
Abstract
Objective
This study aimed to investigate the impact of S100A4-small interfering RNA (S100A4-siRNA) on apoptosis and enhanced radiosensitivity in non-small-cell lung cancer (A549) cells. We also explored the mechanisms of radiosensitization and identified a new target to enhance radiosensitivity and gene therapy for non-small-cell lung cancer.
Methods
RNA interference is a powerful tool for gene silencing. In this study, we constructed an effective siRNA to knock down S100A4. A549 cells were randomly divided into three groups: blank, negative control, and S100A4-siRNA. To investigate the effect of S100A4-siRNA, the expression of S100A4, E-cadherin, and p53 proteins and their messenger RNA (mRNA) was detected by Western blot and quantitative real-time polymerase chain reaction. Transwell chambers were used to assess cell invasion. Cell cycle and apoptosis were analyzed by flow cytometry. Radiosensitivity was determined by colony formation ability.
Results
Our results demonstrate that S100A4-siRNA effectively silenced the S100A4 gene. When siRNA against S100A4 was used, S100A4 protein expression was downregulated, whereas the expressions of E-cadherin and p53 were upregulated. In addition, a clear reduction in S100A4 mRNA levels was noted compared with the blank and negative control groups, whereas E-cadherin and p53 mRNA levels increased. Transfection with S100A4-siRNA significantly reduced the invasiveness of A549 cells. S100A4 silencing induced immediate G2/M arrest in cell cycle studies and increased apoptosis rates in A549 cells. In clonogenic assays, we used a multitarget, single-hit model to detect radiosensitivity after S100A4 knockdown. All parameters (D0, Dq, α, β) indicated that the downregulation of S100A4 enhanced radiosensitivity in A549 cells. Furthermore, S100A4-siRNA upregulated p53 expression, suggesting that S100A4 may promote A549 cell proliferation, invasion, and metastasis by regulating the expression of other proteins. Therefore, siRNA-directed S100A4 knockdown may represent a viable clinical therapy for lung cancer.
Conclusion
S100A4 downregulation potentially enhances the sensitivity of human A549 cells to radiotherapy.
Introduction
Lung cancer is the leading cause of cancer-related deaths in the world, among which non-small-cell lung cancer (NSCLC) accounts for ~85% of the cases.Citation1–Citation3 Radiotherapy coupled with surgery and chemotherapy is the most important method of contemporary NSCLC treatment.Citation4 However, the use of this therapy is also confronted with challenges due to the intrinsic radioresistance of tumor cells. Hence, enhancing the radiosensitivity of resistant tumor cells is a common strategy in the clinical application of radiotherapy.Citation5
Located in the 1q21 human chromosome region, S100A4 (also known as mts1) is a member of the S100 family of transcription factors.Citation6 S100A4 modulates invasion, metastasis, apoptosis, and cell cycle progression of a variety of malignant tumors through different mechanisms. S100A4 promotes tumor cell invasion and metastasis by downregulating intercellular and cell-substratum adhesion, remodeling the extracellular matrix, altering cytoskeletal dynamics, and promoting angiogenesis.Citation7–Citation10 S100A4 has also been implicated in modulating cell cycle progression, likely by suppressing p53.Citation11 S100A4 also decreases proapoptotic gene expression and inhibits apoptosis.Citation12 S100A4 is detectable in numerous cancer types, and its presence is associated with poor prognosis in many malignant tumors, such as breast,Citation13 bladder,Citation14 esophagus,Citation15 and colon.Citation16 Therefore, inhibi tion of S100A4 may be a good antitumor strategy to reduce cancer cell growth.
Ionizing radiations can result in lethal cell damage, which is correlated with DNA damage induction and repair.Citation17 The tumor suppressor gene p53 encodes a transcriptional regulatory protein that plays a crucial role in controlling cell cycle progression and apoptosis. Grigorian et alCitation18 used several in vitro approaches to demonstrate that S100A4 binds to the extreme end of the p53 C-terminal regulatory domain. Mutations in the p53 gene are noted in almost all types of human cancer with a frequency ranging from 20% to 60%.Citation19 Jiang et alCitation20 demonstrated that β-Elemene increases the radiosensitivity of A549 cells and that the mechanism involved may be related to the upregulation of p53 and induction of cellular apoptosis. When p53 is mutated, cells with DNA damage can escape apoptosis and become cancer cells.Citation21
In this study, we designed a short interfering RNA (siRNA) against S100A4 and evaluated the inhibitory effect of siRNA transfection on cell cycle, apoptosis, and invasion in A549 cells. This effort provides insight into the mechanism by which S100A4 promotes motility, invasion, and metastasis in A549 cells to determine whether S100A4-siRNA sensitizes human NSCLC cells to radiotherapy.
