
Abstract
Background
The expression of the urokinase plasminogen activator receptor (uPAR), a glycosylphosphatidylinositol-anchored protein family member, and the activity of its ligand, urokinase-type plasminogen activator (uPA), have been associated with the invasive and metastatic potentials of a variety of human brain tumors through their regulation of extracellular matrix degradation. Domesticated dogs develop naturally occurring brain tumors that share many clinical, phenotypic, molecular, and genetic features with their human counterparts, which has prompted the use of the dogs with spontaneous brain tumors as models to expedite the translation of novel brain tumor therapeutics to humans. There is currently little known regarding the role of the uPA system in canine brain tumorigenesis. The objective of this study was to characterize the expression of uPAR and the activity of uPA in canine brain tumors as justification for the development of uPAR-targeted brain tumor therapeutics in dogs.
Methods
We investigated the expression of uPAR in 37 primary canine brain tumors using immunohistochemistry, Western blotting, real-time quantitative polymerase chain reaction analyses, and by the assay of the activity of uPA using casein–plasminogen zymography.
Results
Expression of uPAR was observed in multiple tumoral microenvironmental niches, including neoplastic cells, stroma, and the vasculature of canine brain tumors. Relative to normal brain tissues, uPAR protein and mRNA expression were significantly greater in canine meningiomas, gliomas, and choroid plexus tumors. Increased activity of uPA was documented in all tumor types.
Conclusions
uPAR is overexpressed and uPA activity increased in canine meningiomas, gliomas, and choroid plexus tumors. This study illustrates the potential of uPAR/uPA molecularly targeted approaches for canine brain tumor therapeutics and reinforces the translational significance of canines with spontaneous brain tumors as models for human disease.
Background
While the pathophysiology of tumor progression and metastasis is complex, a fundamental step in the process is the ability of neoplastic cells to initiate proteolysis.Citation1 Degradation of basement membranes (BM) and the extracellular matrix (ECM) is necessary for cancer cells to breach natural tissue barriers, migrate, invade, and induce microenvironmental remodeling and angiogenesis.Citation1–Citation3 A growing body of evidence has indicated that serine proteases and metalloproteinases are key extracellular proteolytic enzymes that are overexpressed in many solid tumors of several species, including cats, dogs, and humans.Citation3–Citation6
Activation of plasminogen to form plasmin is a central event in the regulation of extra- and pericellular proteolysis in both physiological and pathological states, as plasmin is a nonspecific serine protease capable of degrading most extracellular proteins.Citation3,Citation7 There are two types of plasminogen activators (PAs): tissue-type plasminogen activator (tPA) and urokinase-type plasminogen activator (uPA), both of which can catalyze the conversion of the inactive plasminogen zymogen to plasmin, as well as cause the activation of additional protease zymogens and other latent growth factors.Citation6,Citation7 The principal role of tPA involves the intravascular generation of plasmin for thrombolysis, whereas uPA is predominantly associated with degradation of the ECM.Citation7
The uPA system consists of the serine proteinases uPA and plasmin, two serpin plasminogen activator inhibitors (PAI; PAI-1 and PAI-2), the serpin plasmin inhibitor α2-anti-plasmin, and the cell surface uPA receptor (uPAR), a member of the glycosylphosphatidylinositol (GPI)-anchored protein family.Citation2,Citation3 Participation of uPAR in tumor invasion and metastasis involves the plasminogen activation dependent breakdown of the ECM and attachment of invasive cells to ECM components, which is mediated by the specific interaction of uPAR with its ligand, uPA. However, uPAR has also been implicated in tumor progression through mechanisms that are independent of plasminogen activation including extracellular signal transduction via interactions with low-density lipoprotein receptor-related protein, integrins, vascular endothelial growth factor, epidermal growth factor receptor, or platelet-derived growth factor receptor.Citation2,Citation8
Naturally occurring brain tumors develop in dogs and have been shown to share clinical, phenotypic, and molecular similarities with their human counterparts.Citation9,Citation10 This homology has driven interest in use of dogs with spontaneous brain tumors as models for the development and assessment of novel therapeutics.Citation9,Citation10 Although treatments targeting components of the uPA system have been investigated in human gliomas and meningiomas,Citation11–Citation13 little is currently known about the expression or activity of uPAR/uPA in canine brain tumors. We hypothesized that uPAR/uPA would be overexpressed and active in canine brain tumors when compared with normal brain tissues. This would justify further development and evaluation of uPAR/uPA targeted molecular therapies in dogs.
