
Abstract
Introduction
Major depressive disorder (MDD) is a recurrent, devastating mental disorder, which affects >350 million people worldwide, and exerts substantial public health and financial costs to society. Thus, there is a significant need to discover innovative therapeutics to treat depression efficiently. Stress-induced dysfunction in the subtype of neuronal cells and the change of synaptic plasticity and structural plasticity of nucleus accumbens (NAc) are implicated in depression symptomology. However, the molecular and epigenetic mechanisms and stresses to the NAc pathological changes in depression remain elusive.
Materials and methods
In this study, treatment group mice were treated continually with the chronic unpredictable mild stress (CUMS) until expression of depression-like behaviors were found. Depression was confirmed with sucrose preference, novelty-suppressed feeding, forced swimming, and tail suspension tests. We applied high-throughput RNA sequencing to assess microRNA expression and transcriptional profiles in the NAc tissue from depression-like behaviors mice and control mice. The regulatory network of miRNAs/mRNAs was constructed based on the high-throughput RNA sequence and bioinformatics software predictions.
Results
A total of 17 miRNAs and 10 mRNAs were significantly upregulated in the NAc of CUMS-induced mice with depression-like behaviors, and 12 miRNAs and 29 mRNAs were downregulated. A series of bioinformatics analyses showed that these altered miRNAs predicted target mRNA and differentially expressed mRNAs were significantly enriched in the MAPK signaling pathway, GABAergic synapse, dopaminergic synapse, cytokine–cytokine receptor interaction, axon guidance, regulation of autophagy, and so on. Furthermore, dual luciferase report assay and qRT-PCR results validated the miRNA/mRNA regulatory network.
Conclusion
The deteriorations of GABAergic synapses, dopaminergic synapses, neurotransmitter synthesis, as well as autophagy-associated apoptotic pathway are associated with the molecular pathological mechanism of CUMS-induced depression.
Introduction
Major depressive disorder (MDD) is a common, diverse, widespread, and intermittent neuropsychiatric burden that includes the wide range of symptoms, such as inability to feel pleasure, lack of motivation, retraction from social interaction, cognitive difficulties, and changes in appetite.Citation1,Citation2 Depression affects a significant portion of the world’s population annually; according to the World Health Organization, it is the leading cause of disability in the world.Citation3 Extensive research efforts have been done in past decades; however, the etiology of depression is still indefinable, its diagnosis unclear, and the pharmacotherapy ineffective. This may be due to poor understanding of the molecular pathophysiology of depression.Citation4
It has been established that depression is a collection of diseases, with overlapping causal pathways, beginning with interaction among genetic and environmental factors.Citation5,Citation6 The persistent stress to the genetically susceptible persons directs to the deficits of hypothalamus–pituitary–adrenal axis, neurotrophic factors, monoamine factor, and cytokine, which induce neuron atrophy and dysfunction in brain reward circuits, including the nucleus accumbens (NAc), prefrontal cortex (PFC), ventral tegmental area (VTA), and basolateral amygdala, in the depression patients and animal models with depression-like behaviors.Citation7,Citation8 A number of abnormally regulated genes have been reported in genetically susceptible persons.Citation9–Citation11 However, existing data cannot explain mechanism that mediates pathological changes in depression. Thus, it is important to investigate the potential changes in gene expressions and the regulation of these changes, specifically in brain regions that mediate emotional memory and anxiety.
It has been suggested that NAc is essential for emotion processing and receives dense innervations from mood-related structures including PFC, VTA, hippocampus, amygdala, and hypothalamus.Citation12,Citation13 It is regarded as a neural interface between action and motivation, and has an important role in food intake, sexual behavior, reward-motivated behavior, and stress-related behavior.Citation14 The dysfunction in this region may be correlated to low motivation, interest loss, and anhedonia in depression disorder. Recently, various reports have suggested that synaptic plasticity, structural plasticity changes in NAc medium spiny neurons, and reduced activity are implicated in depression symptomology.Citation15–Citation18 Recently, neuromodulation, an intervention targeting the NAc, has been used in patients diagnosed with treatment-resistant depression.Citation19,Citation20 Furthermore, NAc deep brain motivation has also been linked with anxiolytic and antidepressant effect and enhancement of quality of life in patients diagnosed with severe treatment-resistant depression.Citation21
Gene expression disturbance in the brain is most probably linked with environmental factors which lead to depression.Citation22,Citation23 This assumption has been established by analyzing miRNA in depressive patientsCitation24,Citation25 and in mice with depression-like behaviors.Citation26 A reaction chain from stress to miRNA dysregulation, alternations in mRNA/protein expression, and neuron atrophy and function deficit are proposed to be associated with MDD. However, the molecules acknowledged for major depression are exhibited differently in different reports. The inconsistency may be due to different experimental models in different studies.
This study was designed to characterize the NAc transcriptional and post-transcriptional landscape in mice with depression-like behaviors encouraged by chronic unpredictable mild stress (CUMS). We performed an unbiased, genome-wide bioinformatics analysis of mRNA transcriptome profiles and miRNA in the NAc from CUMS-induced depression mice with the use of high-throughput RNA sequencing. The miRNA–mRNA networks were created to probe and illustrate the complexity of stress-induced miRNA regulation. By using different associated analyses and comparisons, we found signal pathways in the NAc associated with stress-induced depression, in order to present the guidelines for understanding the molecular mechanisms of depression and exploring novel therapeutic strategies for this complex disorder.
Materials and methods
Animals
Male C57BL/6J mice (4 weeks, 16–18 g) were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd (Beijing, China). In this study, all procedures were carried out under the Guideline of National Institutes of Health, USA, for the Care and Use of Laboratory Animals and approved by Animal Care and Use Committee of Shandong University of Traditional Chinese Medicine Institutional (SDUTCM201805311223).
CUMS paradigm
The CUMS paradigms were performed with a minor modification according to the previous studies.Citation7,Citation27 All experimental mice were acclimated to environment and sucrose intake for 1 week. We divided the experimental mice into two groups: control and CUMS. The 3–4 mice in the control groups were housed in one cage, and each mouse in the CUMS group was housed in a single cage. The control group was kept in undisturbed cages. A variate-stressor paradigm was used in the CUMS group.
