR-3156-5p, miR-2110, miR-125a-5p, miR-425-5p, miR-766-3p) in serum and plasma of breast, colorectal, lung, and melanoma cancer patients and in healthy controls. According to the final data of this study, there is a clear indication that circulating miRNAs are present in the circulation at levels that can change according to the cancer status and that only one miRNA, namely miR-21-5p, was consistently increased in the plasma of all cancer patients.Citation27
In this review we summarize recent advances in the identification and characterization of miRNAs in lung cancer and especially the role of miR-21 in lung cancer and how miR-21 may be used to facilitate patient diagnosis, prognosis, and response or resistance to therapy.
miRNAs and biology of lung cancer
A growing number of studies suggest an important involvement of miRNAs in various steps of lung carcinogenesis. Several miRNAs are involved in the regulation of cell cycle and lung cancer growth, including in apoptosis, angiogenesis, invasion, and metastasis by targeting different or similar genes.Citation28–Citation31
Apoptosis
The expression of B-cell lymphoma 2 (BCL-2) family gene, one of the central regulators of apoptosis, is regulated by numerous miRNAs. It has been reported that downregulation of miR-7 and miR-335 mediated induction of apoptosis in A549 cells via targeting BCL-2 which subsequently led to an increase in the activities of apoptotic proteins caspase-3 and caspase-7 in cells with apoptotic nuclei.Citation32,Citation33 Moreover, very recently Huang et alCitation34 showed that miR-181 was down-regulated both in NSCLC tissues and cell lines and that this reduction was associated with increased BCL-2 levels. They suggested that miR-181 exerts its proapoptotic function mainly through targeting BCL-2 expression.
Angiogenesis
Phosphatase and tensin homolog (PTEN), a well-known regulator of Akt pathway, has been reported as a target gene of miR-494.Citation35 In this way, miR-494 has a proangiogenic activity through activation of Akt/eNOS pathway. In contrast, miR-200 inhibits angiogenesis through direct and indirect mechanisms especially by targeting interleukin-8 (IL8) and CXCL1 secreted by the tumor endothelial and cancer cells.Citation36
Invasion and metastasis
miR-21 posttranscriptionally downregulates the expression of the tumor suppressor PTEN and subsequently, stimulates growth and invasion in NSCLC.Citation37 Moreover, inhibition of miR-21 expression reduces proliferation, migration, and invasion of A549 cells by upregulating the expression of the programmed cell death protein 4 (PDCD4).Citation38 Li et alCitation39 showed that the expression of miR-183/96/182 cluster is strongly correlated with miR-200 and epithelial–mesenchymal transition in NSCLC as both of these miRNA families target FOXF2, which can potently regulate epithelial–mesenchymal transition, invasion, and metastasis in lung cancers by transcriptional repression of E-cadherin and miR-200.
miR-21
miR-21 is one of the most commonly observed aberrant miRNAs in human cancers and is one of the first miRNAs to be described as an oncomir. A large-scale miRNA analysis on 540 samples in six different types of solid tumors has shown that miR-21 was the only miRNA upregulated in all cancer types.Citation40 Several studies have shown that miR-21 is upregulated in breast,Citation41,Citation42 ovarian,Citation43 colorectal,Citation44 prostate,Citation45,Citation46 pancreatic,Citation47 thyroid cancer,Citation48 as well as in glioma.Citation49,Citation50 Many studies have also clearly shown that miR-21 is deregulated in lung cancer.Citation51,Citation52
In order to understand the way that miR-21 expression is involved in so many different types of cancer, many functional studies have been performed. These studies predicted and validated a number of target genes of miR-21; most of them are tumor suppressors. The most notable and experimentally validated direct targets of miR-21 are discussed in the following sections and .
Figure 2 Notable and experimentally validated direct targets of miR-21.
