
Abstract
Background
The At-rich interactive domain 1A (ARID1A) is frequently mutated in gastric cancers (GCs) with a poor prognosis. Growing evidence indicates that loss of ARID1A expression leads to activation of the phosphatidylinositol 3-kinase (PI3K)/AKT pathway by AKT phosphorylation. We aim to investigate the different sensitivity for the AKT inhibitor in ARID1A-deficient GC cells.
Methods
After transfection using siRNA or shRNA, the effect of ARID1A knockdown on the PI3K/AKT signaling pathway was evaluated by Western blot analysis. ARID1A-knockdown cells were treated with AKT inhibitor (GSK690693), 5-fluorouracil, or cisplatin, alone or in combination. Viability and apoptosis were analyzed using EZ-CYTOX cell viability assay and flow cytometry, respectively.
Results
ARID1A depletion accelerated the phosphorylation of AKT and S6 in a dose-dependent manner and led to an increased proliferation of MKN-1, MKN-28, and KATO-III GC cells (P<0.001). ARID1A-deficient cells were more vulnerable to GSK690693 in comparison to the controls (P<0.001), even at very low doses. Flow cytometry confirmed the increased apoptosis in ARID1A-deficient cells treated with GSK690693 (0.01 μmol/L; P<0.001). In contrast to our expectations, ARID1A depletion did not cause resistance to 5-fluorouracil or cisplatin. Addition of GSK690693 to the conventional chemotherapy induced more decreased cell viability in ARID1A-knockdown cells (P<0.01).
Conclusion
Loss of ARID1A expression is a surrogate marker for the activation of the AKT signaling pathway and is also a reliable biomarker to predict the response for the AKT inhibitor. We anticipate that appropriate patient selection based on ARID1A expression in the tumor tissue will increase the drug sensitivity for the AKT inhibition and improve the clinical outcome.
Introduction
Despite recent improvements in surgical and chemotherapeutic approaches, about half of the patients diagnosed with advanced gastric cancer (GC) still die from recurrent disease after curative surgery or distant metastasis.Citation1,Citation2 Fluoropyrimidine- or platinum-based conventional cytotoxic chemotherapy is still widely used as the first-line treatment of GC.Citation3 However, nearly all patients develop resistance to these drugs with adverse effects, limiting the efficacy of this approach.Citation4,Citation5 The only targeted therapeutic agent, trastuzumab (Roche, Basel, Switzerland), increases survival by only a few months, even in a selected biomarker-positive population.Citation1 Thus, identification of efficacious therapeutic interventions that sensitize GC cells is required to improve the management of GC patients.
The AT-rich interactive domain 1A (ARID1A) is an ARID family member that encodes a subunit of the Switch/Sucrose nonfermentable chromatin remodeling family.Citation6 ARID1A is involved in regulating diverse cellular processes, including development, differentiation, proliferation, and DNA repair.Citation7 Recent genome-wide sequencing studies demonstrated that ARID1A is frequently mutated in ovarian and endometrial cancers, and also in GCs, ranging from 8% to 33%.Citation8–Citation12 Genetic alterations in ARID1A in GCs are mostly insertion/deletion mutations, which lead to truncation of ARID1A protein.Citation10 Loss of ARID1A protein is considered a poor prognostic factor in a variety of cancers, including GC.Citation13
Growing evidence indicates that loss of ARID1A expression leads to the activation of the phosphatidylinositol 3-kinase (PI3K)/AKT pathway, mainly due to AKT phosphorylation.Citation14–Citation16 This appears to be true in GCs, as we previously demonstrated in the human GC tissues by immunohistochemistry.Citation17 If ARID1A-deficient tumor cells depend on constitutive activation of the AKT pathway, it is assumed that tumor cells lacking ARID1A expression may be more vulnerable to the selective AKT pathway inhibitors.Citation18 In support of this notion, MCF7 breast cancer cells and primary MRC5 cells exhibit significantly increased sensitivity to the AKT inhibitors, MK-2206 and perifosine, as well as the PI3K inhibitor, buparlisib, after ARID1A knockdown.Citation19
In this study, we demonstrate that loss of ARID1A promotes proliferation of cancer cells by activating the AKT signaling pathway in GC cell lines. The promising preclinical activity of selective AKT pathway inhibitors was demonstrated in ARID1A-deficient GC cells.
