
Abstract
MALAT1, an important cancer-associated long noncoding RNA (lncRNA), contributes to the development and progression of several cancers. Disordered expression of MALAT1 has been observed in several cancers, including cervical cancer, breast cancer, and ovarian cancer. However, the exact effects and molecular mechanisms of MALAT1 in ovarian cancer progression are still unknown. Here, we investigated the role of MALAT1 in human ovarian cancer cell lines and clinical tumor samples, in order to determine the function of this molecule. In our research, lncRNA-MALAT1 was specifically upregulated in ovarian cancer cell lines and promoted ovarian cancer-cell growth through targeting microRNA (miR)-506. Knockdown of MALAT1 inhibited the proliferation and DNA synthesis of human ovarian cancer cell in vitro. In addition, miR-506-dependent iASPP regulation was required in MALAT1-induced ovarian cancer-cell growth. These findings indicated that MALAT1 might suppress tumor growth via miR-506-dependent iASPP regulation. Taken together, our data indicated that MALAT1 might be an oncogenic lncRNA that promotes proliferation of ovarian cancer and could be regarded as a therapeutic target in human ovarian cancer.
Keywords:
Introduction
Human genome-sequence data indicate that more than 90% of the DNA sequence is actively transcribed, but only 2% of it encodes protein, and thus the majority of transcripts are referred to as long noncoding RNAs (lncRNAs).Citation1,Citation2 Small ncRNAs, such as microRNAs (miRNAs), have been studied extensively, and their roles in gene regulation and cell function have been elucidated in numerous cancers.Citation2 Recent studies have shown that lncRNAs play important roles in both normal development and diseases, including cancer.Citation3 lncRNAs have emerged as new players in cancer research, and several studies have shown that some lncRNAs function as oncogenes, tumor-suppressor genes, or both, depending on the circumstances.Citation4
Several lncRNAs have been reported to be involved in ovarian cancer. HOTAIR, a cell cycle-associated lncRNA, has been revealed to be overexpressed in ovarian cancer, and significantly correlated with advanced tumor stage, high histological grade, and poor prognosis. Knockdown of HOTAIR could notably suppress cellular proliferation, inhibit invasion, and decrease the secretion of MMP2 and MMP9 in ovarian cancer.Citation5 In addition, Ruan et al reported that the lncRNA SNHG12 gene promoted cell proliferation and migration by upregulating AMOT gene expression in ovarian cancer cells in vivo and in vitro.Citation6
MALAT1 is located on chromosome 11q13, and the expression of MALAT1 has been reported to be upregulated in several cancers, including lung, breast, pancreas, liver, colon, uterus, cervix, and prostate cancer.Citation7 MALAT1 expression is an independent prognostic parameter for survival in several cancers.Citation7 Cai et al reported that the lncRNA MALAT1 plays an oncogenic role in ovarian cancer and may be a promising therapeutic target for the treatment of ovarian cancer patients.Citation8 Moreover, MALAT1 has been reported to be an oncogenic lncRNA that promotes proliferation and metastasis of ovarian cancer via the PI3K–Akt pathway.Citation9 Although emerging evidence has suggested that MALAT1 is associated with ovarian cancer progression and prognosis, the underlying mechanism remains unclear.
iASPP, a member of the ASPP family, was considered as an oncogene that inhibits apoptotic cell death typically after DNA damage. The high expression of iASPP in cancers is associated with worse disease status, therapy resistance, and poor survival of patients.Citation10 iASPP has been reported to be associated with the chemoresistance of ovarian cancer;Citation11 however, whether iASPP is involved in ovarian cancer-cell growth remains unclear.
In this study, we report an interaction between MALAT1 and miRNA (miR)-506, which regulates ovarian cancer-cell growth through direct targeting of iASPP. Our findings provide a novel understanding of the role of MALAT1 and miR-506 in ovarian cancer progression and the mechanism involved.
Materials and methods
Cells and reagents
The human ovarian cancer-cell lines SKOV3, A2780, HO8910, and CAOV3 and normal human ovarian surface epithelial cells (HOSEPiCs) were purchased from the American Type Culture Collection. Trizol reagent and Lipofectamine 2000 were from Thermo Fisher Scientific (Waltham, MA, USA), SYBR green master mix was from Takara (Kyoto, Japan), Dulbecco’s Modified Eagle’s medium and fetal bovine serum were from Thermo Fisher Scientific, and 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium was from Beyotime (Haimen, China).