Materials and methods
Tissue sources
Brain tumor samples included surgical biopsy or resection specimens, necropsy samples, or archival paraffin-embedded specimens from clinical cases presented to the Veterinary Teaching Hospital of the Virginia-Maryland College of Veterinary Medicine. Surgical and necropsy specimens were snap frozen in liquid nitrogen or immersed in Allprotect reagent (Qiagen, Valencia, CA, USA), and stored at -80°C within 30 minutes of collection. Normal brain samples (meninges, cerebral cortex, and choroid plexus) were collected from necropsy and archival paraffin-embedded materials from dogs with no clinical, magnetic resonance imaging, or histopathological evidence of brain disease (n=5). Clients provided written consent for their dog’s tissues to be banked in a biospecimen repository and used for research purposes (IACUC Approval 13-153-CVM). Clinicopathological data were recorded from each canine subject including the breed, age, sex, tumor location, and analyses performed on each tumor (). All tumors were classified and graded by veterinary pathologists according to World Health Organization criteria by examination of hematoxylin and eosin (H&E) and glial fibrillary acidic protein, vimentin, or Olig-2 stained slides prepared from formalin-fixed, paraffin-embedded specimens, as previously reported.Citation9,Citation14,Citation15
Table 1 Clinicopathological characteristics of dogs with primary brain tumors and normal controls
uPAR immunohistochemistry
A 5-μm section was cut from each paraffin-embedded block, mounted on positively charged slides, placed in a drying oven at 40°C for 30 minutes, and then placed in a drying oven at 65°C for 30 minutes. Immunohistochemistry was performed using a procedure modified from a previously described protocol using an automated staining system (BenchMark XT; Ventana Medical Systems, Inc., Tucson, AZ, USA) and an alkaline phosphatase detection method (Enhanced Alkaline Phosphatase Detection Red Kit; Ventana).Citation16 Epitope retrieval was performed with Cell Conditioning 1 Solution (Ventana) at 96°C for 90 minutes, followed by rinsing in reaction buffer (Ventana). Following rinsing, slides were incubated with a murine anti-uPAR antibody (M7294, Clone; Dako, Santa Cruz, CA, USA; 1:25 dilution using antibody dilution buffer; Ventana) at 37°C for 2 hours, and then rinsed three times for 2 minutes each with reaction buffer. Slides were subjected to automated amplification (Amplification Kit; Ventana) for 16 minutes and endogenous biotin blocking (Blocking Kit; Ventana) for 8 minutes prior to the additional of the secondary biotinylated goat antimouse/antirabbit antibody for 30 minutes, and rinsed with reaction buffer. Slides were incubated with a streptavidin–alkaline phosphatase conjugate in Tris buffer with MgCl2 and ZnCl (Ventana) for 30 minutes. Slides were then developed with a naphthol substrate and fast-red chromogen (Fast Red; Ventana) for 15 minutes, counterstained with Richard-Allan Hematoxylin, and dried at room temperature. Negative controls were performed by substituting the primary antibody with antibody dilution buffer (Ventana). Canine tonsils and intestines were used as the positive control tissue.Citation16
uPAR Western blotting (WB)
Canine tumor lysates were prepared in an extraction buffer (150 mM NaCl, 20 mM Tris/HCl [pH 7.4], 0.1% sodium-dodecyl sulfate [SDS], 1% sodium deoxycholate, 1% Triton X-114 with proteinase inhibitory mixture [5 μg/mL aprotinin, 1 μg/mL leupeptin, 1 μg/mL trypsin inhibitor, and 1 mM phenylmethylsulfonyl fluoride]) and incubated for 5 minutes at 37°C. Extracts were then centrifuged at 2500× g for 5 minutes to separate a detergent phase that contains mainly hydrophobic membrane proteins including GPI-anchored uPAR. Human HL-60 leukemia cell extracts prepared in SDS–polyacrylamide gel electrophoresis (PAGE) buffer (Enzo, Farmingdale, NY, USA) were used as a WB-positive control. The total protein concentration was obtained using a bicinchoninic assay kit (BCA-1; Sigma–Aldrich, St Louis, MO, USA), and 20 μg of protein from these samples was separated under nonreducing conditions by 15% SDS–PAGE and transferred to nitrocellulose membranes (Bio-Rad, Hercules, CA, USA). After blocking with phosphate-buffered saline–Tween and 5% nonmilk fat at room temperature for 2 hours, the membrane was blotted with a monoclonal murine anti-uPAR antibody (SAB4200412, 1:500 dilution; Sigma, St Louis, MO, USA) for 1 hour at room temperature. According to the manufacturer, this antibody is cross-reactive with human and canine uPAR. The membrane was rinsed and incubated with goat antimouse horseradish peroxidase secondary antibody (1:1,000 dilution; BD Bioscience, San Jose, CA, USA) for 1 hour at room temperature and developed using an enhanced chemiluminescence detection kit (GE Healthcare Life Sciences, Piscataway, NJ, USA) to allow for relative quantification of uPAR. A rabbit anti-β-actin poly-clonal antibody (Biovision, Mountain View, CA, USA) was used as a protein loading control. Quantitative analyses were performed using a high-resolution digital imaging (Amersham 600; GE Healthcare) and dedicated chemiluminescent software system (Melanie, 2D Gel Package; GE Healthcare).