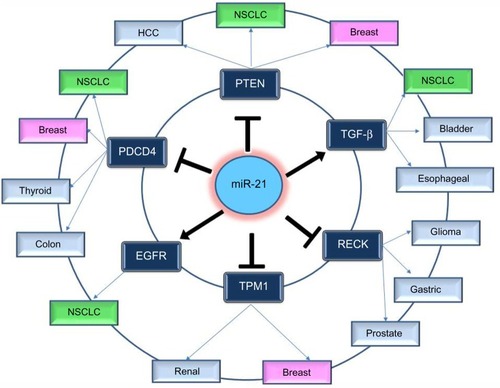
PTEN
Inhibition of miR-21 in cultured hepatocellular carcinoma cells increased the expression of PTEN tumor suppressor, and decreased tumor cell proliferation, migration, and invasion.Citation53 The effect of miR-21 on PTEN expression was also assessed in NSCLC cell lines and in NSCLC tumor samples; it was shown once more that miR-21 represses PTEN and stimulates growth and invasion in NSCLC.Citation37,Citation54,Citation55 High miR-21/low PTEN expression levels indicated a poor tyrosine kinase inhibitor (TKI) clinical response and shorter OS in NSCLC patients and moreover the in vitro assays have showed that miR-21 was upregulated concomitantly to downregulation of PTEN in PC-9 gefitinib resistant cells in comparison with PC-9 cells.Citation56 Overexpression of PTEN gene could also mimic the functions of miR-21 inhibitor in NSCLC cells, while siRNA-mediated PTEN knockdown could reverse the effects on NSCLC cells induced by overexpression of miR-21.Citation54
PDCD4
Knockdown of miR-21 in colon cancer cells led to reduced extravasation and distal metastasis because of the rise of the protein expression levels of PDCD4.Citation44 miR-21 negatively regulates PDCD4 at the posttranscriptional level in colon cancer,Citation44 breast cancer,Citation57 and medullary thyroid carcinoma.Citation58 Moreover, inhibition of miR-21 expression reduces proliferation, migration, and invasion of A549 NSCLC cells by upregulating the expression of PDCD4.Citation38,Citation59
Tropomyosin 1
This gene is a member of the tropomyosin (Tm) family, which plays an important role in cancer cell transformationCitation60 and is a direct target of miR-21. Tropomyosin 1 (TPM1) carries a putative miR-21 binding site, which is responsible for regulation by miR-21 at the translational level.Citation61 Inhibition of miR-21 leads to an increase in TPM1 protein expression levels in breast cancer, but inhibition of TPM1 through miR-21 overexpression results in progressive cell migration and invasion.Citation61 Moreover, the role of miR-21 as regulator in TPM1 was also demonstrated in renal cell carcinoma.Citation62
Transforming growth factor-b
Overexpression of transforming growth factor-b (TGF-β) has been described in several tumor tissues, and it was demonstrated that it is associated with tumor progression and metastasis.Citation63,Citation64 Researches have been performed to study the correlated expression of TGF-β and miR-21 in bladder tumors,Citation65 esophageal cancer,Citation66 and lung cancer.Citation67 In lung cancer, TGF-β1, a central pathological mediator of fibrotic diseases, enhanced miR-21 expression in primary pulmonary fibroblasts, and increasing miR-21 levels promoted the pro-fibrogenic activity of TGF-β1 in fibroblasts.Citation68
Epidermal growth factor receptor pathway
A significant correlation between epidermal growth factor receptor (EGFR) and miR-21 levels in lung carcinoma cell lines and the suppression of miR-21 by an EGFR-TKI suggest that the EGFR signaling is a pathway positively regulating miR-21 expression.Citation51 The association of miR-21 expression with the acquired resistance to EGFR-TKIs in NSCLC cell lines, animal models, and advanced NSCLC patients was further studied by Li et al,Citation69 who found that miR-21 could induce EGFR-TKI resistance through inhibiting PTEN and PDCD4 expression and activating the PI3K/Akt signal pathway. In addition to the miRNA microarray data showing higher level of miR-21 in EGFR mutation cases, the in vitro analyses using NSCLC cell lines also showed that the activated EGFR signaling upregulates miR-21 expression. A statistically significant positive correlation was observed between miR-21 expression level and p-EGFR level in NSCLC cell.Citation51,Citation70
RECK
miR-21 regulation of RECK expression was detected in several types of cancer such as in gliomaCitation71 and in prostate cancer.Citation72 RECK decreases the amount of active MMP-2 and MMP-9 and inhibits metastatic activity in vitro and in vivo through modulation of these MMPs, which are known to be involved in cancer progression.Citation73
miR-21 as a prognostic and diagnostic biomarker in lung cancer
