
Abstract
To date, the role of dopamine D1 receptor (DRD1) polymorphism in schizophrenia remains controversial. We carried out a meta-analysis to determine whether DRD1 polymorphism influences the risk of schizophrenia. We examined whether rs4532 and rs5326 genetic variants are related to the etiology of schizophrenia, using a meta-analysis. Relevant case-control studies were retrieved by database searching and selected according to established inclusion criteria. A total of ten studies were identified and included in our meta-analysis, nine for rs4532, with 1,941 cases and 2,480 controls, and four for rs5326, with 1,285 cases and 1,195 controls. No significant association was found between the rs4532 locus and schizophrenia. For the rs5326 locus, the guanine-adenine (GA) genotype was associated with schizophrenia as a risk factor (for GA vs guanine-guanine [GG], odds ratio [OR] =1.36, 95% confidence interval [CI]: 1.15–1.61, P<0.001). The GA genotype of rs5326 increased the risk of schizophrenia, but there was no association between rs4532 and schizophrenia. These data may provide references for case-control studies in schizophrenia in future.
Keywords:
Introduction
Schizophrenia is a common mental disorder affected by the interaction of multiple genetic and environmental factors, with a median incidence of 15.2/100,000 persons worldwide.Citation1 Evidence from family, adoption, and twins studies supports that its heritability is up to 80%.Citation2 Many efforts have been made over the years, in order to clarify the pathological mechanisms, but a consistent conclusion hasn’t been achieved given that nongenetic factors, such as social and psychological factors, have an impact on the occurrence of schizophrenia and increase the difficulty of the identification of susceptible genes.Citation2 Nevertheless, genome-wide association studies have identified a genetic association by comparing common single nucleotide polymorphisms across the human genome,Citation3 and a number of copy number variants have been suggested as susceptibility factors for schizophrenia.Citation4 To date, dysregulation of dopaminergic neurotransmission has been implicated in the pathogenesis of schizophrenia,Citation5,Citation6 and the reduction of prefrontal cortical dopamine neurotransmission likely leads to schizophrenia.Citation7
Dopamine D1 receptor (DRD1) gene polymorphisms roughly play a role in the occurrence of schizophrenia by affecting the expression of the D1 receptor. Linkage association between DRD1 and schizophrenia has been reported in Chinese,Citation8 American,Citation9 and Portuguese populations.Citation10 Increasing evidence indicates that the polymorphism in the 5′ untranslated region of DRD1 is associated with the mediation of the therapeutic response to antipsychotics,Citation11 Moreover, present studiesCitation12–Citation14 show that deficits in the DRD1 transmission are associated with the cognitive and negative symptoms of schizophrenia and that there is a significant decrease in DRD1 expression in the basal ganglia of schizophrenic patients.Citation15 In contrast, some studies have reported that there was no genetic association between the DRD1 gene and the risk of schizophrenia.Citation16–Citation18 Therefore, the role of the DRD1 gene in schizophrenia remains controversial, and a consistent conclusion has yet to be achieved. The human DRD1 gene rs4532 (A-48G) and rs5326 (G-94A) loci are two commonly studied polymorphisms that are located on the 5′ UTR region. The Rs4532 locus has been associated with therapeutic response to antipsychotics,Citation19 psychotic symptoms, and prefrontal cortex-related cognitive performance.Citation7,Citation20 As for the rs5326 locus, this may act as a short-open-reading frame preceding the receptor cistron and encode a small peptide that inhibits receptor translation.Citation21 Thus, it is necessary and sufficient to estimate the role of rs4532 and rs5326 loci in the pathogenesis of schizophrenia.
There are several possible explanations for this conflicting evidence, such as small sample size, ethnic differences, and publication bias. To systematically synthesize these scattered studies, a potentially useful method is to use meta-analytic techniques, which can combine the results of several studies to generate a single estimate of the major effect with enhanced precision.Citation22 Results from multiple studies are incorporated in an unbiased fashion.Citation22 Meta-analysis is becoming important in psychiatric genetics because of its great improvement and rapid increase of the number and size of data sets. Consequently, in our present study, we carried out a meta-analysis of studies examining the association between DRD1 gene polymorphism and the risk of schizophrenia.
