
Abstract
Endometrial carcinoma (EC) is the most common gynecologic malignancy, but the molecular events involved in the development and progression of EC remain unclear. This study aimed to investigate the role of DNA methyltransferase 1 (DNMT1), a member of DNA methyltransferases, in EC. AN3CA cells were transfected with DNMT1 siRNA. The proliferation, cell cycle, and apoptosis of AN3CA cells were evaluated by Cell Counting Kit-8 (CCK-8) assay and flow cytometry. The expression of related genes was detected by polymerase chain reaction and Western blot analysis. Knockdown of DNMT1 inhibited the proliferation, induced apoptosis, and G0/G1 phase arrest of AN3CA cells. Furthermore, knockdown of DNMT1 upregulated the expression of nuclear factor kappa-B-inhibitor alpha (NF-κBIA) and Bax and downregulated the expression of Bcl-2 and CCND1/2 in AN3CA cells. In conclusion, this study provides the first evidence that knockdown of DNMT1 affects the expression of cell cycle- and apoptosis-associated proteins in EC cells, suggesting the potential of DNMT1 in EC therapy.
Introduction
Endometrial carcinoma (EC) is one of the most common malignancies of the female genital tract. EC is the second most common form of gynecologic malignancy, with >189,000 new cases and 45,000 deaths worldwide each year.Citation1,Citation2 EC patients with advanced or recurrent tumor, regardless of histological subtype, have a poor prognosis. Radiation or hormone therapy could be used as an adjuvant treatment for the majority of women with advanced or recurrent EC.
It is well known that mutation or deletion of coding regions is responsible for gene inactivation. However, a new molecular mechanism of gene silencing has been discovered.Citation3 DNA promoter hypermethylation is a key epigenetic mechanism for the silencing of many genes, including those involved in cell cycle regulation, DNA repair, and apoptosis.Citation4 Hypermethylation of CpG islands may inhibit transcription by recruiting the methyl-CpG-binding domain proteinsCitation5 or by interfering with the recruitment and function of basal transcription factors or transcriptional coactivators.Citation6 DNA methylation is modulated by DNA methyltransferases (DNMTs).Citation7 The human genome includes four genes that code DNMTs (DNMT1, DNMT2, DNMT3A, and DNMT3B) with DNMT1 and DNMT3B being the more potent.Citation8 DNMT1 is responsible for precise duplicating and maintaining the preexisting DNA methylation patterns after replication. DNMT1 could inhibit the transcription of tumor suppressor genes and facilitate tumorigenesis. Meanwhile, inhibition of DNMT1 activity could reduce the hypermethylation of repressive genes and reverse phenotypes of malignant tumor.Citation9 Global DNA hypomethylation is associated with chromosomal instability and is an early event in the neoplastic progression of most human and animal cancers.Citation10
DNMT1 silencing has been shown to inhibit cell proliferation and alter cell cycle progression through cell cycle arrest and apoptosis in cervical cancer.Citation11 Apoptosis is a programmed cell death morphologically characterized by cell shrinkage, chromatin condensation, internucleosomal DNA fragmentation, and the formation of apoptotic bodies. Moreover, cell cycle progression and apoptosis are well regulated by a set of intracellular signal transduction pathways.Citation12 DNMT1 inhibited the transcription of cell proliferation-related tumor suppressors, such as p27 and Bax, which are implicated in the development of several types of cancers.Citation13,Citation14 Several signals from the tumor microenvironment have been shown to trigger cell apoptosis processes via specific pathways involving Bcl-2, Cyclin-D1, and nuclear factor kappa-B (NF-κB), which are important regulators of cell fate in cancer cells.Citation15
In this study, we aimed to investigate the role of DNMT1 in EC. We demonstrated that knockdown of DNMT1 inhibited cell proliferation and induced cell apoptosis by modulating the expression of Bcl-2, Bax, NF-κB-inhibitor alpha (NF-κBIA), and Cyclin (CCN)D1/2. These results suggest that DNMT1 plays an important role in the etiology and progression of EC.