Yanaihara et alCitation74 were the first group that analyzed miRNA microarray data of lung cancer specimens from NSCLC adenocarcinoma patients and indicated that high miR-155 expression was a significantly unfavorable prognostic factor. On the basis of the microarray results, they further selected miR-17, miR-21, and miR-155 to validate in three additional cohorts of NSCLC adenocarcinoma patients and finally suggested that elevated miR-21 expression is associated with worse prognosis in NSCLC patients.Citation75 Additionally, Lu et alCitation76 studied in a cohort of 357 stage I NSCLC patients the expression levels of several miRNAs and found the prognostic value of two miRNA signature. The prognostic value of mature miR-21 in NSCLC was first shown in 2008; in this study it was shown that miR-21 was upregulated in fresh frozen NSCLC tissues in respect to corresponding adjacent noncancerous tissues and that this overexpression correlated with OS.Citation52 Two meta-analyses summarized the data from different studies and concluded that overexpression of miR-21 was an independent prognostic factor for NSCLC.Citation77,Citation78 Moreover, in a meta-analysis study from a total of 141 prognostic miRNAs, it was identified that four miRNAs (miR-155, -21, -34, and let-7) appeared more frequently, and miR-21 and miR-155, especially, were found to be independent prognostic markers associated with unfavorable survival and recurrence in lung cancer.Citation79
Multiple studies have confirmed since then that high expression of miR-21 in plasma and tissues is correlated with worse prognosis in NSCLC patients and could be an independent prognostic marker.Citation30,Citation80–Citation82 However, there are few studies focusing on the potential predictive value of miRNAs for early-stage NSCLC. A study performed by Tang et alCitation80 concluded that overexpression of circulating miR-21 and miR-155 in plasma and lower expression of circulating miR-145 could distinguish lung cancer patients from healthy smokers with 69.4% sensitivity and 78.3% specificity and that this pattern is associated with poor survival. Liu et alCitation81 showed that serum miR-21 may be a potential novel biomarker for the diagnosis of NSCLC as miR-21 has been found to be significantly elevated in the serum of NSCLC patients with lower survival. Studies examining postoperative lung cancer patients also found that miR-21 serum levels significantly decreased in response to successful surgeries, with higher miR-21 expression corresponding to shorter survival time and disease recurrence.Citation82 With regard to the plasma, Geng et alCitation83 showed that five plasma miRNAs (miR-20a, miR-145, miR-21, miR-223, and miR-221) could be used as promising biomarkers in early screening of NSCLC based on the results from plasma samples of 126 early-stage NSCLC patients, 42 noncancerous pulmonary disease patients, and 60 healthy controls.
miR-21 as a predictive biomarker in lung cancer
One of the most important issues facing current cancer research is resistance to treatments such as chemotherapy, radiotherapy, and targeted therapies (). Recent studies are showing that miRNAs can be potentially used as predictive biomarkers in various types of cancer. Blower et alCitation84 first reported that variable miR-21 levels can change sensitivity to the cytotoxic action of drugs in opposite directions suggesting the existence of different mechanisms determining toxicity in cancer cell lines. Several studies have shown that miRNAs can control response to therapy in tumors.Citation85,Citation86 Correlation of miR-21 expression and response to therapy has been studied in many types of cancer. More specifically in: 1) breast cancer where miR-21 overexpression mediates resistance to trastuzumab,Citation87 2) leukemia where miR-21 expression is upregulated in the daunorubicin-resistant leukemia cell line,Citation88 and 3) gastric cancer where miR-21 overexpression induces resistance to cisplatin.Citation89
Table 1 miR-21 controls key biological processes driving to tumorigenesis and therapeutic response in lung cancer
Regarding miRNA expression at the primary tumor, it was reported that NSCLC patients with lower expression of miR-21 seem to have substantial benefits from an adjuvant platinum-based chemotherapy as compared with patients with higher miR-21 expression tumors.Citation90 Additionally, it was found that miR-21 induced the resistance of the NSCLC cell line to cisplatin via targeting the 3′-UTR of PTEN to decrease its expression posttranscriptionally.Citation91 Using miR-21 mimics and its inhibitor, Yang et alCitation91 found that exogenous miR-21 promoted cell survival when exposed to cisplatin treatment, whereas miR-21 inhibition led to the opposite function.