uPAR real-time quantitative polymerase chain reaction (RT-qPCR)
mRNA from canine tumors and brain tissues was isolated using an RNeasy kit (Qiagen) according to the manufacturer’s instructions, and all mRNA samples were stored at −80°C before analyses. An iTaq Universal one-step kit (Bio-Rad) was used to perform the RT-qPCR, with β-actin serving as an internal control. The primers for uPAR (NCBI-ID XM_014119899.1) were 5′-GCCTTACCGAGGTTGTGTGT-3′ (forward) and 5′-CATCCAGGCACTGTTCTTCA-3′ (reverse). The β-actin-specific primers were 5′-CTGGAACGGTGAAGGTGACA-3′ (forward) and 5′-GGGAGAGGACTGGGCCATT-3′ (reverse). The uPAR primers used in this study were created using the Primer3Plus platform.Citation17 The specificity of the uPAR primers and their corresponding amplicons were evaluated in silico using the BLAST online tool.Citation18 uPAR mRNA expression was calculated as the fold-change relative to the β-actin control using comparative CT methodology.Citation9
uPA activity: casein–plasminogen zymography
The activity of uPA in tissues was analyzed by casein–plasminogen zymography using a previously described method.Citation19 Tumor homogenates (10 μg protein/lane) were separated by electrophoresis in 10% polyacrylamide gel copolymerized with 1 mg/mL of casein (Sigma) and 13 μg/mL human plasminogen (Sigma) under nonreducing conditions. Following electrophoretic separation, the gels were washed twice in 2.5% Triton-X 100 for 30 minutes, incubated with reaction buffer (50 mM Tris–HCl, 5 mM CaCl2, and 0.02% NaN3, pH 8.0) at 37°C for 18 hours, and then stained with Coomassie blue (Sigma). Purified, high-molecular weight (50 kDa) canine uPA (DUPA; Molecular Innovations, Novi, MI, USA) and human tPA (ab92637; Abcam, Cambridge, UK) were used as positive controls, and negative controls consisted of samples loaded on gels devoid of plasminogen. Specific inhibition of uPA activity was performed by running parallel gels, which contained 2 mM amiloride (Sigma). Amiloride is a selective inhibitor of uPA but does not affect other PAs.Citation20
Statistical analysis of WB and RT-qPCR
To test the null hypothesis that tumor type does not have an effect on uPAR expression, data were modeled by use of a Kruskal–Wallis one-way analysis of variance. To test the null hypothesis that tumor grade does not have an effect on uPAR expression, Jonckheere trend tests were done to compare tumor grade variability within tumor types. When significant differences were identified, Mann–Whitney tests were used for pair-wise comparisons. A Bonferroni’s multiple comparison adjustment was used to confirm significant results. For all analyses, alpha was set at 0.05.
Results
Canine tissue samples
A total of 37 primary canine brain tumor samples were included in analyses of uPAR expression and uPA activity (). Of 37 samples, 21 were meningiomas, (Grade I, n=14; Grade II, n=7), 7 were astrocytomas (Grade II, n=2; Grade III, n=1; and Grade IV [glioblastoma; GBM], n=4), 6 were oligodendrogliomas (Grade II, n=3; Grade III, n=3), and 3 were choroid plexus tumors (Grade II, n=2; Grade III, n=1).
uPAR immunohistochemistry
In normal brain tissue controls (), uPAR immunoreactivity was observed in rare microglia () and vascular endothelium contained within the cerebral cortex, as well as consistently in the choroid plexus vascular endothelium (). No uPAR immunoreactivity was observed in normal meninges ().
