
Abstract
The design of novel pharmacologic agents as well as their approval for sale in markets all over the world is a tedious and pricey process. Inevitably, oncologic patients commonly experience unwanted effects of new anticancer drugs, while the acquisition of clinical experience for these drugs is largely based on doctor–patient partnership which is not always effective. The repositioning of marketed non-antineoplastic drugs that hopefully exhibit anticancer properties into the field of oncology is a challenging option that gains ground and attracts preclinical and clinical research in an effort to override all these hindrances and minimize the risk for reduced efficacy and/or personalized toxicity. This review aims to present the anticancer properties of drugs used for the management of hypercholesterolemia. A global view of the antitumorigenicity of all marketed antihypercholesterolemic drugs is of major importance, given that atherosclerosis, which is etiologically linked to hypercholesterolemia, is a leading worldwide cause of morbidity and mortality, while hypercholesterolemia and tumorigenesis are known to be interrelated. In vitro, in vivo and clinical literature data accumulated so far outline the mechanistic basis of the antitumor function of these agents and how they could find application at the clinical setting.
Introduction
Many pharmacologic agents that are commercially available find application in multiple fields in everyday clinical practice. Characteristically, the chemotherapeutic agent mitomycin C exhibits multilevel therapeutic exploitability functioning, both for killing cancer cells and regulating wound healing.Citation1,Citation2 Importantly, a growing body of evidence suggests that several pharmacologic agents could be “repositioned” or “repurposed” in the field of oncology, owing to their interference with signaling pathways which are crucial for tumorigenesis. This notion is experimentally (at the preclinical level, clinical level or even at both levels) corroborated for marketed agents currently used for the management of different pathologic entities ranging from pain,Citation3 gastrointestinal disorders,Citation4 depression,Citation5 hypercholesterolemiaCitation6 and diabetes mellitus type IICitation7,Citation8 to human immunodeficiency virusCitation9,Citation10 and parasitic infection.Citation11,Citation12
Herein, for the first time, we concurrently review the antitumor activity of the widely prescribed marketed antihypercholesterolemic drugs, statins and ezetimibe. Notably, although a concernable amount of theoretical evidence regarding the anticancer activity of statins in different types of malignancies has already been published,Citation13,Citation14 to the best of our knowledge, so far, there has been no published work summarizing the anticancer function of both statins and ezetimibe. The fact that hypercholesterolemia is etiologically linked to a leading cause of morbidity and mortality worldwide, that is, atherosclerosisCitation15 and that high blood cholesterol levels are interrelated with tumorigenesisCitation16 underlines the impetus for presenting the anti-tumorigenicity of this category of drugs. Bile acid sequestrants (cholestyramine, colestipol and colesevelam) are also used for the management of hypercholesterolemia. Yet, to the best of our knowledge, no evidence has been provided so far suggesting any anticancer activity for these drugs. The objective of this article is to prompt further basic and/or clinical research regarding the repositioning of the aforementioned or other marketed drugs in cancer therapeutics by presenting the antitumorigenicity of non-vitamin antilipidemic agents that are either synthetic (atorvastatin, fluvastatin, rosuvastatin and ezetimibe) or fermentation derived (lovastatin, simvastatin and pravastatin). Of note, vitamin B3 (niacin), which is known to reduce low-density lipoprotein (LDL)-cholesterol levels,Citation17 participates to antitumor pathways upon its in vivo conversion into nicotinamide.Citation18,Citation19 However, only the antitumor functions of non-vitamin antilipidemic are presented here.
Current clinical use of statins and ezetimibe
Statins are LDL-cholesterol-lowering agents. They serve as 5-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase inhibitors, thereby blocking the rate-limiting step in mevalonate metabolic pathway, which is responsible for the biosynthesis of cholesterol and isoprenoids, exemplified by geranylgeranyl pyrophosphate (GGPP).Citation20 Statins are currently commonly prescribed both to prevent the onset of cardiovascular disease (CVD) as well as to patients who already suffer from CVD, since in the latter case, they reduce the mortality rates.Citation21
Like statins, ezetimibe is an LDL-cholesterol-lowering drug. Yet, it works through a different mechanism to prevent the exogenous uptake of cholesterol as well as its biliary absorption. Hence, until now, ezetimibe finds clinical application as an antihypercholesterolemic agent.Citation22,Citation23 It is used as a second-line option in hypercholesterolemic individuals who do not satisfactorily respond to statin therapy, either alone or in combination with a statin.
Possible therapeutic repurposing
Overview
Hypercholesterolemia has been associated with accelerated tumor growth,Citation16 while statins interfere with the pathways modulating molecules that are crucial for tumorigenesis; the fact that prenylation of the cancer-associated G-protein superfamily members (eg, RAS isoforms and RhoA) is required for their activationCitation20 has instigated researchers to explore the antitumor activity of statins. Although there are many reports indicating that the anticancer function(s) of statins can be reversed by mevalonate pathway intermediates,Citation24,Citation25 it becomes increasingly clear that statins may actually employ far more sophisticated mechanism of action to affect tumor biology.Citation26–Citation30
In striking contrast to statins which have long been known to target oncoproteins (eg, RAS), ezetimibe targets a molecule that is not an oncoprotein. In fact, ezetimibe blocks the intestinal sterol influx transporter Niemann–Pick C1-Like 1 (NPC1L1).Citation22 To the best of our knowledge, however, no evidence has been provided that ezetimibe’s anticancer mechanism of action is NPC1L1-dependent. As it is presented immediately below, ezetimibe’s function goes beyond NPC1L1 regulation to interfere with tumor microvascularization,Citation22 as well. These data underline the fact that the pharmacologic agents that either target the oncoproteins or interfere with different pathways that involve cancer-associated proteins merit research interest as possible anticancer drugs.
Preclinical data
Simvastatin, pravastatin and rosuvastatin
Simvastatin exerts antiproliferative effects on non-small cell lung cancer (NSCLC) A549 cells by impairing the activation of ERK.Citation25,Citation31 These effects are counteracted by fueling the mevalonate pathway with the terpenoid precursor, farnesyl pyrophosphate.Citation25 Downregulation of transforming growth factor-β receptor II (TGF-βRII) has also been documented as a molecular event that is associated with simvastatin-dependent inhibition of ERK.Citation25 This is of major importance, since deregulated TGF-β signaling is a well-known culprit in lung carcinogenesis.Citation32 Other mechanism linked to the simvastatin-mediated deterioration of NSCLC cell viability is the pro-oxidant activity of this drugCitation33 as well as its ability to decrease the expression of antiapoptotic proteins (XIAP, Bcl-2) and to stimulate the expression of proapoptotic ones, including Bax and p53.Citation31,Citation34 Moreover, simvastatin acts in a cytostatic fashion through negating the expression of cell cycle-regulatory molecules (cyclin D1, cyclin-dependent kinase 4 [CDK4]) governing the G1/S phase transition in NSCLC A549 and NCI-H460 cells.Citation34,Citation35 Importantly, experiments indicate that treatment of different cancer cell lines with simvastatin regulates Bcl-2/Bax expression and triggers apoptosis, but leaves the human normal small airway epithelial cells unaffected.Citation36
In highly metastatic triple-negative MDA-MB-231 breast cancer cells, simvastatin functions in an antigrowth manner to inhibit NFkB/PTEN- and NFkB/BclXL-dependent routes.Citation37 Also, the metastatic potency of these cells is antagonized by simvastatin or lovastatin that upregulate the expression of p53 which, in turn, represses the expression of the adhesion molecule, CD44.Citation38 Another mechanism by which simvastatin decreases the migratory and invasive potential of these cells is by interfering with a Rho GTPase/Hippo-YAP pathway, which modulates the expression of the oncogenic protein CD168 (hyaluronan-mediated motility receptor) whose gene promoter is responsive to the transcription coactivator YAP and TEAD family transcription factors.Citation39 In breast cancer cells which are positive for HER2, simvastatin seems to be a promising anticancer agent that shuts off the expression of HER2 through upregulating its negative transcriptional modulator, namely, PEA3.Citation24 In MCF-7 breast cancer cells (these cells are ER-positive and HER2-negative), simvastatin and fluvastatin stimulate apoptosis through positively modulating the expression of inducible nitric oxide synthase (iNOS) at the transcriptional level.Citation40 In ER-negative breast cancer cells, simvastatin displays synergistic antigrowth function with the cardiac glycoside, digitoxin.Citation41
Experimental evidence pinpoints to an interrelation between the retinoblastoma protein (RB) status and the tamoxifen resistance phenotype. In fact, analyses have shown that in a molecular background where the tumor-suppressive retinoblastoma protein (pRb) axis is dysfunctional (eg, in SaOS2 osteosarcoma cells), the interference with the assembly of the minichromosome maintenance (MCM) complex and DNA replication causes more extensive DNA damages than in the presence of functional pRb (eg, in U2OS osteosarcoma cells).Citation42 Besides, premenopausal ER-positive breast cancer patients carrying a functional pRb are benefited from adjuvant tamoxifen treatment, in striking contrast to premenopausal ER-positive breast cancer patients with nonfunctional pRb, who do not experience any benefit.Citation43 Simvastatin negatively affects MCM7 expression, resulting in the elevation of the DNA damage marker, γH2AX. Remarkably, the combined treatment of tamoxifen-resistant human breast cancer cells (these cells express pRb at lower levels in comparison to their wild-type counterparts) with simvastatin and tamoxifen evokes a genotoxic signal, which is efficient to trigger apoptosis.Citation42
In prostate cancer cells characterized by androgen-independent growth, simvastatin displays synergistic toxicity with another mevalonate pathway inhibitor, the bisphosphonate alendronate. Unlike statins that deplete isoprenoid biosynthetic route products by inhibiting HMG-CoA reductase, alendronate and other bisphosphonates inhibit the mevalonate pathway at the level of farnesyl diphosphate synthase by virtue of molecular mimicry. Hence, at low doses, simvastatin along with alendronate decreases the viability of these cells. This event is accompanied by the stimulation of the c-JUN and the induction of apoptosis. However, this is not the case in androgen-dependent prostate cancer cells, where no such anticancer synergy is observed.Citation44 When prostate cancer cells are incubated with conditioned medium obtained from mouse calvaria, insulin-like growth factor-1 receptor (IGF1R) is upregulated. Simvastatin abolishes this induction of IGF1R. Given that this receptor mediates the prosurvival effects of IGF-1 which is released by the bone matrix, this molecular event is of pivotal importance. Moreover, simvastatin acts in a synergistic manner with the IGF1R antagonist NVP-AEW541 to trigger apoptosis in prostate cancer cells. These findings indicate the value of simvastatin in the modulation of the dynamic interactions between prostate cancer cells and bone microenvironment in prostate cancer skeletal metastases.Citation45 Another antimetastatic property of simvastatin is its ability to disrupt the interaction among the adhesion molecules expressed on prostate cancer cells (avβ3 integrin) and vascular endothelial cells (ICAM-1) as well as to preserve the endothelial barrier integrity.Citation46
Simvastatin exhibits cytostatic effects (G0/G1 arrest) in a peroxisome proliferator-activated receptor-γ (PPAR-γ)-dependent manner, as well as antimigratory and anti-invasive properties in bladder cancer cells. Further, in these cells, simvastatin controls the expression of epithelial-to-mesenchymal transition (EMT)-associated molecules, exemplified by E-cadherin, which is elevated, and β-catenin, which is decreased upon exposure to simvastatin.Citation47
In leiomyoma cells, the ability of simvastatin to trigger apoptosis has been attributed to the targeting of calcium-permeable voltage-gated channels,Citation48 while in renal cancer cells, the proapoptotic and antimetastatic functions of this drug have been linked to perturbed mammalian target of rapamycin (mTOR), JAK2/STAT3 and ERK signaling. Colon cancer cell viability is reduced by simvastatin through the downregulation of the prosurvival factors Bcl-xL and Bcl-2 and by the suppression of the expression of the apoptosis inhibitors, cIAP1 and cFLIP.Citation49 In a seemingly paradoxical manner, the sustained activation of ERK by simvastatin is required for drug-induced apoptosis in HT-29 human colon cancer cells, whereas, in contrast, IGF-1-triggered ERK/Akt activation is counteracted by simvastatin that stimulates apoptosis.Citation50