It is now well established that EGFR TKIs, such as gefitinib and erlotinib, constitute the best therapeutic option in first, second, and maintenance setting for NSCLC patients carrying EGFR mutations.Citation92 Unfortunately, half of the patients who initially respond to EGFR-TKI acquire resistance to EGFR-TKI within the 1st year, and in 50% of these cases, a secondary EGFR-T790M gatekeeper mutation has been identified.Citation92,Citation93 miR-21 overexpression was found to reduce sensitivity to gefitinib in PC9 cells by downregulating PTEN and activating Akt and ERK pathways, which enhanced invasive ability and induced apoptosis.Citation56 Furthermore, in the same study, Shen et alCitation56 investigated whether alteration of miR-21/PTEN expression correlates with TKI sensitivity using 47 NSCLC tumor samples from TKI-treated patients. As expected, patients with progressive disease had significantly higher miR-21 levels and lower PTEN expression than patients having partial response and stable disease. These findings are all in agreement and suggest involvement of high miR-21/low PTEN expression in modulation of TKI sensitivity in NSCLC.Citation56
In lung cancer, very recently, Jeon et alCitation94 have shown that overexpression of miR-21, miR-30, and miR-100 could modulate the response to tumor necrosis factor-related apoptosis-inducing ligand by targeting fundamental tumor suppressor genes such as caspase-8, caspase-3, TRAF-7, and FoxO3a and oncogenes such as PI3K and cyclins. They report that in the resistant cells, downmodulation of caspase-8 and TRAF-7 by miR-21 and miR-100 activates NF-κB, which in turn, by a positive feedback loop, induces the transcriptional activation of miR-21 and miR-100.Citation94
Circulating miR-21 in serum was recently shown to be an independent prognostic indicator for NSCLC patients who developed resistance to EGFR-TKI since it was found to be significantly increased in disease progression; high levels of circulating miR-21 is a poor prognostic factor for advanced NSCLC patients treated with EGFR-TKIs.Citation69 Plasma miR-21 was identified as an early detection and chemosensitivity biomarker in patients with advanced NSCLC.Citation95 In this study, Wei et alCitation95 demonstrated that plasma levels of miR-21 were found to be significantly higher in the stable and progressive disease group than in the partial chemotherapeutic response group (cisplatin or carboplatin based), whereas no statistical significance was found between the partial chemotherapeutic response group and the control group. miR-21 overexpression in plasma of NSCLC patients was found to be an independent predictor of response to gefitinib; patients with higher expression of miR-21 in plasma had a significant improvement in OS following an adjuvant therapy with gefitinib compared to those with low expression.Citation96 Plasma miR-21 expression is associated with resistance to platinum-based chemotherapy in NSCLC; downregulation of miR-21 by a locked nucleic acid (LNA) anti-miR-21 in vitro and in vivo enhanced the sensitivity of A549 cells to cis-diamminedichloride platinum II (DDP) through apoptotic signaling.Citation97
Regarding resistance to radiotherapy, Liu et alCitation98 reported that upregulation of miR-21 could inhibit the sensitivity of NSCLC cells to irradiation, while downregulation of miR-21 could significantly enhance the sensitivity of NSCLC cells to irradiation. The potential molecular mechanism of this resistance was further studied by Ma et al,Citation99 suggesting that the effects of miR-21 overexpression on A549 cell apoptosis enhanced by irradiation depend on the activation of PI3K/Akt signaling pathway.
It is important to point out that according to recent findings, exosomes also seem to get involved in the resistance to therapy in lung cancer. As Xiao et alCitation100 showed, exposure of A549 cells to DDP could cause cells to release more exosomes than in normal conditions, and the interaction of these exosomes with other A549 cells could increase the resistance of these A549 cells to DDP. When exposed to DDP, A549 cells were stimulated and the expression level of miR-21 increased significantly both in exosomes and in cancer cells.
miR-21 as a therapeutic target in lung cancer
Restoring or downregulating the expression levels of specific miRNAs could be a promising approach to treat cancer; there are two ways to develop miRNA-based therapeutics: 1) miRNA mimics known as “miRNA replacement therapy”, which are used to restore a loss of function of tumor suppressor genes and 2) miRNA antagonists (antagomirs), which are synthetic RNAs complementary to a specific targeted miRNA that block its function and allow in this way its target mRNA to be translated.Citation101 This approach has a very limited use for clinical applications till now.
The first miRNA replacement therapy, MRX34, is a liposome formulate miR-34 mimic that is intravenously injected.Citation102 This potential drug has entered human clinical trials for patients with unresectable primary liver cancer or solid cancers with liver involvement (http://www.clinicaltrials.gov, NCT01829971). Besides, Santaris Pharma is conducting an early clinical trial, which is currently ongoing, using LNA-anti-miR-122 (PSC-3649) in healthy human subjects. The results of these above-cited studies will provide valuable information on the pharmacokinetic and safety profile of these potential drugs. Finally, inhibition of miR-21 was chosen as one of the promising therapeutic strategies for treating hepatocellular carcinoma, and a miR-21 inhibitor (RG-012) is currently being developed by Regulus Therapeutics.