Figure 1 Immunohistochemical evaluation of uPAR expression in control canine brain tissues (A–F) and brain tumors (G–R).
Abbreviations: GBM, glioblastoma; H&E, hematoxylin and eosin; NB, normal brain (cerebral cortex); NCP, normal choroid plexus; NM, normal meninges; uPAR, urokinase plasminogen activator receptor; CII, grade II choroid plexus tumor; OligoIII, grade III oligodendroglioma; MenII, grade II meningioma; MenI, grade I meningioma.
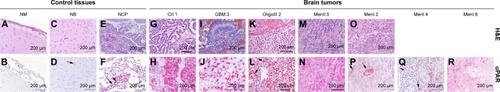
The neoplastic epithelium in each of the choroid plexus tumors () examined displayed diffuse intense cytoplasmic uPAR immunoreactivity. Immunoreactivity of supporting fibrovascular stromal cells was observed only in the choroid plexus carcinoma.
All astrocytomas (7/7) and oligodendrogliomas (6/6) were immunoreactive to uPAR (). Tumor cells in oligodendrogliomas and astrocytomas demonstrated cytoplasmic patterns of uPAR immunoreactivity. In all high-grade glioma samples, >30% of neoplastic cells were immunoreactive to uPAR (). The tumor vas-culature within both astrocytomas and oligodendrogliomas demonstrated regionally variable immunoreactivity to uPAR but was universally marked in hyperplastic tumor vasculature (). In the two necropsy-derived GBM specimens, the most intense staining of the neoplastic cells and tumor vasculature was observed adjacent to areas of tumor invasion of the neuropil (data not shown).
Among meningiomas (), 19 of 21 demonstrated immunoreactivity to uPAR, including 12 of 14 Grade I tumors and 7 of 7 Grade II tumors. In 6 of 14 Grade I meningiomas, uPAR immunoreactivity was predominantly localized to the endothelial cytoplasm within the tumor vasculature (), with <5% of neoplastic cells demonstrating uPAR immunoreactivity in these cases. In 6 of 14 Grade I and 7 of 7 Grade II meningiomas, mildly to markedly intense cytoplasmic immunoreactivity was observed in tumor, stromal, and vascular endothelial cells. In Grade I meningiomas, uPAR immunoreactivity in these tumor cellular populations was mild and multifocal in 4 of 6 cases (), whereas in Grade II meningiomas uPAR immunoreactivity was intense and diffuse in 4 of 7 cases ().
uPAR WB
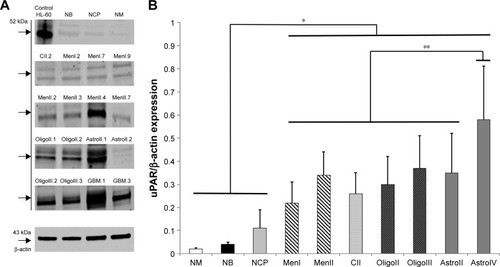
WB documented expression of uPAR protein in all control and tumor tissues examined. In the positive HL-60 control as well as in some of the canine normal brain and tumor samples, two distinct bands were detected (), one at a MW of 52 kDa and another at 46 kDa, corresponding to uPAR isoforms 1 and 3, respectively. Compared with normal brain tissues (), the expression of uPAR was significantly greater in all types of brain tumors, and uPAR expression in GBM was higher compared with all other tumor types and grades.
Figure 2 (A) Representative Western blots of uPAR protein in canine normal brain tissues and brain tumors. The numeral after each tumor type and grade indicates the specific sample depicted. (B) uPAR protein expression compared with β-actin concentrations in normal canine brain tissues and tumors.
