
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Circular RNAs are a large class of noncoding RNA that have shown huge capabilities as gene regulators. Recent evidence suggest that circular RNAs are associated with many diseases, especially cancer. However, little attention has been focused on the expression and function of circular RNA in gastric cancer (GC). In this study, we demonstrate that the expression of circ-104916 is downregulated in GC tissues and cell lines. A lower expression of circ-104916 appeared in deeper invasion depth, higher tumor stage and more frequent lymphatic metastasis patients. Overexpression of circ-104916 effectively inhibited the proliferation, migration and invasion abilities of GC cells in vitro. Western blot showed that circ-104916 overexpression upregulated E-cadherin and downregulated N-cadherin, Vimentin and Slug, indicating that circ-104916 was involved in the epithelial–mesenchymal transition process. Our results revealed that circ-104916 might be a novel potential tumor suppressor and biomarker of GC.
Introduction
Gastric cancer (GC) is the third leading cause of cancer death worldwide, which affects >400,000 patients/year in China.Citation1 More than 80% of the Chinese patients were diagnosed at advanced stages, on which surgery with curative intent required extended lymphadenectomy. Laparoscopic distal gastrectomy had been proved to be safe for advanced GC.Citation2 Even with advanced systematic therapy, the 5-year overall survival of patients with late-stage GC was not >20%.Citation3 Therefore, it is of great significance to investigate the mechanism of GC progression and metastasis.
Circular RNA (circRNA) was first detected over 2 decades ago, which was considered a byproduct of splicing errors.Citation4 With the development of high-throughput RNA sequencing and bioinformatics, it was clear that circular RNA was abundant and stable in eukaryotic cell.Citation5,Citation6 Moreover, studies had found out that circular RNA expression was dysregulated in pancreatic cancer,Citation7 ovarian cancer,Citation8 bladder cancerCitation9 and hepatocellular cancer.Citation10 CircRNA could act as microRNAs sponge to regulate gene expression. The most famous circular RNA was CDR1as, which contained >70 conventional binding sites and functioned as a super sponge for miR-7.Citation11 Recent studies showed that CDR1as was a risk factor of hepatic microvascular invasion in hepatocellular carcinoma (HCC)Citation12 and knockdown of CDR1as suppressed the HCC cell proliferation and invasion.Citation13 However, the expression and function of circular RNA in GC are still unknown.
In our previous study, via microarray, we found that the expression of circ-104916 was reduced in GC.Citation14 In this study, we found that circ-104916 was significantly downregulated in GC tissues by expending the sample size. In vitro studies showed that circ-104916 could suppress the proliferation, migration and invasion capabilities of GC cells. This is the first study to investigate the role of circ-104916 in GC, which suggests that circ-104916 may be a potential tumor suppressor and biomarker of GC.
Materials and methods
Tissues samples
Human samples including paired cancer specimens and adjacent normal tissues were obtained from the patients who underwent surgical resection from November 2015 to May 2016 at Nanfang Hospital, Southern Medical University. All fresh samples were immediately frozen after resection and stored at −80°C until use. Tumors were staged according to the TNM staging classification of stomach carcinoma of American Joint Committee on Cancer (AJCC; 7th ed, 2010). None of the patients had received chemotherapy or radiotherapy before surgery. The study was approved by the Clinical Research Ethics Committee of Nanfang Hospital of Southern Medical University. Written informed consent was obtained from all participants.
RNA extraction and quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNA was extracted from the cells and tissues using TRIzol reagent (Takara, Tokyo, Japan) according to the manufacturer’s protocol. After RNA extraction, reverse transcriptase (Takara) was used for synthesizing cDNA according to the manufacturer’s instructions. The expression level of the circRNAs was evaluated by quantitative PCR (qPCR) using SYBR Green assay. Specific divergent primers were designed to amplify the circular transcripts. PCR was performed in 10 μL reaction volume, including 2 μL of cDNA, 5 μL of 2× Master Mix (Takara), 0.4 μL of forward primer (10 μM), 0.4 μL of reverse primer (10 μM) and 2.2 μL of double distilled water. The reaction was set at 95°C for 10 min for pre-denaturation and then at 95°C for 10 s and at 60°C for 60 s repeating 40 cycles. β-actin was used as a reference. We used the following primer sequences: circ-104916 – forward: 5′-GCTCGGTGACCTTGGTCTGG-3′ and reverse: 5′-GCGTGTTGGGATGCCTCTGT-3′ and β-actin – forward: 5′-TGGATCAGCAAGCAGGAGTA-3′ and reverse: 5′-TCGGCCACATTGTGAACTT-3′. Both target and reference were amplified in triplicate wells. The relative level of each circRNA was calculated using the method.