Further, simvastatin increases the cytotoxic effects of irradiation combined with the anti-epidermal growth factor receptor (anti-EGFR) antibody cetuximab in head and neck squamous cell carcinoma (HNSCC) cells. Experimental data indicate that this antitumor activity is mechanistically associated with the suppression of ERK1/2-, STAT3- and AKT-dependent pathways and the induction of apoptotic cell death.Citation51 In human lymphoma and melanoma cells, the antitumor function of simvastatin is attributed both to its ability to sensitize to bortezomib-mediated apoptosis or to induce apoptosis, respectively, as well as to its ability to impinge on the expression of CDK inhibitors (p21 Waf1/Cip1, p27 Kip1), thereby halting cell cycle progression.Citation52,Citation53 In melanoma cells, simvastatin as well as lovastatin and a statin that is not used at the clinical setting (mevastatin) are known to exhibit antiproliferative and anti-invasive properties.Citation54 On the other hand, in multiple myeloma cells, simvastatin can induce both apoptosis and necrosis.Citation55 Also, simvastatin enhances the chemosensitivity of multiple myeloma cells by disrupting an HMG-CoA/GGPP/Rho/ROCK pathway, which confers cell adhesion-mediated drug resistance (CAM-DR) to them.Citation56
A mechanism by which simvastatin triggers apoptosis in SH-SY5Y neuroblastoma cells is through causing depletion of dolichol phosphate, which is produced throughout the mevalonate pathway. This isoprenoid compound serves as a sugar donor in the multistep process of protein N-glycosylation in the endoplasmic reticulum (ER). Impaired protein N-glycosylation results in the activation of the unfolded protein response (UPR)-related molecules BiP and CHOP, as well as in cellular stress and apoptotic death. Of note, this proapoptotic effect of simvastatin is dolichol-reversible. In addition, in neuroblastoma and rhabdomyosarcoma cells, simvastatin-mediated dolichol depletion precludes the full glycosylation of ABCB1 transporter, a protein that has been linked to the phenomenon of multidrug resistance in cancer cells. Also, simvastatin causes a drop in the transportation rates of ABCB1 in neuroblastoma cells.Citation57
Interference of simvastatin with the Rho GTPase/Hippo-YAP/CD168 pathway is not only documented in vitro as mentioned above,Citation39 but also in human breast cancer xenografts. The importance of this finding is underlined by the fact that CD168 along with YAP is found to be overexpressed in clinical specimens from patients with invasive breast cancer.Citation39 Also, in MDA-MB-231 xenografts, simvastatin prevents the occurrence of bone metastasis, and this has been attributed to an inverse relationship between the expression of CD44 and p53, which is associated with drug administration.Citation38 In immunodeficient mouse strains xenografted with tamoxifen-resistant breast cancer cells, simvastatin acts as a negative modulator of the replication licensing factor (RLF), MCM7. Simvastatin restores the sensitivity of tamoxifen-resistant tumors to tamoxifen and suppresses tumor growth.Citation42 In animal models of triple-negative breast cancer (TNBC), simvastatin has been found to engage a FOXO3a-mediated pathway to decrease the metastatic potency of malignant cells.Citation58 Another notable in vivo antitumor activity of simvastatin is that it inhibits lung cancer metastasis to the bones in murine xenografts.Citation31 Moreover, simvastatin exhibits synergistic anticancer effects in mouse HNSCC xenografts, in conjunction with radiotherapy and cetuximab.Citation51
The antitumor properties of simvastatin are also evident in human renal carcinoma xenografts.Citation59 Liposomal simvastatin administration yielded encouraging results in mice with melanoma. Mechanistically, the observed antitumor effect was demonstrated to be related to the ability of simvastatin to oppose the pro-oxidant function of tumor-associated macrophages (TAMs).Citation60 This is also the case of pravastatin, which, upon administration in a liposomal formulation to melanoma-bearing mice, exhibits anti-inflammatory properties. Moreover, liposomal pravastatin was reported to upregulate major histocompatibility complex (MHC) class I expression in tumors, thereby possibly positively influencing tumor immunosurveillance.Citation61
Simvastatin opposes the development of tumors in the mouse model of colitis-associated colon cancer and exhibits proapoptotic and antiangiogenic effects in colon cancer xenografts.Citation49 In addition, simvastatin acts in synergy with bevacizumab to inhibit the growth of human colorectal cancer (CRC) xenografts and their ability to give rise to metastases.Citation62 More recently, simvastatin has been reported to serve as an antiangiogenic factor in CRC in vivo by regulating a HER2/VEGF signaling pathway.Citation63
Moreover, simvastatin prevents virus-driven lymphomagenesis in immunodeficient miceCitation64 and functions to impede mammary tumorigenesis in rodents induced by irradiation at the earliest stages.Citation65 In the bone marrow of immunodeficient mice, simvastatin reduces the clonogenic capacity of human myeloblastic leukemia cells.Citation66 In rhabdomyosarcoma xenografts, clinically relevant doses of simvastatin inhibit tumor growth, while in xenograft models of neuroblastoma and rhabdomyosarcoma, simvastatin displays synergistic proapoptotic effects with cyclophosphamide.Citation57
Aside from simvastatin, rosuvastatin and pravastatin also exhibit antitumor function. Rosuvastatin, has been reported to suppress the activity of arginase (an urea cycle enzyme) and to reduce the levels of polyamines, both in the serum and tumor tissue in mouse models of breast cancer.Citation67 These data add another level of complexity to statins’ antitumor mechanism of action, given that arginase, its substrate arginine and polyamines participate in perplexed signaling pathways and are involved in tumor biology.Citation68–Citation71 On the other hand, pravastatin displays remarkable chemopreventive efficacy in rodent models of chemical-induced colon carcinogenesis.Citation72,Citation73
Lovastatin
Lovastatin stimulates caspase-dependent apoptotic death of melanoma cells in a GGPP-reversible manner.Citation74 Aside from melanoma cells, lovastatin induces apoptosis in a wide range of human cancer cells, including medulloblastoma and malignant mesothelioma cells.Citation75–Citation80
Lovastatin triggers apoptosis in different lung cancer cell lines by affecting Bax levels and stimulating p53.Citation81 Further, lovastatin overrides resistance to gefitinib-induced apoptosis in NSCLC cells carrying mutant K-RAS by positively and negatively modulating the expression of Bax and Bcl-2, respectively,Citation82 while it inhibits NSCLC cell adhesion and migration through depleting lipid rafts from cholesterol.Citation83 Of note, the RLF MCM2 has been identified as a target of putative therapeutic activity of lovastatin in human NSCLC. Actually, exposure of NSCLC cells to lovastatin results in the decrease of MCM2 levels in a JNK-dependent fashion.Citation84
In colon cancer cell lines, lovastatin potentiates the cytotoxic effects of commonly used chemotherapeutics such as cisplatin and 5-fluorouracil (5-FU) by upregulating the expression of Bax and suppressing Bcl-2.Citation85 Celecoxib-induced apoptosis in colon cancer cells is increased by lovastatin,Citation86 while in colon cancer cells, NSCLC cells as well as in SCC9 cells, this statin potentiates the cytotoxicity of gefitinib.Citation87
Lovastatin along with simvastatin induces cell cycle arrest and stimulates apoptosis in prostate cancer cells by affecting the expression of a series of cell cycle regulators (D-type cyclins, pRb and the CDK inhibitors, p21 Waf1/Cip1 and p27 Kip1) and through activating a RhoA/caspase-dependent pathway.Citation88 In these cells, lovastatin has also been reported to decrease E2F1 levels to trigger apoptosis.Citation89 Antitumor properties of lovastatin have also been reported in glioblastoma cells, where lovastatin exhibits antiproliferative and anti-invasive properties by inhibiting small GTPase (Rac1 and H-Ras) isoprenylation.Citation90
Another mechanism by which lovastatin indirectly displays anticancer function is through prevention of the adenosine triphosphate (ATP)-dependent extrusion of the antileukemic drug imatinib from the cytosol of CD34 antigen-expressing chronic myeloid leukemia (CML) cells by multidrug resistant transporters, namely, ABCG2 and ABCB1. These effects of lovastatin are cholesterol-dependent.Citation91 In human B lymphoma cells, the antiproliferative effects of lovastatin have been attributed to its antioxidant potency as well as to its ability to suppress the expression of the calcium-permeable channel transient receptor potential canonical 6 (TRPC6).Citation92
An antibody microarray analysis showed that the expression of a set of 37 proteins is altered in response to lovastatin in MDA-MB-231 triple-negative aggressive breast cancer cells. Remarkably, the apoptosis-related proteins, death receptor 3 (DR3) and caspase-7, as well as the tumor-suppressor, p57Kip2, were found to be upregulated among other proteins, whereas the cell-cycle regulatory protein CDK4 and apoptosis-inducing factor (AIF) were downregulated. Transglutaminase II (a mediator of N-cadherin switching) and hypoxia-inducible factor 1α (HIF-1α) that are associated with the EMT phenomenon and matrix metalloproteinase (MMP)-7, which is known to be related with metastasis, were also reported to be negatively modulated by lovastatin in these cells.Citation93 In leukemia cells of human origin, lovastatin displays synergistic effects with the clinically used antimetabolite, cytarabine.Citation94
Intriguingly, lovastatin dampens the dimerization of the cancer-associated molecule EGFR and downstream Akt-dependent signaling in a GGPP-reversible fashion. This leads to the suppression of global protein synthesis in squamous cell carcinoma cells.Citation95 In addition, lovastatin may prevent tumorigenesis, which takes place on the ground of chronic inflammation, given that in vitro experimentation showed that this statin markedly prevents the oxidative damage of DNA evoked by stimulated phagocytes.Citation96 Also, Ames testing suggests that lovastatin as well as atorvastatin may exhibit chemopreventive activity, since both these drugs inhibit the action of mutagens.Citation97
In vivo, lovastatin is known to enhance the antimelanoma effects of cisplatin at the preclinical levelCitation98 and increase the anticancer function of tumor necrosis factor-αCitation99 and doxorubicin in different mouse tumor models.Citation99 In a mouse model of ascites tumor, lovastatin triggers apoptosis in Dalton’s lymphoma ascitic cells,Citation100 while in rats, this drug halts metastatic dissemination of fibrosarcoma and lymphoma cells.Citation101,Citation102 Besides, lovastatin reduces the metastatic potential of mouse mammary tumors and melanomas.Citation103,Citation104 Further, in a manner that is unrelated to K-RAS mutational status or expression, lovastatin inhibits nitrosamine-induced tumorigenesis in murine lungs.Citation105 Of note, in different murine tumor models, lovastatin is known to protect the myocardium from doxorubicin-induced cell damage.Citation99 In tumors of HER2/neu transgenes, lovastatin decreases the population of macrophages exhibiting an M2-like phenotype; a cell population which is known to control various aspects of carcinogenesis. In contrast, the population of effector T lymphocytes increases upon lovastatin treatment. At the same time, tumors get more oxygen and they become more accessible to anticancer drugs by virtue of lovastatin’s property to trigger vascular remodeling in tumors.Citation27
Atorvastatin
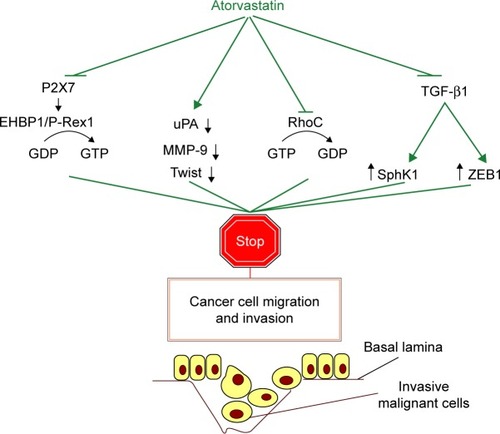
In vitro experimentation using human pancreatic cancer cells demonstrated that atorvastatin displays antitumor synergy with the FTI tipifarnib and the cyclooxygenase-2 (COX-2) inhibitor celecoxib.Citation106 In HNSCC cells, atorvastatin displays inhibitory activity on RhoC, a small GTPase and metastasis marker in HNSCC that has to be prenylated in order to be functional. Atorvastatin impairs the proliferation of HNSCC cells and reduces their motility and invasiveness.Citation107 In A549 cells, a TGF-β1/sphingosine kinase 1 (SphK1) axis has been recently characterized. Inhibition of this axis by atorvastatin is associated with the partial suppression of the EMT phenomenon and, hence, mitigation of the metastatic potency of these NSCLC cells. In addition, atorvastatin prevents the TGF-β1–mediated induction of the EMT-promoting transcription factor, ZEB1. This molecular event is also associated with the suppression of the migratory activity of A549 cells by atorvastatin.Citation108
EH domain-binding protein 1 (EHBP1) has been found to serve as a mediator of atorvastatin’s purinergic (P2X7), signaling-dependent, anti-invasive activity in prostate cancer cells, which are positive for the tumor suppressor protein PTEN. In these cells, EHBP1 physically associates with the invasiveness-related guanine nucleotide exchange factor (GEF), P-Rex1. In other cancer cell lines, statins also suppress invasive growth through interfering with a P2X7-dependent pathway that modulates the nuclear phospho-Akt levels. The importance of these findings is substantiated by population genetic data suggesting that a loss of function allele coding for a P2X7 receptor variant is correlated with nonaggressive cancer, in contrast to the common allele which is associated with aggressiveness.Citation109 Mechanistically, the antimigratory/anti-invasive function of atorvastatin in breast cancer principally involves suppression of the expression of urokinase-type plasminogen activator (uPA), MMP-9 and the basic helix–loop–helix (bHLH) transcription factor, Twist.Citation29 The different signaling routes used by atorvastatin to suppress the migratory and invasive potential of malignant cells in vitro are collectively depicted in . Importantly, atorvastatin dampens the expression of the RLF MCM7 and induces genomic damage and apoptosis in tumor cells in a pRb-deficient background to a greater extent than in the presence of pRb in a similar way as simvastatin does, both in an in vitro and an in vivo setting.Citation6
Figure 1 Signaling pathways mechanistically involved in atorvastatin’s ability to inhibit the migratory and invasive potential of different types of cancer cells in vitro.