Abbreviations: GBM, glioblastoma; NM, normal meninges; NB, normal brain (cerebral cortex); NCP, normal choroid plexus; uPAR, urokinase plasminogen activator receptor; MenI, grade I meningioma; MenII, grade II meningioma; CII, grade II choroid plexus tumor; OligoII, grade II oligodendroglioma; OligoIII, grade III oligodendroglioma; AstroII, grade II astrocytoma; AstroIV, grade IV astrocytoma.
uPAR RT-qPCR
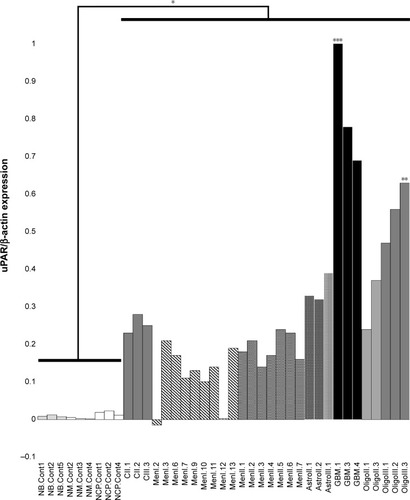
Patterns of uPAR gene expression were similar to what was observed in WB analyses, with uPAR expression in all types and grades of brain tumors being significantly increased compared with control brain tissues (P<0.002). The highest uPAR gene expression was seen in high-grade gliomas (). Expression of uPAR in astrocytomas and oligodendrogliomas was higher compared with meningiomas (P<0.02) and choroid plexus tumors (P<0.04). No significant differences in uPAR expression were observed between meningiomas and choroid plexus tumors (P=0.43), or between astrocytomas and oligodendrogliomas (P=0.22). Differences in uPAR expression among tumor grades were observed, with expression in GBM being significantly greater than in Grade II and III astrocytomas (combined, P=0.004), and expression in Grade III oligodendrogliomas was also greater than in Grade II oligodendrogliomas (P=0.009). No significant differences in uPAR gene expression were observed between grades of meningiomas (P=0.67).
Figure 3 Expression of urokinase plasminogen activator receptor (uPAR) mRNA in canine primary brain tumors as determined by real-time quantitative polymerase chain reaction.
Abbreviations: NB.Cont, normal brain control; MN.Cont, normal meninges control; NCP.Cont, normal choroid plexus control; CII, grade II choroid plexus tumor; CIII, grade III choroid plexus tumor; MenI, grade I meningioma; MenII, grade II meningioma; AstroII, grade II astrocytoma; AstroIII, grade III astrocytoma; GBM, glioblastoma; OligoII, grade II oligodendroglioma; OligoIII, grade III oligodendroglioma.
uPA activity: casein–plasminogen zymography
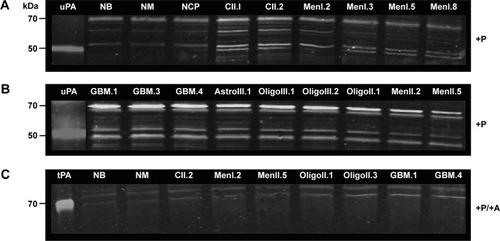
Casein–plasminogen zymography demonstrated lysis bands at molecular weights of 45–55 kDa, corresponding to uPA, and 65–70 kDa, identified as tPA, in all tissue samples (). In the presence of amiloride, inhibition of uPA lysis bands occurred at molecular weights of ~45–60 kDa, indicating specific uPA proteolytic activity in all tissue samples (). Qualitatively, uPA activity was detected in all tumor samples, with comparably little activity detected in normal brain tissues.
Figure 4 uPA activity in normal canine brain tissues and brain tumors as determined with casein–plasminogen zymography.
Abbreviations: NB, normal brain; NCP, normal choroid plexus; NM, normal meninges; +P, plasminogen-treated gel; MenI, grade I meningioma; MenII, grade II meningioma; CII, grade II choroid plexus tumor; OligoII, grade II oligodendroglioma; OligoIII, grade III oligodendroglioma; AstroII, grade II astrocytoma; AstroIII, grade III astrocytoma; GBM, glioblastoma; +P/+A, plasminogen and amiloride-treated gel; tPA, purified human tissue plasminogen activator control; uPA, urokinase-type plasminogen activator control.
Discussion
Meningiomas, gliomas, and choroid plexus tumors account for ~50%, 35%, and 10% of all canine primary brain tumors, respectively.Citation21–Citation23 Similarities in the clinical behavior, morphology, molecular signatures, and genetic aberrations have been observed between human and canine brain tumors, which has resulted in the emergence of the brain tumor bearing dog as a model for translational research.Citation9,Citation10,Citation24 While these tumors represent a phenotypically, clinically, and genomically heterogeneous group of cancers, a shared pathophysiological denominator that contributes to the challenges associated with their treatment is their ability to invade the brain parenchyma.Citation8 In this study, using several complementary methods, we have demonstrated that uPAR protein and mRNA are overexpressed and uPA activity is present in the most commonly encountered canine primary brain tumors.