Materials and methods
Cell culture
The study was approved by the ethical committee of Jinshan Hospital of Fudan University. The human lung adenocarcinoma cell line A549 was obtained from the Chinese Academy of Science (Shanghai, People’s Republic of China). Cells were maintained at 37°C in a humidified incubator under 5% CO2 and cultured in RPMI-1640 medium (Sigma-Aldrich Co., St Louis, MO, USA) supplemented with 100 U/mL penicillin, 100 μg/mL streptomycin, and 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific, Waltham, MA, USA). The medium was replaced every 2 or 3 days. Cells in the logarithmic growth phase were used to perform the experi ments described in the following section.
Transfection of siRNA
S100A4-siRNA and nontargeting negative control (NC) siRNA were purchased from GenePharm (Shanghai, People’s Republic of China). The presence of siRNA sequences was confirmed by DNA sequencing. The following sequences were used: S100A4, sense: 5′-UGUCCACCUUCCACAAGUATT-3′ and antisense: 5′-UACUUGUGGAAGGUGGACATT-3′; NC, sense: 5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense: 5′-ACGUGACACGUUCGGAGAATT-3′. Opti-MEM and Lipofectamine 2000 were purchased from the Cell Signaling Technology Corporation (Danvers, MA, USA). A549 cells in logarithmic growth phase were seeded in six-well culture plates. When the cells reached 50% confluence, 20 μM of S100A4-siRNA was transfected with 5 μL of Lipofectamine 2000 and 250 μL of serum-free Opti-MEM medium without antibiotics according to the manufacturer’s instructions. After incubation for 6 hours, the medium was replaced with the standard culture medium as previously described. After an additional 42-hour incubation, cells were used for the experiments described in the following section. The siRNA silencing experiment was repeated thrice.
Western blot analysis
After cell lysis, total protein was collected and run on 12% Bis-Tris gradient gels (Thermo Fisher Scientific) and electropho retically transferred onto polyacrylamide membranes (Thermo Fisher Scientific). Nonspecific binding sites were blocked by incubating the membranes for 1 hour at 37°C with 5% nonfat dried milk in Tris-buffered saline containing 0.05% Tween-20 (TBST). Membranes were incubated overnight at 4°C with the following primary antibodies: anti-S100A4 (1:1,000; Santa Cruz Biotechnology Inc., CA, USA), anti-p53 (1:1,000; Santa Cruz Biotechnology Inc.), anti-E-cadherin (1:400; Santa Cruz Biotechnology Inc.), and anti-β-actin (1:1,000; Abcam, Cambridge, UK). Then, membranes were additionally washed and incubated with a goat anti-rabbit secondary antibody (1:20,000; Abcam). Bands were visualized using an enhanced chemiluminescence system (ECL, Pierce, Rockford, IL, USA).
Quantitative real-time polymerase chain reaction
Total RNA was extracted using TRIzol reagent (Thermo Fisher Scientific) according to the manufacturer’s instructions. RNA concentrations were determined by an ultraviolet spectrophotometer (SHIMADZU UV-2450; Shimadzu Corporation, Kyots, Japan). Subsequently, 2 μg of total RNA was reverse transcribed to produce single-stranded DNA using Super-Script II (Thermo Fisher Scientific). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using the ABI Prism 7700 Sequence Detection System (PerkinElmer Inc., Waltham, MA, USA). The primer sequences were: 5′-AGCAACTTGGACAGCAACAG-3′ (forward) and 5′-TCTTCCTGGGCTGCTTATCT-3′ (reverse) for S100A4; 5′-AGGCCTTGGAACTCAAGGAT-3′ (forward) and 5′-CCCTTTTTGGACTTCAGGTG-3′ (reverse) for p53; 5′-CCAAAGCCTCAGGTCATAAACA-3′ (forward) and 5′-TTCTTGGGTTGGGTCGTTGTAC-3′ (reverse) for E-cadherin. Control PCR was performed using the following primers for GAPDH: 5′-CATCTTCTTTTGCGTCGCCA-3′ (forward) and 5′-TTAAAAGCAGCCCTGGTGACC-3′ (reverse). Quantitative PCR (Q-PCR) assays were performed in triplicate, and the mean values were used to calculate mRNA expression. Gene expression was normalized to GAPDH mRNA. The results were calculated and presented as 2−ΔΔCT.