Abbreviations: ATP, adenosine triphosphate; bHLH, basic helix–loop–helix; EMT, epithelial-to-mesenchymal transition; GDP, guanosine diphosphate; GEF, guanine-nucleotide exchange factor; GTP, guanosine triphosphate; HNSCC, head and neck squamous cell carcinoma; MMP, matrix metalloproteinase; NSCLC, non-small cell lung cancer; PTEN, phosphate and tensin homolog deleted on chromosome 10; SphK1, sphingosine kinase 1; uPA, urokinase-type plasminogen activator.
In Pankras/p53 mice which are engineered to carry KrasG12D and p53R172 mutations, thereby genetically and phenotypically resembling the human pancreatic carcinogenesis, nutritional supplementation with atorvastatin resulted in a decrease of the proliferation index of the tumors and their shrinkage. In fact, treatment with atorvastatin or the FTI tipifarnib (R115777) prevented protein prenylation and halted the growth of malignant cells. Importantly, atorvastatin decreased neutrophil infiltration in chronic pancreatitis near the tumor in transgenes.Citation28 The latter finding is important given, the interrelation between inflammation and tumorigenesis.Citation110,Citation111
In addition, analysis in pancreatic tumor xenografts in mice demonstrated that atorvastatin displays antitumor synergy with the COX-2 inhibitor celecoxib and tipifarnib.Citation106 In rats, combined administration of atorvastatin with either naproxen or sulindac, two marketed nonsteroidal anti-inflammatory drugs (NSAIDs), could markedly inhibit inflammatory signaling and lower the incidence of colon adenocarcinomas.Citation112 In line with the latter finding, atorvastatin acts in synergism with the COX-2 inhibitor celecoxib to suppress tumorigenesis in a prostate cancer xenograft mouse modelCitation113 or the progression of the androgen-dependent pattern of prostate tumor growth to androgen independenceCitation114 as well as the formation of intestinal polyps in the mouse familial adenomatous polyposis (FAP) model APCmin.Citation115
Further, there are reports indicating the in vivo antitumor function of atorvastatin in head and neck cancer, MYC-induced lymphoma and breast cancer. In mice with head and neck cancer, atorvastatin suppresses tumor angiogenesis and the occurrence of distant metastases.Citation107 The antitumor activity of atorvastatin in murine conditional transgenes for modeling MYC-driven lymphomagenesis is mechanistically associated with the suppression of the Ras/ERK/MYC axis upon the pharmacologic inhibition of HMG-CoA reductase.Citation116 In an orthotopic model of breast cancer metastasis, a micelle-based nanosystem for the delivery of atorvastatin inhibited distant metastases of breast cancer cells.Citation29
Fluvastatin
The fluorinated compound fluvastatin displays selective toxicity toward malignant rat glial cells, but it does not affect normal neurons. Its multilevel antitumor activity is molecularly associated with the stimulation of the JNK1/2 axis, the suppression of ERK1/2-dependent signaling, as well as the impaired secretion of VEGF and MMP-9.Citation117 Paradoxically, statins are not only involved in the inhibition of GTPase isoprenylation, but also function as glycosylation inhibitors to suppress tumor growth. In fact, fluvastatin inhibits glycosylation of the class III receptor tyrosine kinase FMS-like tyrosine kinase-3 (FLT3), which commonly harbors activating mutations in acute myeloid leukemia (AML) and cannot be successfully targeted. This molecular event is associated with hampered MAPK and AKT activation and killing of cells expressing mutant FLT3 (FLT3/ITD).Citation26 This is a good example of how a class of pharmacologic agents somehow interferes with more than one type of protein’s posttranslational modification to achieve their pleiotropic biologic effects. Regarding the antitumor activity of statins in hematologic malignancies, evidence has been provided for the combined antileukemic effects of fluvastatin with all-trans retinoic acid (ATRA).Citation118
In renal cancer cells, fluvastatin’s antitumor activity has been suggested to be mediated by the derepression of the function of a negative modulator of translation initiation, namely, programmed cell death 4 (PDCD4), through the inhibition of the mTOR.Citation119 Interestingly, fluvastatin as well as lovastatin reduce EGF-stimulated human pancreatic cancer cell invasion by the inhibition of RhoA isoprenylation.Citation120 Of note, the stimulation of the production of the antitumor cytokine interleukin-18 by human monocytes is thought to account for an antitumor mechanism of action of all the aforementioned statins (fluvastatin, simvastatin and pravastatin).Citation121
Fluvastatin is known to confer a survival benefit to leukemic mice with FLT3/ITD.Citation26 In addition, fluvastatin negatively influences the ability of mouse renal cancer cells to give rise to metastases in the lungs of mice.Citation122
Preclinical data
Ezetimibe
A study carried out by Solomon et al indicates that not only statins, but also ezetimibe exhibit antitumor activity. Actually, ezetimibe was reported to act independently, but additively with the application of a low fat/no cholesterol (LFNC) diet in retarding tumor growth in severe combined immunodeficiency (SCID) mice implanted with human prostate tumors. Moreover, it was demonstrated that ezetimibe and LFNC exhibit additive, independent inhibitory effects on LNCaP cell proliferation and viability, as well as on tumor microvascularization through upregulating the expression of thrombospondin 1 (TSP-1), a well-established antiangiogenic factor.Citation22 More experimentation is required to shed light on whether ezetimibe can halt the growth of other cancer cell types as well.
Clinical data
Data from Phase I and II clinical trials support that monthly administration of lovastatin at relatively high doses (25 mg/kg/day) for a week followed by a 3-week pause in patients with prostate cancer or central nervous system (CNS) tumors causes, in most cases, ubiquinone-reversible myopathy, but yields only minor and partial responses.Citation123,Citation124 Almost no objective responses were reported in patients with advanced gastric adenocarcinoma who received 7-day courses of lovastatin at a dose of 35 mg/kg/day every 28 days.Citation125 However, a clinically relevant dose of pravastatin (40 mg/day) was found to extend life expectancy by two times in patients with advanced hepatocellular carcinoma (HCC) who had previously undergone transcatheter arterial embolization (TAE) followed by 5-FU administration, without causing myalgia, indicating its usefulness in adjuvant treatment.Citation126 In line with this, chemoembolization is more efficacious in improving survival in HCC patients when it is combined with pravastatin at a dose of 20–40 mg/day.Citation127
A Phase II clinical study yielded promising results with clinically relevant doses of simvastatin (40 mg/day) combined with the conventional chemotherapeutic scheme FOLFIRI (folinic acid, 5-FU and irinotecan) in patients with metastatic CRC.Citation128 Of note, certain monoclonal antibodies targeting the EGFR, exemplified by cetuximab, have been shown to be of clinical benefit, provided that KRAS does not harbor mutations. Hence, it has been hypothesized that simvastatin, which interferes with RAS posttranslational modification (prenylation), might restore the sensitivity to cetuximab. Unfortunately, this hypothesis is not valid, as reported by a Phase II clinical study in previously treated, metastatic CRC patients. Specifically, only 4 out of 18 patients who were enrolled into the study experienced disease stabilization at 12.5 weeks after cetuximab was first administered. This does not suggest any clinical applicability of simvastatin as a KRAS modulator and, hence, an agent that would restore sensitivity to cetuximab in metastatic CRC patients carrying KRAS mutations.Citation129 This also holds true for another anti-EGFR monoclonal antibody, namely, panitumumab, as indicated by another Phase II clinical study.Citation130 An analysis retrospectively assessing the data stemming from the CAIRO2 Phase III clinical trial reported that the usage of statins does not beneficially influence either progression-free survival (PFS) or overall survival of metastatic CRC patients with the KRAS mutant gene who received chemotherapeutic agents in conjunction with the monoclonal antibodies, cetuximab and bevacizumab.Citation131 On the other hand, in Danish individuals suffering from different types of cancer, the use of statin reduces the rates of malignancy-associated death, as shown by a retrospective analysis.Citation132 Considering the latter analysis, however, some phenomena including the preferential prescription of statins to nonsmokers by some general practitioners or other comorbidities in statin users (eg, CVD) may have led to data misinterpretation and/or incorrect inferences.Citation133,Citation134
Hopefully, certain statins, in a manner that depends upon their physicochemical properties (ie, their hydrophobicity), increase the PFS rates of patients suffering from the most life-threatening type of primary breast cancer; inflammatory breast cancer, as demonstrated in an observational study.Citation135 This is in line with the results from a Phase II study where it was found that the preoperative administration of high doses of atorvastatin manages to decrease the proliferation of tumor cells that express HMG-CoA reductase in women with primary invasive breast cancer.Citation136 In window trials enrolling breast cancer patients, comparison of the transcriptome before and after treatment with high dose (80 mg/day) of atorvastatin for 14 days prior to surgery has shown that the mRNA levels of molecules associated with immune response, apoptosis (GADD45B) and MAPK signaling (eg, the DUSP1 phosphatase) are statistically significantly affected,Citation137 while this atorvastatin dosage regimen also modulates cyclin D1 and the oncosuppressor p27Kip1 proteins in breast cancer, negatively and positively, respectively.Citation138 Remarkably, a pilot perioperative study demonstrated that fluvastatin at clinically attainable dosages (20 or 80 mg/day) exhibits potent chemopreventive properties in high-grade early-stage breast cancer patients.Citation139
In a cohort of white postmenopausal women, the self-reported consumption of simvastatin, lovastatin as well as other lipophilic statins over a follow-up period of 10.5 years was associated with an elevated risk for non-melanoma skin cancer (NMSC), irrespective of other factors including sunlight exposure, vitamin D intake, smoking and body mass index (BMI). Therefore, researchers did not exclude the scenario of a protumorigenic – possibly immunomodulatory – activity of statins and recommended that subjects with a family history of NMSC or those who are carcinogenesis prone due to their skin type should avoid the consumption of the aforementioned statins.Citation140
Intriguingly, the results of other previous studies regarding the putative link between NMSC and statin usage are discrepant, both incriminating this category of antihypercholesterolemic agents for NMSCCitation141–Citation144 and pinpointing to a protective role of statins against the development of cancerCitation145–Citation147 or even not suggesting any statistically significant relationship between them.Citation148–Citation151 In accordance with the latter scenario, patients suffering from metastatic CRC who received chemotherapy and then were enrolled into a Phase III study in South Korea did not experience any clinical benefit (PFS) when 40 mg of simvastatin were added to a XELIRI/FOLFIRI chemotherapy regimen.Citation152 Based on the statistics on statin prescription (simvastatin ranked first in statin prescription in the UKCitation153) and the fact that atorvastatin is one of the most extensively studded statins as an antitumor agent at the clinical basis, summarizes selected completed or active clinical trials evaluating the usage of these two antihypercholesterolemic agents in different types of malignancy or precancerous conditions.