The CUMS protocol consisted of 13 stressors including social isolation, food and water deprivation, empty bottles, soiled cage, restraint space, circadian disturbance, tilted cage, stroboscope, cold (4°C), rotating cage, wet cage, white noise, and space reduction (). This experiment was maintained for 4 weeks until the mice expressed depression-like behavior phenotype.
Behavioral assessments
Behavioral tests were conducted to assess whether the CUMS induced mice expressed depressed phenotype. The novelty-suppressed feeding test (NST) and sucrose preference test (SPT) were used to assess the anhedonia, and the tail suspension test (TST) and forced swimming test (FST) were used to estimate the behavioral despair. Behavioral assessment experiments were performed in the light cycle with a sound-proof behavioral facility.
TST and FST
TST and FST were carried out as previously reported.Citation28
SPT
SPT was conducted with a minor modification according to previous studies.Citation29,Citation30
NST
Mice were observed for NST after 24-hour food deprivation (water was provided ad libitum). Details of the experiment have been previously reported.Citation30
RNA purification and sequencing
The protocol of RNA purification and sequencing has already been reported in our previously published study.Citation7 In brief, if there were depression-like behaviors 24 hours after treatment (CUMS), both control and CUMS mice were anesthetized by using isoflurane and decapitated by guillotine. NAc samples were dissected and immediately placed on the ice-cold glass slides. These tissues were placed into the frozen vials that contained RNA and stored at -80°C for subsequent analyses. Total RNA was isolated from the colon with the RNAiso Plus Kit (TaKaRa, Dalian, China). After extracting RNA, the UV absorbance was used to measure RNA purity and quality. Absorbance at 260 nm was used to measure the amount of nucleic acid present in the sample. Absorbance at 280 nm were used to estimate the amount of protein in the sample. Absorbance at 230 nm was used to determine the amount of other contaminants that may be present in the samples, such as guanidine thiocyanate. The A260/A280 ratios ranging from 1.9 to 2.1 were acceptable ratios for purity in this experiment, and the requirement for A260/A230 ratios was >2.0. RNA concentration was calculated using the 260 nm reading. The samples with total RNA amount >10 µg, the concentration >200 ng/µL, were selected for the construction of transcriptome and small RNA libraries, respectively. The sequencings were done with the use of BGISEQ-500 platform (BGI, Shenzhen, China). The average reading length of two libraries was about 100 bp (pair-end) and 50 bp (single-end).
Bioinformatics
Bioinformatics analysis of transcriptome, miRNA expression profile, and integrated miRNA/mRNA network have been reported in our previous report.Citation7,Citation30
Quantitative RT-PCR for the validations of miRNA and mRNA
To validate the RNA sequencing data, qRT-PCR was used to detect five miRNAs and five mRNAs which were significantly different between control and mice with depression-like behaviors (n=12/group), in which the tissues were included as those samples for sequencing (all gene primers are listed in ). Briefly, total RNA was harvested from the NAc with the RNAiso Plus Kit (TaKaRa). Then, RNA was reverse-transcribed into cDNA with PrimeScript™ RT reagent Kit (TaKaRa). qRT-PCR was carried out using SYBR® Premix Ex Taq™ kit (TaKaRa) on biosystems QuantStudio 7 Flex (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s protocol. The relative expression level of mRNAs in the NAc was normalized to GAPDH. The relative miRNA expression in the NAc was normalized to U6. The 2-ΔΔCt method was used to calculate the qRT-PCR results. Each sample of qRT-PCR was repeated in three replications.
Dual luciferase reporter assay
The 3′-untranslated region (UTR) sequence of Gad1 mRNA was amplified and fused into the NotI and XhoI sites of psiCHECK2. The site-directed mutation of the detected miR-144-3p targeting site of Gad1 mRNA 3′-UTR was constructed with the guideline of Site-Directed Mutagenesis Kit (Stratagene, La Jolla, CA, USA). The miRNA mimics and the negative control (primer sequences are listed in ) were synthesized in Guangzhou Rui Bo Biological Technology Co., Ltd. HEK 293 T cells (24-well plates) were transfected with reporter vectors together with miR-144-3p mimics and inhibitor or the corresponding miR-NC. The renilla luciferase activity was measured and normalized to firefly luciferase activity at 48 hours post-transfection by using Dual-Glo® Luciferase Assay System (E2920; Promega Corporation, Fitchburg, WI, USA). Each experiment was performed in triplicate.
Statistical analyses
The data of the behavior tests, gene analyses, and luciferase activity are presented as mean ± SEM. The association between miRNAs and prediction was investigated by Pearson’s correlation coefficients. Unpaired Student’s t-test was used to evaluate the statistical difference between the two groups. P<0.05 was considered statistically significant.
Results
CUMS induces depression-like behaviors
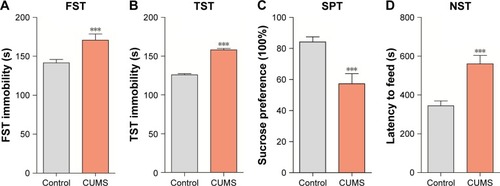
The mood states of CUMS-treated or control group mice were tested by NST, SPT, TST, and FST. Mice exposed to CUMS showed extensively improved immobility in FST () and TST () compared to control group, which demonstrated that those mice were in the status of helplessness and hopelessness. Stressed mice also displayed significantly reduced sucrose preference () and increased latency to feed in the NST (), which showed that those mice were in the mood states of anhedonia and anxiety. Our data indicated that the CUMS paradigm induced depression-like behaviors.