Dysregulation of the uPAR interactome has been implicated as an important component of the pathogenesis of numerous human central nervous system disorders including Alzheimer’s disease, autism, brain tumors, and epilepsy.Citation24 In human primary brain tumors, uPAR participates in numerous plasminogen activation dependent cell surface oncogenic processes including cellular invasion, migration, and distant metastatic seeding that are mediated via degradation of the BM and ECM.Citation1,Citation13,Citation25 Through its interactions with integrins, uPAR also regulates cell-signaling mechanisms involved in tumor cell proliferation, chemotaxis, angiogenesis, and protection from apoptosis.Citation2,Citation25 For systemic malignancies, it has also recently been hypothesized that uPAR upregulation in circulating tumor cells may be the switch that triggers dormant tumor cells to proliferate and initiate the brain metastatic cascade.Citation26 Our demonstration of uPAR expression in the tumor, stromal, and endothelial cellular fractions of multiple tumor types also suggests that uPAR may play a fundamental role in canine brain tumorigenesis.
The malignant potential of human meningiomas and gliomas has been correlated with uPAR expression, which parallels our finding of significantly increased uPAR expression in canine high-grade gliomas compared with other tumor types and grades.Citation1,Citation8,Citation13
Previous studies have shown that robust uPAR expression occurs at the invasive edges of malignant gliomas.Citation25 In a limited number of cases, we observed intense uPAR immunoreactivity at the interface of GBM where it effaced the neuropil. The role of uPAR-mediated proteolytic cleavage in canine glioma invasion requires further investigation.
Investigations in human brain tumors have demonstrated that uPAR is a pharmacologically tractable molecular target for diagnostic and therapeutic purposes.Citation12,Citation27
The results of this study also indicate that uPAR is an attractive target for exploitation in dogs with spontaneous brain tumors, which are a faithful animal model of human disease.Citation9,Citation10 The results reported here served as the justification for our development of a recombinant uPAR-targeted oncolytic Newcastle disease virus (NDV) for use in canine brain tumors. By genetically modifying a native LaSota NDV strain as previously described, such that the LaSota fusion (F) protein cleavage site is replaced with a uPA cleavage site,Citation28,Citation29 we have generated the LaSota–uPA recombinant NDV strain the F protein of which is cleavable exclusively by uPAR. A Phase I clinical trial investigating LaSota–uPA in canine meningiomas has been initiated.Citation30
Conclusion
We have shown that uPAR is overexpressed in multiple microenvironmental compartments in canine meningiomas, gliomas, and choroid plexus tumors. This study provides a rationale to develop and assess uPAR-targeted therapeutic strategies in naturally occurring canine brain tumors.
Acknowledgments
Portions of this study were funded by the Wallace H. Coulter Foundation Career Grant, the Virginia Biosciences Health Research Corporation, the Virginia Veterinary Memorial Fund, R01 CA139099/CA/NCI NIH HHS, and R21 AI070528/AI/NIAID NIH HHS. We acknowledge and appreciate the contributions of our friend and colleague, Dr Subbiah Elankumaran, who passed away on September 2, 2015, prior to completion of the article.
Disclosure
The authors report no conflicts of interest in this work.