Tissue specimens
Thirty paired ovarian cancer specimens and corresponding adjacent nontumor tissues were collected from tumor surgical resections in Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University (Guangzhou, China). All human tissues were obtained with written informed consent, and this study was approved by the Clinical Research Ethics Committee of Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University.
Cell transfection
Both the miRNA mimics and small-interfering RNAs (siRNAs) were synthesized by GenePharma (Shanghai, China). Oligonucleotide and plasmid transfection were conducted by using the Lipofectamine 2000 transfection reagent, followed by the protocol recommended by the manufacturer. After 48-hour transfection, the cells were collected and used for further investigations.
MTT assay
Following transfection, cell proliferation was assessed using MTT assay (Promega Corporation, Fitchburg, WI, USA), according to the manufacturer’s instructions. Cells were seeded into 96-well plates at a density of 5,000 cells per well. After 24 hours, cells were transfected with 100 nM miR-506 mimic/miR-506 negative control (NC), si-MALAT1/si-NC, or si-iASPP/si-NC. After an additional 24 hours, 20 μL of 5 mg/mL MTT (Sigma-Aldrich, St Louis, MO, USA) was added into each well and incubated for 4 hours in a humidified incubator. The supernatant was then discarded, and 200 μL of dimethyl sulfoxide was added to each well to dissolve the formazan. Optical density was measured at 490 nm. The viability of the cells without any treatment was defined as 100%, and the viability of other groups was calculated separately from that of the sham cells.
BrdU incorporation assay
Bromodeoxyuridine (BrdU) assays were performed to determine DNA synthesis at 24 and 48 hours after transfecting SKOV3 and A2780 cells with miR-506 mimic/miR-506 NC, si-MALAT1/si-NC, or si-iASPP/si-NC. The infected cells were cultured for 24 or 48 hours and incubated with a final concentration of 10 μM BrdU (BD Biosciences, San Jose, CA, USA) for 2–24 hours. At the end of the incubation period, the medium was removed, the cells were fixed for 30 minutes at room temperature, incubated with peroxidase-coupled anti-BrdU antibody (Sigma-Aldrich) for 60 minutes at room temperature, washed with phosphate-buffered saline, incubated with peroxidase substrate (tetramethylbenzidine) for 30 minutes, and absorbance values were measured at 450 nm.
RNA extraction and real-time PCR
Total RNA was isolated by Trizol reagent following the manufacturer’s instructions. After RNA extraction, RNA samples were reverse-transcribed with high-capacity complementary (cDNA) reverse-transcription kit (Thermo Fisher Scientific). Fast Start Universal SYBR Green Master (Hoffman-La Roche, Basel, Switzerland) was applied for quantitative reverse-transcription polymerase chain reaction (PCR). The following primers were used to detect the expression of MALAT1 and GAPDH (internal control): MALAT1 (sense), 5′-AAAGCAAGGTCTCCCCACAAG-3′; MALAT1 (antisense), 5-GGTCTGTGCTAGATCAAAAGGCA-3′; GAPDH (sense), 5′-AGAAGGCTGGGGCT-CATTTG-3′; and GAPDH (antisense), 5′-AGGGGCCATCCACAGTCTTC-3′. The relative fold changes of candidate genes were analyzed by using the 2−ΔΔCT method.
Western blot
Cell lysates were lysed by radioimmunoprecipitation-assay buffer (Sigma-Aldrich) with a complete protease-inhibitor cocktail (Roche, USA). Cell lysates were transferred to a 1.5 mL tube and kept at −20°C before use. Sodium dodecyl sulfate polyacrylamide-gel electrophoresis was conducted to separate the cellular proteins. All the cellular proteins within this study were separated by 5% stacking gel and 10% running gel. The molecular weight of candidate proteins was determined according to prestained SeeBlue rainbow marker (Thermo Fisher Scientific), loaded in parallel. The membranes were probed with the following antibodies: MALAT1 (Santa Cruz Biotechnology, Dallas, TX, USA), iASPP (Abcam, Cambridge, UK), and β-actin (Sigma-Aldrich). The blots were detected with Kodak film developer (FujiFilm, Tokyo, Japan).