Cell culture
Human GC cell lines (AGS, MKN-28, NCI-N87 and MKN-45) and human gastric mucosa epithelial cells (GES-1) were propagated in Roswell Park Memorial Institute (RPMI) 1640 (HyClone, Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS; Capricorn, Ebsdorfergrund, Germany).
Circular RNA plasmid construction and transfection
We constructed the circular RNA plasmid used in this study. Human circ-104916 cDNA was synthesized by GENEWIZ (Suzhou, China) and cloned into pcDNA3.1. We used a 651-bp DNA fragment corresponding to exon 1, exons 3–6 and exon 8 of the NEK6 gene and added 1 kb upstream and 200 bp downstream to the nonlinear splice sites. In addition, an 800-bp DNA stretch was added upstream of the splice acceptor site and inserted downstream in the reverse orientation.Citation11 The GC cells were seeded at six-well plates and then transfected at 48 h with pcDNA-hsa-circ-104916 (4 μg) or control plasmid (4 μg) using Lipofectamine 3000 Transfection Reagent according to the manufacturer’s protocol (Thermo Fisher Scientific, Waltham, MA, USA).
Cell proliferation assay
Cell proliferation was assayed using the Cell Counting Kit-8 (CCK-8) assay (Dojindo Laboratories, Kumamoto, Japan) according to the manufacturer’s protocol. The transfected cells were plated in 96-well plates (1,500 cells/well). Cell proliferation was detected every 24 h according to the manufacturer’s protocol. Briefly, 10 μL of CCK 8 solution was added to each well and incubated for 2 h at 37°C. The solution was then measured spectrophotometrically at 450 nm.
In vitro cell migration and invasion assays
A total of 1×105 BGC823 and MGC803 cells transfected for 48 h with 4 μg pcDNA-hsa-circ-104916 or pcDNA3.1 were plated in the upper chamber of transwell assay inserts (Corning Incorporated, Corning, NY, USA), which contained 200 μL of serum-free RPMI 1640 with a membrane (8 mm pores) to test migration. The lower chambers were filled with RPMI 1640 containing 10% FBS. The cells on the filter surface were fixed with methanol, stained with crystal violet and photographed with a digital microscope after 24 h. The cell numbers were calculated in five random fields for each chamber.
The transfected cells (1×106) were plated in the top chamber (Corning Incorporated) that was precoated with Matrigel (BD Biosciences, San Jose, CA, USA) in 200 μL serum-free RPMI 1640 to test cell invasion. There was 750 μL of 10% FBS-RPMI 1640 in the bottom chambers. The invasion function was determined after 48 h.
Western blot assay
The cellular proteins were extracted 48 h after transfection. The cells in each well were lyzed in 50 μL of lysis buffer containing radioimmunoprecipitation assay (RIPA; 49.5 μL) and protease inhibitor cocktail (0.5 μL). Then, 30 μg of protein was resolved by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) analysis and transferred onto a polyvinylidene difluoride (PVDF) membrane. The primary antibodies used were anti-E-cadherin (1:1,000; Abcam, Cambridge, UK), anti-N-cadherin (1:5,000; Abcam), anti-Vimentin (1:1,000; Abcam), anti-Slug (1:1,000; Abcam), anti-Snail (1:1,000; Abcam) and anti-GAPDH (1:10,000; Proteintech group, Inc., Chicago, IL, USA). The membranes were washed and then incubated with goat anti-rabbit horseradish peroxidase (HRP)-conjugated secondary antibody (1:10,000; Abcam) for 2 h at room temperature. The blots were visualized by enhanced chemilu-minescence detection.