Cell invasion assay
The transwell device containing microporous 8 μm membranes (Corning Incorporated, Corning, NY, USA) was placed in 24-well plates. Then, 1×104 cells/mL were seeded in the upper chambers and incubated at 37°C with 5% CO2 for 24 hours. Then, the cells were fixed and stained with 0.1% crystal violet. We counted the number of transmembrane cells under an optical microscope, selected three high-power fields at random, and assessed each field of vision to measure the invasion of tumor cells in vitro. Cells were counted and compared among groups. The experiments were repeated thrice.
Colony forming assay
This assay was used to measure the effect of S100A4 silencing on radiation sensitivity of A549 cells. A549 cells were randomly divided into three groups as described earlier: control, NC, and siRNA. All groups were irradiated with X-ray doses of 0, 2, 4, 6, 8, and 10 Gy. The following number of cells were plated for each radiation dose: 500 cells for 0 Gy, 1,000 for 2 Gy, 1,500 for 4 Gy, 2,000 cells for 6 Gy, 3,000 for 8 Gy, and 4,000 for 10 Gy. After 24-hour irradiation, the cells were suspended and incubated for ~14 days in a humidified atmosphere containing 5% CO2 at 37°C with fresh media being replaced every 3 days. Cell colonies were fixed with 80% cold methanol and then stained with 0.4% crystal violet for 1 hour. The colony formation units were evaluated by counting the number of colonies/well in a 5× magnification field. Clones of at least 50 cells were counted as one colony. Survival curves were plotted as the log of survival fraction. The experiment was repeated thrice to obtain the average colony formation rate. A standard radiation survival curve was constructed and the parameters D0, Dq, α (the probability of target cell damage occurs when an ionizing particle breaks through the DNA double strand breaks), and β (the probability of target cell damage occurs when two ionized particles break through the DNA double strand breaks) were calculated with the multitarget, single-hit model. D0 is the dose required to reduce the fraction of surviving cells to 37% of its previous value, and Dq is the repair capacity of the cells after radiation.
Cell cycle analysis
Cell cycle phases were analyzed by measuring the DNA fragment staining with propidium iodide (PI, Sigma-Aldrich Co.) and used according to the manufacturer’s instruction. A549 cells in the exponential growth phase were plated in six-well plates, divided, and treated as noted earlier. The cells were harvested and centrifuged at 24, 48, and 72 hours after exposure to10 Gy irradiation. Cell pellets were washed twice with phosphate-buffered saline. The cells were fixed with 70% ethanol at 4°C overnight and incubated with 200 μL DNAse-free RNAse (Sigma-Aldrich Co.) for 30 minutes at room temperature in the dark. Then, the cells were analyzed immediately by flow cytometry with a FACScanTM system using the CellQuestTM software (Version 3.3) (BD Biosciences, San Jose, CA, USA).
Apoptosis assay
A549 cells grown in six-well plates were divided and treated as mentioned earlier. All the cells in each group were harvested at 24, 48, and 72 hours after irradiation. The experiments were performed with an annexin V apoptosis detection kit (BD Biosciences). The cell pellets were resuspended in 100 mL of binding buffer and stained with 5 mL each of annexin V and PI staining solution for 15 minutes. Then, the samples were analyzed by flow cytometry on a FACScan (Becton Dickinson, Mountain View, CA, USA) using the Cell Quest program.
Statistical analysis
All values were expressed as mean ± standard deviation (SD). The statistics package SPSS 21.0 (SPSS Inc., Chicago, IL, USA) was used for all statistics analyses and mapping. A multitarget, single-hit model was used to fit cells to a survival curve. For all statistical analyses, differences were considered significant if P<0.05.