Methods
Identification and eligibility of relevant studies
To identify studies eligible for our meta-analysis, three online electronic databases (PubMed, Web of Science, and China National Knowledge Infrastructure [CNKI]) were searched (the last search update was December 2013). The following key words were used in the literature search: DRD1, dopamine receptor 1, dopamine D1 receptor, dopamine receptor D1, schizophrenia, polymorphism, and variation. The reference lists from identified articles and potentially relevant review articles were also screened to identify additional relevant studies. Studies included in this meta-analysis met the following inclusion criteria: (1) used a case-control design; (2) included patients with schizophrenia and healthy controls; (3) offered available allele or genotype frequencies; (4) the most recent of studies with the same or overlapping data published by the same authors; and (5) published in an English language, peer-reviewed journal. Major reasons for exclusion of studies were: (1) no control population; (2) duplicate of earlier publication; and (3) no usable genotype frequency data.
Data extraction
According to the inclusion criteria listed above, two of the authors extracted information from all eligible publications independently. If there were missing data, we contacted the responsible authors to retrieve additional data not contained in the original paper. Any disagreement was resolved through discussion until the two authors reached a consensus. The following data were included from each study: the first author’s last name, publication year, region, race, study design, source of controls, diagnostic criteria, detection method, and numbers of genotypes between cases and controls.
Statistical analysis
The strength of the association between two loci and schizophrenia was measured by odd ratios (OR), with 95% confidence intervals (CI). Pooled effect sizes across studies were performed by a random effects model, which evaluates the likely effect size across different populations and takes heterogeneity across studies into account. The random effects model is different from a fixed effects model, which evaluates the most likely effect size from multiple studies, hypothesizing that they are sampled from a single population, but it can be biased by high heterogeneity across studies.Citation22
The degree of heterogeneity between studies was performed using a Q-statistic.Citation23,Citation24 A P-value >0.05 for the Q-test indicated a lack of heterogeneity, and P-value <0.05 indicated an existence of heterogeneity. The ICitation2 is the proportion of observed variance in effect sizes attributable to the true differences among studies. A conventional interpretation of ICitation2 is that it defines bounds for low (<25%), moderate (~50%), and high (>75%) heterogeneity.Citation24,Citation25 Subgroup analysis was performed by type of region.
An estimate of publication bias was performed by visual inspection of a funnel plot, in which the standard error of log (OR) of each study was plotted against its log (OR). An asymmetric plot indicated a possible publication bias, and the degree of asymmetry was tested using Egger’s test (P<0.05 was considered to indicate significant publication bias).Citation26
Sensitivity analysis was carried out to assess the potential influences of any single study on the pooled effect size. This was performed by sequentially omitting each study within each meta-analysis, in order to check for significant alterations to the pooled effect sizes. All statistical tests were two-sided. The meta-analysis was performed using Stata version 10.0 (StataCorp LP, College Station, TX, USA).
Results
The search strategy retrieved 44 potentially relevant studies. According to the inclusion criteria, ten articles were identified and included in our meta-analysis (). Among the ten eligible studies, nine studies,Citation16,Citation17,Citation27–Citation33 with 1,941 cases and 2,480 controls, were included in the meta-analysis for the rs4532 locus and four studies,Citation17,Citation32–Citation34 with 1,285 cases and 1,195 controls, in the analysis for rs5326 locus in .