Luciferase-reporter assay
With the Lipofectamine 2000 transfection reagent, SKOV3 cells cultured in 24-well plates were cotransfected with luciferase-reporter plasmids and miRNA mimics, as well as the internal control: purified respiratory syncytial virus (pRSV)–β-galactosidase vector. At 48-hour posttransfection, SKOV3 cells were lysed by lysis buffer (25 mM Tris phosphate, 1% Triton X-100, 1 mM dithiothreitol, 2 mM ethylenediaminetetraacetic acid, 10% glycerol, pH 27.8). After centrifugation at 14,000 rpm for 3 minutes, the supernatant was transferred to a new 1.5 mL tube. Luciferase activity was monitored by mixing 50 μL supernatant with 50 μL luciferase-assay buffer using a GloMax 20/20 luminometer (Promega). β-Galactosidase activity from the pRSV–β-galactosidase vector was used for normalization of the luminescence levels. O-nitrophenyl-β-galactoside (ONPG) colorimetric assays were performed to measure β-galactosidase activity. Supernatant (50 μL) from the aforementioned cell extract was mixed with 100 μL of ONPG solution (0.666 mg/mL ONPG, 40 mM NaH2PO4, 60 mM Na2HPO4, 10 mM KCl, 1 mM MgSO4, 2% β-mercaptoethanol). β-Galactosidase activity was evaluated by the measurement of O-nitrophenol using an enzyme-linked immunosorbent-assay plate reader (Bio-Rad Laboratories, Hercules, CA, USA) at a wavelength of 450 nm.
Statistics
Experimental results are presented as mean ± standard deviation. Comparisons between two groups were conducted using two-tailed Student’s t-test, and differences were considered statistically significant when P<0.05.
Results
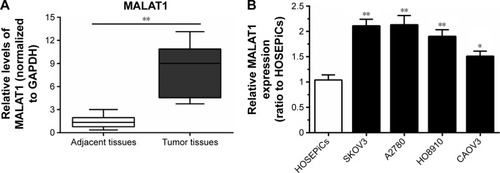
lncRNA-MALAT1 specifically upregulated in ovarian cancer tissues and cell lines
Initially, the expression levels of MALAT1 in 30 paired samples (ovarian cancer specimens and corresponding adjacent nontumor tissues) were examined using real-time PCR. Results showed that MALAT1 expression was significantly higher in tumor tissues than adjacent tissues (). Then, MALAT1 expressions in four human ovarian cancer cell lines – SKOV3, A2780, HO8910, and CAOV3 – and normal HOSEPiCs were determined using real-time PCR. Results showed that in all human ovarian cancer cell lines, expression levels of MALAT1 were upregulated compared to HOSEPiCs ().
Figure 1 Long noncoding RNA MALAT1 was specifically upregulated in ovarian cancer tissues and cell lines.
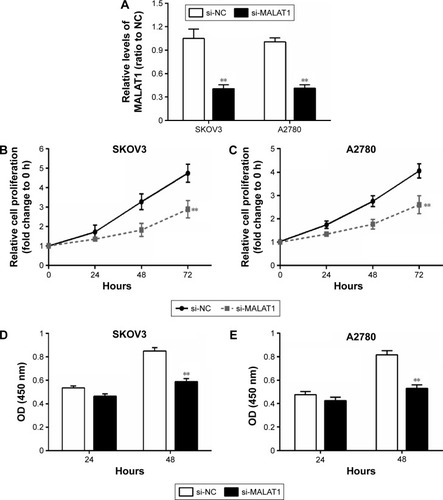
MALAT1 promoted ovarian cancer-cell growth
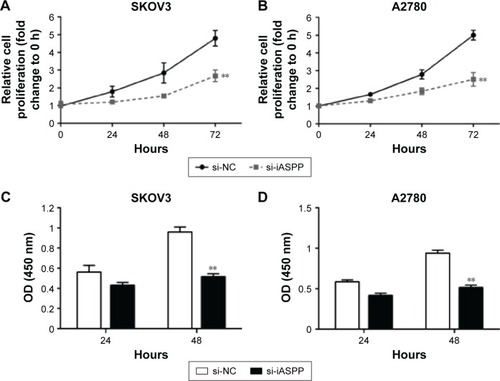
Next, we investigated the association of MALAT1 expression with ovarian cancer-cell proliferation and DNA synthesis. MALAT1 knockdown was achieved by si-MALAT1, and inhibitory efficiency was verified by real-time PCR (). Two ovarian cancer-cell lines, SKOV3 and A2780, were transfected with si-NC or si-MALAT1, and then cell proliferation and DNA synthesis were determined by MTT and BrdU assays. MTT assays revealed that knockdown of MALAT1 significantly attenuated the proliferation of both the SKOV3 and A2780 cell lines over time compared with the si-NC group (). BrdU assay revealed that knockdown of MALAT1 markedly reduced DNA synthesis of both the SKOV3 and A2780 cell lines () compared with the si-NC group. Taken together, these data revealed that lncRNA-MALAT1 was specifically upregulated in ovarian cancer tissues and cell lines and promoted ovarian cancer-cell growth and DNA synthesis.