Statistical analysis
All statistical analyses were performed using SPSS 23.0 software (IBM Corporation, Armonk, NY, USA). Data were expressed as mean ± standard error of the mean (SEM) from at least three separate experiments. Differences between groups were analyzed using Student’s t-test analysis. Correlation between the clinicopathological data and circ-104916 was analyzed using chi-square test. P-value <0.05 was considered as statistically significant.
Results
Downregulation of circNEK6 in human GC
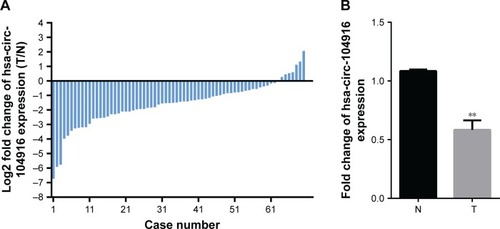
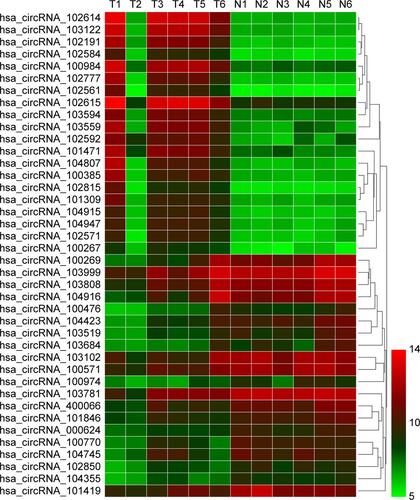
In our previous study, the microarray results suggested that the expression of circRNAs in gastric carcinoma tissues was abnormal compared with adjacent normal tissues (Figure S1). The expression of circ-104916 was reduced in GC tissues. To confirm hsa-circ-104916 expression in GC, qRT-PCR was performed in 70 pairs of human GC specimens and their adjacent noncancerous tissue samples. More than 50% reduction in circ-104916 expression was observed in 65.7% (46/70) of GC tissues (). The expression of circ-104916 was significantly lower in GC tissues than in the corresponding adjacent noncancerous tissues (). Theses results suggested that circ-104916 was downregulated in GC tissues, which was consistent with our microarray result.
Figure 1 Circ-104916 expression in GC tissues.
Abbreviation: qPCR, quantitative polymerase chain reaction.
Correlation with the clinicopathological data and expression of circ-104916
To assess the coloration of circ-104916 expression with clinicopathological data, the level of circ-104916 in tumor tissue was categorized as low group or high group in relation to the media value of relative circ-104916 expression (0.36-fold, tumors/no cancers). Clinicopathological factors were analyzed in the high or low groups. The expression of circ-104916 was significantly associated with invasion depth, tumor stage, lymphatic metastasis and nerve invasion (). However, there was no significant correlation between circ-104916 expression and other clinicopathological factors, such as age, histological grade and vascular invasion. Moreover, a lower expression of circ-104916 appeared in deeper invasion depth, higher tumor stage and more frequently lymphatic metastasis patients (). Taking together, our results showed that circ-104916 expression was associated with clinicopathological features and might be a potential biomarker of GC.
Figure 2 Clinicopathological factors with the expression level of circ-104916.