Materials and methods
Cell culture and transfection
The human EC cell lines HEC-1B, Kle, Ishikawa, and AN3CA were obtained from the Chinese Academy of Sciences Committee Type Culture Collection (Shanghai, People’s Republic of China). Cells were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM)/F12 (11030; Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS; 16000-44; Gibco) in a humidified atmosphere containing 5% CO2 at 37°C. DNMT1 siRNA and its control siRNA negative control (NC) were purchased from Sangon Technologies (Shanghai, People’s Republic of China) and transfected into cells using Lipofectamine 2000 (Thermo Fisher Scientific) according to the manufacturer’s protocol.
Quantitative reverse transcription polymerase chain reaction (RT-PCR)
Total RNA was extracted from the cells using TRIzol reagent (Thermo Fisher Scientific) and reverse-transcribed into cDNA using the one-step PrimeScript RT Reagent Kit (TaKaRa Bio Inc., Kusatsu, Japan). cDNA was then used for real-time quantitative polymerase chain reaction (qPCR) using SYBR Premix Ex Taq on an Eppendorf RealPlex Mastercycler. The primers used were as follows: NF-κBIA forward 5′-CTCCGAGACTTTCGAGGAAATAC-3′, reverse 5′-GCCATTGTAGTTGGTAGCCTTCA-3′; β-actin forward 5′-TCATGGGTGTGAACCATGTGAA-3′, reverse 5′-GGCATGGACTGTGGTCATGAG-3′; p65 forward 5′-CCTGGAGCAGGCTATCAGTC-3′, reverse 5′-ATCTTGAGCTCGGCAGTGTT-3′; Bcl-2 forward 5′-CGCGACTCCTGATTCATT-3′, reverse 5′-TGCATTCTTGGAGGG-3′; Bax forward 5′-CCAAGAAGCTGAGCGAGTGTC-3′, reverse 5′-TGAGGACTCCAGCCACAAAGA-3′; CCND1 forward 5′-ATGTTCGTGGCCTCTAAGATGA-3′, reverse 5′-CAGGTTCCACTTGAGCTTGTTC-3′; CCND2 forward 5′-AAGAATTCCTCCTCAATAGCCTGCAGCAGTA-3′, and reverse 5′-GCGGGATATCGACCTGTGAGAATTCGAT-3′. Relative mRNA levels were calculated using the 2(−ΔCt) method, and data were obtained in triplicate from three independent experiments.
Western blot analysis
Cells were washed with ice-cold phosphate-buffered saline (PBS) and lysed in ice-cold RIPA Lysis Buffer (Thermo Fisher Scientific) on ice for 30 min. Total protein was measured using the Bio-Rad protein assay reagent according to the manufacturer’s protocol. Proteins (20 mg) were separated on 10% polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes. The membranes were blocked with 5% bovine serum albumin (BSA)/PBS for 1 h and then incubated overnight at 4°C with primary antibodies against DNMT1 (1:1,000 dilution; Abcam, Cambridge, UK), p65 (1:500 dilution; Sangon Technologies), NF-κBIA (1:500 dilution; Sangon Technologies), Caspase-2 ((1:1,000 dilution; Sangon Technologies), Bcl-2 (1:1,000 dilution; Sangon Technologies), Bax (1:500 dilution; Sangon Technologies), CCND1 (1:1,000 dilution; Sangon Technologies), CCND2 (1:1,000 dilution; Sangon Technologies), and GAPDH (1:1,000 dilution; Cell Signaling Technology, Danvers, MA, USA). The membranes were washed three times with PBS and then incubated with peroxidase-linked secondary antibody (1:1,000 dilution) for 1 h at room temperature. The signals were developed using an electro-chemi-luminescence kit (Pierce™, Thermo Fisher Scientific) and analyzed using the TotalLab software.
Cell proliferation assay
Cell Counting Kit-8 (CCK-8; Beyotime Institute of Biotechnology) was used for the detection of cell proliferation. In 96-well plates, 103 cells/well were incubated in DMEM for 48 h prior to the addition of 10 µL of CCK-8 solution. The mixture was incubated at 37°C for 4 h, and then, the absorbance in each well was measured using an enzyme-linked immunosorbent assay reader (Bio-Rad Laboratories, Hercules, CA, USA) at 450 nm.