Numerous studies in lung cancer cells lines use an LNA-based miR-21 (anti-miR-21) in order to knock down miR-21 expression. Anti-miR-21 significantly inhibited growth and induced death in A549 cells through apoptotic signalingCitation51,Citation97 and protected against metastasis of human lung cancer cells through inhibition of NADPH oxidase.Citation103 In the in vivo study, a xenograft tumor assay was designed to manipulate the effect of anti-miR-21, meanwhile, tumor growth curve was recorded and immunohistochemistry of the xenograft tumor was performed.Citation73 As expected, miR-21 inhibitor suppresses NSCLC growth in the xenograft model. Moreover, Xu et alCitation97 transfected A549 cells with LNA anti-miR-21 or scramble control into female nude mice followed by treatment with DDP or phosphate buffer saline and noticed that the size of the implanted tumors (A549 transfected with LNA anti-miR-21) treated with DDP were reduced in comparison with the controls, this enhanced the evidence that downregulation of miR-21 by LNA anti-miR-21 increased the in vivo chemosensitivity. As experiments with the xenograft carcinoma model indicate that one transient transfection with anti-miR-21 is sufficient to cause substantial inhibition of tumor growth, this raises the possibility that anti-miR-21 may have potential therapeutic value.
Future perspectives
miRNA-based therapeutics could be a remarkably smart way for targeting therapy in cancer, since one miRNA in charge of a specific function or mechanism can simultaneously target several downstream genes. miR-21 overexpression is involved in many oncogenic pathways of lung cancer and other malignancies. The development of a targeted therapeutic that aims to silence miR-21 seems to be very promising. However, there are some limitations such as how to reach a sufficient dose within the cells in order to achieve efficient miRNA inhibition and which is the right dose of miR-21. However, most studies so far have been performed in vitro, in vivo studies based on systemic or localized target delivery of anti-miR-21 are still required to be carried out. As miR-21 is overexpressed in most human tumors, therapeutic delivery of anti-miR-21 may still be beneficial for a large number of cancers for which no cure is available. However, there is no doubt that significant work remains to be done, and it is necessary to wait for the pharmacokinetic results of the clinical trials in order to evaluate the safety and the efficiency of these molecules.
Disclosure
The authors report no conflicts of interest in this work.
References
- SiegelRNaishadhamDJemalACancer statistics, 2013CA Cancer J Clin201363113023335087
- HerbstRSHeymachJVLippmanSMLung cancerN Engl J Med20083591367138018815398
- SpiraAEttingerDSMultidisciplinary management of lung cancerN Engl J Med200435037939214736930
- CalinGACroceCMMicroRNA signatures in human cancersNat Rev Cancer2006685786617060945
- StefaniGSlackFJSmall non-coding RNAs in animal developmentNat Rev Mol Cell Biol2008921923018270516
- LeeRCFeinbaumRLAmbrosVThe C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14Cell1993758438548252621
- RuvkunGMolecular biology. Glimpses of a tiny RNA worldScience200129479779911679654
- Di LevaGGarofaloMCroceCMMicroRNAs in cancerAnnu Rev Pathol2014928731424079833
- LimLPLauNCGarrett-EngelePMicroarray analysis shows that some microRNAs downregulate large numbers of target mRNAsNature200543376977315685193
- LewisBPBurgeCBBartelDPConserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targetsCell2005120152015652477
- RajewskyNMicroRNA target predictions in animalsNat Genet200638S8S1316736023
- LeeYKimMHanJMicroRNA genes are transcribed by RNA polymerase IIEMBO J2004234051406015372072
- LeeYAhnCHanJThe nuclear RNase III Drosha initiates microRNA processingNature200342541541914508493
- YiRQinYMacaraIGCullenBRExportin-5 mediates the nuclear export of pre-microRNAs and short hairpin RNAsGenes Dev2003173011301614681208
- HutvágnerGMcLachlanJPasquinelliAEBálintETuschlTZamorePDA cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNAScience200129383483811452083
- PrattAJMacRaeIJThe RNA-induced silencing complex: a versatile gene-silencing machineJ Biol Chem2009284178971790119342379
- LinSLChangDYingSYAsymmetry of intronic pre-miRNA structures in functional RISC assemblyGene2005356323816005165