Materials and methods
Cell lines and cell culture
MKN-1, MKN-28, MKN-45, KATO-III, AGS, MKN-74, and NCI-N87 human GC cell lines were purchased from the Korean Cell Line Bank and were maintained in RPMI 1640 (Hyclone, South Lagan, UT, USA) supplemented with 10% fetal bovine serum, 1% penicillin, and streptomycin at 37°C in a humidified atmosphere containing 5% CO2.
Reagents and antibodies
The following reagents and antibodies were used: GSK690693 (Cell Signalling Technology, Danvers, MA, USA), 5-fluorouracil (5-FU) and cisplatin (both from Sigma-Aldrich, St Louis, MO, USA), ARID1A, p-AKT, AKT, p-S6, poly-ADP ribose polymerase (1:1,000; Cell Signalling Technology), and β-actin (1:5,000; Santa Cruz Biotechnology, Santa Cruz, CA, USA).
Western blot analysis
After treatment with the specified drugs, cells were washed with PBS and lysed in the lysis buffer. Lysates were centrifuged at 13,000 rpm for 20 min at 4°C and then incubated on ice for 20 min. Protein concentration was determined by the Bradford assay (Bio-Rad, Hercules, CA, USA). Equal amounts of protein from each sample were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto polyvinylidene difluoride membrane (EMD Millipore, Billerica, MA, USA). The immunoblots were blocked by incubation in 5% skim milk, 25 mM Tris-HCl (pH 8.0), 150 mM NaCl, and 0.1% Tween 20 for 1 hour at room temperature. The membrane was then incubated with primary antibodies on a shaker overnight at 4°C. Protein detection was performed using the ECL kit (Abclon, Seoul, Korea).
RNA interference
ARID1A depletion was done by transfection of MKN-1, MKN-28, and KATO-III cells with SMARTpool: ON-TARGETplus ARID1A siRNA (L-017263-00-0005; GE Dharmacon, Lafayette, CO, USA). siRNAs were transfected using DharmaFECT1 (GE Dharmacon). In addition, MKN-1, MKN-28, and KATO-III cells were infected with a lentivirus encoding a nonspecific (NS) scramble shRNA (#1864; Addgene, Cambridge, MA, USA), Mission ARID1A shRNA#2 (NM00615.3-TRCN0000059089; Sigma-Aldrich), or Mission ARID1A shRNA#3 (NM00615.3-TRCN0000059090; Sigma-Aldrich) using polybrene (EMD Millipore). After infection, cells were selected with puromycin (Sigma-Aldrich) and ARID1A expression levels were checked by Western blot.
Cell viability assay
After 48-hour exposure of specified drugs, cells were analyzed using EZ-CYTOX cell viability assay kit (Daeillab Services, Seoul, Korea). Cells were plated in 96-well plates. EZ-CYTOX solution was added to each well of the plates and incubated at 37°C for 1.5–2 hours. The absorbance was measured at 450 nm.
Cell apoptosis
To detect synergistic effects of the chemotherapeutic drugs on cell apoptosis, treated cells were analyzed using the fluorescein isothiocyanate-Annexin V apoptosis kit (BD Biosciences, San Jose, CA, USA). The results were analyzed by flow cytometry using an FACSAria III device (BD Biosciences). All the experiments were repeated independently in triplicate.
Statistical analysis
Statistical analysis was performed using GraphPad Prism version 5.0 (GraphPad Software Inc., La Jolla, CA, USA). The half inhibitory concentration (IC50) values were calculated with an F-test. For two-group comparisons, a two-tailed unpaired t-test was used. For multiple group comparisons, one-way analysis of variance was used. Two-sided P<0.05 was considered statistically significant.