Cell cycle analysis
Cells were seeded at a density of 2×105 cells/mL in six-well plates. After overnight incubation, the cells were exposed to 100 nM of DNMT1 siRNA for 48 h. The cells were fixed in 70% ethanol for 16 h at 4°C, and the fixed cells were washed with PBS and resuspended in 1 mL of propidium iodide (PI) staining solution for incubation at 37°C for 15 min. The stained cells were analyzed to determine the fraction of cell population in various cell cycle stages using a flow cytometer (BD FACSCanto II; BD Biosciences, Franklin Lakes, NJ, USA).
Apoptosis analysis
Cells were seeded at a density of 2×105 cells/mL in six-well plates. After overnight incubation, the cells were exposed to 100 nM of DNMT1 siRNA for 48 h. The cells were fixed in 70% ethanol for 16 h at 4°C, and the fixed cells were washed with PBS and subjected to Annexin V-fluorescein isothiocyanate (FITC)/PI staining using Annexin V-FITC/PI Apoptosis Detection Kit (Sangon Technologies). The stained cells were analyzed to determine the fraction of cell population in various cell cycle stages using a flow cytometer (BD FACSCanto II).
Statistical analysis
Data were shown as the mean ± SD, and the differences were evaluated using one-way analysis of variance (ANOVA) for three-group comparisons and Student’s t-tests for two-group comparisons. All statistical analyses were performed using the SPSS 13.0 software package. P<0.05 was considered to be statistically significant.
Results
DNMT1 expression is high in poorly differentiated EC cell lines
In four EC cell lines, DNMT1 mRNA and protein expression levels were higher in poorly differentiated EC cell lines (AN3CA and HEC-1B) than in well-differentiated EC cell lines (Ishikawa and Kle) (). We selected AN3CA and HEC-1B cell lines for following experiments. Compared to siRNA NC-transfected cells, the mRNA level of DNMT1 in DNMT1 siRNA-transfected cells was significantly decreased (P<0.01) (). Moreover, the protein level of DNMT1 was significantly decreased in DNMT1 siRNA-transfected cells (). These data indicate that DNMT1 siRNA could effectively knockdown the expression of DNMT1 in EC cells.
Figure 1 Differential DNMT1 expression in endometrial cancer cell lines.
Abbreviations: EC, endometrial carcinoma; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
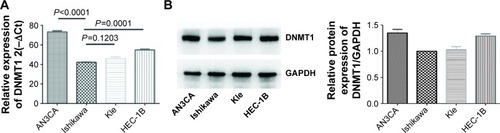
Figure 2 DNMT1 knockdown inhibits the proliferation of EC cells.
Abbreviations: CCK-8, Cell Counting Kit-8; EC, endometrial carcinoma; Con, control; NC, negative control; OD, optical density; ISK, Ishikawa; PCR, polymerase chain reaction; qPCR, real-time quantitative PCR.
Knockdown of DNMT1 inhibits the proliferation of EC cells
To understand the role of DNMT1 in EC cells, we examined the effect of DNMT1 knockdown on EC cell proliferation. CCK-8 assay showed that cell proliferation was significantly inhibited in AN3CA and HEC-1B cells transfected with DNMT1 siRNA compared to the cells transfected with siRNA NC (). These data suggest that DNMT1 promotes the proliferation of EC cells.
Knockdown of DNMT1 induces cell cycle arrest at G0/G1 phase in EC cells
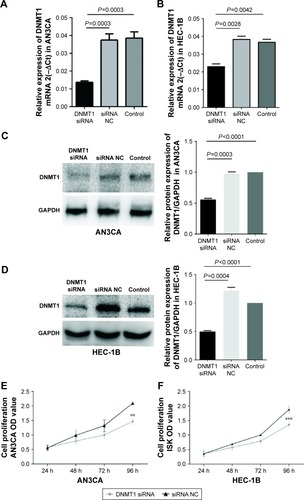
To explore the possible mechanisms by which DNMT1 promotes EC cell proliferation, the effect of DNMT1 silencing on cell cycle progression of AN3CA cells was analyzed by flow cytometry. The results showed that knockdown of DNMT1 in AN3CA cells induced an increase in the percentage of cells in G0/G1 phase (from 82.07%±0.80% in the siRNA NC group to 85.77%±0.29% in the DNMT1 siRNA group, P<0.05), parallel with a decrease in the percentage of cells in S phase (from 17.17%±0.59% in siRNA NC group to 13.87%±0.40% in DNMT1 siRNA group, P<0.05) (). These results suggest that DNMT1 knockdown inhibits EC cell proliferation through inducing G0/G1 cell cycle arrest.