Figure 1 CUMS leads the mice to express depression-like behaviors. Mice were subjected to the adaptation for a week, the CUMS for 4 weeks. (A) The values of immobile time in the FST were 170.68±1.28 seconds in CUMS-treated mice (n=12) and 141.60±1.8 seconds in controls (P<0.01). (B) The values of immobile time in the TST were 158.18±1.79 seconds in CUMS-treated mice and 124.94±0.86 seconds in controls (P<0.001). (C) The SPT values were 57.22%±1.9% in CUMS-treated mice (n=12) and 84.23%±0.92% in control mice (P<0.001). (D) The latency to eat food is 560.55±12.25 seconds in CUMS-treated mice and 344.58±7.22 in controls (P<0.001). The results are expressed as mean ± SEM. n=12 per group, ***P<0.001 compared with control.
Overall qualities of RNA-sequencing data set
In this study, we used RNA-sequencing and assessed miRNA expression and transcriptional profiles in NAc sample from mice with depression-like behaviors and control mice (n=4 per group). About 32,489,852–35,179,878 raw sequence reads were produced in the small RNA library. After trimming and filtering of reads with low quality and adaptor, a total of 29,759,815–32,525,311 clean small RNA reads were obtained (). The high qualities of small RNA sequencing and transcriptome data were further used for analysis. In parallel, RNAs from 45,123,294 to 45,175,652 raw sequence reads of about 100 bp were obtained from eight mRNA library Illumina sequencing. After filtering the reads that contained N with adaptor sequence and low quality, about 1.2 GB clean reads from each library were generated and mapped, which were equivalently about 75.82%–80.62% of the total reads from the University of California Santa Cruz mm10 (). In conclusion, these results from the high qualities of small RNA sequencing and transcriptome data can be used for further analysis.
miRNA expression profiles in NAc from CUMS-induced depression mice
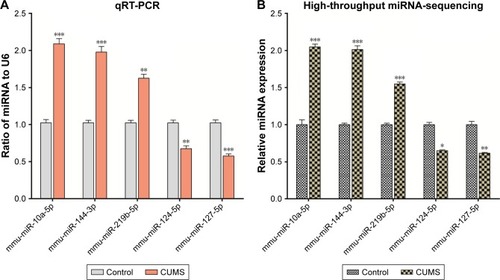
The miRNA expression profile has been given in with their expression change >1.5 fold and P-value <0.05. Seventeen miRNAs were significantly upregulated in the NAc of mice with CUMS-induced depression-like behaviors and 12 miRNAs were downregulated. These upregulated miRNAs included certain known miRNAs (mmu-miR-378d, mmu-miR-34b-5p, mmu-miR-34c-3p, mmu-miR-194-5p, mmu-miR-206-3p, mmu-miR-10a-5p, mmu-miR-144-3p, mmu-let-7a-1-3p, mmu-miR-34c-5p, mmu-miR-34b-3p, mmu-miR-21a-5p, mmu-miR-10b-5p, mmu-miR-31-5p, mmu-miR-219b-5p, and mmu-miR-338-3p), as well as two novel miRNAs (novel_mir42 and novel_mir39). Whereas, the downregulated miRNAs included mmu-miR-7a-2-3p, mmu-miR-124-5p, mmu-miR-351-5p, mmu-miR-127-5p, mmu-miR-671-5p, mmu-miR-335-3p, mmu-miR-377-3p, mmu-miR-574-5p, mmu-miR-200b-5p mmu-miR-486a-5p, mmu-miR-532-3p, and novel_mir29.
Table 1 miRNAs with quantitative change over 1.5-folds and their characteristics
In order to confirm the results of miRNA sequencing analysis, three upregulated miRNAs (mmu-miR-10a-5p, mmu-miR-144-3p, and mmu-miR-219b-5p) and two downregulated miRNAs (mmu-miR-124-5p and mmu-miR-127-5p) were selected for performing qRT-PCR. Consistent with high-throughput sequencing, these miR-NAs from mice with CUMS-induced depression-like behaviors are significantly changed in qRT-PCR compared with those from control mice (). These consistent results from the miRNA sequencing () and qRT-PCR supported the validation of our study.
Figure 2 The validation of differentially expressed miRNAs in the NAc from mice with CUMS-induced depression-like behaviors and controls. Three upregulated miRNAs and two downregulated miRNAs were involved in different cellular functions and were selected for qRT-PCR analysis. (A) qRT-PCR was used to analyze the relative values of mmu-miR-10a-5p, mmu-miR-144-3p, mmu-miR-219b-5p, mmu-miR-124-5p, and mmu-miR-127-5p from mice with CUMS-induced depression-like behaviors and controls (n=12 per group), in which the samples were used from those tissues for high-throughput sequencing. (B) The relative level of mmu-miR-10a-5p, mmu-miR-144-3p, mmu-miR-219b-5p, mmu-miR-124-5p, and mmu-miR-127-5p from mice with CUMS-induced depression-like behaviors (n=4) and controls (n=4), which were analyzed by high-throughput miRNA sequencing. U6 was set as the internal control. The relative values for control mice were normalized to be 1. The data are expressed as mean ± SEM. *P<0.1, **P<0.01, ***P<0.001.
Transcriptional profiles in the NAc from mice with CUMS-induced depression behaviors
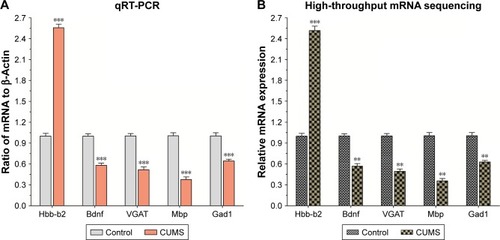
mRNAs in the NAc from mice with depression-like behaviors and controls were quantified by sequencing total RNAs. We computed Reads Per Kilobase Million (RPKM) values for those genes. Genes with lower expression level (RPKM <5) were removed, thus 16,918 mRNAs were left for differential expression. The mRNA expression profile has been given in in which alterations in their expressions were >1.5-fold in mice with depression-like behaviors and controls with P-value <0.05, which was a criterion to ensure differentially expressed genes. Twenty-nine mRNAs were significantly downregulated in the NAc from mice with CUMS-induced depression-like behaviors and 10 mRNAs were upregulated (). The decreased expression of mRNAs in the NAc of mice with CUMS-induced depression-like behaviors included Bdnf, Gad1, Atg13, Slc32a1 (VGAT), Gad2, Th, Mbp, and so on. Whereas, compared to control mice, Aspa, Atg2b, Hba-a1, Hba-a2, Hbb-b2, and Tnfsf12 mRNAs were significantly increased in the NAc of mice with CUMS-induced depression-like behaviors.