Figure 2 MALAT1 promoted ovarian cancer-cell growth.
Abbreviations: si, small interfering; PCR, polymerase chain reaction; NC, negative control; OD, optical density.
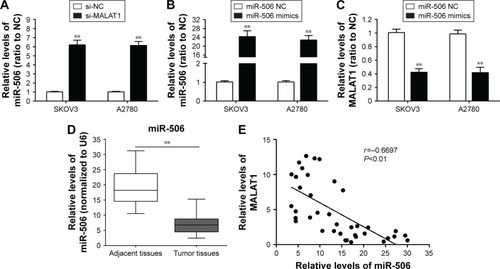
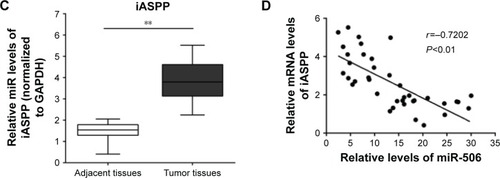
miR-506 expression downregulated in ovarian cancer tissues and negatively correlated with lncRNA-MALAT1
According to previous studies, the miR-376 family plays a suppressive role in ovarian cancer.Citation12,Citation13 To investigate further the mechanism by which MALAT1 regulates ovarian cancer-cell growth, we detected the association of miR-506 with MALAT1. Results from real-time PCR showed that the expression levels of miR-506 were significantly upregulated after knockdown of MALAT1 compared with the si-NC group (). Then, miR-506 mimics were used to achieve miR-506 overexpression, and expression levels of miR-506 were verified by real-time PCR in the SKOV3 and A2780 cell lines (). The expression levels of MALAT1 were determined by real-time PCR in response to miR-506 overexpression. Results showed that MALAT1 expression was decreased in response to miR-506 overexpression compared with the miR-506 NC group (). Moreover, expression levels of miR-506 were significantly downregulated in tumor tissues compared with adjacent normal tissues (). We examined the potential correlation among the RNA-expression levels of MALAT1 and miR-506, and an inverse correlation between MALAT1 and miR-506 expressions was observed ().
Figure 3 miR-506 expression was downregulated in ovarian cancer tissues and negatively correlated with lncRNA-MALAT1.
Abbreviations: miR, microRNA; lncRNA, long noncoding RNA; PCR, polymerase chain reaction; si, small interfering; NC, negative control.
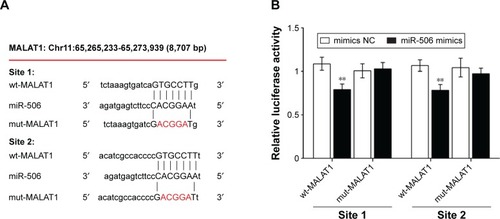
MALAT1 regulated miR-506 by direct targeting
To detect the mechanism by which MALAT1 regulates miR-506, a wild-type (wt)-MALAT1 3′-untranslated region (UTR) luciferase-reporter vector (wt-MALAT1) and a mutant (mut)-MALAT1 3′-UTR luciferase-reporter vector (mut-MALAT1) were constructed by sequentially mutating the two predicted miR-506-binding sites in the MALAT1 3′-UTR (). We cotransfected the wt-MALAT1/mut-MALAT1 vectors and miR-506 NC/miR-506 mimics into SKOV3 cells. The luciferase activity of the wt-MALAT1 3′-UTR luciferase-reporter vector was significantly reduced in miR-506 mimic-transfected cells compared to control groups (). Moreover, miR-506-mediated repression of MALAT1 3′-UTR luciferase-reporter activity was abolished by mutation of the putative miR-506-binding site in the MALAT1 3′-UTR ().