Table 1 Selected active or completed clinical trials on simvastatin and atorvastatin for cancer therapeutics and/or cancer prevention
More clinical trials are necessary to evaluate the antitumor function of statins. The results from the clinical trials that have already been completed or are still ongoing (NCT02026583, NCT01992042) are, therefore, much awaited. Regarding ezetimibe, yet, no clinical evidence has been provided supporting its antitumor function. The relationship between ezetimibe and tumorigenesis is actually obscure.Citation154 A multinational study, namely, SEAS, investigating the occurrence of cancer in patients with aortic stenosis who received placebo compared to those who received simvastatin (40 mg daily) and ezetimibe (10 mg daily) reported that the latter are at higher risk for carcinogenesis.Citation155 However, no such association was reported in a subsequent study enrolling a SEAS trial cohort from the UK, Denmark and Scandinavia.Citation156 Clinical trials evaluating the possible anticancer therapeutic potency of ezetimibe in oncologic patients are necessary to enable its future repositioning in the field of oncology.
Considerations for using and repurposing antihypercholesterolemic drugs
In many studies, the anticancer function of statins on various cancer cell lines has been demonstrated, using them at clinically irrelevant doses.Citation157–Citation159 In other cases, however, statins display their anticancer properties at dosages that are clinically attainable. For instance, the IC50 (ie, 50% of growth suppression) of lovastatin in various cancer human cells is within the concentration range 0.3–2 μM.Citation160 According to data stemming from experiments in animals, 2–4 μM of lovastatin in serum is safe and toxicity becomes evident only at a concentration of 20–25 μM.Citation161 Clinical data indicate that in cancer patients, even high plasma concentrations of lovastatin (up to 12 mM) administered for a short period (≤4 days) are well tolerated.Citation159
Statins display additivity and synergism with conventional chemo drugs and monoclonal antibodies (eg, antimetabolites, cisplatin, doxorubicin and cetuximab).Citation85,Citation94,Citation98,Citation162–Citation164 In addition, there is accumulating evidence that statins act at low doses in synergism with NSAIDs as chemopreventive agents.Citation165 Moreover, owing to the phenomenon of biotransformation by cytochrome P450 (CYP) 3A4 (expressed in gut and liver), the concentrations of statins required to achieve their antitumor effects may be actually much lower than the concentrations used for in vitro experiments.Citation158 Hence, all these data open the possibility for the future use of statins as adjuvant agents for the treatment of cancer at dose schemes close to the current therapeutic anticholesterolemic dose.
Some people receiving statins experience myotoxicity and/or hepatotoxicity. In addition, coadministration of statins with certain drugs (including clarithromycin, erythromycin, azole antifungals and cyclosporine A) which inhibit CYP3A4 – the major statin-metabolizing CYP isoform – is prohibitive, since these agents increase the serum statin concentration, thereby exacerbating myopathy.Citation166,Citation167 Therefore, frequent monitoring of serum transaminases is highly recommendable, while statin-induced reduction of serum Q10 (a mevalonate pathway product) levels that has been associated with myopathy could be restored by ubiquinone supplementation.Citation168 Great caution should also be exercised with regard to those patients who receive medication that is incompatible with statins exemplified by cyclosporine and macrolide antibiotics that inhibit CYP3A4.Citation169,Citation170 Regarding statin myotoxicity, particular caution should be paid in case that these drugs will eventually be repositioned in the field of oncology since malignancy and myopathy have been interrelated, while myopathy may also be ascribed to steroid consumption by cancer patients.Citation171,Citation172 Conceivably, in cases of myopathic patients, the use of statins as antitumor drugs would not be recommended. A major concern is that subjects suffering from type II diabetes mellitus who consume atorvastatin are likely to develop atorvastatin-induced autoimmune myositis (AtorAIM). Hallmarks of AtorAIM are: 1) detection of anti-HMG-CoA reductase autoantibodies in the serum of patients, 2) abnormal creatine kinase (CK) levels even when the administration of atorvastatin is being discontinued and 3) responsiveness to the intravenous immunoglobin (IVIG) induction therapy, although AtorAIM patients do not respond to corticosteroids.Citation173
Since ezetimibe is devoid of the severe adverse effects of statins (eg, hepatotoxicity, myotoxicity) and has been proved to be safe and well tolerated, either in monotherapy or in combinational hypolipidemic therapy,Citation174–Citation176 it would be of great interest if future clinical trials would evaluate it as an antitumor agent. However, it should be noted that, ezetimibe in combination with cholesterol diet stimulates NFkB and local intestinal inflammation in guinea pigs.Citation177 Given the interrelation between inflammation and intestinal cancer,Citation178 more studies are needed to address whether ezetimibe could fuel intestinal carcinogenesis in humans under certain circumstances.
Conclusion and future perspectives
The putative usage of the pharmacologic agents discussed here, especially statins, in the field of oncology is not based on the same rationale as those for the mechanism of action of either chemotherapeuticsCitation179–Citation181 or targeted anticancer drugs.Citation182 Statins were first isolated and characterized as HMG-CoA reductase inhibitors in 1970s, while lovastatin was the first statin that gained Food and Drug Administration approval as an antihypercholesterolemic drug in 1987. Hence, researchers started to make efforts to target cholesterol synthesis as a blood cholesterol-lowering strategy, even though two decades ago, published dataCitation183 indicated an interrelation between the biosynthetic pathway of cholesterol and cellular transformation. In early 1990s, it was hypothesized that the blockage of mevalonate pathway could suppress cancer cell growth.Citation184 However, accruing experimental data presented above suggest that statins may be exploited as antitumor agents owing not solely to their isoprenoid biosynthesis-blocking property. Actually, statins are somehow molecularly “endowed” with pleiotropism, and therefore, they exhibit multilevel antitumor function(s) associated with the modulation of divergent signaling pathways, including cell life and death decisions,Citation31 cell-to-cell adhesion,Citation46 EMT,Citation32 protein N-glycosylation, the UPR,Citation57 DNA replication licensing,Citation42 TAMsCitation60 or even angiogenesis.Citation63 Thus, it becomes increasingly clear that statins act as versatile anticancer bulletins targeting pathways far beyond those that are known to link cholesterol and lipoprotein metabolism to carcinogenesis.Citation185 Versatility at the signaling and physiologic level also holds true for the blood cholesterol-lowering drug ezetimibe, which displays an antiangiogenic mode of antitumor action.Citation22
Additional clinical data are needed for statins to assess their antitumor potency. The results of ongoing clinical studies (eg, NCT01992042 and NCT02026583) are much awaited. It would also be very interesting to evaluate the antitumor properties of ezetimibe in oncologic patients, given that no such evidence has been provided for this antihypercholesterolemic agent so far, even though it is devoid of the major adverse effects associated with the use of statins.
A darling enterprise of the future is to delineate the mechanism(s) of the anticancer activity of non-antineoplastic marketed agents, which, in a similar manner to statins, are currently used for managing CVD – either prophylactically or therapeutically – given that CVD and cancer rank as the two leading causes of mortality globally.Citation186 Aspirin, similar to statins, is a widely prescribed NSAID with well-established antitumor properties.Citation9,Citation187 Caffeine is commonly combined with aspirin in commercially available formulations and is also well known to exhibit antitumor function.Citation188–Citation190 Serum proteomics data (Figure S1) indicate that at the clinical level, administration of aspirin combined with caffeine compared to aspirin alone may affect (either positively or negatively) the levels of proteins in human serum that have been linked to cancer biology, cancer prognosis/diagnosis and prediction of the efficacy of anticancer medication. These proteins are TTR,Citation191 RBP4,Citation192 HpCitation193 and ApoA1.Citation194 These hints would be very interesting to be experimentally validated in the future. Paradoxically enough, although several experimental data have been provided so far regarding the multilevel pharmacologic activity of aspirin or caffeine alone,Citation9,Citation195–Citation198 only less amount of data has been published regarding the effects of aspirin combined with caffeine.Citation199,Citation200 Most importantly, research so far has focused on the analgesic properties of this combinational scheme. To the best of our knowledge, there is no evidence addressing whether such a pharmacologic combination is associated with superior antitumor function in comparison to that of either agent alone. If the scenario for anticancer synergism holds true, it would be interesting to delineate the signaling pathway(s) that is/are involved in the synergistic effect. Intriguingly, caffeine is an inhibitor of the activity of the master DNA damage response (DDR) kinases ATM and ATR,Citation201 while an interplay between COX and ATR has been described. Actually, COX positively regulates ATR levels through a COX/PGE2/ERK/ATR axis in cancer cells and this pathway has been functionally associated with the acquisition of resistance to genotoxic agents.Citation202 Thus, a major future challenge would be to assess whether aspirin combined with caffeine displays antitumor synergism and which molecular circuits are affected in the presence of both these agents with regard to tumorigenesis. Besides, another scenario which is worthy of investigation is a putative synergism between an NSAID such as aspirin and atorvastatin or another statin against tumorigenesis, or even the possible antitumorigenic synergism of a triple combination of an NSAID, a statin and caffeine, given that atorvastatin and celecoxib are known to display anticancer synergism.Citation113 A synergism of such a double or triple combination in the prevention of androgen-independent pattern of growth of prostate cancer is also possible and merits investigation, as indicated by the former findings.Citation114
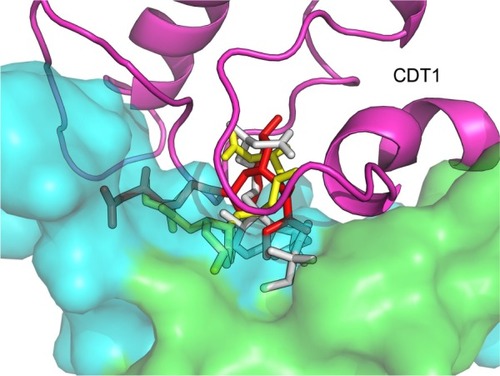
Finally, another future challenge is to identify new cancer-associated molecular targets of statins by exploiting the predictive power of bioinformatics tools in order to develop novel antitumor therapeutic schemes. Consistent with this, an in silico analysis indicates that simvastatin not only serves as a negative modulator of MCM7 expression as it was previously mentioned,Citation6,Citation42 but also may physically interact with another RLF, the human protein CDT1. Specifically, bioinformatics indicate that simvastatin possibly binds at one up to three different binding sites on human CDT1 and predict that this putative binding of the drug interferes with the intermolecular interaction between CDT1 and its physical inhibitor, geminin (). The importance of this prediction is substantiated by previous findings showing that silencing the expression of geminin is an emerging cancer cell-eliminating strategy irrespective of the p53 status which leaves normal or immortalized cells unaffected. This methodology is based on the geminin silencing-induced DNA re-replication and the consequent induction of genomic damage and programmed cell death. However, as it is reported, not all malignant human cells are equally affected by geminin silencing. In fact, HeLa and skin melanoma cells are not sensitive to geminin silencing.Citation203 Conceivably, an attractive goal is to carry out in vitro assays to address whether simvastatin and/or another statin physically interferes with the binding of geminin at CDT1 as in silico analysis predicts. If this assumption holds true, it would be very interesting to assess the physiologic relevance of this interference. In case simvastatin or another statin hinders the CDT1:geminin interaction, it is plausible that the inhibition of geminin activity and statin treatment could exhibit synergistic cancer cell-killing effects in cancer cells that are sensitive to geminin silencing. In addition, geminin silencing-resistant cells could be sensitized in the presence of simvastatin. This testable prediction merits experimental validation that could open new avenues in cancer therapeutics.