Results
ARID1A knockdown activates AKT pathway and promotes growth of GC cells
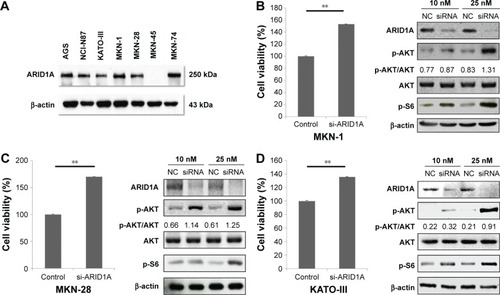
ARID1A protein levels in different GC cell lines were evaluated using Western blotting. Most of the GC cells showed intact ARID1A expression, while MKN-45 was ARID1A deficient (). Then, we selected MKN-1, MKN-28, and KATO-III and silenced endogenous ARID1A using siRNAs in these cells. The siRNA remained as effective till 48 hours after transfection. ARID1A depletion increased cell proliferation compared to the controls (P<0.001), and it accelerated the phosphorylation of AKT and its downstream S6 proteins in a dose-dependent manner ().
Figure 1 Cellular proliferation by AKT phosphorylation is induced by ARID1A knockdown.
ARID1A-deficient GC cells are vulnerable to select AKT inhibitors
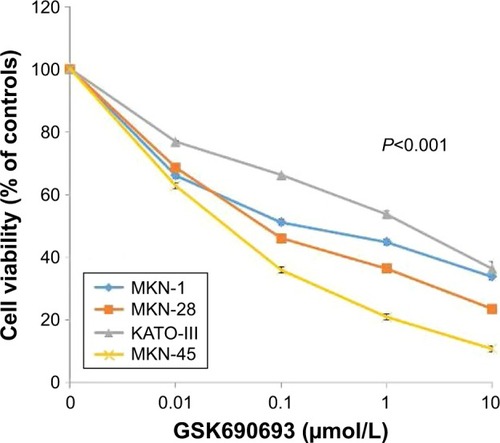
After transfection with control-siRNA and siARID1A, GC cells were incubated with increasing concentrations (0.01–10 μmol/L) of the AKT inhibitor, GSK690693, for 48 hours, followed by cell viability detection (). ARID1A-deficient cells were more vulnerable to the AKT inhibitor compared to the controls (P<0.001), even at very low doses. Consistent with this, ARID1A-deficient MKN-45 cells were most sensitive to GSK690693. The IC50 value of the wild-type (ARID1A-loss) MKN-45 was 0.043, while the IC50 values of the wild-type (ARID1A-intact) MKN-1, MKN-28, and KATO-III cells were 0.132, 0.084, and 4.521, respectively (P<0.001; ).
Figure 2 ARID1A depletion leads to increased sensitivity toward AKT pathway inhibitors.
Abbreviation: NC, normal control.
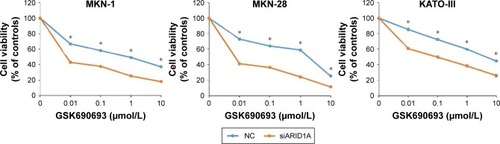
Figure 3 Loss of ARID1A expression is associated with high sensitivity to the AKT inhibitor in gastric cancer cell lines.
Abbreviation: IC50, half inhibitory concentration.
AKT inhibition leads to increased apoptosis in ARID1A-deficient cells
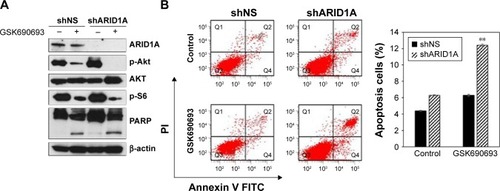
Treatment with the GSK690693 (10 μmol/L) completely abrogated p-Akt induced by ARID1A knockdown in ARID1A-deficient MKN-28 cells and led to reduced p-S6, in contrast to the controls (). Poly-ADP ribose polymerase cleavage was increased in ARID1A-knockdown cells treated with GSK690693. Flow cytometry confirmed the increased apoptosis in ARID1A-deficient cells treated with even low dose (0.01 μmol/L) of GSK690693, compared to the controls (P<0.001; ).