Table 2 mRNAs with quantitative change over 1.5-folds and their characteristics
Based on the bioinformatics analysis of Kyoto Encyclopedia of Genes and Genomes (KEGG) database, these differentially expressed genes (DEGs) were significantly enriched in the following signaling pathways and processes, such as the MAPK signaling pathway, GABAergic synapse, dopaminergic synapse, neuroactive ligand–receptor interaction, glutamate metabolism, cytokine–cytokine receptor interaction, axon guidance, regulation of autophagy, apoptosis, and so on (). Thus, the GABAergic synapses, dopaminergic synapses, axon guidance, neurotrophin signaling pathway, neurotransmitter synthesis, autophagy-associated apoptotic pathway, and neural-immune process in the NAc were associated with the pathogenesis of depression. It was noteworthy that the unregulated genes were clustered in hemoglobin genes (Hba-a1, Hba-a2, and Hbb-b2).
Table 3 Signaling pathways identified by KEGG function analysis based on DEG data
In order to verify the differentially expressed genes data, five significantly changed mRNAs involved in different cellular functions were selected for performing qRT-PCR from the NAc tissues that had been used for the mRNA sequencing. The level of Hbb-b2 (P<0.001) gene expression was increased, and the expressions of Bdnf, Slc32a1(VGAT), Mbp, and Gad1 were decreased in mice with CUMS-induced depression-like behaviors compared with that in the control mice (all P<0.001, ). The consistent results obtained by gene sequencing () and qRT-PCR supported our research validation.
Figure 3 The validation of differentially expressed mRNAs in the NAc from mice with CUMS-induced depression-like behaviors and controls.
Abbreviations: CUMS, chronic unpredictable mild stress; NAc, nucleus accumbens; SEM, standard error of the mean.
Integrated miRNA/mRNAregulatory networks
The level of mRNA expression in the cells was affected by miRNAs, through which the bindings of miRNAs with their dicers degrade mRNAs and weaken their translations. If the downregulated mRNAs in the NAc from mice with depression-like behaviors were caused by miRNAs, their corresponding miRNAs will be upregulated. To test this hypothesis and validate our data about mRNA changes, we performed a series of bioinformatics analyses.
The significantly changed miRNAs predicted target mRNAs well match the measures by mRNA sequencing based on the three databases (Targetscan, RNA22, and miRDB) about the complex interactions between miRNAs and mRNAs. shows the alternated miRNAs and their predicted-target mRNAs. For instance, the unregulated mmu-miR-144-3p predicted the downregulated Gad1 and Nr3c1 as the target mRNAs. The altered mRNAs and their corresponding miRNAs are shown in . Among those predicted altered mRNAs, Bdnf and mRNAs could be predicted as the four changed miRNAs (mmu-let-7a-1-3p, mmu-miR-10a-5p, mmu-miR-206-3p, and mmu-miR-10b-5p). From and –, we can find that the altered mRNAs and miRNAs were simultaneously and reversely changed. The consistent results by associatively sequencing mRNAs and miRNAs validated our analyses and strengthen our conclusion.
Table 4 The changed miRNAs predict target mRNAs
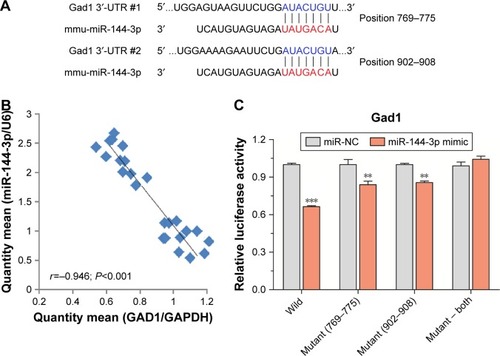
miRNA-144-3p can bind to the GAD1 mRNApredicted region
To confirm the silico prediction, we selected miRNA-144-3p to examine whether they targeted Gad1 by qRT-PCR and dual luciferase reporter assay. Two areas of Gad1 3′-UTRs (769–775 and 902–908) were predicted as the miRNA-144-3p binding site according to the three databases (). The results of qRT-PCR analysis showed that there were negative correlations between miRNA-144-3p and Gad1 mRNA expression in NAc tissue from mice with depression-like behaviors and controls (r=-0.946; P<0.001; ). Furthermore, in dual luciferase report assay, we constructed luciferase reporter plasmids, which contained the two wild-type or corresponding mutant 3′-UTRs of the predicted binding sites. These reporter constructs were transfected into HEK293T cells. After miRNA-144-3p mimics or its negative controls were incubated, the relative activities of luciferase reporter for the 3′-UTR 769–775 and 902–908 of Gad1 mRNA were significantly lowered by the mimics of miRNA-144-3p, which were reversed by mutating the binding sites of miRNA-144-3p. Interestingly, this phenomenon can also be eliminated by mutating both the binding sites of Gad1, which appeared to have the synergistic effect (). These results suggested that miR-144-3p might act as negative regulators of translation through binding to the two sites of Gad1 3′-UTR, which also supported our analyses for the prediction of miRNA target genes.
Figure 4 miRNA-144-3p can bind to the Gad1 mRNA predicted region.
Abbreviations: NC, negative control; SEM, standard error of the mean.
Discussion
MDD is a recurrent, devastating mental disorder, which affects >350 million people worldwide, and exerts substantial public health and financial costs to society. Thus, there is a significant need to discover innovative therapeutics to treat depression efficiently. Growing evidence indicates that dysfunctions in brain reward circuitry are major causes of mood disorders including depression. In the present study, we performed transcriptome analysis in CUMS model and reported differentially expressed genes and miRNAs in the NAc, a key brain region of reward circuitry.