Results
Effect of S100A4-siRNA on S100A4, E-cadherin, and P53 proteins and their mRNAs in A549 cells
As shown by the Western blot in , the expression of S100A4 protein was downregulated when siRNA against S100A4 was used, whereas the expressions of E-cadherin and p53 were upregulated. The nontargeting siRNA did not significantly affect S100A4, E-cadherin, or p53 protein expression, as demonstrated by comparison of the expression of these proteins between the blank and NC groups. S100A4 protein was reduced both at 24 and 48 hours after transfection but was markedly reduced at 48 hours (), while E-cadherin and p53 proteins were elevated at 48 hours (). Moreover, after transfection of A549 cells with S100A4-siRNA, mRNA levels were detected in a semiquantitative and quantitative manner using qRT-PCR. A clear reduction in S100A4 mRNA levels was observed compared with the blank and NC group, whereas the E-cadherin and p53 mRNA levels were elevated ().
Figure 1 Analysis of S100A4, E-cadherin, and p53 after S100A4-siRNA treatment in A549 cell lines.
Abbreviations: h, hours; siRNA, small interfering RNA; NC, negative control.
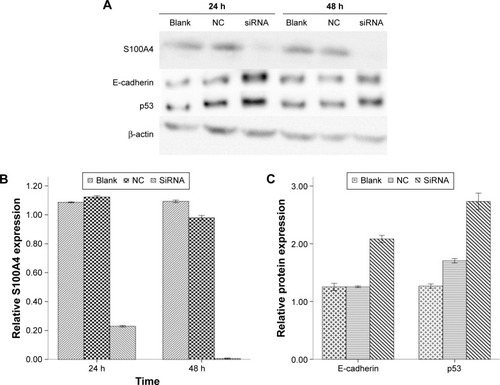
Figure 2 S100A4, E-cadherin, and p53 mRNA expression levels by quantitative real-time PCR both at 24 hours (A) and 48 hours (B).
Abbreviations: mRNA, messenger RNA; PCR, polymerase chain reaction; siRNA, small interfering RNA; NC, negative control.
Invasion assay
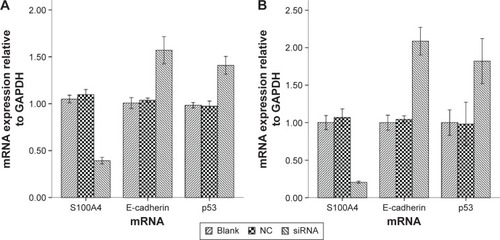
We investigated the invasive capacity of A549 cells after S100A4 silencing. Transwell assays applying Matrigel-coated filters revealed the number of A549 cells penetrating through the artificial basement membrane. The results suggest that S100A4 knockdown cells exhibited markedly reduced invasion compared with the blank and NC groups. Our data suggest that S100A4 plays an important role in cell invasion ().
Figure 3 S100A4 targeting siRNA reduces the invasion of A549 cells.
Abbreviations: siRNA, small interfering RNA; NC, negative control; SD, standard deviation.
Effect of S100A4-siRNA on colony formation by A549 cells
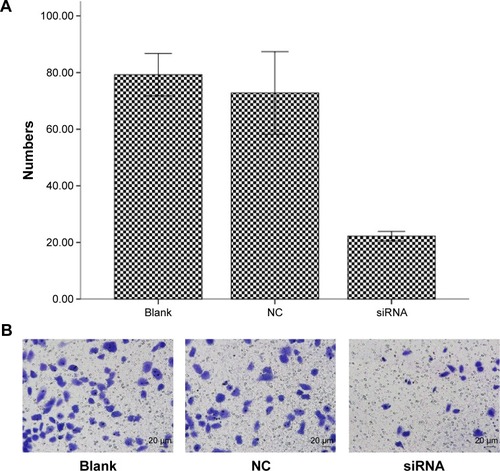
To assess the radiosensitivity after knockdown of S100A4, the multitarget, single-hit model was applied to fit the dose-survival curves. We observed that the cells transfected with NC siRNA exhibited radiosensitivity similar to that exhibited by control cells. Specifically, D0, Dq, sensitization enhancement ratio (SER), and survival rate of irradiated 2Gy cells (SF2) were comparable between both experimental groups as shown in . In terms of radio biological parameters, D0 equaled 3.13 Gy, and the SER, calculated by D0 value, equaled 1.34 Gy in the S100A4-siRNA group. All parameters (D0, Dq, α, β) indicated that transfection with S100A4-siRNA significantly (P<0.05) enhanced the radiosensitivity of A549 cells.