Figure 4 MALAT1 regulated miR-506 by direct targeting.
Abbreviations: miR, microRNA; wt, wild type; UTR, untranslated region; mut, mutant; NC, negative control.
miR-506 inhibited ovarian cancer-cell growth
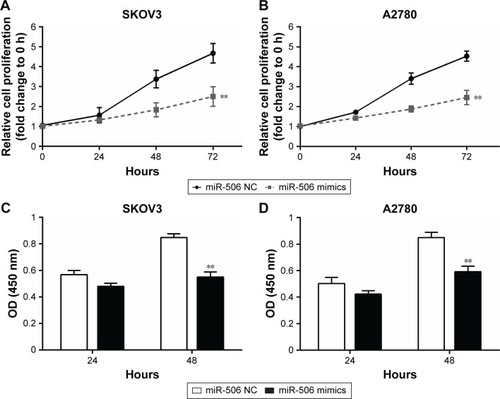
It has been indicated that miR-506 functions as a tumor suppressor in several cancers.Citation14,Citation15 Here, MTT assay was used to determine ovarian cancer-cell growth in response to miR-506 overexpression. It was observed that cell proliferations of both the SKOV3 and A2780 cell lines were reduced when miR-506 was overexpressed compared with the miR-506 NC group (). Results from BrdU assays indicated that miR-506 overexpression markedly reduced DNA synthesis of both the SKOV3 and A2780 cell lines compared with the miR-506 NC group ().
Figure 5 miR-506 inhibited ovarian cancer-cell growth.
Abbreviations: miR, microRNA; NC, negative control; OD, optical density.
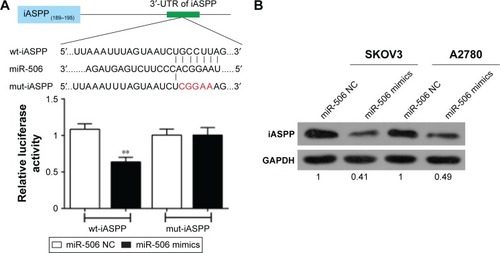
miR-506 regulated iASPP expression by direct targeting
iASPP has been reported to be the direct target of miR-376c, one member of the miR-376 family.Citation12 In order to investigate the association of miR-506 with iASPP in ovarian cancer, we created a wt-iASPP 3′-UTR luciferase reporter vector (wt-iASPP), as well as a mut-iASPP 3′-UTR luciferase reporter vector (mut-iASPP) by sequentially mutating the predicted miR-506-binding site in the iASPP 3′-UTR (). We cotransfected the wt-iASPP/mut-iASPP vectors and miR-506 NC/miR-506 mimics into SKOV3 cells. The luciferase activity of the iASPP 3′-UTR luciferase-reporter vector was significantly reduced in miR-506 mimic-transfected cells compared to control groups (). Moreover, miR-506-mediated repression of iASPP 3′-UTR luciferase-reporter activity was abolished by mutation of the putative miR-506-binding site in the iASPP 3′-UTR (). Next, results from Western blot showed that the expression of iASPP was significantly downregulated by miR-506 overexpression in both the SKOV3 and A2780 cell lines (). Moreover, expression levels of iASPP were significantly upregulated in tumor tissues compared with adjacent normal tissues (). We examined the potential correlation among the RNA-expression levels of iASPP and miR-506, and an inverse correlation between iASPP and miR-506 expressions was observed ().
Figure 6 miR-506 regulated iASPP expression by direct targeting.
Abbreviations: miR, microRNA; wt, wild type; UTR, untranslated region; mut, mutant; NC, negative control.