The differentially expressed mRNAs were significantly enriched in GABAergic synapses, dopaminergic synapses, axon guidance, neurotrophin signaling pathway, neurotransmitter synthesis, autophagy-associated apoptotic pathway, and neural-immune process. It is noteworthy that the upregulated mRNAs were clustered in hemoglobin genes. In addition, target miRNAs and mRNAs were upregulated. The inline results from sequencing miRNAs and mRNAs ( and ) as well as qRT-PCR ( and ) fortify our results. Moreover, the consistent results of qRT-PCR and dual luciferase reporter assays also strengthen the bio-informatics analysis of miRNA target prediction (). To the best of our knowledge, here for the first time, we reveal the molecular mechanism of the NAc pathophysiology in CUMS-induced depression.
Various research reports have highlighted the significance of NAc in MDD.Citation12,Citation31–Citation35 In humans, deep brain stimulation of the NAc has been shown to alleviate anhedonic symptoms of depression.Citation36 However, the NAc is a heterogeneous structure comprising different cell types, various dopamine receptors, and other neuromodulatory signaling. These characteristics made it difficult to elucidate the precise role of the NAc circuitry in depressive behaviors. Increasing evidence supports the reduction of synaptic transmission and GABAergic neurons in the NAc associated with depression.Citation37–Citation39 Vulnerable mice exhibit an elevated stubby spine NAc that is associated with high occurrence of miniature excitatory post-synaptic current.Citation40 Electrophysiological record of NAc excitatory diffusion followed stress expose were unbalanced reduction.Citation41 Also, in our previous study, we found that GABAergic neurons were attributed as inhibitory synapse outputs, excitatory, and excitability synapse reception in CUMS-induced depression mice.Citation8 Such indications are completely in-line with our observations (–). As GABAergic neurons regulated the coordination and encoding ability of excitatory neurons,Citation42,Citation43 the attenuation of GABAergic neurons led to the dysfunction of neuronal networks in the NAc. Furthermore, our analyses indicated the functional defect in dopaminergic synapses, axon guidance, neurotrophin signaling pathway, and neuroactive ligand–receptor interaction. As was known to us, depression was characterized as low self-esteem and anhedonia, which might have occurred due to dysfunction of the synapses and neurons in the reward circuits, such as the synaptic transmission deficit and the degeneration of the axons in the NAc and medial PFC.Citation44 Taken together, these results provided insight into mechanisms underlying the physiological absence of the reward circuits in psychiatric disorders for example, bipolar disorder, MDD, attention schizophrenia and post-traumatic stress disorder.
Earlier studies indicated downregulation in the genes of MAPK signaling pathway, neurotransmitter synthesis and myelin-associated protein in rodent animal model of depression-like and depressive patients with the use of qRT-PCR, microarray or in situ hybridization.Citation45–Citation49 These indications were also consistent with the current study in CUMS-induced depression-like mice. In addition, our analysis further implies autophagy-associated apoptotic pathway and neural-immune process, which may bring new insight into molecular mechanism essential depression. These latest findings might be due to the use of advanced mRNA sequencing method.
In this study, the leading observations were that the upregulated mRNAs were clustered in hemoglobin genes. Hemoglobin expression is high in erythrocytes. It has a key role in oxidative stress, response to injury, and neuronal respiration.Citation50,Citation51 Cells can use neuronal hemoglobin to produce hemoglobin derived peptides action on opioid and cannabin receptors.Citation52 Chronic peripheral inflammation was contributed to increase the frontal expression of hemoglobin genes in ischemic and intracerebral hemorrhage patients.Citation53,Citation54 Upregulation of hemoglobin genes (Hbb-b1, Hba-a1, and Hba-a2) has also been found in the medial PFC of mice with CUMS-induced depression-like behaviors in our previous study.Citation7 Human HbF and HbA2 were associated with disease progression in bipolar disorder, which had a shielding function of HbA2 against postpartum episodes.Citation55 Thus, the elevation in hemoglobin genes was found in diverse pathological states. Consistent results of related studies reveal hemoglobin genes as the potential markers of chronic mild stress-induced depression.
miRNAs are a group of single-stranded small non-coding RNA molecules with 19–22 nucleotides, which could post-transcriptionally degrade mRNA or inhibit the translation of mRNA via binding the 3′-UTR section of mRNAs and furthermore influence the expression of target genes.Citation56 In some cases, miRNAs might trigger translation or even control transcription level by binding to precise gene promoters. The role of miRNAs in depression is mainly explained from preclinical studies which showed the role of miRNAs in neurogenesis, neurodevelopment, and synaptic plasticity.Citation57,Citation58 Accumulated facts imply that stress is a significant risk factor in depression and can differentially regulate miRNA-mediated expression of mRNA transcripts in the brain of rodent.Citation59–Citation61 Additionally, mRNAs down-regulation in the mice NAc are rooted by miRNAs, and their related miRNAs will also be upregulated. To validate the hypothesis and strengthen our findings about mRNA changes, we investigated the miRNA expression in NAc by their sequencings from mice with depression-like behaviors.
In our current study, we found that some known miRNAs were significantly changed in mice with CUMS-induced depression-like behaviors (), which degraded mRNAs listed in . In other words, the analysis from miRNA sequencing was consistent with the analysis from mRNA sequencing. Furthermore, these dysregulation miRNAs were reported in previous studies by qRT-PCR, microarray, or situ hybridization in rodent depression animal model or depressive patients.Citation62–Citation68 We next performed a series of bioinformatics analyses for integrated miRNA/mRNA regulatory networks ( and ). For instance, the unregulated mmu-miR-144-3p predicted the downregulated Gad1 and Nr3c1 as the target mRNAs. Whereas, the downregulated mmu-miR-124-5p predicted the unregulated Aspa and Irf7 as the target mRNAs. Among those predicted altered mRNAs, Bdnf mRNA could be predicted as the four changed miRNAs (mmu-let-7a-1-3p, mmu-miR-10a-5p, mmu-miR-206-3p, and mmu-miR-10b-5p).