Table 1 Baseline characteristics of the studies included in meta-analysis
Table 2 Genotype frequencies of rs4532 and rs5326 of all studies included in the meta-analysis
Meta-analysis result
lists the main results of pooled OR for the loci of rs4532 and rs5326 and schizophrenia. No significant association was found between the rs4532 locus and schizophrenia (for adenine-guanine [AG] vs adenine-adenine [AA] [OR =0.76, 95% CI: 0.54–1.07, P=0.115] or guanine-guanine [GG] vs AA [OR =0.90, 95% CI: 0.64–1.26, P=0.526]). For the rs4532 locus in the allele contrast model (guanine [G] vs adenine [A]), OR =0.95, 95% CI: 0.76–1.20, P=0.665. For the rs5326 locus, the GA genotype was associated with schizophrenia, as a risk factor (for GA vs GG, OR =1.36, 95% CI: 1.15–1.61, P<0.001), while no association between the AA genotype and schizophrenia was observed (for AA vs GG, OR =0.89, 95% CI: 0.51–1.54, P=0.675). For rs5326 locus in the allele contrast model (A vs G), OR =1.24, 95% CI: 1.16–1.32, P<0.001. The A allele was associated with schizophrenia as a risk factor. The forest plot is shown in .
Figure 1 Forest plot for association between rs4532 and rs5326 loci and schizophrenia.
Abbreviations: CI, confidence interval; OR, odds ratio; A, adenine; G, guanine.
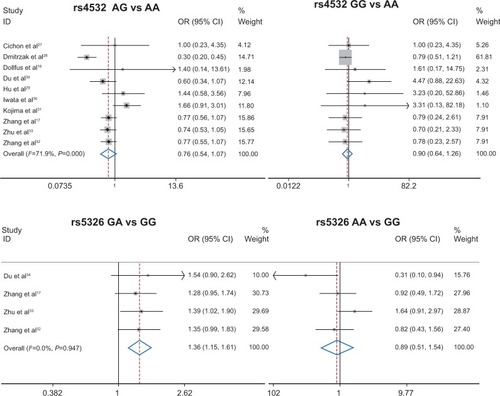
Table 3 Main results of pooled ORs for rs4532 and rs5326
Subgroup analysis
Examination of heterogeneity among the rs4532 locus group across studies was significant. However, the genotype frequency of rs4532 locus had diverse distribution among different races. We carried out the subgroup analysis, by race, in the rs4532 locus group. Races were divided into a Mongolian subgroup (including Chinese and Japanese) and Caucasian subgroup (including German, Polish, and French). lists the results of the subgroup analysis. No significant association was found between the rs4532 locus and schizophrenia in the Mongolian and Caucasian subgroups. Examination of heterogeneity across studies was also not significant, which showed that the heterogeneity among the rs4532 locus group was generated by different races.
Table 4 Subgroup analysis between rs4532 and the risk of schizophrenia
Sensitivity analysis
A sensitivity analysis was carried out for each meta-analysis to assess the influence of every single study. There were no significant differences in the corresponding pooled ORs when removing one study at the time from each meta-analysis, indicating that our results were stable and reliable.
Publication bias
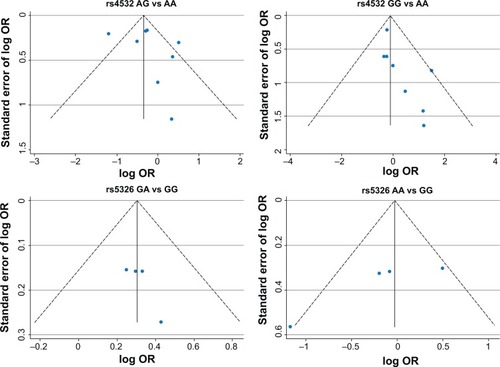
A funnel plot was performed to assess publication bias of the literature. The funnel plot is shown in . Egger’s test was used to supply the statistical evidence for funnel plot symmetry. For rs4532 locus, the results did not show any evidence of publication bias (for AG vs AA [t=0.82, P=0.439] or for GG vs AA [t=2.35, P=0.051]). For the rs5326 locus, the results did not show any evidence of publication bias (for GA vs GG [t=1.94, P=0.192] of for GA vs GG [t=−2.31, P=0.147]).