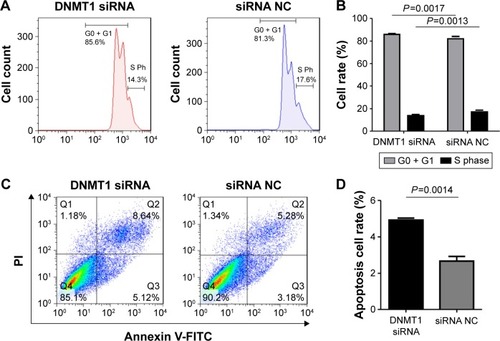
Figure 3 DNMT1 knockdown induces cell cycle arrest and apoptosis of AN3CA cells.
Abbreviations: PI, propidium iodide; FITC, fluorescein isothiocyanate; NC, negative control; Ph, phase.
Knockdown of DNMT1 induces apoptosis in EC cells
Furthermore, we examined the effect of DNMT1 silencing on the apoptosis of AN3CA cells. Flow cytometry of AN3CA cells stained with Annexin-V-FITC showed that the proportion of early apoptotic cells was increased in AN3CA cells transfected with DNMT1 siRNA (4.93%±0.20%) compared to that in cells transfected with siRNA NC (2.66%±0.45%) (). Our results suggest that DNMT1 inhibits the apoptosis of EC cells.
Knockdown of DNMT1 alters the expression of cell cycle- and apoptosis-related proteins in EC cells
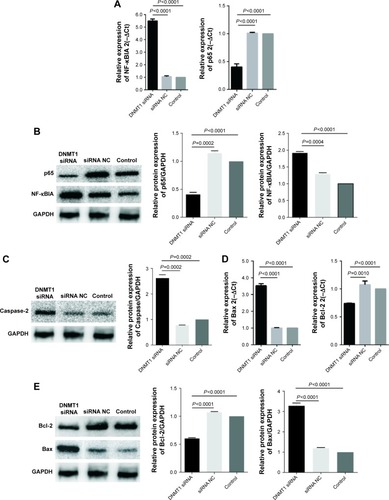
To understand the molecular mechanism by which DNMT1 modulates EC cell cycle and apoptosis, we detected the expression of a variety of proteins involved in cell cycle and apoptosis. qPCR and Western blot analyses showed that NF-κBIA and Caspase-2 were upregulated and p65 was downregulated in DNMT1 siRNA-transfected AN3CA cells (). Moreover, Bax was upregulated and Bcl-2 was downregulated in AN3CA cells after the knockdown of DNMT1 (). These results reveal that DNMT1 regulates these apoptosis-related genes to promote the apoptosis of EC cells.
Figure 4 DNMT1 knockdown alters the expression of apoptosis-related proteins.
Abbreviations: NF-κBIA, nuclear factor kappa-B-inhibitor alpha; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; NC, negative control; PCR, polymerase chain reaction; qPCR, real-time quantitative PCR.
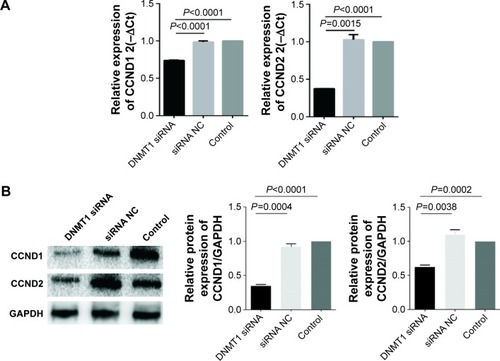
Furthermore, we found that mRNA and protein levels of CCND1 and CCND2 were significantly decreased in DNMT1 siRNA-transfected AN3CA cells compared to that in siRNA NC-transfected cells (). These data indicate that DNMT1 upregulates CCND1 and CCND2 to enhance the proliferation of EC cells.
Figure 5 DNMT1 knockdown downregulates CCNDs.