Figure 4 AKT inhibition leads to increased apoptosis in ARID1A-deficient cells.
Abbreviations: FITC, fluorescein isothiocyanate; PARP, poly-ADP ribose polymerase.
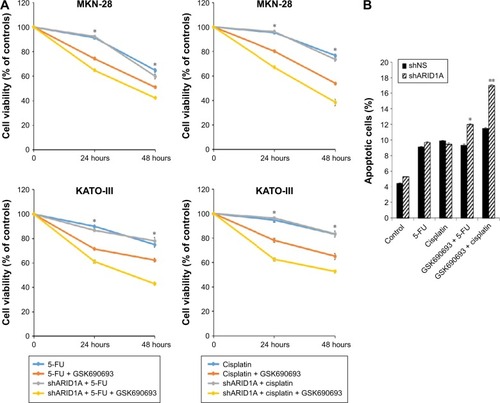
Addition of AKT inhibitors to conventional chemotherapy exerts increased antitumor activity in ARID1A-deficient cancer cells
Since ARID1A knockdown increased the proliferation of cancer cells by activating AKT signaling, we hypothesized that ARID1A-depleted cells might be resistant to conventional chemotherapy. The antiproliferative effect of conventional chemotherapy was examined in ARID1A-depleted GC cells. 5-FU or cisplatin was applied at different concentrations (10–60 μmol/L) for 48 hours to MKN-1, MKN-28, and KATO-III cells transfected with control-shRNA and shARID1A. In contrast to our expectations, ARID1A depletion did not cause resistance to these drugs (). Next, to further explore the synergistic effect of GSK690693 with conventional chemotherapy, the cells were treated with 5-FU (10 μmol/L) or cisplatin (10 μmol/L) in the presence of a minimal drug concentration of GSK690693 (0.01 μmol/L). Compared with single agent alone, addition of GSK690693 to the conventional chemotherapy induced more decreased cell viability in ARID1A-knockdown MKN-28 and KATO-III cells, compared to wild-type cells (P<0.01; ). Consistent with this observation, GSK690693 in combination with 5-FU or cisplatin induced a significant increase in apoptosis compared with 5-FU or cisplatin alone in ARID1A-knockdown cells (P<0.01; )
Figure 5 Loss of ARID1A expression did not induce resistance to the conventional chemotherapy.
Abbreviations: 5-FU, 5-fluorouracil; GC, gastric cancer.
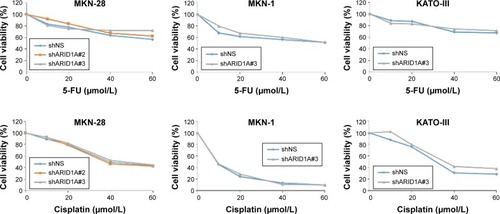
Figure 6 Addition of AKT inhibitors to conventional chemotherapy increases antitumor activity in ARID1A-deficient cancer cells.
Abbreviation: 5-FU, 5-fluorouracil.
Discussion
We demonstrate that loss of ARID1A induces cellular proliferation by activating the PI3K/AKT signaling pathway. ARID1A-deficient cancer cells were much more vulnerable to the AKT inhibitor, possibly due to the more significant abrogation of p-AKT and its downstream node p-S6 in ARID1A-depleted cancer cells treated with GSK690693. ARID1A knockdown sensitizes MCF7 breast cancer cells and HGC-27/SGC-7901 GC cells to PI3K and AKT inhibition.Citation19,Citation20 We confirmed this using different GC cell lines, suggesting that this phenomenon is ubiquitous regardless of cancer types. In addition, combination treatment of AKT inhibitor with conventional chemotherapy had an additive effect in ARID1A-knockdown GC cells, which might have utility in the future.