Figure 2 Docking of simvastatin on the human DNA replication licensing factor CDT1.
Supplementary material
Figure S1 Proteome profiling indicating alterations in the expression levels of human proteins evoked by the consumption of aspirin alone or combined with caffeine at clinically attainable doses.
Notes: Proteomic analysis in the serum of a 33-year-old healthy female individual 16 hours after the consumption of 500 mg of a marketed (ASPIRIN 500MG/TAB, Bayer Hellas A.B.E.E., Marousi 151 25, Attiki, Greece) tablet of aspirin (“A”) or a marketed (KALMALINE AC [500+30]MG/TAB, Coup A.B.E.E., Agias Varvaras 53, P.C. 17235 Athens, Greece) tablet containing 500 mg of aspirin and 30 mg of caffeine (“A+B”). None of these pharmacologic agents or even a placebo pill was consumed by the volunteer in the case of “control” serum analysis, while the individual voluntarily did not consume any food or beverage containing caffeine (chocolate, tea and so on) at least 24 hours before any of the three distinct blood sample collections, which took place throughout a 3-day period (ie, 1st, 2nd and 3rd days when control, sample A and sample A+B were collected). Approximately 8 mL of blood was collected in serum separating tubes (BD Vacutainer SST II 8.5 mL Advance Tubes-367958; Scientific Diagnostics Ltd, Attiki, Greece), and 2 mL of serum obtained after sample centrifugation for 15 min at 3,000 rpm was transferred into cryovials (Greiner Bio-One GmbH 5 mL U-Bottom cryovials CE-IVD; Bioline Scientific, Athens, Greece) and stored at −20°C, until their transfer in dry ice shortly after (1 day) the completion of the 3-day sample collecting period. All the experimental procedures associated with 2D-DIGE from protein extraction and sample labeling with CyDye DIGE fluors (blue, green and red for “Control”, “A” and “A+B” samples, respectively) to spot picking and protein identification by mass spectrometry were performed at Applied Biomics (Hayward, CA, USA) according to the established protocol as it has been described elsewhere.Citation1 Shown here (A) are 2D-DIGE gel images from overlay of each paired sample, A/control, A+B/control and A+B/A. The protein spots shown in yellow color means that protein expression is similar in the paired samples (two-color image overlay). White color (B) means that the protein expression is the same in all these samples (three-color image overlay). For the aforementioned three paired comparisons, a fold ≥1.3 was used as a cut-off value in the DeCyder analysis and most significant changed spots were selected. All the 39 spots were circled and numbered in the triple color overlay gel image with the estimated MW and isoelectric point (pI) grid (C). Each spot number was assigned (Assigned ID/protein ID) if the ratio of changes in at least one paired sample was ≥1.3 fold. Some of the statistically significant spot ratios corresponding to the expression of the 15 proteins which were mostly affected (either positively or negatively) in the A+B/A paired sample were chosen for identification by mass spectrometry. The rationale for focusing on this category of proteins is based on the fact that these proteins indicate the differential biologic outcome of aspirin when it is received in combination with caffeine (“A+B” or the numerator of the fraction) or not (“A” or the denominator of the fraction), respectively. Mass spectrometry revealed that the ID of four randomly chosen spots (C, shown as boxed values of the “A+B” paired sample) corresponding to the 15 most influenced proteins in the A+B/A paired sample is TTR (protein expression ratio 3, 5, spot number 20), RBP4 (protein expression ratio 1, 5, spot number 27), Hp (protein expression ratio 1, 71, spot number 30) and ApoA1 (protein expression ratio −2, 1, spot number 39). The role of these proteins in tumorigenesis and/or cancer diagnosis/prognosis is discussed in the text.
Abbreviations: 2D-DIGE, two-dimensional difference in gel electrophoresis; ApoA1, apolipoprotein A1; Hp, haptoglobin; MW, molecular weight; TTR, transthyretin; RBP4, retinol-binding protein 4.
![Figure S1 Proteome profiling indicating alterations in the expression levels of human proteins evoked by the consumption of aspirin alone or combined with caffeine at clinically attainable doses.Notes: Proteomic analysis in the serum of a 33-year-old healthy female individual 16 hours after the consumption of 500 mg of a marketed (ASPIRIN 500MG/TAB, Bayer Hellas A.B.E.E., Marousi 151 25, Attiki, Greece) tablet of aspirin (“A”) or a marketed (KALMALINE AC [500+30]MG/TAB, Coup A.B.E.E., Agias Varvaras 53, P.C. 17235 Athens, Greece) tablet containing 500 mg of aspirin and 30 mg of caffeine (“A+B”). None of these pharmacologic agents or even a placebo pill was consumed by the volunteer in the case of “control” serum analysis, while the individual voluntarily did not consume any food or beverage containing caffeine (chocolate, tea and so on) at least 24 hours before any of the three distinct blood sample collections, which took place throughout a 3-day period (ie, 1st, 2nd and 3rd days when control, sample A and sample A+B were collected). Approximately 8 mL of blood was collected in serum separating tubes (BD Vacutainer SST II 8.5 mL Advance Tubes-367958; Scientific Diagnostics Ltd, Attiki, Greece), and 2 mL of serum obtained after sample centrifugation for 15 min at 3,000 rpm was transferred into cryovials (Greiner Bio-One GmbH 5 mL U-Bottom cryovials CE-IVD; Bioline Scientific, Athens, Greece) and stored at −20°C, until their transfer in dry ice shortly after (1 day) the completion of the 3-day sample collecting period. All the experimental procedures associated with 2D-DIGE from protein extraction and sample labeling with CyDye DIGE fluors (blue, green and red for “Control”, “A” and “A+B” samples, respectively) to spot picking and protein identification by mass spectrometry were performed at Applied Biomics (Hayward, CA, USA) according to the established protocol as it has been described elsewhere.Citation1 Shown here (A) are 2D-DIGE gel images from overlay of each paired sample, A/control, A+B/control and A+B/A. The protein spots shown in yellow color means that protein expression is similar in the paired samples (two-color image overlay). White color (B) means that the protein expression is the same in all these samples (three-color image overlay). For the aforementioned three paired comparisons, a fold ≥1.3 was used as a cut-off value in the DeCyder analysis and most significant changed spots were selected. All the 39 spots were circled and numbered in the triple color overlay gel image with the estimated MW and isoelectric point (pI) grid (C). Each spot number was assigned (Assigned ID/protein ID) if the ratio of changes in at least one paired sample was ≥1.3 fold. Some of the statistically significant spot ratios corresponding to the expression of the 15 proteins which were mostly affected (either positively or negatively) in the A+B/A paired sample were chosen for identification by mass spectrometry. The rationale for focusing on this category of proteins is based on the fact that these proteins indicate the differential biologic outcome of aspirin when it is received in combination with caffeine (“A+B” or the numerator of the fraction) or not (“A” or the denominator of the fraction), respectively. Mass spectrometry revealed that the ID of four randomly chosen spots (C, shown as boxed values of the “A+B” paired sample) corresponding to the 15 most influenced proteins in the A+B/A paired sample is TTR (protein expression ratio 3, 5, spot number 20), RBP4 (protein expression ratio 1, 5, spot number 27), Hp (protein expression ratio 1, 71, spot number 30) and ApoA1 (protein expression ratio −2, 1, spot number 39). The role of these proteins in tumorigenesis and/or cancer diagnosis/prognosis is discussed in the text.Abbreviations: 2D-DIGE, two-dimensional difference in gel electrophoresis; ApoA1, apolipoprotein A1; Hp, haptoglobin; MW, molecular weight; TTR, transthyretin; RBP4, retinol-binding protein 4.](/cms/asset/6cf2fcc0-224a-43fc-968d-641ff348752d/dott_a_12195605_sf0001_c.jpg)
Reference
- ClementCCCannizzoESNastkeMDAn expanded self-antigen peptidome is carried by the human lymph as compared to the plasmaPLoS One201053e986320360855
Disclosure
The authors report no conflicts of interest in this work.