Discussion
In recent years, numerous studies have shown an association between cell cycle regulation and cancer and inhibition of cell cycle has become an appreciated target for the management of cancer.Citation16 Our study showed that knockdown of DNMT1 exerted strong growth inhibitory effects on human EC cells by the perturbation of G1–S phase transition. CCNDs are crucially involved in G1–S phase transition of cell cycle progression, and their overexpression is associated with various cancers.Citation17,Citation18 Indeed, we found that DNMT1 knockdown led to a decreased expression of CCND1 and CCND2 in EC cells, which may explain how DNMT1 promotes EC cell proliferation. A study demonstrated that hyper-methylation of CCND was associated with the loss of mRNA expression and tumor development in several kinds of cancer, such as prostate cancer.Citation19 A recent study showed that DNMT1 protein interacted with β catenin and regulated Wnt signaling in colorectal cancer cells.Citation20 CCND1 and CCND2 are the downstream targets of Wnt/β catenin signaling and the important effectors to enhance cell cycle progression. Therefore, it is possible that the knockdown of DNMT1 decreased the stability and activity of β catenin and downregulated the expression of CCND1 and CCND2, leading to cell cycle arrest. Further exploration of the mechanism of action of DNMT1 in regulating cell cycle progression and apoptosis will deepen our understanding of the role of DNMT1 in tumorigenesis.
NF-κB family of transcription factors include five members, such as p50, p52, RelA (also known as p65), RelB, and c-Rel, which regulate the expression of various genes involved in inflammatory, antiapoptotic, and immune responses. The activation of NF-κB in cancer cells promotes cancer development and progression, and NF-κB has been thought to be a target for cancer therapy. NF-κB promotes cell survival by inhibiting apoptosis.Citation21 Our results demonstrate a decreased expression of p65 in EC cells with subsequent increased expression of NF-κBIA after the knockdown of DNMT1. We postulate that the loss of DNMT1-mediated downregulation of NF-κB activity could lead to the inhibition of cell growth and induction of apoptosis. We confirmed that DNMT1 knockdown significantly upregulated the expression of Caspase-2 in EC cells, which may contribute to apoptosis.
Members of the Bcl-2 family of proteins are the critical regulators of apoptotic pathway.Citation22 Bcl-2 is an upstream effector in the apoptotic pathway and is a potent suppressor of apoptosis.Citation23 Bcl-2 is found at inappropriately high levels in more than half of all human tumors. Bcl-2 has been shown to form a heterodimer with the proapoptotic member Bax and neutralize its proapoptotic effects. Therefore, the ratio of Bax/Bcl-2 is a crucial factor in determining whether cells will undergo apoptosis under conditions that promote cell death.Citation24 In our study, a decrease in Bcl-2 protein expression was observed in DNMT1 siRNA-transfected EC cells, while the protein expression of Bax was upregulated; hence, the ratio of Bax to Bcl-2 was altered in favor of apoptosis. Our results suggest that the upregulation of Bax and the downregulation of Bcl-2 may be another molecular mechanism through which knockdown of DNMT1 induces apoptosis.
Consistent with our results in EC cells with DNMT1 silencing, we obtained similar results in AN3CA cells treated with 5-aza-dC, the DNA methyl-transferase inhibitor.Citation25 Notably, a previous study reported that DNMT1 was overexpressed in endometrial tumor tissues and DNMT1 expression level was correlated with tumor progression.Citation26 These data suggest the potential of DNMT1 as a new target for EC treatment. However, in vivo studies on animal models of EC are needed before clinical trials on DNMT1 inhibitors.
Conclusion
DNMT1 is aberrantly overexpressed in aggressive and poorly differentiated EC cells. Knockdown of DNMT1 significantly inhibited EC cell proliferation, mostly by inducing cell cycle arrest at G0/G1 phase and inducing apoptosis. Knockdown of DNMT1 could downregulate CCNDs and NF-κB, decrease Bcl-2/Bax ratio, and upregulate Caspase-2. Our study suggests that DNMT1 is a potential therapeutic target for EC.
Acknowledgments
This study was sponsored by Natural Science Foundation of Shanghai (No. 16ZR1427100), the National Natural Science Foundation of China (No. 81602280 and No. 81602281).
Disclosure
The authors report no conflicts of interest in this work.