The PI3K/AKT pathway is frequently altered in cancers, promoting growth, proliferation, and survival.Citation21,Citation22 Targeting its major nodes, such as PI3K or AKT, may represent a key therapeutic opportunity.Citation21 Since the PI3K/AKT pathway has also been implicated in carcinogenesis and progression of GC, clinical investigations have targeted the PI3K/AKT pathway in patients with GC.Citation23 However, the clinical efficacy of these inhibitors as monotherapy in particular has, so far, been limited, showing very low response rate.Citation24 These failures possibly stem from the lack of appropriate patient selection based on a reliable biomarker.
Here, we present ARID1A as a surrogate marker for the activation of AKT pathway and may also anticipate more improved clinical response when patients with ARID1A-negative GC are treated with AKT inhibitors. Moreover, unlike other protein biomarkers, the expression status of ARID1A in tumor tissue can be clearly distinguished and leaves no concern for inter-observer variability in most cases.Citation17
PI3K/AKT has been implicated for chemotherapy resistance, including resistance for trastuzumab treatment.Citation25 A clinical investigation revealed that the loss of ARID1A expression was related to shorter progression-free survival and chemoresistance in ovarian clear cell carcinoma.Citation26 ARID1A gene silencing reduced the sensitivity of ovarian clear cell carcinoma to cisplatin.Citation27 However, in contrast to our expectation, ARID1A-deficient GC cells did not show resistance when treated with conventional chemotherapy, such as 5-FU and cisplatin. Viability of ARID1A-knockdown MKN-28 cells treated with paclitaxel was also similar to that of wild-type cells (data not shown). The exact relationship between ARID1A loss and chemoresistance needs to be investigated further.
The limitations of our study are that only one AKT inhibitor was used and the lack of in vivo experiments. Further assessment of the mechanism of action could shed light on the importance and clinical utility of using AKT inhibitors for treating patients having ARID1A-deficient GC, either singly or in combination.
Conclusion
Loss of ARID1A expression is a surrogate marker for the activation of AKT signaling pathway and also a reliable biomarker to predict the response for the AKT inhibitor. We anticipate that appropriate patient selection based on ARID1A expression in the tumor tissue will increase the drug sensitivity for AKT inhibition and improve the clinical outcome. Future mechanistic and in vivo studies should be performed prior to exploring this pathway in clinical trials.
Acknowledgments
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF), funded by the Ministry of Education (2017R1C1B2003970). This research was also supported by the Bio & Medical Technology Development Program of the NRF funded by the Korean government, MSIP (2015M3A9B6073827).
Disclosure
The authors report no conflicts of interest in this work.
References
- BangYJVan CutsemEFeyereislovaATrastuzumab in combination with chemotherapy versus chemotherapy alone for treatment of HER2-positive advanced gastric or gastro-oesophageal junction cancer (ToGA): a phase 3, open-label, randomised controlled trialLancet2010376974268769720728210
- LeeJLimDHKimSPhase III trial comparing capecitabine plus cisplatin versus capecitabine plus cisplatin with concurrent capecitabine radiotherapy in completely resected gastric cancer with D2 lymph node dissection: the ARTIST trialJ Clin Oncol201230326827322184384
- WadhwaRSongSLeeJSYaoYWeiQAjaniJAGastric cancer-molecular and clinical dimensionsNat Rev Clin Oncol2013101164365524061039
- CunninghamDStarlingNRaoSCapecitabine and oxaliplatin for advanced esophagogastric cancerN Engl J Med20083581364618172173