References
- SimpsonCBJamesJCThe efficacy of mitomycin-C in the treatment of laryngotracheal stenosisLaryngoscope2006116101923192517003706
- MearzaAAAslanidesIMUses and complications of mitomycin C in ophthalmologyExpert Opin Drug Saf200761273217181449
- LuYLiuXFLiuTRCelecoxib exerts antitumor effects in HL-60 acute leukemia cells and inhibits autophagy by affecting lysosome functionBiomed Pharmacother2016841551155727884749
- Vila-LeaheyAOldfordSAMarignaniPAWangJHaidlIDMarshallJSRanitidine modifies myeloid cell populations and inhibits breast tumor development and spread in miceOncoimmunology201657e115159127622015
- CharlesEHammadiMKischelPThe antidepressant fluoxetine induces necrosis by energy depletion and mitochondrial calcium overloadOncotarget2017823181319627911858
- LiJLiuJLiangZSimvastatin and Atorvastatin inhibit DNA replication licensing factor MCM7 and effectively suppress RB-deficient tumors growthCell Death Dis201783e2673
- PapanagnouPStivarouTTsironiMUnexploited antineoplastic effects of commercially available anti-diabetic drugsPharmaceuticals (Basel)201692
- EricesRCubillosSAravenaRDiabetic concentrations of metformin inhibit platelet-mediated ovarian cancer cell progressionOncotarget2017813208652088028209916
- PapanagnouPBaltopoulosPTsironiMMarketed nonsteroidal anti-inflammatory agents, antihypertensives, and human immunodeficiency virus protease inhibitors: as-yet-unused weapons of the oncologists’ arsenalTher Clin Risk Manag20151180781926056460
- JensenKBikasAPatelANelfinavir inhibits proliferation and induces DNA damage in thyroid cancer cellsEndocr Relat Cancer201724314715628137980
- RussoAPellosiDSPagliaraVBiotin-targeted Pluronic(®) P123/F127 mixed micelles delivering niclosamide: A repositioning strategy to treat drug-resistant lung cancer cellsInt J Pharm2016511112713927374195
- AhnSYYangJHKimNHAnti-helminthic niclosamide inhibits Ras-driven oncogenic transformation via activation of GSK-3Oncotarget2017819318563186328418865
- MeiZLiangMLiLZhangYWangQYangWEffects of statins on cancer mortality and progression: A systematic review and meta-analysis of 95 cohorts including 1,111,407 individualsInt J Cancer201714051068108127859151
- CaiXHuXTanXMetformin induced AMPK activation, G0/G1 Phase cell cycle arrest and the inhibition of growth of esophageal squamous cell carcinomas in vitro and in vivoPLoS One2015107e013334926196392
- WeberCNoelsHAtherosclerosis: current pathogenesis and therapeutic optionsNat Med201117111410142222064431
- PeltonKCoticchiaCMCuratoloASHypercholesterolemia induces angiogenesis and accelerates growth of breast tumors in vivoAm J Pathol201418472099211024952430
- FarnierMChenEJohnson-LevonasAOMcCrary SiskCMitchelYBEffects of extended-release niacin/laropiprant, simvastatin, and the combination on correlations between apolipoprotein B, LDL cholesterol, and non-HDL cholesterol in patients with dyslipidemiaVasc Health Risk Manag20141027929024855368
- ItzhakiOGreenbergEShalmonBNicotinamide inhibits vasculogenic mimicry, an alternative vascularization pathway observed in highly aggressive melanomaPLoS One201382e5716023451174
- ElangovanSPathaniaRRamachandranSThe niacin/butyrate receptor GPR109A suppresses mammary tumorigenesis by inhibiting cell survivalCancer Res20147441166117824371223
- GaoJLiaoJYangGYCAAX-box protein, prenylation process and carcinogenesisAm J Transl Res20091331232519956441
- SchoolingCMAu YeungSLFreemanGCowlingBJThe effect of statins on testosterone in men and women, a systematic review and meta-analysis of randomized controlled trialsBMC Med2013115723448151
- SolomonKRPeltonKBoucherKEzetimibe is an inhibitor of tumor angiogenesisAm J Pathol200917431017102619179610
- YuCCLaiWTShihKCEfficacy, safety and tolerability of ongoing statin plus ezetimibe versus doubling the ongoing statin dose in hypercholesterolemic Taiwanese patients: an open-label, randomized clinical trialBMC Res Notes2012525122621316
- ZhaoZCaoXPanYShaSZhaoTZhangTSimvastatin downregulates HER2 via upregulation of PEA3 to induce cell death in HER2-positive breast cancer cellsOncol Res2012205–618719523581225
- ShangLJiaSSJiangHMWangHXuWHLvCJSimvastatin downregulates expression of TGF-betaRII and inhibits proliferation of A549 cells via ERKTumour Biol20153664819482425631750
- WilliamsABLiLNguyenBBrownPLevisMSmallDFluvastatin inhibits FLT3 glycosylation in human and murine cells and prolongs survival of mice with FLT3/ITD leukemiaBlood2012120153069307922927251
- MiraECarmona-RodriguezLTardaguilaMA lovastatin-elicited genetic program inhibits M2 macrophage polarization and enhances T cell infiltration into spontaneous mouse mammary tumorsOncotarget20134122288230124317954
- LiaoJChungYTYangALAtorvastatin inhibits pancreatic carcinogenesis and increases survival in LSL-KrasG12D-LSL-Trp53R172H-Pdx1-Cre miceMol Carcinog201352973975022549877
- XuPYuHZhangZHydrogen-bonded and reduction-responsive micelles loading atorvastatin for therapy of breast cancer metastasisBiomaterials201435267574758724899527
- MatusewiczLMeissnerJToporkiewiczMSikorskiAFThe effect of statins on cancer cells–reviewTumour Biol20153674889490426002574
- LiuHWangZLiYLiWChenYSimvastatin prevents proliferation and bone metastases of lung adenocarcinoma in vitro and in vivoNeoplasma201360324024623373992
- WangLYangHLeiZRepression of TIF1gamma by SOX2 promotes TGF-beta-induced epithelial-mesenchymal transition in non-small-cell lung cancerOncogene201635786787725961934
- LiYFuJYuanXHuCSimvastatin inhibits the proliferation of A549 lung cancer cells through oxidative stress and up-regulation of SOD2Die Pharmazie201469861061425158572
- YuXPanYMaHLiWSimvastatin inhibits proliferation and induces apoptosis in human lung cancer cellsOncol Res201320835135723924855
- LiangYWChangCCHungCMChenTYHuangTYHsuYCPreclinical activity of simvastatin induces cell cycle arrest in G1 via blockade of cyclin D-Cdk4 expression in Non-Small Cell Lung Cancer (NSCLC)Int J Mol Sci20131435806581623481641
- SpampanatoCDe MariaSSarnataroMSimvastatin inhibits cancer cell growth by inducing apoptosis correlated to activation of Bax and down-regulation of BCL-2 gene expressionInt J Oncol201240493594122134829
- Ghosh-ChoudhuryNMandalCCGhosh-ChoudhuryNGhosh ChoudhuryGSimvastatin induces derepression of PTEN expression via NFkappaB to inhibit breast cancer cell growthCell Signal201022574975820060890
- MandalCCGhosh-ChoudhuryNYonedaTChoudhuryGGGhosh-ChoudhuryNSimvastatin prevents skeletal metastasis of breast cancer by an antagonistic interplay between p53 and CD44J Biol Chem201128613113141132721199873
- WangZWuYWangHInterplay of mevalonate and Hippo pathways regulates RHAMM transcription via YAP to modulate breast cancer cell motilityProc Natl Acad Sci U S A20141111E89E9824367099
- KotamrajuSWilliamsCLKalyanaramanBStatin-induced breast cancer cell death: role of inducible nitric oxide and arginase-dependent pathwaysCancer Res200767157386739417671209
- EinbondLSWuHASanduCDigitoxin enhances the growth inhibitory effects of thapsigargin and simvastatin on ER negative human breast cancer cellsFitoterapia201610914615426691294
- LiangZLiWLiuJSimvastatin suppresses the DNA replication licensing factor MCM7 and inhibits the growth of tamoxifen-resistant breast cancer cellsSci Rep201774177628150753
- LehnSFernoMJirstromKRydenLLandbergGA non-functional retinoblastoma tumor suppressor (RB) pathway in premenopausal breast cancer is associated with resistance to tamoxifenCell Cycle201110695696221358261
- RogersMKalraSMoukharskayaJSynergistic growth inhibition of PC3 prostate cancer cells with low-dose combinations of simvastatin and alendronateAnticancer Res20153541851185925862838
- NordstrandALundholmMLarssonALernerUHWidmarkAWikstromPInhibition of the insulin-like growth factor-1 receptor enhances effects of simvastatin on prostate cancer cells in co-culture with boneCancer Microenviron20136323124023335094
- Al-HuseinBGocASomanathPRSuppression of interactions between prostate tumor cell-surface integrin and endothelial ICAM-1 by simvastatin inhibits micrometastasisJ Cell Physiol2013228112139214823559257
- WangGCaoRWangYSimvastatin induces cell cycle arrest and inhibits proliferation of bladder cancer cells via PPARgamma signalling pathwaySci Rep201663578327779188
- BorahayMAKilicGSYallampalliCSimvastatin potently induces calcium-dependent apoptosis of human leiomyoma cellsJ Biol Chem201428951350753508625359773
- ChoSJKimJSKimJMLeeJYJungHCSongISSimvastatin induces apoptosis in human colon cancer cells and in tumor xenografts, and attenuates colitis-associated colon cancer in miceInt J Cancer2008123495195718521906
- JangHJHongEMParkSWStatin induces apoptosis of human colon cancer cells and downregulation of insulin-like growth factor 1 receptor via proapoptotic ERK activationOncol Lett201612125025627347133
- de LlobetLIBaroMMesiaRBalartJSimvastatin enhances the effects of radiotherapy and cetuximab on a cell line (FaDu) derived from a squamous cell carcinoma of head and neckTransl Oncol20147451352225171892
- FuchsDBergesCOpelzGDanielVNaujokatCHMG-CoA reductase inhibitor simvastatin overcomes bortezomib-induced apoptosis resistance by disrupting a geranylgeranyl pyrophosphate-dependent survival pathwayBiochem Biophys Res Commun2008374230931418625202
- SaitoASaitoNMolWSimvastatin inhibits growth via apoptosis and the induction of cell cycle arrest in human melanoma cellsMelanoma Res2008182859418337644
- GlynnSAO’SullivanDEustaceAJClynesMO’DonovanNThe 3-hydroxy-3-methylglutaryl-coenzyme A reductase inhibitors, simvastatin, lovastatin and mevastatin inhibit proliferation and invasion of melanoma cellsBMC Cancer20088918199328
- GronichNDruckerLShapiroHRadnayJYarkoniSLishnerMSimvastatin induces death of multiple myeloma cell linesJ Investig Med2004525335344
- SchmidmaierRBaumannPSimsekMDayyaniFEmmerichBMeinhardtGThe HMG-CoA reductase inhibitor simvastatin overcomes cell adhesion-mediated drug resistance in multiple myeloma by geranylgeranylation of Rho protein and activation of Rho kinaseBlood200410461825183215161667
- AtilBBerger-SieczkowskiEBardyJWernerMHoheneggerMIn vitro and in vivo downregulation of the ATP binding cassette transporter B1 by the HMG-CoA reductase inhibitor simvastatinNaunyn Schmiedeberg’s Arch Pharmacol20163891173226319048
- WolfeARDebebBGLacerdaLSimvastatin prevents triple-negative breast cancer metastasis in pre-clinical models through regulation of FOXO3aBreast Cancer Res Treat2015154349550826590814
- FangZTangYFangJSimvastatin inhibits renal cancer cell growth and metastasis via AKT/mTOR, ERK and JAK2/STAT3 pathwayPLoS One201385e6282323690956
- AlupeiMCLicareteEPatrasLBanciuMLiposomal simvastatin inhibits tumor growth via targeting tumor-associated macrophages-mediated oxidative stressCancer Lett20153562 Pt B94695225444912
- CoimbraMBanciuMFensMHLiposomal pravastatin inhibits tumor growth by targeting cancer-related inflammationJ Control Release2010148330331020869410
- LeeSJLeeILeeJParkCKangWKStatins, 3-hydroxy-3-methylglutaryl coenzyme A reductase inhibitors, potentiate the anti-angiogenic effects of bevacizumab by suppressing angiopoietin2, BiP, and Hsp90alpha in human colorectal cancerBr J Cancer2014111349750524945998