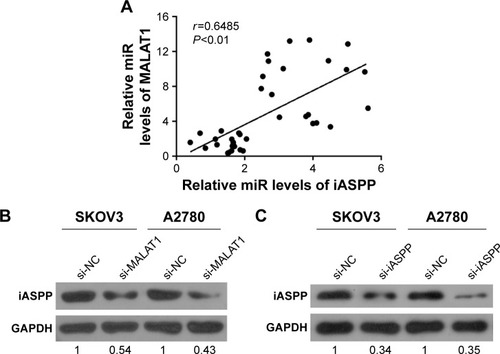
MALAT1 was positively correlated with iASPP
To investigate further the role and correlation of MALAT1 and iASPP in regulating human ovarian cancer proliferation, we determined the potential correlation between the RNA-expression levels of MALAT1 and iASPP. A positive correlation between the expression levels of MALAT1 and iASPP was observed (). Western blot results showed that protein contents of iASPP in the SKOV3 and A2780 cell lines were significantly decreased in response to MALAT1 knockdown by si-MALAT1 or iASPP inhibition by si-iASPP ().
Figure 7 MALAT1 was positively correlated with iASPP.
Abbreviations: si, small interfering; NC, negative control.
iASPP promoted ovarian cancer-cell growth
To investigate the effect of iASPP on ovarian cancer-cell growth, we determined ovarian cancer-cell growth in response to iASPP knockdown. According to MTT-assay results, ovarian cancer-cell growth was attenuated in response to iASPP inhibition by si-iASPP (). Moreover, BrdU results showed that DNA synthesis of both the SKOV3 and A2780 cell lines was reduced after iASPP inhibition ().
Figure 8 iASPP promoted ovarian cancer-cell growth.
Abbreviations: si, small interfering; NC, negative control; OD, optical density.
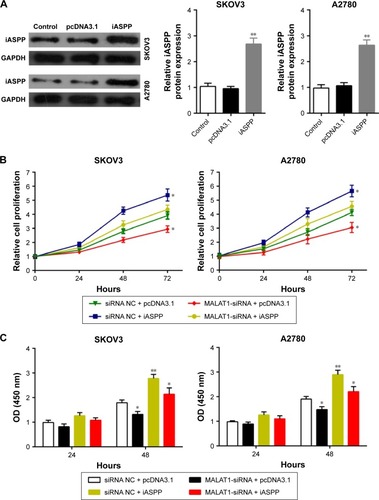
MALAT1 regulated ovarian cancer-cell growth through iASPP
Since we had demonstrated that MALAT1 was positively correlated with iASPP and that iASPP promoted ovarian cancer-cell growth, we further investigated whether MALAT1 could regulate MALAT1 through iASPP. Transfection of plasmid cDNA (pcDNA)-3.1/iASPP into SKOV3 and A2780 cell lines was generated to achieve iASPP overexpression. Transfection efficiency was verified using Western blot assay (). Then, si-NC/si-MALAT1 and pcDNA3.1/iASPP were cotransfected into the SKOV3 and A2780 cell lines. MTT and BrdU assays were used to determine cell growth. Results showed that the growth of both the SKOV3 and A2780 cell lines was significantly reduced in response to MALAT1 knockdown by si-MALAT1, while it was promoted in response to iASPP overexpression; the inhibitory effect of si-MALAT1 on SKOV3 and A2780 cell growth was partly restored by iASPP overexpression (). Similar results were observed in BrdU assay: the DNA synthesis of both the SKOV3 and A2780 cell lines was significantly reduced in response to MALAT1 knockdown by si-MALAT1, while it was promoted in response to iASPP overexpression; the inhibitory effect of si-MALAT1 on SKOV3 and A2780 cell DNA synthesis was partly restored by iASPP overexpression (). In sum, MALAT1 regulated ovarian cancer-cell growth through iASPP.
Figure 9 MALAT1 regulated ovarian cancer cell growth through iASPP.
Abbreviations: pcDNA, plasmid complementary DNA; si, small interfering; NC, negative control; OD, optical density.