- CervantesARodaDTarazonaNRoselloSPerez-FidalgoJACurrent questions for the treatment of advanced gastric cancerCancer Treat Rev2013391606723102520
- DallasPBPacchioneSWilskerDBowrinVKobayashiRMoranEThe human SWI-SNF complex protein p270 is an ARID family member with non-sequence-specific DNA binding activityMol Cell Biol20002093137314610757798
- ReismanDGlarosSThompsonEAThe SWI/SNF complex and cancerOncogene200928141653166819234488
- Cancer Genome Atlas Research, NetworkKandothCSchultzNIntegrated genomic characterization of endometrial carcinomaNature20134977447677323636398
- WiegandKCShahSPAl-AghaOMARID1A mutations in endometriosis-associated ovarian carcinomasN Engl J Med2010363161532154320942669
- WangKKanJYuenSTExome sequencing identifies frequent mutation of ARID1A in molecular subtypes of gastric cancerNat Genet201143121219122322037554
- WangKYuenSTXuJWhole-genome sequencing and comprehensive molecular profiling identify new driver mutations in gastric cancerNat Genet201446657358224816253
- ZangZJCutcutacheIPoonSLExome sequencing of gastric adenocarcinoma identifies recurrent somatic mutations in cell adhesion and chromatin remodeling genesNat Genet201244557057422484628
- YangLWeiSZhaoRWuYQiuHXiongHLoss of ARID1A expression predicts poor survival prognosis in gastric cancer: a systematic meta-analysis from 14 studiesSci Rep201662891927354232
- LiangHCheungLWLiJWhole-exome sequencing combined with functional genomics reveals novel candidate driver cancer genes in endometrial cancerGenome Res201222112120212923028188
- BosseTter HaarNTSeeberLMLoss of ARID1A expression and its relationship with PI3K-Akt pathway alterations, TP53 and microsatellite instability in endometrial cancerMod Pathol201326111525153523702729
- WiegandKCHennessyBTLeungSA functional proteogenomic analysis of endometrioid and clear cell carcinomas using reverse phase protein array and mutation analysis: protein expression is histotype-specific and loss of ARID1A/BAF250a is associated with AKT phosphorylationBMC Cancer20141412024559118
- KimYBHamIHHurHLeeDVarious ARID1A expression patterns and their clinical significance in gastric cancersHum Pathol201649617026826411
- SamartzisEPNoskeADedesKJFinkDImeschPARID1A mutations and PI3K/AKT pathway alterations in endometriosis and endometriosis-associated ovarian carcinomasInt J Mol Sci2013149188241884924036443
- SamartzisEPGutscheKDedesKJFinkDStuckiMImeschPLoss of ARID1A expression sensitizes cancer cells to PI3K- and AKT-inhibitionOncotarget20145145295530324979463
- ZhangQYanHBWangJChromatin remodeling gene AT-rich interactive domain-containing protein 1A suppresses gastric cancer cell proliferation by targeting PIK3CA and PDK1Oncotarget2016729461274614127323812
- DienstmannRRodonJSerraVTaberneroJPicking the point of inhibition: a comparative review of PI3K/AKT/mTOR pathway inhibitorsMol Cancer Ther20141351021103124748656
- CourtneyKDCorcoranRBEngelmanJAThe PI3K pathway as drug target in human cancerJ Clin Oncol20102861075108320085938
- TranPNguyenCKlempnerSJTargeting the phosphatidylinositol-3-kinase pathway in gastric cancer: can omics improve outcomes?Int Neurourol J201620Suppl 2S131S14027915478
- RamanathanRKMcDonoughSLKenneckeHFPhase 2 study of MK-2206, an allosteric inhibitor of AKT, as second-line therapy for advanced gastric and gastroesophageal junction cancer: A SWOG cooperative group trial (S1005)Cancer2015121132193219725827820
- GajriaDChandarlapatySHER2-amplified breast cancer: mechanisms of trastuzumab resistance and novel targeted therapiesExpert Rev Anticancer Ther201111226327521342044
- KatagiriANakayamaKRahmanMTLoss of ARID1A expression is related to shorter progression-free survival and chemoresistance in ovarian clear cell carcinomaMod Pathol201225228228822101352
- LyuCZhangYZhouXLangJARID1A gene silencing reduces the sensitivity of ovarian clear cell carcinoma to cisplatinExp Ther Med20161264067407128105136