Figure 4 Fitting of dose-survival curves using a multitarget, single-hit model demonstrating the impact of S100A4-siRNA on A549 cells.
A549 cell cycle arrest at the G2/M phase
In the present study, comparison of cell cycle progression after irradiation was conducted by flow cytometry analysis. The effects of S100A4-siRNA on A549 cell cycle progression are presented in . We observed cell cycle arrest at the G2/M phase in cells that were transfected with siRNA against S100A4. The G2/M cell population was significantly different when compared with cells that were transfected with NC siRNA and blank after 10 Gy radiation at 24, 48, and 72 hours (P<0.05), and G2/M arrest was most evident at 24 hours ().
Figure 5 Cell cycle assessment by flow cytometry.
Abbreviations: h, hours; siRNA, small interfering RNA; NC, negative control; S, synthesis; G0, stationary phase; G1, first gap; M, mitosis; G2, second gap.
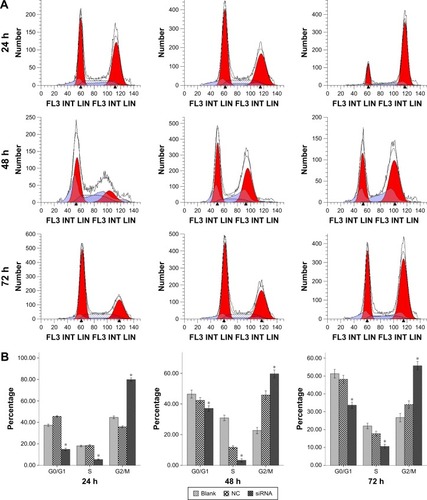
Table 1 Cell cycle at different hours
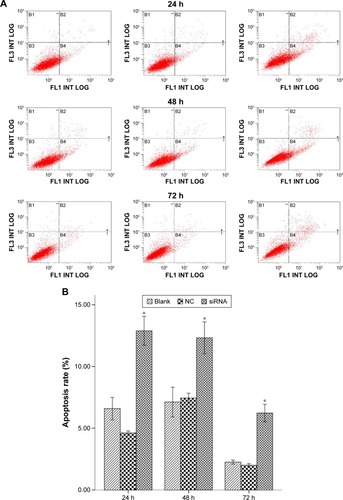
Effect of S100A4-siRNA on apoptosis
After flow cytometry analysis, annexin V-positive cells were used to indicate apoptosis. The apoptotic index for the S100A4-siRNA was significantly increased compared with both NC and blank groups at 24, 48, and 72 hours (, P<0.01).
Figure 6 Flow cytometry analysis for apoptotic cells with annexin V/PI.
Abbreviations: h, hours; siRNA, small interfering RNA; NC, negative control; SD, standard deviation; PI, propidium iodide.
Discussion
S100A4 is a well-established marker and mediator of metastatic disease and tumor growth. We demonstrated that the S100A4 gene is abundantly expressed at both the protein and mRNA levels. It has been previously demonstrated in other cell types that S100A4 downregulation may alter the expression of matrix metalloproteinases (MMPs), E-cadherin, and p53.Citation22,Citation23 The mechanism by which S100A4 protein modulates expression of the E-cadherin and p53 genes in lung cancer cells is not known. Our results suggest that a transcriptional regulation mechanism is involved. In this study, we confirmed the positive correlation between S100A4, p53, and E-cadherin expression in A549 cells. Using an A549 cell line expressing S100A4-siRNA, we demonstrated that knockdown of S100A4 resulted in a decreased expression of p53 and E-cadherin protein and their mRNAs in A549 cells. Our results suggested the possibility that the S100A4 gene could be used as a potential therapeutic target through RNA silencing.
Mahon et alCitation24 demonstrated that knockdown of S100A4 results in an increase in the sensitivity of pancreatic ductal adenocarcinoma cell lines to gemcitabine as well as an increase in the number of apoptotic cells. However, no study has explored the possible correlation between S100A4 expression and lung cancer radiosensitivity to date.