- LiGZhengJXuBLingJQiuWWangYSimvastatin inhibits tumor angiogenesis in HER2-overexpressing human colorectal cancerBiomed Pharmacother20178541842427887848
- CohenJIHMG CoA reductase inhibitors (statins) to treat Epstein-Barr virus-driven lymphomaBr J Cancer20059291593159815856040
- InanoHSuzukiKOnodaMWakabayashiKAnti-carcinogenic activity of simvastatin during the promotion phase of radiation-induced mammary tumorigenesis of ratsCarcinogenesis1997189172317279328167
- ClutterbuckRDMillarBCPowlesRLInhibitory effect of simvastatin on the proliferation of human myeloid leukaemia cells in severe combined immunodeficient (SCID) miceBr J Haematol199810225225279695968
- ErbasHBalOCakirEEffect of rosuvastatin on arginase enzyme activity and polyamine production in experimental breast cancerBalkan Med J2015321899525759778
- PolatMFTaysiSPolatSBoyukABakanEElevated serum arginase activity levels in patients with breast cancerSurg Today200333965566112928840
- MorrisSMJrArginine metabolism: boundaries of our knowledgeJ Nutr20071376 Suppl 21602s1609s17513435
- NowotarskiSLWosterPMCaseroRAJrPolyamines and cancer: implications for chemotherapy and chemopreventionExpert Rev Mol Med201315e323432971
- PlunkettWArginine addiction in AMLBlood2015125263971397226113532
- NarisawaTFukauraYTanidaNHasebeMItoMAizawaRChemopreventive efficacy of low dose of pravastatin, an HMG-CoA reductase inhibitor, on 1,2-dimethylhydrazine-induced colon carcinogenesis in ICR miceTohoku J Exp Med199618021311389111762
- NarisawaTMorotomiMFukauraYHasebeMItoMAizawaRChemoprevention by pravastatin, a 3-hydroxy-3-methylglutaryl-coenzyme A reductase inhibitor, of N-methyl-N-nitrosourea-induced colon carcinogenesis in F344 ratsJpn J Cancer Res19968787988048797885
- ShellmanYGRibbleDMillerLLovastatin-induced apoptosis in human melanoma cell linesMelanoma Res2005152838915846140
- DimitroulakosJYegerHHMG-CoA reductase mediates the biological effects of retinoic acid on human neuroblastoma cells: lovastatin specifically targets P-glycoprotein-expressing cellsNat Med1996233263338612233
- RubinsJBGreatensTKratzkeRATanATPolunovskyVABittermanPLovastatin induces apoptosis in malignant mesothelioma cellsAm J Respir Crit Care Med19981575 Pt 1161616229603146
- DimitroulakosJNohynekDBackwayKLIncreased sensitivity of acute myeloid leukemias to lovastatin-induced apoptosis: A potential therapeutic approachBlood1999934130813189949174
- MacaulayRJWangWDimitroulakosJBeckerLEYegerHLovastatin-induced apoptosis of human medulloblastoma cell lines in vitroJ Neurooncol199942111110360474
- DimitroulakosJYeLYBenzaquenMDifferential sensitivity of various pediatric cancers and squamous cell carcinomas to lovastatin-induced apoptosis: therapeutic implicationsClin Cancer Res20017115816711205904
- OgunwobiOOBealesILStatins inhibit proliferation and induce apoptosis in Barrett’s esophageal adenocarcinoma cellsAm J Gastroenterol2008103482583718371146
- MaksimovaEYieTARomWNIn vitro mechanisms of lovastatin on lung cancer cell lines as a potential chemopreventive agentLung20081861455418034278
- ParkIHKimJYJungJIHanJYLovastatin overcomes gefitinib resistance in human non-small cell lung cancer cells with K-Ras mutationsInvest New Drugs201028679179919760159
- JeonJHKimSKKimHJChangJAhnCMChangYSLipid raft modulation inhibits NSCLC cell migration through delocalization of the focal adhesion complexLung Cancer201069216517119945766
- ZhangXTengYYangFMCM2 is a therapeutic target of lovastatin in human non-small cell lung carcinomasOncol Rep20153352599260525738322
- AgarwalBBhendwalSHalmosBMossSFRameyWGHoltPRLovastatin augments apoptosis induced by chemotherapeutic agents in colon cancer cellsClin Cancer Res1999582223222910473109
- SwamyMVCoomaIReddyBSRaoCVLamin B, caspase-3 activity, and apoptosis induction by a combination of HMG-CoA reductase inhibitor and COX-2 inhibitors: a novel approach in developing effective chemopreventive regimensInt J Oncol200220475375911894121
- ManthaAJHansonJEGossGLagardeAELorimerIADimitroulakosJTargeting the mevalonate pathway inhibits the function of the epidermal growth factor receptorClin Cancer Res20051162398240715788691
- HoqueAChenHXuXCStatin induces apoptosis and cell growth arrest in prostate cancer cellsCancer Epidemiol Biomarkers Prev2008171889418199714
- ParkCLeeIKangWKLovastatin-induced E2F-1 modulation and its effect on prostate cancer cell deathCarcinogenesis200122101727173111577016
- AfshordelSKernBClasohmJLovastatin and perillyl alcohol inhibit glioma cell invasion, migration, and proliferation – impact of Ras-/Rho-prenylationPharmacol Res201591697725497898
- Glodkowska-MrowkaEMrowkaPBasakGWStatins inhibit ABCB1 and ABCG2 drug transporter activity in chronic myeloid leukemia cells and potentiate antileukemic effects of imatinibExp Hematol201442643944724667683
- SongXLiuBCLuXYLovastatin inhibits human B lymphoma cell proliferation by reducing intracellular ROS and TRPC6 expressionBiochim Biophys Acta20141843589490124518247
- YangTYaoHHeGEffects of lovastatin on MDA-MB-231 breast cancer cells: an antibody microarray analysisJ Cancer20167219219926819643
- HolsteinSAHohlRJInteraction of cytosine arabinoside and lovastatin in human leukemia cellsLeuk Res200125865166011397469
- ZhaoTTLe FrancoisBGGossGDingKBradburyPADimitroulakosJLovastatin inhibits EGFR dimerization and AKT activation in squamous cell carcinoma cells: potential regulation by targeting rho proteinsOncogene201029334682469220562912
- WeitbergABThe antioxidant effect of lovastatin on phagocyte-induced DNA damage: implications for cancer preventionJ Exp Clin Cancer Res200726458358618365556
- AjithTASojaMA comparative study on the antimutagenicity of atorvastatin and lovastatin against directly acting mutagensCell Biol Toxicol200622426927416642263
- FeleszkoWZagozdzonRGolabJJakobisiakMPotentiated antitumour effects of cisplatin and lovastatin against MmB16 melanoma in miceEur J Cancer19983434064119640231
- FeleszkoWBalkowiecEZSieberthELovastatin and tumor necrosis factor-alpha exhibit potentiated antitumor effects against Ha-ras-transformed murine tumor via inhibition of tumor-induced angiogenesisInt J Cancer199981456056710225445
- AjithTAHarikumarKBThasnaHSabuMCBabithaNVProapoptotic and antitumor activities of the HMG-CoA reductase inhibitor, lovastatin, against Dalton’s lymphoma ascites tumor in miceClin Chim Acta20063661–232232816380106
- MatarPRozadosVRRoggeroEAScharovskyOGLovastatin inhibits tumor growth and metastasis development of a rat fibrosarcomaCancer Biother Radiopharm199813538739310851430
- MatarPRozadosVRBindaMMRoggeroEABonfilRDScharovskyOGInhibitory effect of Lovastatin on spontaneous metastases derived from a rat lymphomaClin Exp Metastasis1999171192510390143
- JaniJPSpechtSStemmlerNMetastasis of B16F10 mouse melanoma inhibited by lovastatin, an inhibitor of cholesterol biosynthesisInvasion Metastasis19931363143247860224
- AloPLViscaPMarciAMangoniABottiCDi TondoUExpression of fatty acid synthase (FAS) as a predictor of recurrence in stage I breast carcinoma patientsCancer19967734744828630954
- HawkMACesenKTSiglinJCStonerGDRuchRJInhibition of lung tumor cell growth in vitro and mouse lung tumor formation by lovastatinCancer Lett19961091–22172229020924
- DingNCuiXXGaoZA triple combination of atorvastatin, celecoxib and tipifarnib strongly inhibits pancreatic cancer cells and xenograft pancreatic tumorsInt J Oncol20144462139214524647860
- IslamMSharmaSKumarBTeknosTNAtorvastatin inhibits RhoC function and limits head and neck cancer metastasisOral Oncol201349877878623648017
- FanZJiangHWangZQuJAtorvastatin partially inhibits the epithelial-mesenchymal transition in A549 cells induced by TGF-beta1 by attenuating the upregulation of SphK1Oncol Rep20163621016102227349500
- GhalaliAWiklundFZhengHSteniusUHogbergJAtorvastatin prevents ATP-driven invasiveness via P2X7 and EHBP1 signaling in PTEN-expressing prostate cancer cellsCarcinogenesis20143571547155524451147
- AldertonGKTumour immunology: suppressing tumorigenic inflammationNat Rev Cancer2012124228
- BrisoEMGuinea-ViniegraJBakiriLInflammation-mediated skin tumorigenesis induced by epidermal c-FosGenes Dev201327181959197324029918
- SuhNReddyBSDeCastroACombination of atorvastatin with sulindac or naproxen profoundly inhibits colonic adenocarcinomas by suppressing the p65/beta-catenin/cyclin D1 signaling pathway in ratsCancer Prev Res (Phila)20114111895190221764859
- ZhengXCuiXXAvilaGEAtorvastatin and celecoxib inhibit prostate PC-3 tumors in immunodeficient miceClin Cancer Res20071318 Pt 15480548717875778
- ZhengXCuiXXGaoZAtorvastatin and celecoxib in combination inhibits the progression of androgen-dependent LNCaP xenograft prostate tumors to androgen independenceCancer Prev Res (Phila)20103111412420051379
- SwamyMVPatlollaJMSteeleVEKopelovichLReddyBSRaoCVChemoprevention of familial adenomatous polyposis by low doses of atorvastatin and celecoxib given individually and in combination to APCMin miceCancer Res200666147370737716849589
- ShachafCMPerezODYoussefSInhibition of HMGcoA reductase by atorvastatin prevents and reverses MYC-induced lymphomagenesisBlood200711072674268417622571
- Slawinska-BrychAZdzisinskaBKandefer-SzerszenMFluvastatin inhibits growth and alters the malignant phenotype of the C6 glioma cell linePharmacol Rep201466112112924905317
- SassanoAKatsoulidisEAnticoGSuppressive effects of statins on acute promyelocytic leukemia cellsCancer Res20076794524453217483369
- WoodardJSassanoAHayNPlataniasLCStatin-dependent suppression of the Akt/mammalian target of rapamycin signaling cascade and programmed cell death 4 up-regulation in renal cell carcinomaClin Cancer Res200814144640464918628479
- KusamaTMukaiMIwasakiTInhibition of epidermal growth factor-induced RhoA translocation and invasion of human pancreatic cancer cells by 3-hydroxy-3-methylglutaryl-coenzyme a reductase inhibitorsCancer Res200161124885489111406567
- TakahashiHKWeitz-SchmidtGIwagakiHYoshinoTTanakaNNishiboriMHypothesis: the antitumor activities of statins may be mediated by IL-18J Leukocyte Biol200680221521616735695
- HoriguchiASumitomoMAsakumaJAsanoTAsanoTHayakawaM3-hydroxy-3-methylglutaryl-coenzyme a reductase inhibitor, fluvastatin, as a novel agent for prophylaxis of renal cancer metastasisClin Cancer Res200410248648865515623649
- ThibaultASamidDTompkinsACPhase I study of lovastatin, an inhibitor of the mevalonate pathway, in patients with cancerClin Cancer Res1996234834919816194
- LarnerJJaneJLawsEPackerRMyersCShaffreyMA phase I–II trial of lovastatin for anaplastic astrocytoma and glioblastoma multiformeAm J Clin Oncol19982165795839856659
- KimWSKimMMChoiHJPhase II study of high-dose lovastatin in patients with advanced gastric adenocarcinomaInvest New Drugs2001191818311291836
- KawataSYamasakiENagaseTEffect of pravastatin on survival in patients with advanced hepatocellular carcinoma. A randomized controlled trialBr J Cancer200184788689111286466
- GrafHJungstCStraubGChemoembolization combined with pravastatin improves survival in patients with hepatocellular carcinomaDigestion2008781343818797167
- LeeJJungKHParkYSSimvastatin plus irinotecan, 5-fluorouracil, and leucovorin (FOLFIRI) as first-line chemotherapy in metastatic colorectal patients: a multicenter phase II studyCancer Chemother Pharmacol200964465766319169686