Discussion
LncRNA-MALAT1 has been reported to be upregulated in many cancers.Citation16 Initially, we looked at MALAT1 miRNA expression in ovarian cancer tissues and cell lines compared with adjacent tissues and a normal cell line and found that MALAT1 expression was significantly higher in ovarian cancer tissues and cell lines. After siRNA knockdown of MALAT1, cell proliferation and DNA synthesis were significantly decreased in ovarian cancer-cell lines. These results suggested that MALAT1 functions as an oncogene in ovarian cancer, consistent with results from Hirata et al, where MALAT1 promoted human renal cancer-cell growth.Citation17 The molecular mechanism by which MALAT1 promotes cancer-cell proliferation varies by cancer species. MALAT1 may promote colorectal cancer-tumor development via its target protein AKAP9.Citation18 In the context of kidney clear-cell carcinoma, MALAT1 promote kidney clear-cell carcinoma proliferation and metastasis through sponging miR-200s in vitro and in vivo.Citation19 miR-200s can partly reverse MALAT1’s stimulation on proliferation and metastasis in kidney clear-cell carcinoma.Citation19 Moreover, a significant negative correlation exists between miR-101 or miR-217 and MALAT1 in esophageal squamous cell carcinoma, and silencing of MALAT1 inhibits proliferation, migration, and invasion of esophageal squamous cell carcinoma cells.Citation20 These studies suggest this lncRNA–miRNA interaction plays an important role in cancer progression through regulation of cancer growth and metastasis.
In the present study, we revealed the regulation of miR-506 by MALAT1 for the first time. Knockdown of MALAT1 upregulated miR-506, while forced miR-506 over-expression inhibited the expression of MALAT1. miR-506 has been regarded as a tumor-suppressive miRNA, and has been reported to be involved in hepatocellular carcinoma, melanoma, and glioblastoma.Citation14,Citation15,Citation21,Citation22 Studies have revealed that miRNAs, miR-506 included, mapped to the 14q32 locus regulate proliferation, apoptosis, migration, and invasion in cancer cells.Citation21,Citation23 In the presents study, we revealed that miR-506 regulates ovarian cancer growth and DNA synthesis by targeting iASPP. Since we have also demonstrated that MALAT1 downregulates miR-506 by direct targeting in ovarian cancer cell lines, as exhibited by real-time PCR assays and luciferase-reporter gene assays, we assumed that MALAT1 may regulate iASPP through miR-506.
To detect the correlation of MALAT1 and iASPP in ovarian cancer-cell growth regulation, siRNA has been transfected into ovarian cancer cell lines to knock down MALAT1 or iASPP. As expected, iASPP was downregulated by both MALAT1 knockdown and si-iASPP. In addition, the proliferation and DNA synthesis of ovarian cancer-cell lines were reduced after iASPP inhibition, consistent with Yang et al, where miR-205 regulated cell proliferation, invasion, and migration by targeting TGFα in osteosarcoma, and enforced expression of TGFα sufficiently restored the effects of miR-205 on cell proliferation, invasion, and migration.Citation24 Moreover, iASPP ectopic overexpression was achieved to investigate whether it could restore the effect of MALAT1 on ovarian cancer-cell growth. As exhibited by MTT and BrdU assays, the growth and DNA synthesis of both the SKOV3 and A2780 cell lines were significantly reduced in response to MALAT1 knockdown by si-MALAT1, while they were promoted in response to iASPP overexpression; the inhibitory effect of si-MALAT1 on SKOV3 and A2780 cell growth and DNA synthesis was partly restored by iASPP overexpression. Taken together, MALAT1 regulates ovarian cancer-cell proliferation and DNA synthesis through the miR-506–iASPP axis. In conclusion, MALAT1/miR-506/iASPP axis plays a key role in ovarian cancer growth, and provides a potential therapeutic application in ovarian cancer patients.
Acknowledgments
This work was supported by grant 2013-163 from the Key Laboratory of Malignant Tumor Molecular Mechanism and Translational Medicine of Guangzhou Bureau of Science and Information Technology and grant KLB09001 from the Key Laboratory of Malignant Tumor Gene Regulation and Target Therapy of Guangdong Higher Education Institutes.
Disclosure
The authors report no conflicts of interest in this work.