RNA interference (RNAi) reduced mRNA and protein levels of specific genes through posttranscriptional gene silencing mechanisms. This technology is highly efficient, specific, and minimally toxic. This technology is used in functional genomic studies and for therapeutic gene regulation.Citation25,Citation26 In recent years, focus on the impact of S100A4 knockdown on malignant tumors has increased. For example, Ma et alCitation6 reported that siRNA-directed knockdown of S100A4 decreases proliferation and invasiveness of osteosarcoma cells in experiments. Li et alCitation27 demonstrated that S100A4 siRNA inhibits human pancreatic cancer cell invasion in vitro. However, methods to sensitize lung cancer cells to the effects of radiation have remained unclear to date.
Clonogenic survival analysis revealed that S100A4 knockdown markedly potentiated the decrease in NSCLC cell survival induced by radiation. The clonogenic assay acted by modulating the extracellular activity of MMPs.Citation28
Cell cycle regulation is also an important factor in radio-sensitivity. Radiosensitivity can vary in different phases of the cell cycle. Cells are most sensitive to radiation during the G2/M phase, less sensitive during the G1 phase, and least sensitive near the end of the S phase.Citation29 Our study demonstrated that S100A4-siRNA altered the cell cycle with S phase reduction and G2/M phase arrest. These results indicate that S100A4-siRNA acts on the G2/M transition checkpoint of the cell cycle. The cells arrested in the G2/M phase tend to undergo radiation-induced apoptosis; therefore, S100A4-siRNA may be a contributing factor to increased radiosensitivity.
Apoptosis is important in mediating radiosensitivity. Resistance to radiotherapy is a major limiting factor in the treatment of cancer, and abnormal regulation of apoptosis is a contributing factor.Citation30,Citation31 In the present study, radiation alone induced minimal cell apoptosis. When radiation was applied in combination with S100A4-siRNA, the apoptotic index significantly increased, suggesting that S100A4-siRNA sensitized A549 cells to radiation, at least in part, by enhancing radiation-induced apoptosis.
Conclusion
This study further substantiates the proposed role of S100A4 knockdown in potentiating the sensitivity of A549 cells to X-rays by assessing the effect of S100A4 knockdown, which resulted in an increase in cells arrested at the G2/M phase, an increase in cell apoptosis, and a decrease in cell survival. Consequently, S100A4 downregulation potentially could serve as an important anticancer therapy.
Author contributions
All authors read and approved the final manuscript for publication. All authors contributed toward data analysis, drafting, and revising the paper and agree to be accountable for all aspects of the work.
Acknowledgments
This research was supported by a grant from the Science and Technology Council of the Jinshan District, Shanghai, People’s Republic of China (2013-3-02). We thank Dr Liu Bo for performing FACScan analysis.
Disclosure
The authors report no conflicts of interest in this work.
References
- GajraANewmanNGambleGPAbrahamNZKohmanLJGrazianoSLImpact of tumor size on survival in stage IA non-small cell lung cancer: a case for subdividing stage IA diseaseLung Cancer2003421515714512187
- SiegelRNaishadhamDJemalACancer statistics, 2013CA Cancer J Clin2013631113023335087
- SechlerMCizmicADAvasaralaSNon-small-cell lung cancer: molecular targeted therapy and personalized medicine – drug resistance, mechanisms, and strategiesPharmgenomics Pers Med201361253623690695
- KrauseSDebusJNeuhofDRadiotherapyRecent Results Cancer Res201118318328529121509690
- TerrisDJHoEYIbrahimHZEstimating DNA repair by sequential evaluation of head and neck tumor radiation sensitivity using comet assayArch Otolaryngol Head Neck Surg2002128669870212049567
- MaXYangYWangYAnGLvGSmall interfering RNA-directed knockdown of S100A4 decreases proliferation and invasiveness of osteosarcoma cellsCancer Lett2010299217118120855150