- BaasJMKrensLLten TijeAJSafety and efficacy of the addition of simvastatin to cetuximab in previously treated KRAS mutant metastatic colorectal cancer patientsInvest New Drugs20153361242124726386973
- BaasJMKrensLLBosMMSafety and efficacy of the addition of simvastatin to panitumumab in previously treated KRAS mutant metastatic colorectal cancer patientsAnticancer Drugs201526887287726053280
- KrensLLSimkensLHBaasJMStatin use is not associated with improved progression free survival in cetuximab treated KRAS mutant metastatic colorectal cancer patients: results from the CAIRO2 studyPLoS One2014911e11220125375154
- NielsenSFNordestgaardBGBojesenSEStatin use and reduced cancer-related mortalityN Engl J Med20133686576577
- KlopCDriessenJHde VriesFStatin use and reduced cancer-related mortalityN Engl J Med20133686574
- KopelEMaorEGoldenbergIStatin use and reduced cancer-related mortalityN Engl J Med2013368657557623388015
- BrewerTMMasudaHLiuDDStatin use in primary inflammatory breast cancer: a cohort studyBr J Cancer2013109231832423820253
- BjarnadottirORomeroQBendahlPOTargeting HMG-CoA reductase with statins in a window-of-opportunity breast cancer trialBreast Cancer Res Treat2013138249950823471651
- BjarnadottirOKimbungSJohanssonIGlobal transcriptional changes following statin treatment in breast cancerClin Cancer Res201521153402341125840970
- FeldtMBjarnadottirOKimbungSStatin-induced antiproliferative effects via cyclin D1 and p27 in a window-of-opportunity breast cancer trialJ Transl Med20151313325925673
- GarwoodERKumarASBaehnerFLFluvastatin reduces proliferation and increases apoptosis in women with high grade breast cancerBreast Cancer Res Treat2010119113714419728082
- WangAStefanickMLKapphahnKRelation of statin use with non-melanoma skin cancer: prospective results from the Women’s Health InitiativeBr J Cancer2016114331432026742009
- Randomised trial of cholesterol lowering in 4444 patients with coronary heart disease: the Scandinavian Simvastatin Survival Study (4S)Lancet19943448934138313897968073
- MRC/BHF Heart Protection Study of cholesterol lowering with simvastatin in 20,536 high-risk individuals: a randomised placebo-controlled trialLancet2002360932672212114036
- KuoppalaJLamminpaaAPukkalaEStatins and cancer: a systematic review and meta-analysisEur J Cancer200844152122213218707867
- ArnspangSPottegardAFriisSStatin use and risk of non-melanoma skin cancer: a nationwide study in DenmarkBr J Cancer2015112115315625290087
- BlaisLDesgagneALeLorierJ3-Hydroxy-3-methylglutaryl coenzyme A reductase inhibitors and the risk of cancer: a nested case-control studyArch Intern Med2000160152363236810927735
- GraafMRBeiderbeckABEgbertsACRichelDJGuchelaarHJThe risk of cancer in users of statinsJ Clin Oncol200422122388239415197200
- PetoREmbersonJLandrayMAnalyses of cancer data from three ezetimibe trialsN Engl J Med2008359131357136618765432
- BjerreLMLeLorierJDo statins cause cancer? A meta-analysis of large randomized clinical trialsAm J Med2001110971672311403756
- AsgariMMTangJEpsteinEHJrStatin use and risk of basal cell carcinomaJ Am Acad Dermatol2009611667219464071
- HaukkaJSankilaRKlaukkaTIncidence of cancer and statin usage–record linkage studyInt J Cancer2010126127928419739258
- LiXWuXBChenQStatin use is not associated with reduced risk of skin cancer: a meta-analysisBr J Cancer2014110380280724366301
- LimSHKimTWHongYSA randomised, double-blind, placebo-controlled multi-centre phase III trial of XELIRI/FOLFIRI plus simvastatin for patients with metastatic colorectal cancerBr J Cancer2015113101421142626505681
- WallaceAChinnDRubinGTaking simvastatin in the morning compared with in the evening: randomised controlled trialBMJ2003327741878814525878
- DrazenJMD’AgostinoRBWareJHMorrisseySCurfmanGDEzetimibe and cancer–an uncertain associationN Engl J Med2008359131398139918765434
- RosseboABPedersenTRBomanKIntensive lipid lowering with simvastatin and ezetimibe in aortic stenosisN Engl J Med2008359131343135618765433
- GreenARameyDREmneusMIncidence of cancer and mortality in patients from the Simvastatin and Ezetimibe in Aortic Stenosis (SEAS) trialAm J Cardiol2014114101518152225267716
- MarcelliMCunninghamGRHaidacherSJCaspase-7 is activated during lovastatin-induced apoptosis of the prostate cancer cell line LNCaPCancer Res199858176839426061
- GbelcovaHLenicekMZelenkaJDifferences in antitumor effects of various statins on human pancreatic cancerInt J Cancer200812261214122118027870
- BurkeLPKukolyCAStatins induce lethal effects in acute myeloblastic leukemia [corrected] cells within 72 hoursLeuk Lymphoma200849232233018231920
- PrasannaPThibaultALiuLSamidDLipid metabolism as a target for brain cancer therapy: synergistic activity of lovastatin and sodium phenylacetate against human glioma cellsJ Neurochem19966627107168592143
- KornbrustDJMacDonaldJSPeterCPToxicity of the HMG-coenzyme A reductase inhibitor, lovastatin, to rabbitsJ Pharmacol Exp Ther198924824985052918466
- FeleszkoWMlynarczukIBalkowiec-IskraEZLovastatin potentiates antitumor activity and attenuates cardiotoxicity of doxorubicin in three tumor models in miceClin Cancer Res2000652044205210815931
- LishnerMBar-SefAElisAFabianIEffect of simvastatin alone and in combination with cytosine arabinoside on the proliferation of myeloid leukemia cell linesJ Investig Med2001494319324
- LeeJLeeIHanBEffect of simvastatin on cetuximab resistance in human colorectal cancer with KRAS mutationsJ Natl Cancer Inst2011103867468821398618
- XiaoHYangCSCombination regimen with statins and NSAIDs: a promising strategy for cancer chemopreventionInt J Cancer2008123598399018548583
- MaronDJFazioSLintonMFCurrent perspectives on statinsCirculation2000101220721310637210
- FarmerJATorre-AmioneGComparative tolerability of the HMG-CoA reductase inhibitorsDrug Safety200023319721311005703
- NawarskasJJHMG-CoA reductase inhibitors and coenzyme Q10Cardiology Rev20051327679
- WilliamsDFeelyJPharmacokinetic-pharmacodynamic drug interactions with HMG-CoA reductase inhibitorsClin Pharmacokinet200241534337012036392
- MoldenEAnderssonKSSimvastatin-associated rhabdomyolysis after coadministration of macrolide antibiotics in two patientsPharmacotherapy200727460360717381388
- BatchelorTTTaylorLPThalerHTPosnerJBDeAngelisLMSteroid myopathy in cancer patientsNeurology1997485123412389153449
- GopaluniSThomasLMShahCWhen myopathy strikes in a cancer patientGastrointest Cancer Res: GCR20093521221420084165
- TroyanovYLandon-CardinalOFritzlerMJAtorvastatin-induced necrotizing autoimmune myositis: An emerging dominant entity in patients with autoimmune myositis presenting with a pure polymyositis phenotypeMedicine (Baltimore)2017963e569428099331
- TellierPL’ezetimibe : de la pharmacologie aux essais thérapeutiques [Ezetimibe: from pharmacology to clinical trials]Ann Endocrinol (Paris)2003646442447 French15067249
- JeuLChengJWPharmacology and therapeutics of ezetimibe (SCH 58,235), a cholesterol-absorption inhibitorClin Ther20032592352238714604738
- YatskarLFisherEASchwartzbardAEzetimibe: rationale and role in the management of hypercholesterolemiaClin Cardiol2006292525516506638
- FraunbergerPGroneEGroneHJDrexelHWalliAKEzetimibe reduces cholesterol content and NF-kappaB activation in liver but not in intestinal tissue in guinea pigsJ Inflamm (London)201714328167864
- SakuraiTHigashitsujiHKashidaHThe oncoprotein gankyrin promotes the development of colitis-associated cancer through activation of STAT3Oncotarget2017815247622477628160571
- AssarafYGMolecular basis of antifolate resistanceCancer Metastasis Rev200726115318117333344
- OronskyBTReidTKnoxSJScicinskiJJThe scarlet letter of alkylation: a mini review of selective alkylating agentsTransl Oncol20125422622922937173
- YaredJATkaczukKHUpdate on taxane development: new analogs and new formulationsDrug Des Devel Ther20126371384
- BrownCTargeted therapy: an elusive cancer targetNature20165377620S106S10827626779
- WarburgOOn the origin of cancer cellsScience (New York, NY)19561233191309314
- BuchwaldHCholesterol inhibition, cancer, and chemotherapyLancet (London, England)1992339880211541156
- CruzPMMoHMcConathyWJSabnisNLackoAGThe role of cholesterol metabolism and cholesterol transport in carcinogenesis: a review of scientific findings, relevant to future cancer therapeuticsFront Pharmacol2013411924093019
- AuneDKeumNGiovannucciEWhole grain consumption and risk of cardiovascular disease, cancer, and all cause and cause specific mortality: systematic review and dose-response meta-analysis of prospective studiesBMJ2016353i271627301975
- ShiCZhangNFengYCaoJChenXLiuBAspirin inhibits IKK-beta-mediated prostate cancer cell invasion by targeting matrix metalloproteinase-9 and Urokinase-type plasminogen activatorCell Physiol Biochem20174141313132428278500
- JiangJLanYQZhangTThe in vitro effects of caffeine on viability, cycle cycle profiles, proliferation, and apoptosis of glioblastomasEur Rev Med Pharmacol Sci201519173201320726400523
- DongSKongJKongJLow concentration of caffeine inhibits the progression of the hepatocellular carcinoma via Akt signaling pathwayAnticancer Agents Med Chem201515448449225666502
- ChenYChouWCDingYMWuYCCaffeine inhibits migration in glioma cells through the ROCK-FAK pathwayCell Physiol Biochem20143361888189825011614
- LiuLLiuJDaiSReduced transthyretin expression in sera of lung cancerCancer Sci200798101617162417683510
- NoyNLiLAbolaMVBergerNAIs retinol binding protein 4 a link between adiposity and cancer?Horm Mol Biol Clin Investig20152323946
- ZhaoCAnnamalaiLGuoCCirculating haptoglobin is an independent prognostic factor in the sera of patients with epithelial ovarian cancerNeoplasia2007911717325738
- Zamanian-DaryoushMDiDonatoJAApolipoprotein A-I and cancerFront Pharmacol2015626526617517
- MareeAOFitzgeraldDJAspirin and coronary artery diseaseThromb Haemost20049261175118115583721
- BarrettPJCaseyEKSisungCGaebler-SpiraDCaffeine as a neurostimulant: Two pediatric acquired brain injury casesJ Pediatr Rehabil Med20103322923221791853
- TazzeoTBatesGRomanHNCaffeine relaxes smooth muscle through actin depolymerizationAm J Physiol Lung Cell Mol Physiol20123034L334L34222683573
- KolahdouzanMHamadehMJThe neuroprotective effects of caffeine in neurodegenerative diseasesCNS Neurosci Ther201723427229028317317
- EngelhardtGMauzABPairetMRole of caffeine in combined analgesic drugs from the point of view of experimental pharmacologyArzneimittelforschung19974789179279296278
- PiniLAGuerzoniSCainazzoMCiccareseMPrudenzanoMPLivreaPComparison of tolerability and efficacy of a combination of paracetamol + caffeine and sumatriptan in the treatment of migraine attack: a randomized, double-blind, double-dummy, cross-over studyJ Headache Pain201213866967523054063
- CortezDCaffeine inhibits checkpoint responses without inhibiting the ataxia-telangiectasia-mutated (ATM) and ATM- and Rad3-related (ATR) protein kinasesJ Biol Chem200327839371393714512847089
- KimYMLeeEJParkSYChoKHKimJYPyoHCyclooxygenase-2 up-regulates ataxia telangiectasia and Rad3 related through extracellular signal-regulated kinase activationMol Cancer Res2009771158116819584262
- ZhuWDepamphilisMLSelective killing of cancer cells by suppression of geminin activityCancer Res200969114870487719487297