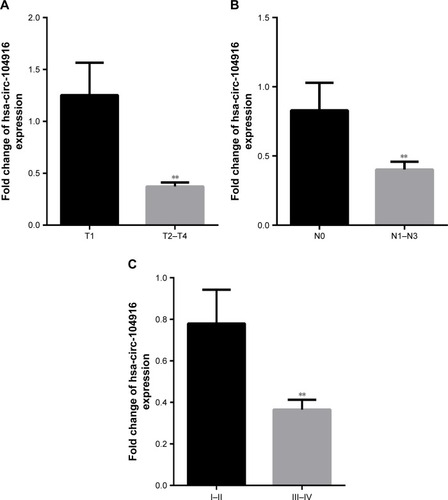
Table 1 Correlation with the clinicopathological data and expression of circ-104916
Circ-104916 suppresses proliferation of GC cells in vitro
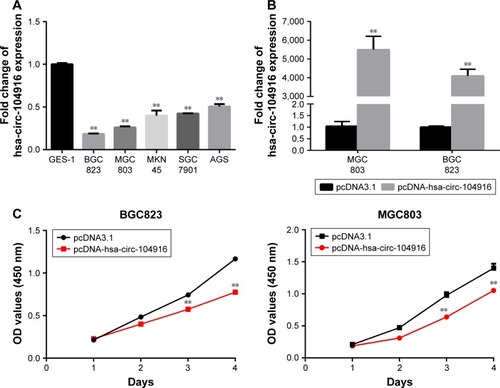
To detect the function of circ-104916 in the progression of GC, we performed gain of function assay. First, we analyzed circ-104916 expression in human GC cell lines. The expression of circ-104916 in AGS, MKN45, BGC823, MGC803 and 7901 was significantly lower than that in the normal gastric mucosa cell line GES-1 (), which was consistent with our clinical findings that human GC tissues had low levels of circ-104916. According to the endogenous expression of circ-104916 in five GC cell lines, we transfected pcDNA-hsa-circ-104916 or negative control in BGC823 and MGC803 cell lines. After 48 h of treatment, the expression of circ-104916 was effectively increased (). Then, we assessed the effect of circ-104916 on the proliferation of GC cells though CCK-8 assay. The result showed that overexpression of circ-104916 could suppress the proliferation of BGC823 and MGC803 cell lines (). Our results indicated that circ-104916 was downregulated in GC cells and could inhibit the proliferation of GC cells.
Figure 3 Circ-104916 overexpression suppressed the proliferation of GC cells.
Abbreviations: GC, gastric cancer; CCK-8, Cell Counting Kit-8; OD, optical density; SEM, standard error of the mean.
Circ-104916 suppresses GC cell migration and invasion
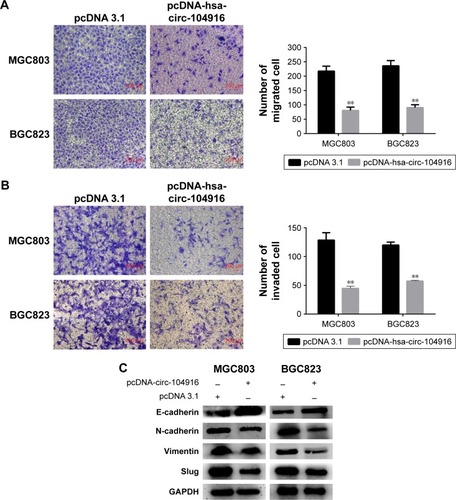
Migration and invasion assays were used to determine the change of migration and invasion capability of BGC823 and MGC803 cells, which were both transfected with pcDNA-circ-104916. The migration assay indicated that overexpression of circ-104916 significantly suppressed the migration capability of BGC823 and MGC803 cells (). Meanwhile, the invasion assay showed that overexpression of circ-104916 could inhibit the invasion capability of BGC823 and MGC803 cells (). These results suggested that circ-104916 could inhibit the migration and invasion of GC cells.
Figure 4 Circ-104916 inhibited the migration and invasion of GC cells.
Abbreviations: GC, gastric cancer; EMT, epithelial–mesenchymal transition.
Circ-104916 suppresses GC cell invasion and migration via the epithelial–mesenchymal transition (EMT) process
To further explore the underlying mechanism of decreased invasion and migration by circ-104916, EMT molecules were evaluated. The upregulation of E-cadherin was observed in circ-104916 overexpression of GC cells. Additionally, the downregulation of N-cadherin and Vimentin were also observed in circ-104916 overexpression of GC cells. The decreased expression of Slug was accompanied with circ-104916 overexpression (). In conclusion, our results suggested that circ-104916 might suppress GC cell migration and invasion via inhibiting the EMT process.