- LakshmiMSParkerCSherbetGVExpression of the transmembrane glycoprotein CD44 and metastasis associated 18A2/MTS1 gene in B16 murine melanoma cellsAnticancer Res1997175A345134559413186
- LakshmiMSParkerCSherbetGVMetastasis-associated MTS1and NM23 genes affect tubulin polymerisation in B16 melanomas: a possible mechanism of their regulation of metastatic behaviour of tumorsAnticancer Res19931322993038390799
- LiZHSpektorAVarlamovaOBresnickARMts1 regulates the assembly of nonmuscle myosin-IIABiochemistry20034248142581426614640694
- de Silva RudlandSMartinLRoshaniallCAssociation of S100A4 and osteopontin with specific prognostic factors and survival of patients with minimally invasive breast cancerClin Cancer Res20061241192120016489073
- ChenHFernigDGRudlandPSSparksAWilkinsonMCBarracloughRBinding to intracellular targets of the metastasis-inducing protein, S100A (p9Ka)Biochem Biophys Res Commun200128651212121711527429
- Schmidt-HansenBKlingelhöferJGrum-SchwensenBFunctional significance of metastasis-inducing S100A4 (Mts1) in tumor–stroma interplayJ Biol Chem200427923244982450415047714
- CabezónTCelisJESkibshøjIExpression of S100A4 by a variety of cell types present in the tumor microenvironment of human breast cancerInt J Cancer200712171433144417565747
- DaviesBRO’DonnellMDurkanGCExpression of S100A4 protein is associated with metastasis and reduced survival in human bladder cancerJ Pathol2002196329229911857492
- NinomiyaIOhtaTFushidaSIncreased expression of S100A4 and its prognostic significance in esophageal squamous cell carcinomaInt J Oncol200118471572011251165
- TaylorSHerringtonSPrimeWRudlandPSBarracloughRS100A4 (P9Ka) protein in colon carcinoma and liver metastases: association with carcinoma cells and T-lymphocytesBr J Cancer200286340941611875708
- RodemannHPMolecular radiation biology: perspectives for radiation oncologyRadiother Oncol200992329329819726094
- GrigorianMAndresenSTulchinskyETumor suppressor p53 protein is a new target for the metastasis-associated Mts1/S100A4 protein: functional consequences of their interactionJ Biol Chem200127625226992270811278647
- HollsteinMRiceKGreenblattMSDatabase of p53 gene somatic mutations in human tumors and cell linesNucleic Acids Res19942217355135557937055
- JiangHMaSFengJIn vitro study of radiosensitization by β-Elemene in A549 cell line from adenocarcinoma of lungChinese-German J Clin Oncol2009811215
- LaiPBChiTYChenGGDifferent levels of p53 induced either apoptosis or cell cycle arrest in a doxycycline-regulated hepatocellular carcinoma cell line in vitroApoptosis200712238739317191126
- BoyeKMaelandsmoGMS100A4 and metastasis: a small actor playing many rolesAm J Pathol2010176252853520019188
- BergeGMaelandsmoGMEvaluation of potential interactions between the metastasis-associated protein S100A4 and the tumor suppressor protein p53Amino Acids201141486387320177948
- MahonPCBarilPBhaktaVS100A4 contributes to the suppression of BNIP3 expression, chemoresistance, and inhibition of apoptosis in pancreatic cancerCancer Res200767146786679517638890
- DykxhoornDMPalliserDLiebermanJThe silent treatment: siRNAs as small molecule drugsGene Ther200613654155216397510
- GazzanigaPGradiloneAGiulianiLExpression and prognostic significance of LIVIN, SURVIVIN and other apoptosis-related genes in the progression of superficial bladder cancerAnn Oncol2003141859012488298
- LiNSongMMChenXHLiuLHLiFSS100A4 siRNA inhibits human pancreatic cancer cell invision in vitroBiomed Environ Sci201225446547023026527
- KikuchiNHoriuchiAOsadaRNuclear expression of S100A4 is associated with aggressive behavior of epithelial ovarian carcinoma: an important autocrine/paracrine factor in tumor progressionCancer Sci200697101061106916984379
- PawlikTMKeyomarsiKRole of cell cycle in mediating sensitivity to radiotherapyInt J Radiat Oncol Biol Phys200459492894215234026
- XieYFShengWXiangJZhangHYeZYangJAdenovirus-mediated ING4 expression suppresses pancreatic carcinoma cell growth via induction of cell-cycle alteration, apoptosis, and inhibition of tumor angiogenesisCancer Biother Radiopharm200924226126919409049
- YiTLiHWangXWuZEnhancement radiosensitization of breast cancer cells by deguelinCancer Biother Radiopharm200823335536218593368