References
- SiegelRLMillerKDJemalACancer statistics, 2015CA Cancer J Clin201565152925559415
- TsikourasPBouchlariotouSVrachnisNEndometrial cancer: molecular and therapeutic aspectsEur J Obstet Gynecol Reprod Biol201316911923433742
- ChometPSCytosine methylation in gene-silencing mechanismsCurr Opin Cell Biol1991334384431892655
- JonesPABaylinSBThe fundamental role of epigenetic events in cancerNat Rev Genet20023641542812042769
- BogdanovićOVeenstraGJCDNA methylation and methyl-CpG binding proteins: developmental requirements and functionChromosoma2009118554956519506892
- BallestarEEstellerMMethyl-CpG-binding proteins in cancer: blaming the DNA methylation messengerBiochem Cell Biol200583337438415959563
- OkanoMBellDWHaberDALiEDNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian developmentCell199999324725710555141
- OkanoMXieSLiECloning and characterization of a family of novel mammalian DNA (cytosine-5) methyltransferasesNat Genet19981932192209662389
- BestorTHThe DNA methyltransferases of mammalsHum Mol Genet20009162395240211005794
- FigueiredoJCGrauMVWallaceKGlobal DNA hypomethylation (LINE-1) in the normal colon and lifestyle characteristics, dietary and genetic factorsCancer Epidemiol Biomarkers Prev20091841041104919336559
- ZhangYChenF-QSunY-HZhouS-YLiT-YChenREffects of DNMT1 silencing on malignant phenotype and methylated gene expression in cervical cancer cellsJ Exp Clin Cancer Res2011309821999220
- EomH-JChoiJp38 MAPK activation, DNA damage, cell cycle arrest and apoptosis as mechanisms of toxicity of silver nanoparticles in Jurkat T cellsEnviron Sci Technol201044218337834220932003
- PengD-FKanaiYSawadaMDNA methylation of multiple tumor-related genes in association with overexpression of DNA methyltransferase 1 (DNMT1) during multistage carcinogenesis of the pancreasCarcinogenesis20062761160116816537562
- SowińskaAJagodzinskiPPRNA interference-mediated knockdown of DNMT1 and DNMT3B induces CXCL12 expression in MCF-7 breast cancer and AsPC1 pancreatic carcinoma cell linesCancer Lett2007255115315917532557
- GuptaSAfaqFMukhtarHInvolvement of nuclear factor-kappa B, Bax and Bcl-2 in induction of cell cycle arrest and apoptosis by apigenin in human prostate carcinoma cellsOncogene200221233727373812032841
- PorterTRRichardsFMHoulstonRSContribution of Cyclin D1 (CCND1) and E-cadherin (CDH1) polymorphisms to familial and sporadic colorectal cancerOncogene200221121928193311896626
- CasimiroMCDi SanteGJuXCyclin D1 promotes androgen-dependent DNA damage repair in prostate cancer cellsCancer Res201676232933826582866
- RahalOMPabonaJMPKellyTSuppression of Wnt1-induced mammary tumor growth and lower serum insulin in offspring exposed to maternal blueberry diet suggest early dietary influence on developmental programmingCarcinogenesis201334246447423144318
- HenriqueRCostaLVCerveiraNHypermethylation of Cyclin D2 is associated with loss of mRNA expression and tumor development in prostate cancerJ Mol Med2006841191191817016690
- SongJDuZRavaszMDongBWangZEwingRMA protein interaction between β-catenin and Dnmt1 regulates wnt signaling and DNA methylation in colorectal cancer cellsMol Cancer Res201513696998125753001
- BoursVBonizziGBentires-AljMNF-κB activation in response to toxical and therapeutical agents: role in inflammation and cancer treatmentToxicology20001531–3273811090945
- AdamsJMCorySThe Bcl-2 protein family: arbiters of cell survivalScience19982815381132213269735050
- LiuZDingYYeNWildCChenHZhouJDirect activation of bax protein for cancer therapyMed Res Rev201636231334126395559
- KroemerGThe proto-oncogene Bcl-2 and its role in regulating apoptosisNat Med1997366146209176486
- LiBLLuWLuCCpG island hypermethylation-associated silencing of microRNAs promotes human endometrial cancerCancer Cell Int20131314423680357
- IkedaSImuraJSuzukiKProtein expression, mRNA expression and gene amplification of DNA methyltransferase 1 in endometrial tumor tissuesMol Clin Oncol20131342342924649186