Discussion
In recent years, with the rapid development of RNA sequencing, researches have discovered that circRNA is abundant and especially expresses in a cell type or development stage.Citation5 Circular RNAs can modulate cellular physiology as microRNA sponges. For instance, the circ-ITCH suppresses esophageal squamous cell carcinoma via the Wnt/β-catenin pathway.Citation15 Circ-001569 could promote proliferation and invasion of colorectal cancer via target mir-145.Citation16 Previous studies also elaborated that circRNA could affect cell processes by forming RNA–protein complexes.Citation17,Citation18 For example, Circ-Foxo3 not only could block cell-cycle progression via forming ternary complexes with p21 and CDK2Citation17 but also induces cell apoptosis via binding MDM2 and P53.Citation18 Recent studies revealed that circular RNA could regulate transcription or splicing. Circular intronic RNA may have a cis-regulatory role on their parent coding genes.Citation19 Thus, circRNA may be a core participant in physiology and pathology processes. To the best of our knowledge, there were few studies about circular RNA in GC. Only one study showed that circ-002059 in GC was significantly associated with TNM stage and distal metastasis.Citation20 However, the effect and function of circRNA in GC remains unknown.
In our study, via microarray, we first found that circ-104916 was significantly lower in the GC tissue than the adjacent normal tissues. Through database CircBase,Citation21 we figured out that circ-104916 was generated from chromosomal region 9q33.3, which was back spliced by exon 1, exon 3, exon 4, exon 5, exon 6 and exon 8 of NEK6. Mature circ-104916 transcript is a circular RNA molecule of 651nt. Then, we validated the expression of circ-104916 in 70 matched samples. The result was coincident with the microarray finding. Moreover, deeper invasion depth, higher tumor stage and more frequent lymphatic metastasis patients had a lower expression of circ-104916. These results indicated that circ-104916 might be a potential biomarker for GC.
In in vitro experiments, the results suggested that circ-104916 could suppress the proliferation, migration and invasion abilities of GC cells. Therefore, circ-104916 may be a potential tumor suppressor and play a core role in GC invasion and metastasis. EMT is a critical cellular progress in cancer metastasis.Citation22 What is more, a previous study had reported that hundreds of circRNAs were regulated during human EMT.Citation23 The key events in EMT are dissolution of the epithelial cell–cell junction, downregulation of an epithelial gene expression signature and activation of genes that help to define the mesenchymal phenotype and increased cell protrusions and motility.Citation24 By Western blot, we found that the epithelial molecule (E-cadherin) was upregulated and the mesenchymal molecule (N-cadherin, Vimentin) was down-regulated after circ-104916 overexpression. Slug (also known as Snail2) is a member of the SNAIL family of zinc finger transcriptional repressors, which can repress the E-cadherin expression.Citation25 In our data, circ-104916 could decrease Slug expression. Taking together, the results suggested that circ-104916 might have epithelial-specific functions through decreasing the Slug expression.
Our study has several limitations. First, we did not perform loss of function because of the low efficiency of siRNA resulting from the high GC rate around the back-splice sites. Second, these conclusions were based on the responses of two cell lines and may not reflect the process in the organism. The conformance in mice needs to be accomplished in the future.
In summary, our data first found that circ-104916 was downregulated in GC tissues and the expression of circ-104916 was related to invasion depth, tumor stage and lymphatic metastasis. The overexpression of circ-104916 suppressed the migration and invasion of GC cells through altering the EMT process. Our research suggested that circ-10496 might be a novel potential tumor suppressor and biomarker of GC.
Acknowledgments
The support of Zhen Han in data analysis was essential in this paper. This work was supported by the grants from Guangdong Provincial Science and Technology Key Project (2014A020215014), the Research Fund of Public Welfare in the Health Industry, the National Health and Family Planning Commission of China (201402015) and the Key Clinical Specialty Discipline Construction Program, National Natural Science Foundation of China (81672446 and 81600510).
Supplementary material
Figure S1 Hierarchical clustering analysis of the most up- and down regulated circRNAs.
Notes: Each column represents a sample, and each row represents a circRNA. Red strip represents a high relative expression, and green strip represents a low relative expression. T represents GC tissues and N represents normal gastric mucosa.