Table 5 The changed mRNAs are regulated by miRNAs
To verify the silico prediction, we selected miRNA-144-3p to test whether they targeted Gad1 by dual luciferase reporter assay. Our results demonstrated that miR-144-3p acted as negative regulators of translation through attaching with the two sites of Gad1 3′-UTR. miR-144-3p role still remained unclear in depressive disorders. But, some biological mechanisms can explain our judgments. miR-144-3p has extensive expression nature which is enriched in human brain, additionally, also enriched in malignant hematopoietic and normal cells and tissues.Citation69 miR-144-3p is very conserved and has various envisaged targets in both mice and humans. Numerous investigations have revealed that miR-144-3p was implicated against response to stress, aging diseases, and mood stabilizer treatment.Citation7,Citation70,Citation71 miR-144-3p targeting signaling pathways include Nrf2, Wnt/β-catenin, and MAKP pathways.Citation72–Citation74 In addition, miR-144-3p may suppress ataxin 1 (ATXN1) expression in human cells, and interestingly, the Genetic Association Database illustrated that ATXN1 is linked with psychological disorders.Citation75 In terms of response to stress, miR-144-3p level is notably elevated in depressed patients compared with healthy young adults. These studies support that miR-144-3p-mediated target gene expressions were involved in the processes of the pathogenesis of depression.
The quantitative alterations of miRNAs in some reports without the alterations of their targeted mRNAs in others may be caused by the following reasons. The changes in miRNAs may not reach the threshold to regulate their targeted mRNAs.Citation76 circRNAs act as miRNA sponges and positive regulators of miRNA-targeted genes.Citation77 On the other hand, the quantitative changes of mRNAs without the alteration of their corresponding miRNAs imply that the altered mRNAs are likely regulated by other epigenetic mechanisms, such as DNA methylation and repressive histone modification in the promoters.Citation78–Citation80 Therefore, the associated analysis of miRNA/mRNAs in the brain regions with depression-related dysfunction may validate the data and strengthen the conclusion.
Conclusion
In summary, we performed a series of bioinformatics analyses for RNA sequencing data in the NAc tissue from the CUMS-induced depression mice. The worsening of dopaminergic synapses, GABAergic synapses, and neurotransmitter syntheses and autophagy-associated apoptotic pathway are associated with the molecular pathological mechanism of CUMS-induced depression. Our analyses support the string from stress to neuron atrophy through miRNA/mRNA regulatory network and provide the guidelines for developing novel therapeutic strategies for this complex disorder.
Author contributions
All authors contributed toward data analysis, drafting and critically revising the paper, approved the final version to be published, and agree to be accountable for all aspects of the work.
Acknowledgments
This work was supported by the Shandong Co-Innovation Center of Classic TCM Formula, Scientific Innovation Team of Shandong University of Traditional Chinese Medicine, the National Natural Science Foundation of China (81673852 and 81603419), Shandong Province University Scientific Research Project (J18KZ014), Shandong Medical Health Technology Development Plan Project (2018WS203), and Student Research Training Program of Shandong University of TCM (2018090 and 2018093).
Supplementary materials
Table S1 Chronic unpredictable mild stress procedure
Table S2 qRT-PCR prime information
Table S3 3′-Untranslated region (UTR) and site-directed mutation prime sequence Gad1
Table S4 Filtering small RNA library raw data and quality control
Table S5 Filtering transcriptome raw data and alignment
Disclosure
The authors report no conflicts of interest in this work.
References
- AnthesEDepression: a change of mindNature2014515752618518725391944
- SmithKMental health: a world of depressionNature20145157526180181
- GaynesBNWardenDTrivediMHWisniewskiSRFavaMRushAJWhat did STAR*D teach us? Results from a large-scale, practical, clinical trial for patients with depressionPsychiatric Services200960111439144519880458
- MaKZhangHBalochZPathogenetic and therapeutic applications of tumor necrosis factor-α (TNF-α) in major depressive disorder: a systematic reviewInt J Mol Sci2016175733751
- Converge ConsortiumSparse whole-genome sequencing identifies two loci for major depressive disorderNature2015523756258859126176920
- PriceJLDrevetsWCNeural circuits underlying the pathophysiology of mood disordersTrends Cogn Sci2012161617122197477
- MaKGuoLXuACuiSWangJ-HMolecular mechanism for stress-induced depression assessed by sequencing miRNA and mRNA in medial prefrontal cortexPLoS One2016117e015909327427907
- ZhuZWangGMaKCuiSWangJ-HGABAergic neurons in nucleus accumbens are correlated to resilience and vulnerability to chronic stress for major depressionOncotarget2017822359333594528415589
- BoppSKHeilbronnerUSchlattmannPLeptin gene polymorphisms are associated with weight gain during lithium augmentation in patients with major depressionEur Neuropsychopharmacol201929221122130554862
- KangH-JBaeK-YKimS-WMethylation of the glucocorticoid receptor gene associated with depression in patients with acute coronary syndromePsychoneuroendocrinology2019101424930408722
- PanDXuYZhangLGene expression profile in peripheral blood mononuclear cells of postpartum depression patientsSci Rep2018811013929973662
- KnowlandDLimBKCircuit-based frameworks of depressive behaviors: the role of reward circuitry and beyondPharmacol Biochem Behav2018174425229309799