- NahviAShoemakerCJGreenRAn expanded seed sequence definition accounts for full regulation of the hid 3′ UTR by bantam miRNARNA20091581482219286629
- HaMKimVNRegulation of microRNA biogenesisNat Rev Mol Cell Biol20141550952425027649
- ArroyoJDChevilletJRKrohEMArgonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasmaProc Natl Acad Sci U S A20111085003500821383194
- ValadiHEkstromKBossiosASjostrandMLeeJJLotvallJOExosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cellsNat Cell Biol2007965465917486113
- MeloSASugimotoHO’ConnellJTCancer exosomes perform cell-independent microRNA biogenesis and promote tumorigenesisCancer Cell20142670772125446899
- CortezMABueso-RamosCFerdinJLopez-BeresteinGSoodAKCalinGAMicroRNAs in body fluids – the mix of hormones and biomarkersNat Rev Clin Oncol2011846747721647195
- ChenXBaYMaLCharacterization of microRNAs in serum: a novel class of biomarkers for diagnosis of cancer and other diseasesCell Res200818997100618766170
- MitchellPSParkinRKKrohEMCirculating microRNAs as stable blood-based markers for cancer detectionProc Natl Acad Sci U S A2008105105131051818663219
- SourvinouISMarkouALianidouESQuantification of circulating miRNAs in plasma: effect of preanalytical and analytical parameters on their isolation and stabilityJ Mol Diagn20131582783423988620
- FerracinMLupiniLSalamonIAbsolute quantification of cell-free microRNAs in cancer patientsOncotarget20156145451455526036630
- NicolosoMSSpizzoRShimizuMRossiSCalinGAMicroRNAs – the micro steering wheel of tumour metastasesNat Rev Cancer2009929330219262572
- WhiteNMFatoohiEMetiasMJungKStephanCYousefGMMetastamirs: a stepping stone towards improved cancer managementNat Rev Clin Oncol20118758421045789
- MarkouASourvinouIVorkasPAYousefGMLianidouEClinical evaluation of microRNA expression profiling in non small cell lung cancerLung Cancer20138138839623756108
- LinSGregoryRIMicroRNA biogenesis pathways in cancerNat Rev Cancer20151532133325998712
- XiongSZhengYJiangPLiuRLiuXChuYMicroRNA-7 inhibits the growth of human non-small cell lung cancer A549 cells through targeting BCL-2Int J Biol Sci2011780581421750649
- WangHLiMZhangREffect of miR-335 upregulation on the apoptosis and invasion of lung cancer cell A549 and H1299Tumour Biol2013343101310923740614
- HuangPYeBYangYShiJZhaoHMicroRNA-181 functions as a tumor suppressor in non-small cell lung cancer (NSCLC) by targeting Bcl-2Tumour Biol2015363381338725524579
- MaoGLiuYFangXTumor-derived microRNA-494 promotes angiogenesis in non-small cell lung cancerAngiogenesis201518337338226040900
- PecotCVRupaimooleRYangDTumour angiogenesis regulation by the miR-200 familyNat Commun20134242724018975
- ZhangJGWangJJZhaoFLiuQJiangKYangGHMicroRNA-21 (miR-21) represses tumor suppressor PTEN and promotes growth and invasion in non-small cell lung cancer (NSCLC)Clin Chim Acta201041184685220223231
- YangYMengHPengQDownregulation of microRNA-21 expression restrains non-small cell lung cancer cell proliferation and migration through upregulation of programmed cell death 4Cancer Gene Ther201522232925477028
- LiJTanQYanMmiRNA-200c inhibits invasion and metastasis of human non-small cell lung cancer by directly targeting ubiquitin specific peptidase 25Mol Cancer20141316624997798
- VoliniaSCalinGALiuCGA microRNA expression signature of human solid tumors defines cancer gene targetsProc Natl Acad Sci U S A20061032257226116461460
- IorioMVFerracinMLiuCGMicroRNA gene expression deregulation in human breast cancerCancer Res2005657065707016103053
- YanLXHuangXFShaoQMicroRNA miR-21 overexpression in human breast cancer is associated with advanced clinical stage, lymph node metastasis and patient poor prognosisRNA2008142348236018812439
- IorioMVVisoneRDi LevaGMicroRNA signatures in human ovarian cancerCancer Res2007678699870717875710
- AsanganiIARasheedSANikolovaDAMicroRNA-21 (miR-21) post-transcriptionally downregulates tumor suppressor Pdcd4 and stimulates invasion, intravasation and metastasis in colorectal cancerOncogene2008272128213617968323
- RibasJNiXHaffnerMmiR-21: an androgen receptor-regulated microRNA that promotes hormone-dependent and hormone-independent prostate cancer growthCancer Res2009697165716919738047
- FoliniMGandelliniPLongoniNmiR-21: an oncomir on strike in prostate cancerMol Cancer201091220092645