Figure 2 Funnel plot for evaluating the publication bias in schizophrenia.
Discussion
This was a meta-analysis testing the association between the genetic polymorphisms of the DRD1 gene loci of rs4532 and rs5326 and the risk of schizophrenia. In our study, the GA genotype of the rs5326 locus increased the risk of schizophrenia, and the A allele was also associated with schizophrenia as a risk factor, while no association was observed between rs4532 locus and schizophrenia.
Dopamine receptors play a pivotal role in regulating emotion, cognition, and motor behavior, as well as in neuroendocrine signaling. Prefrontal dopamine D1 receptor signaling is required for temporal control and for guiding intentional movements.Citation35 Abnormal modulation of dopamine receptor signaling and disorder of dopaminergic nerve function may cause a series of neuropsychiatric defects.Citation36 The DRD1 gene has been considered a potential susceptibility gene for various neuropsychiatric illnesses.Citation9,Citation37,Citation38 Furthermore, several previous studies have reported that the DRD1 gene was associated with alcohol dependence,Citation39 hyperactivity disorders,Citation40 nicotine addiction,Citation41,Citation42 and other behavioral disorders.
The DRD1 gene rs4532 and rs5326 loci are located in the 5′ untranslated region, where the nucleotide sequence is involved in transcription but is not being translated into amino acids. A previous meta-analysisCitation43 indicated that the DRD1 gene might contain a genuine susceptibility allele, for which the OR for rs4532 reached 1.18 (95% CI: 1.01–1.38). Furthermore, rs4532 was associated with antipsychotic treatment in schizophrenia,Citation11,Citation19 and also to working memory and cognitive behavior related to the prefrontal cortex.Citation20 These results suggest that rs4532 likely has a potential link with the etiology of schizophrenia. Rs4532, due to a G to A substitution, may act as a short open reading frame preceding the receptor cistron and encode a small peptide that inhibits receptor translation.Citation21 However, the positive or negative association between the rs5326 locus and schizophrenia hasn’t been determined.Citation32,Citation34
Heterogeneity is usually a concern in a meta-analysis. In our study, evidence of heterogeneity was observed in the rs4532 group. This might have been partially due to different races because the genotype frequencies changed greatly in different regions. When subgroup analysis was carried out to explain the heterogeneity, we found that differences in races could lead to the heterogeneity among the studies. The funnel plot was not completely symmetrical, which indicated the potential presence of publication bias. This could have resulted from the following causes: firstly, negative results were less often published compared with positive results, which made it hard for us to collect them for analysis; second, the literature collection wasn’t comprehensive, and the number of published studies in our meta-analysis wasn’t sufficiently large; and third, studies written in other languages were difficult to collect and analyze.
Several limitations in our study should be noted. First, the sample sizes of several studies included were fairly small and lacked a strong statistical power. Furthermore, the factors of sexCitation38 and age may affect the incidence of the disease. In addition, we didn’t consider the role of different environments, and it is possible that environment may also influence the risk of schizophrenia. Further, epigenetic mechanisms altering the chromatin structure, such as histone acetylation and deoxyribonucleic acid methylation, may mediate effects of environmental factors in transcriptional regulation of specific genes and may be a prominent factor in gene–environmental interaction.Citation44 Thus, further studies are needed to investigate the effect of genetic susceptibility and environment in different ethnicities and between males and females, to fully attest to our results. In the future, research into the role of DRD1 gene polymorphisms in schizophrenia endophenotypes could be useful.
In conclusion, our meta-analysis demonstrated that the A allele of the DRD1 rs5326 polymorphism might be a potential risk factor in schizophrenia. As the number of eligible studies was limited in this meta-analysis, these results need further investigation.
Author contributions
Yuqing Pan participated in the design of the study and performed the statistical analysis. Jun Yao carried out the studies and drafted the manuscript. Baojie Wang conceived the study and participated in its design and coordination. All authors contributed toward the drafting and revising of the final manuscript.