- RussoSJNestlerEJThe brain reward circuitry in mood disordersNat Rev Neurosci201314960962523942470
- FrancisTCLoboMKEmerging role for nucleus accumbens medium spiny neuron subtypes in depressionBiol Psychiatry201781864565327871668
- KravitzAVTyeLDKreitzerACDistinct roles for direct and indirect pathway striatal neurons in reinforcementNat Neurosci201215681681822544310
- AnackerCScholzJO’DonnellKJNeuroanatomic differences associated with stress susceptibility and resilienceBiol Psychiatry2016791084084926422005
- ChandraRLenzJDGancarzAMOptogenetic inhibition of D1R containing nucleus accumbens neurons alters cocaine-mediated regulation of Tiam1Front Mol Neurosci201361323745104
- ChandraRFrancisTCKonkalmattPOpposing role for EGR3 in nucleus accumbens cell subtypes in cocaine actionJ Neurosci201535207927793725995477
- FrancisTCChandraRFriendDMNucleus accumbens medium spiny neuron subtypes mediate depression-related outcomes to social defeat stressBiol Psychiatry201577321222225173629
- DelaloyeSHoltzheimerPEDeep brain stimulation in the treatment of depressionDialogues Clin Neurosci2014161839124733973
- FriedmanAKWalshJJJuarezBEnhancing depression mechanisms in midbrain dopamine neurons achieves homeostatic resilienceScience2014344618131331924744379
- FatemiSHFolsomTDRooneyRJThurasPDExpression of GABAA α2-, β1- and ε-receptors are altered significantly in the lateral cerebellum of subjects with schizophrenia, major depression and bipolar disorderTransl Psychiatry201339e30324022508
- LopizzoNBocchio ChiavettoLCattaneNGene–Environment Interaction in Major Depression: Focus on Experience-Dependent Biological SystemsFront Psychiatry2015686826005424
- MaheuMLopezJPCrapperLDavoliMATureckiGMechawarNMicroRNA regulation of central glial cell line-derived neurotrophic factor (GDNF) signalling in depressionTransl Psychiatry201552e51125689572
- SausESoriaVEscaramísGGenetic variants and abnormal processing of pre-miR-182, a circadian clock modulator, in major depression patients with late insomniaHum Mol Genet201019204017402520656788
- DwivediYRoyBLugliGRizaviHZhangHSmalheiserNRChronic corticosterone-mediated dysregulation of microRNA network in prefrontal cortex of rats: relevance to depression pathophysiologyTransl Psychiatry2015511e68226575223
- XuACuiSWangJ-HIncoordination among subcellular compartments is associated with depression-like behavior induced by chronic mild stressInt J Neuropsychopharmacol2016195pyv12226506857
- TangJXueWXiaBInvolvement of normalized NMDA receptor and mTOR-related signaling in rapid antidepressant effects of Yueju and ketamine on chronically stressed miceScientific Reports2015511357326315757
- SeoJ-SWeiJQinLKimYYanZGreengardPCellular and molecular basis for stress-induced depressionMol Psychiatry201722101440144727457815
- MaKXuACuiSSunM-RXueY-CWangJ-HImpaired GABA synthesis, uptake and release are associated with depression-like behaviors induced by chronic mild stressTransl Psychiatry2016610e91027701406
- BagotRCPariseEMPeñaCJVentral hippocampal afferents to the nucleus accumbens regulate susceptibility to depressionNat Comm2015617062
- NestlerEJRole of the brain’s reward circuitry in depression: transcriptional mechanismsInt Rev Neurobiol201512415117026472529
- GoldenSAChristoffelDJHeshmatiMEpigenetic regulation of Rac1 induces synaptic remodeling in stress disorders and depressionNat Med201319333734423416703
- RobisonAJVialouVSunH-SFluoxetine epigenetically alters the CaMKIIα promoter in nucleus accumbens to regulate ΔFosB binding and antidepressant effectsNeuropsychopharmacology20143951178118624240473
- HorschigJMSmoldersRBonnefondMDirected communication between nucleus accumbens and neocortex in humans is differentially supported by synchronization in the theta and alpha bandPLoS One2015109e013868526394404
- SchlaepferTECohenMXFrickCDeep brain stimulation to reward circuitry alleviates anhedonia in refractory major depressionNeuropsychopharmacology200833236837717429407
- KohnomiSKonishiSMultiple actions of a D3 dopamine receptor agonist, PD128907, on GABAergic inhibitory transmission between medium spiny neurons in mouse nucleus accumbens shellNeurosci Lett2015600172126033184
- YuJYanYLiK-LNucleus accumbens feedforward inhibition circuit promotes cocaine self-administrationProc Nat Acad Sci201711441E8750E875928973852
- AdermarkLSöderpalmBBurkhardtJMBrain region specific modulation of ethanol-induced depression of GABAergic neurons in the brain reward system by the nicotine receptor antagonist mecamylamineAlcohol201448545546124961541
- ChristoffelDJGoldenSADumitriuDIκB kinase regulates social defeat stress-induced synaptic and behavioral plasticityJ Neurosci201131131432121209217
- LimBKHuangKWGrueterBARothwellPEMalenkaRCAnhedonia requires MC4R-mediated synaptic adaptations in nucleus accumbensNature2012487740618318922785313
- LuscherBFuchsTGABAergic control of depression-related brain statesAdvances in pharmacology2015739714425637439
- GaoS-FKlompAWuJ-LSwaabDFBaoA-MReduced GAD(65/67) immunoreactivity in the hypothalamic paraventricular nucleus in depression: a postmortem studyJ Affect Disord20131491–342242523312397
- ProulxCDHikosakaOMalinowRReward processing by the lateral habenula in normal and depressive behaviorsNat Neurosci20141791146115225157511
- ElizaldeNPastorPMGarcia-García ÁlvaroLRegulation of markers of synaptic function in mouse models of depression: chronic mild stress and decreased expression of VGlut1J Neurochem20101951314
- MalemudCJMillerAHPro-inflammatory cytokine-induced SAPK/MAPK and JAK/STAT in rheumatoid arthritis and the new anti-depression drugsExpert Opin Ther Targets200812217118318208366
- CollinsLMDownerEJToulouseANolanYMMitogen-Activated Protein Kinase Phosphatase (MKP)-1 in nervous system development and diseaseMol Neurobiol20155131158116724957007