- DillhoffMLiuJFrankelWCroceCBloomstonMMicroRNA-21 is overexpressed in pancreatic cancer and a potential predictor of survivalJ Gastrointest Surg2008122171217618642050
- SondermannAAndreghettoFMMoulatletACmiR-9 and miR-21 as prognostic biomarkers for recurrence in papillary thyroid cancerClin Exp Metastasis201532652153026007293
- ZhouXZhangJJiaQReduction of miR-21 induces glioma cell apoptosis via activating caspase 9 and 3Oncol Rep20102419520120514462
- QuintavalleCDonnarummaEIaboniMEffect of miR-21 and miR-30b/c on TRAIL-induced apoptosis in glioma cellsOncogene2013324001400822964638
- SeikeMGotoAOkanoTmiR-21 is an EGFR-regulated anti-apoptotic factor in lung cancer in never-smokersProc Natl Acad Sci U S A2009106120851209019597153
- MarkouATsarouchaEGKaklamanisLFotinouMGeorgouliasVLianidouESPrognostic value of mature microRNA-21 and microRNA-205 overexpression in non-small cell lung cancer by quantitative real-time RT-PCRClin Chem2008541696170418719201
- MengFHensonRWehbe-JanekHGhoshalKJacobSTPatelTMicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancerGastroenterology200713364765817681183
- LiuZLWangHLiuJWangZXMicroRNA-21 (miR-21) expression promotes growth, metastasis, and chemo- or radioresistance in non-small cell lung cancer cells by targeting PTENMol Cell Biochem20133721–2354522956424
- ZhangWBaiWZhangWmiR-21 suppresses the anticancer activities of curcumin by targeting PTEN gene in human non-small cell lung cancer A549 cellsClin Transl Oncol201416870871324293118
- ShenHZhuFLiuJAlteration in miR-21/PTEN expression modulates gefitinib resistance in non-small cell lung cancerPLoS One20149e10330525058005
- ZhuSWuHWuFNieDShengSMoYYMicroRNA-21 targets tumor suppressor genes in invasion and metastasisCell Res20081835035918270520
- PennelliGGaluppiniFBarolloSThe PDCD4/miR-21 pathway in medullary thyroid carcinomaHum Pathol201546505725316501
- LuZLiuMStribinskisVMicroRNA-21 promotes cell transformation by targeting the programmed cell death 4 geneOncogene2008274373437918372920
- HelfmanDMFlynnPKhanPSaeedATropomyosin as a regulator of cancer cell transformationAdv Exp Med Biol200864412413119209818
- ZhuSSiMLWuHMoYYMicroRNA-21 targets the tumor suppressor gene tropomyosin 1 (TPM1)J Biol Chem2007282143281433617363372
- LvLHuangFMaoHMicroRNA-21 is overexpressed in renal cell carcinomaInt J Biol Markers20132820120723558936
- GorschSMMemoliVAStukelTAImmunohistochemical staining for TGF-β1 associates with disease progression in human breast cancerCancer Res199252694969521458485
- WeidnerNSempleJPWelchWRFolkmanJTumor angiogenesis and metastasis – correlation in invasive breast carcinomaN Engl J Med199132418
- MonfaredHZiaeeSAHashemitabarMCo-regulated expression of TGF-β Variants and miR-21 in bladder cancerUrol J20131098198724078506
- ZhangYPanTZhongXChengCNicotine upregulates microRNA-21 and promotes TGF-β-dependent epithelial-mesenchymal transition of esophageal cancer cellsTumour Biol2014357063707224756761
- YaoQCaoSLiCMengeshaAKongBWeiMMicro-RNA-21 regulates TGF-β-induced myofibroblast differentiation by targeting PDCD4 in tumor-stroma interactionInt J Cancer20111281783179220533548
- LiuGFriggeriAYangYmiR-21 mediates fibrogenic activation of pulmonary fibroblasts and lung fibrosisJ Exp Med20102071589159720643828
- LiBRenSLiXmiR-21 overexpression is associated with acquired resistance of EGFR-TKI in non-small cell lung cancerLung Cancer20148314615324331411
- MaXKumarMChoudhurySNLoss of the miR-21 allele elevates the expression of its target genes and reduces tumorigenesisProc Natl Acad Sci U S A2011108101441024921646541
- GabrielyGWurdingerTKesariSMicroRNA 21 promotes glioma invasion by targeting matrix metalloproteinase regulatorsMol Cell Biol2008285369538018591254
- HaffnerMCMosbrugerTEsopiDMTracking the clonal origin of lethal prostate cancerJ Clin Invest20131234918492224135135
- Alexius-LindgrenMAnderssonELindstedtIEngströmWThe RECK gene and biological malignancy – its significance in angiogenesis and inhibition of matrix metalloproteinasesAnticancer Res2014343867387325075007
- YanaiharaNCaplenNBowmanEUnique microRNA molecular profiles in lung cancer diagnosis and prognosisCancer Cell20069318919816530703
- SaitoMSchetterAJMollerupSThe association of microRNA expression with prognosis and progression in early-stage, non-small cell lung adenocarcinoma: a retrospective analysis of three cohortsClin Cancer Res2011171875188221350005