Abbreviation: GC, gastric cancer.
Disclosure
The authors report no conflicts of interest in this work.
References
- JemalABrayFCenterMMFerlayJWardEFormanDGlobal cancer statisticsCA Cancer J Clin2011612699021296855
- HuYHuangCSunYMorbidity and mortality of laparoscopic versus open D2 distal gastrectomy for advanced gastric cancer: a randomized controlled trialJ Clin Oncol201634121350135726903580
- QiXLiuYWangWManagement of advanced gastric cancer: an overview of major findings from meta-analysisOncotarget2016747781807820527655725
- SangerHLKlotzGRiesnerDGrossHJKleinschmidtAKViroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structuresProc Natl Acad Sci U S A19767311385238561069269
- MemczakSJensMElefsiniotiACircular RNAs are a large class of animal RNAs with regulatory potencyNature2013495744133333823446348
- QiXZhangDHWuNXiaoJHWangXMaWceRNA in cancer: possible functions and clinical implicationsJ Med Genet2015521071071826358722
- QuSSongWYangXMicroarray expression profile of circular RNAs in human pancreatic ductal adenocarcinomaGenom Data2015538538726484292
- AhmedIKaredathTAndrewsSSAltered expression pattern of circular RNAs in primary and metastatic sites of epithelial ovarian carcinomaOncotarget2016724363663638127119352
- ZhongZLvMChenJScreening differential circular RNA expression profiles reveals the regulatory role of circTCF25-miR-103a-3p/miR-107-CDK6 pathway in bladder carcinomaSci Rep201663091927484176
- ShangXLiGLiuHComprehensive circular RNA profiling reveals that hsa_circ_0005075, a new circular RNA biomarker, is involved in hepatocellular carcinoma developmentMedicine (Baltimore)20169522e381127258521
- HansenTBJensenTIClausenBHNatural RNA circles function as efficient microRNA spongesNature2013495744138438823446346
- XuLZhangMZhengXYiPLanCXuMThe circular RNA ciRS-7 (Cdr1as) acts as a risk factor of hepatic microvascular invasion in hepatocellular carcinomaJ Cancer Res Clin Oncol20171431172727614453
- YuLGongXSunLZhouQLuBZhuLThe circular RNA Cdr1as act as an oncogene in hepatocellular carcinoma through targeting miR-7 expressionPLoS One2016117e015834727391479
- ZhangYLiJYuJCircular RNAs signature predicts the early recurrence of stage III gastric cancer after radical surgeryOncotarget2017814229362294328206972
- LiFZhangLLiWCircular RNA ITCH has inhibitory effect on ESCC by suppressing the Wnt/β-catenin pathwayOncotarget2015686001601325749389
- XieHRenXXinSEmerging roles of circRNA_001569 targeting miR-145 in the proliferation and invasion of colorectal cancerOncotarget2016718266802669127058418
- DuWWYangWLiuEYangZDhaliwalPYangBBFoxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2Nucleic Acids Res20164462846285826861625
- DuWWFangLYangWInduction of tumor apoptosis through a circular RNA enhancing Foxo3 activityCell Death Differ201724235737027886165
- ZhangYZhangXOChenTCircular intronic long noncoding RNAsMol Cell201351679280624035497
- LiPChenSChenHUsing circular RNA as a novel type of biomarker in the screening of gastric cancerClin Chim Acta201544413213625689795
- GlažarPPapavasileiouPRajewskyNcircBase: a database for circular RNAsRNA201420111666167025234927
- NietoMAEpithelial plasticity: a common theme in embryonic and cancer cellsScience20133426159123485024202173
- ConnSJPillmanKAToubiaJThe RNA binding protein quaking regulates formation of circRNAsCell201516061125113425768908
- QiXZhangLLuXNew insights into the epithelial-to-mesenchymal transition in cancerTrends Pharmacol Sci201637424624826837734
- HajraKMChenDYFearonERThe SLUG zinc-finger protein represses E-cadherin in breast cancerCancer Res20026261613161811912130