Acknowledgments
Special thanks to Hao Pang for critically reviewing the manuscript for important intellectual content.
Disclosure
The authors report no conflicts of interest in this work.
References
- McGrathJSahaSChantDWelhamJSchizophrenia: a concise overview of incidence, prevalence, and mortalityEpidemiol Rev200830677618480098
- SullivanPFKendlerKSNealeMCSchizophrenia as a complex trait: evidence from a meta-analysis of twin studiesArch Gen Psychiatry200360121187119214662550
- SchierdingWCutfieldWSO’SullivanJMThe missing story behind Genome Wide Association Studies: single nucleotide polymorphisms in gene deserts have a story to tellFront Genet201453924600475
- ReesEWaltersJTGeorgievaLAnalysis of copy number variations at 15 schizophrenia-associated lociBr J Psychiatry2014204210811424311552
- HowesODKapurSThe dopamine hypothesis of schizophrenia: version III – the final common pathwaySchizophr Bull200935354956219325164
- Abi-DarghamAMeyerJMSchizophrenia: the role of dopamine and glutamateJ Clin Psychiatry201475327427524717380
- RybakowskiJKBorkowskaACzerskiPMKapelskiPDmitrzak-WeglarzMHauserJAn association study of dopamine receptors polymorphisms and the Wisconsin Card Sorting Test in schizophreniaJ Neural Transm2005112111575158215785860
- ZhengYWangXGuNA two-stage linkage analysis of Chinese schizophrenia pedigrees in 10 target chromosomesBiochem Biophys Res Commun200634241049105716510121
- EscamillaMAOntiverosANicoliniHA genome-wide scan for schizophrenia and psychosis susceptibility loci in families of Mexican and Central American ancestryAm J Med Genet B Neuropsychiatr Genet2007144B219319917044102
- SklarPPatoMTKirbyAGenome-wide scan in Portuguese Island families identifies 5q31–5q35 as a susceptibility locus for schizophrenia and psychosisMol Psychiatry20049221321814699422
- HwangRShinkaiTDe LucaVAssociation study of four dopamine D1 receptor gene polymorphisms and clozapine treatment responseJ Psychopharmacol200721771872717092969
- OkuboYSuharaTSuzukiKDecreased prefrontal dopamine D1 receptors in schizophrenia revealed by PETNature199738566176346369024661
- KarlssonPFardeLHalldinCSedvallGPET study of D(1) dopamine receptor binding in neuroleptic-naive patients with schizophreniaAm J Psychiatry2002159576176711986129
- Abi-DarghamAMawlawiOLombardoIPrefrontal dopamine D1 receptors and working memory in schizophreniaJ Neurosci20022293708371911978847
- SedvallGPauliSKarlssonPPET imaging of neuroreceptors in schizophreniaEur Neuropsychopharmacol19955SupplS25S30
- DollfusSCampionDVasseTAssociation study between dopamine D1, D2, D3, and D4 receptor genes and schizophrenia defined by several diagnostic systemsBiol Psychiatry19964054194218874845
- ZhangCXieBDuYSGene-gene interaction between DNMT3B and DRD1 in schizophreniaZhonghua Yi Xue Za Zhi2010904330593062 Chinese21211326
- HoweASLeungTBani-FatemiALack of association between dopamine-β hydroxylase gene and a history of suicide attempt in schizophrenia: comparison of molecular and statistical haplotype analysesPsychiatr Genet201424311011524710129
- PotkinSGBasileVSJinYD1 receptor alleles predict PET metabolic correlates of clinical response to clozapineMol Psychiatry20038110911312556915
- WilliamsGVCastnerSAUnder the curve: critical issues for elucidating D1 receptor function in working memoryNeuroscience2006139126327616310964
- FungMMRanaBKTangCMDopamine D1 receptor (DRD1) genetic polymorphism: pleiotropic effects on heritable renal traitsKidney Int200976101070108019675531