References
- LiottaLAStetler-StevensonWGTumor invasion and metastasis: an imbalance of positive and negative regulationCancer Res199151Suppl 1850545059
- NohHHongSHuangSRole of urokinase receptor in tumor progression and developmentTheranostics20133748749523843896
- DanoKAndreasenPAGrondahl-HansenKKristensenPNielsenLSSkriverLPlasminogen activators, tissue degradation and cancerAdv Cancer Res1985441392662930999
- LeonardiLQuattriniIRopertoFBenassiMSProtease expression in giant cell tumour of bone: a comparative study on feline and human samplesRes Vet Sci201395231031523668898
- SantosALopesCMarquesRMImmunohistochemical analysis of urokinase plasminogen activator and its prognosis in canine mammary tumorsVet J20111891434820598596
- KwaaanHCMazarAPMcMahonBJThe apparent uPA/PAI-1 paradox in cancer: more than meets the eyeSemin Thromb Hemost201339438239123532574
- CollenDLijnenHRBasic and clinical aspects of fibrinolysis and thrombolysisBlood19917812311431241742478
- GilderASJonesJAHuJSoluble urokinase receptor is released selectively by glioblastoma cells that express epidermal growth factor receptor variant III and promotes tumor cell migration and invasionJ Biol Chem201529024147981480925837250
- DebinskiWDickinsonPJRossmeislJHRobertsonJGiboDNew agents for targeting of IL-13RA2 expressed in primary human and canine brain tumorsPLoS One2013810e7771924147065
- LeBlancAKMazckoCBrownDECreation of an NCI comparative brain tumor consortium: informing the translation of new knowledge from canine to human brain tumor patientsNeuro Oncol20161891209121827179361
- MohanPMChintalaSKMohanamSAdenovirus-mediated delivery of antisense gene to urokinase-type plasminogen activator suppresses glioma invasion and tumor growthCancer Res199959143369337310416596
- ValleraDALiCJinNPanoskaltsis-MortariAHallWATargeting urokinase-type plasminogen activator receptor on human glioblastoma tumors with diphtheria toxin fusion protein DTATJ Natl Cancer Inst200294859760611959893
- KargiotisOChettyCGogineniVuPA/uPAR downregulation inhibits radiation-induced migration, invasion, and angiogenesis in IOMM-Lee meningioma cells and decreased tumor growth in vivoInt J Oncol200833593794718949356
- RossmeislJHRobertsonJLZimmermanKLHigginsMAGeigerDACyclooxygenase-2 (COX-2) expression in canine intracranial meningiomasVet Comp Oncol20097317318019691646
- RossmeislJHJonesJCZimmermanKLRobertsonJLSurvival time following hospital discharge in dogs with palliatively treated primary brain tumorsJ Am Vet Med Assoc2013242219319823276095
- FaleiroMRToledoDCRodriguesMMPuPAR expression in canine normal prostate and with proliferative disordersCi Anim Bras Goiania2013142237244
- UntergasserANijveenHRaoXBisselingTGeurtsRLeunissenJAMPrimer3Plus, an enhanced web interface to Primer3Nucleic Acids Res200735W71W7417485472
- AltschulSFMaddenTLSchafferAAGapped BLAST and PSI-BLAST: a new generation of protein database search programsNucleic Acids Res19972517338934029254694
- KimTDSongKSLiGActivity and expression of urokinasetype plasminogen activator and matrix metalloproteinases in human colorectal cancerBMC Cancer2006621116916471
- JankunJSkrzypczak-JankunEMolecular basis of specific inhibition of urokinase plasminogen activator by amilorideCancer Biochem Biophys1999171–210912310738907
- DickinsonPJAdvances in diagnostic and treatment modalities for intracranial tumorsJ Vet Int Med201428411651185
- SongRBViteCHBradleyCWCrossJRPostmortem evaluation of 435 cases of intracranial neoplasia in dogs and relationship of neoplasm with breed, age, and body weightJ Vet Intern Med20132751143115223865437
- SnyderJMShoferFSVan WinkleTJMassicotteCCanine intracranial primary neoplasia: 173 cases (1986–2003)J Vet Intern Med200620366967516734106
- ArchintiMBrittoMEdenGFurlanFMurphyRDegryseBThe urokinase receptor in the central nervous systemCNS Neurol Disord Drug Targets201110227129420874700
- MohanamSSawayaREYamamotoMBrunerJMNicholsonGLRaoJSProteolysis and invasiveness of brain tumors: role of urokinase-type plasminogen activator receptorJ Neurooncol19942221531607745467
- VishnoiMPeddibhotlaSYinWThe isolation and characterization of CTC subsets related to breast cancer dormancySci Rep201551753326631983
- HirataKTamakiNuPAR as a glioma imaging targetJ Nuc Med2016572169170
- KimSHWanasenNPalduraiAXiaoSCollinsPLSamalSKNewcastle disease virus fusion protein is the major contributor to protective immunity of genotype-matched vaccinePLoS One201388e7402224015313
- ShobanaRSamalSKElankumaranSProstate-specific antigen-retargeted recombinant Newcastle disease virus for prostate cancer virotherapyJ Virol20138773792380023345509
- Phase I Clinical Trial of Recombinant Newcastle Disease Virus for Canine Intracranial MeningiomasVirginiaCollege of Virginia-Maryland Veterinary Medicine Available from: http://www.vetmed.vt.edu/clinical-trials/current-studies/meningioma.aspAccessed March 13, 2016