References
- DjebaliSDavisCAMerkelALandscape of transcription in human cellsNature2012489741410110822955620
- Martens-UzunovaESBöttcherRCroceCMJensterGVisakorpiTCalinGALong noncoding RNA in prostate, bladder, and kidney cancerEur Urol20146561140115124373479
- PontingCPOliverPLReikWEvolution and functions of long noncoding RNAsCell2009136462964119239885
- ZhouSWangJZhangZAn emerging understanding of long noncoding RNAs in kidney cancerJ Cancer Res Clin Oncol2014140121989199524816785
- WangBSuYYangQOverexpression of long non-coding RNA HOTAIR promotes tumor growth and metastasis in human osteosarcomaMol Cells201538543244025728753
- RuanWWangPFengSXueYLiYLong non-coding RNA small nucleolar RNA host gene 12 (SNHG12) promotes cell proliferation and migration by upregulating angiomotin gene expression in human osteosarcoma cellsTumour Biol20163734065407326486328
- ShiXSunMLiuHYaoYSongYLong non-coding RNAs: a new frontier in the study of human diseasesCancer Lett2013339215916623791884
- CaiXLiuYYangWLong noncoding RNA MALAT1 as a potential therapeutic target in osteosarcomaJ Orthop Res201634693294126575981
- DongYLiangGYuanBYangCGaoRZhouXMALAT1 promotes the proliferation and metastasis of osteosarcoma cells by activating the PI3K/Akt pathwayTumour Biol20153631477148625431257
- LiYAhmadASarkarFHASPP and iASPP: Implication in cancer development and progressionCell Mol Biol (Noisy-le-grand)20156162826518890
- JiangLSiuMKWongOGiASPP and chemoresistance in ovarian cancers: effects on paclitaxel-mediated mitotic catastropheClin Cancer Res201117216924693321926165
- JinYPengDShenYMicroRNA-376c inhibits cell proliferation and invasion in osteosarcoma by targeting to transforming growth factor-alphaDNA Cell Biol201332630230923631646
- DuanZChoyEHarmonDMicroRNA-199a-3p is downregulated in human osteosarcoma and regulates cell proliferation and migrationMol Cancer Ther20111081337134521666078
- ZhengYYinLChenHmiR-376a suppresses proliferation and induces apoptosis in hepatocellular carcinomaFEBS Lett2012586162396240322684007
- FellenbergJSährHKunzPRestoration of miR-127-3p and miR-376a-3p counteracts the neoplastic phenotype of giant cell tumor of bone derived stromal cells by targeting COA1, GLE1 and PDIA6Cancer Lett2016371113414126655997
- GutschnerTHämmerleMDiederichsSMALAT1: a paradigm for long noncoding RNA function in cancerJ Mol Med (Berl)201391779180123529762
- HirataHHinodaYShahryariVLong noncoding RNA MALAT1 promotes aggressive renal cell carcinoma through Ezh2 and interacts with miR-205Cancer Res20157571322133125600645
- YangMHHuZYXuCMALAT1 promotes colorectal cancer cell proliferation/migration/invasion via PRKA kinase anchor protein 9Biochim Biophys Acta20151852116617425446987
- XiaoHTangKLiuPLncRNA MALAT1 functions as a competing endogenous RNA to regulate ZEB2 expression by sponging miR-200s in clear cell kidney carcinomaOncotarget2015635380053801526461224
- WangXLiMWangZSilencing of long noncoding RNA MALAT1 by miR-101 and miR-217 inhibits proliferation, migration, and invasion of esophageal squamous cell carcinoma cellsJ Biol Chem201529073925393525538231
- ZehaviLAvrahamRBarzilaiASilencing of a large microRNA cluster on human chromosome 14q32 in melanoma: biological effects of mir-376a and mir-376c on insulin growth factor 1 receptorMol Cancer2012114422747855
- ChoudhuryYTayFCLamDHAttenuated adenosine-to-inosine editing of microRNA-376a* promotes invasiveness of glioblastoma cellsJ Clin Invest2012122114059407623093778
- FormosaAMarkertEKLenaAMMicroRNAs, miR-154, miR-299-5p, miR-376a, miR-376c, miR-377, miR-381, miR-487b, miR-485-3p, miR-495 and miR-654-3p, mapped to the 14q32.31 locus, regulate proliferation, apoptosis, migration and invasion in metastatic prostate cancer cellsOncogene201433445173518224166498
- YangGZhangPLvALiuYWangGMiR-205 functions as a tumor suppressor via targeting TGF-α in osteosarcomaExp Mol Pathol2016100116016626708425