- LuYGovindanRWangLMicroRNA profiling and prediction of recurrence/relapse-free survival in stage I lung cancerCarcinogenesis2012331046105422331473
- ZhuWXuBMicroRNA-21 identified as predictor of cancer outcome: a meta-analysisPLoS One20149e10337325098165
- MaXLLiuLLiuXXPrognostic role of microRNA-21 in non-small cell lung cancer: a meta-analysisAsian Pac J Cancer Prev2012132329233422901216
- YangMShenHQiuCHigh expression of miR-21 and miR-155 predicts recurrence and unfavourable survival in non-small cell lung cancerEur J Cancer20134960461523099007
- TangDShenYWangMIdentification of plasma microRNAs as novel noninvasive biomarkers for early detection of lung cancerEur J Cancer Prev20132254054823462458
- LiuXGZhuWYHuangYYHigh expression of serum miR-21 and tumor miR-200c associated with poor prognosis in patients with lung cancerMed Oncol20122961862621516486
- LeeJHVoortmanJDingemansAMMicroRNA expression and clinical outcome of small cell lung cancerPLoS One20116e2130021731696
- GengQFanTZhangBWangWXuYHuHFive microRNAs in plasma as novel biomarkers for screening of early-stage non-small cell lung cancerRespir Res20141514925421010
- BlowerPEChungJHVerducciJSMicroRNAs modulate the chemosensitivity of tumor cellsMol Cancer Ther200871918187804
- DonahueTRNguyenAHMoughanJStromal microRNA-21 levels predict response to 5-fluorouracil in patients with pancreatic cancerJ Surg Oncol201411095295925132574
- OueNAnamiKSchetterAJHigh miR-21 expression from FFPE tissues is associated with poor survival and response to adjuvant chemotherapy in colon cancerInt J Cancer20141341926193424122631
- GongCYaoYWangYUp-regulation of miR-21 mediates resistance to trastuzumab therapy for breast cancerJ Biol Chem2011286191271913721471222
- BaiHXuRCaoZWeiDWangCInvolvement of miR-21 in resistance to daunorubicin by regulating PTEN expression in the leukaemia K562 cell lineFEBS Lett201158540240821187093
- YangSMHuangCLiXFYuMZHeYLiJmiR-21 confers cisplatin resistance in gastric cancer cells by regulating PTENToxicology201330616216823466500
- GaoWLuXLiuLXuJFengDShuYmiRNA-21: a biomarker predictive for platinum-based adjuvant chemotherapy response in patients with non-small cell lung cancerCancer Biol Ther20121333034022237007
- YangZFangSDiYYingWTanYGuWModulation of NF-κB/miR-21/PTEN pathway sensitizes non-small cell lung cancer to cisplatinPLoS One201510e012154725799148
- BellDWGoreIOkimotoRAInherited susceptibility to lung cancer may be associated with the T790M drug resistance mutation in EGFRNat Genet2005371315131616258541
- KobayashiSBoggonTJDayaramTEGFR mutation and resistance of non-small-cell lung cancer to gefitinibN Engl J Med200535278679215728811
- JeonYJMiddletonJKimTA set of NF-κB-regulated microRNAs induces acquired TRAIL resistance in Lung cancerProc Natl Acad Sci U S A201511226E3355E336426080425
- WeiJGaoWZhuCJIdentification of plasma microRNA-21 as a biomarker for early detection and chemosensitivity of non-small cell lung cancerChin J Cancer20113040741421627863
- ShenYTangDYaoRMicroRNA expression profiles associated with survival, disease progression, and response to gefitinib in completely resected non-small-cell lung cancer with EGFR mutationMed Oncol20133075024198203
- XuLHuangYChenDDownregulation of miR-21 increases cisplatin sensitivity of non-small cell lung cancerCancer Genet201420721422024906642
- LiuZLWangHLiuJWangZXMicroRNA-21 (miR-21) expression promotes growth, metastasis, and chemo- or radioresistance in non-small cell lung cancer cells by targeting PTENMol Cell Biochem2013372354522956424
- MaYXiaHLiuYLiMSilencing miR-21 sensitizes non-small cell lung cancer A549 cells to ionizing radiation through inhibition of PI3K/AktBiomed Res Int2014201461786824804226
- XiaoXYuSLiSExosomes: decreased sensitivity of lung cancer A549 cells to cisplatinPLoS One20149e8953424586853
- KrützfeldtJRajewskyNBraichRSilencing of microRNAs with “antagomirs”Nature200543868568916258535
- Mirna TherapeuticsPress release: Mirna therapeutics is first to advance MicroRNA into the clinic for cancer Available from: http://www.mirnarx.comAccessed September 7, 2015
- YanSLiuGPeiCInhibition of NADPH oxidase protects against metastasis of human lung cancer by decreasing microRNA-21Anticancer Drugs20152638839825563770