- LippielloPHoxhaESperanzaLThe 5-HT7 receptor triggers cerebellar long-term synaptic depression via PKC-MAPKNeuropharmacology201610142643826482421
- KériSSzabóCKelemenOBlood biomarkers of depression track clinical changes during cognitive-behavioral therapyJ Affect Disord201416411812224856564
- RichterFMeurersBHZhuCMedvedevaVPChesseletM-FNeurons express hemoglobin alpha- and beta-chains in rat and human brainsJ Comp Neurol2009515553854719479992
- SchelshornDWSchneiderAKuschinskyWExpression of hemoglobin in rodent neuronsJ Cereb Blood Flow Metab200929358559519116637
- GelmanJSSironiJCastroLMFerroESFrickerLDHemopressins and other hemoglobin-derived peptides in mouse brain: comparison between brain, blood, and heart peptidome and regulation in Cpefat/fat miceJ Neurochem2010113487188020202081
- HeYHuaYLiuWHuHKeepRFXiGEffects of cerebral ischemia on neuronal hemoglobinJ Cereb Blood Flow Metab200929359660519066615
- SarlusHWangXCedazo-MinguezASchultzbergMOpricaMChronic airway-induced allergy in mice modifies gene expression in the brain toward insulin resistance and inflammatory responsesJ Neuroinflammation20131019923915208
- InceBGuloksuzSAltınbaşKOralETAlpkanLRAltinozMAMinor hemoglobins HbA2 and HbF associate with disease severity in bipolar disorder with a likely protective role of HbA2 against postpartum episodesJ Affect Disord2013151140540823856286
- TavakolizadehJRoshanaeiKSalmaninejadAMicroRNAs and exosomes in depression: potential diagnostic biomarkersJ Cell Biochem201811953783379729236313
- DwivediYmicroRNAs as biomarker in depression pathogenesisAnn Psychiatry Ment Health201311100326894231
- SaavedraKMolina-MárquezASaavedraNZambranoTSalazarLEpigenetic modifications of major depressive disorderInt J Mol Sci20161781279
- SerafiniGPompiliMHansenKFThe involvement of microR-NAs in major depression, suicidal behavior, and related disorders: a focus on miR-185 and miR-491-3pCell Mol Neurobiol2014341173024213247
- BritesDFernandesANeuroinflammation and depression: microglia activation, extracellular microvesicles and microRNA dysregulationFront Cell Neurosci2015947626733805
- MaffiolettiETarditoDGennarelliMBocchio-ChiavettoLMicro spies from the brain to the periphery: new clues from studies on microRNAs in neuropsychiatric disordersFront Cell Neurosci20148827524653674
- DwivediYEmerging role of microRNAs in major depressive disorder: diagnosis and therapeutic implicationsDialogues in Clin Neurosci20141614361
- MiaoZMaoFLiangJSzyfMWangYSunZSAnxiety-related behaviours associated with microRNA-206-3p and BDNF expression in pregnant female mice following psychological social stressMol Neurobiol20185521097111128092086
- YanHFangMLiuX-YRole of microRNAs in stroke and poststroke depressionSci World J20132013416
- LiuQSunNNZzWFanDHCaoMQChaihu-Shugan-San exerts an antidepressive effect by downregulating miR-124 and releasing inhibition of the MAPK14 and GRIA3 signaling pathwaysNeural Regener Res2018135837845
- HiguchiFUchidaSYamagataHHippocampal microRNA-124 enhances chronic stress resilience in miceJ Neurosci201636277253726727383599
- RoyBDunbarMSheltonRCDwivediYIdentification of MicroRNA-124-3p as a putative epigenetic signature of major depressive disorderNeuropsychopharmacology201742486487527577603
- HussainNZhuWJiangCDown-regulation of miR-10a-5p in synoviocytes contributes to TBX5-controlled joint inflammationJ Cell Mol Med201822124125028782180
- Pienimaeki-RoemerAKonovalovaTMusriMMTranscriptomic profiling of platelet senescence and platelet extracellular vesiclesTransfusion201757114415628026009
- LiuB-BLuoLLiuX-LGengDLiuQYiL-T7-chlorokynurenic acid (7-CTKA) produces rapid antidepressant-like effects: through regulating hippocampal microRNA expressions involved in TrkB-ERK/Akt signaling pathways in mice exposed to chronic unpredictable mild stressPsychopharmacology2015232354155025034119
- GiridharanVVThandavarayanRAFriesGRNewer insights into the role of miRNA a tiny genetic tool in psychiatric disorders: focus on post-traumatic stress disorderTransl Psychiatry2016611e95427845777
- LanFYuHHuMXiaTYueXmiR-144-3p exerts anti-tumor effects in glioblastoma by targeting c-MetJ Neurochem2015135227428626250785
- FengYLiNMaHBeiBHanYChenGUndescribed phenylethyl flavones isolated from Patrinia villosa show cytoprotective properties via the modulation of the mir-144-3p/Nrf2 pathwayPhytochemistry2018153283529859331
- LiJSunPYueZZhangDYouKWangJmiR-144-3p induces cell cycle arrest and apoptosis in pancreatic cancer cells by targeting proline-rich protein 11 expression via the mitogen-activated protein kinase signaling pathwayDNA and Cell Biology201736861962628574724
- PersengievSKondovaIOttingNKoeppenAHBontropREGenome-wide analysis of miRNA expression reveals a potential role for miR-144 in brain aging and spinocerebellar ataxia pathogenesisNeurobiol Aging201132122316e17-2
- DananMSchwartzSEdelheitSSorekRTranscriptome-wide discovery of circular RNAs in ArchaeaNucleic Acids Res20124073131314222140119
- MemczakSJensMElefsiniotiACircular RNAs are a large class of animal RNAs with regulatory potencyNature2013495744133333823446348
- ScalaGMarwahVKinaretPSundJFortinoVGrecoDIntegration of genome-wide mRNA and miRNA expression, and DNA methylation data of three cell lines exposed to ten carbon nanomaterialsData in Brief2018191046105730228994
- SijenTMolecular approaches for forensic cell type identification: on mRNA, miRNA, DNA methylation and microbial markersForensic Sci Int Genet201518213225488609
- DoeckeJDWangYBaggerlyKCo-localized genomic regulation of miRNA and mRNA via DNA methylation affects survival in multiple tumor typesCancer Genetics20162091046347327810075