- MunafòMRFlintJMeta-analysis of genetic association studiesTrends Genet200420943944415313553
- ZintzarasEIoannidisJPHeterogeneity testing in meta-analysis of genome searchesGenet Epidemiol200528212313715593093
- HigginsJPThompsonSGDeeksJJAltmanDGMeasuring inconsistency in meta-analysesBMJ2003327741455756012958120
- HigginsJPThompsonSGQuantifying heterogeneity in a meta-analysisStat Med200221111539155812111919
- EggerMBias in meta-analysis detected by a simple, graphical testBMJ199731571096296349310563
- CichonSNöthenMMRietschelMKörnerJProppingPSingle-strand conformation analysis (SSCA) of the dopamine D1 receptor gene (DRD1) reveals no significant mutation in patients with schizophrenia and manic depressionBiol Psychiatry199436128508537893850
- Dmitrzak-WeglarzMRybakowskiJKSlopienADopamine receptor D1 gene −48A/G polymorphism is associated with bipolar illness but not with schizophrenia in a Polish populationNeuropsychobiology2006531465016397404
- HuYWangGWangHLiuZTangJDistribution of dopamine D1 receptor −48A/G gene polymorphism of healthy Han population in ChinaChinese Journal Of Clinical Rehabilitation2005920152153
- IwataYMatsumotoHMinabeYEarly-onset schizophrenia and dopamine-related gene polymorphismAm J Med Genet B Neuropsychiatr Genet2003116B1232612497608
- KojimaHOhmoriOShinkaiTTeraoTSuzukiAbeKDopamine D1 receptor gene polymorphism and schizophrenia in JapanAm J Med Genet199988211611910206227
- ZhangCFangYXieBNo genetic association between dopamine D1 receptor gene and [early onset] schizophreniaPsychiatry Res2010177335035320382433
- ZhuFYanCXWangQAn association study between dopamine D1 receptor gene polymorphisms and the risk of schizophreniaBrain Res2011142010611321955727
- DuZZhangYChengYZhuC−94 G/A polymorphism in the dopamine D1 receptor gene is associated with schizophrenia in a Chinese Han population from Shandong provinceNeural Regen Res2011061914841487
- NarayananNSLandBBSolderJEDeisserothKDiLeoneRJPrefrontal D1 dopamine signaling is required for temporal controlProc Natl Acad Sci U S A201210950207262073123185016
- GiraultJAGreengardThe neurobiology of dopamine signalingArch Neurol200461564164415148138
- NiXTrakaloJMMundoELinkage disequilibrium between dopamine D1 receptor gene (DRD1) and bipolar disorderBiol Psychiatry200252121144115012488059
- HoenickaJGarridoEPonceGSexually dimorphic interaction between the DRD1 and COMT genes in schizophreniaAm J Med Genet B Neuropsychiatr Genet2010153B494895420127886
- BatelPHouchiHDaoustMRamozNNaassilaMGorwoodA haplotype of the DRD1 gene is associated with alcohol dependenceAlcohol Clin Exp Res200832456757218341651
- BobbAJAddingtonAMSidranskyESupport for association between ADHD and two candidate genes: NET1 and DRD1Am J Med Genet B Neuropsychiatr Genet2005134B1677215717291
- HuangWMaJZPayneTJBeutenJDupontRTLiMDSignificant association of DRD1 with nicotine dependenceHum Genet2008123213314018092181
- NovakGLeBlancMZaiCAssociation of polymorphisms in the BDNF, DRD1 and DRD3 genes with tobacco smoking in schizophreniaAnn Hum Genet201074429129820456319
- AllenNCBagadeSMcQueenMBSystematic meta-analyses and field synopsis of genetic association studies in schizophrenia: the SzGene databaseNat Genet200840782783418583979
- SchmittAMalchowBHasanAFalkaiPThe impact of environmental factors in severe psychiatric disordersFront Neurosci201481924574956