
Abstract
Background
Nuclear enriched abundant transcript 1 (NEAT1) has been demonstrated to act as a tumor inhibitor in many cancers. However, the role of NEAT1 in the development of ovarian cancer (OC) remains far from being elaborated. Hence, the aim of this study is to investigate the expression and function of NEAT1 in OC.
Materials and methods
The expression level of NEAT1 was determined by quantitative real-time polymerase chain reaction in OC cell lines. MTT assay, caspase-3 activity assay, and flow cytometry analysis were conducted to investigate the effects of NEAT1, miR-34a-5p, or B-cell lymphoma-2 (BCL2) on OC cell proliferation and apoptosis. Luciferase reporter assay was used to confirm the interaction of NEAT1, BCL2, and miR-34a-5p in OC cells.
Results
NEAT1 was significantly upregulated in OC cell lines. NEAT1 overexpression promoted proliferation by increasing the proportion of cells in S phase and suppressed apoptosis of OC cells, while knockdown of NEAT1 had the opposite effect. In addition, NEAT1 was demonstrated to directly interact with miR-34a-5p and exert its oncogenic role in OC by negatively regulating miR-34a-5p. Moreover, miR-34a-5p could directly target BCL2 and suppressed its expression. miR-34a-5p overexpression suppressed OC cell proliferation and triggered apoptosis by targeting BCL2. Furthermore, NEAT1 knockdown suppressed BCL2 expression, while anti-miR-34a-5p dramatically abated the inhibitory effect of si-NEAT1 on BCL2 expression.
Conclusion
NEAT1 regulated proliferation and apoptosis of OC cells by miR-34a-5p/BCL2, providing a potential therapeutic approach for the treatment of OC patients.
Introduction
Ovarian cancer (OC) is one of the deadliest gynecologic malignancies and a leading cause of cancer-related death in the female population worldwide, with >230,000 new cases diagnosed and 140,000 deaths of women occurring every year.Citation1,Citation2 Although considerable research efforts have been made toward the diagnosis and treatment of OC in the last two decades, the clinical outcomes are still unsatisfactory in advanced OC patients.Citation3 Since no obvious symptoms are presented in patients with early OC, the majority (up to 70%) of OC patients are estimated to be diagnosed at an advanced stage (stages III and IV), with an overall 5-year survival rate of ~30%.Citation4 Therefore, there is an urgent need to identify potential indicators and elucidate the molecular mechanism involved in OC in order to develop more effective therapies for human OC.
Human genome sequence data indicates that the majority of transcripts are referred to as ncRNAs, which can be characterized into two distinct groups: miRNAs and lncRNAs.Citation5 miRNAs have been extensively elucidated to exert significant biologic functions in numerous cancers.Citation6,Citation7 It has also been proposed that lncRNAs are able to participate in various physiologic and pathologic processes through regulation of coding genes at the transcriptional, posttranscriptional, or epigenetic levels in diverse malignant tumors.Citation8 Increasing evidence has suggested that lncRNAs serve as oncogenes or tumor-suppressive genes in different type of tumors depending on the circumstance.Citation9 Nuclear enriched abundant transcript 1 (NEAT1), a ~3.2 kb novel lncRNA localized to the nucleus, serves as a core structural component of paraspeckle suborganelles.Citation10,Citation11 NEAT1 has been reported to be highly expressed and to function as an oncogene in several human malignancies including non-small cell lung cancer (NSCLC), glioma, and breast cancer.Citation12–Citation14 More importantly, a previous study demonstrated that NEAT1 was upregulated in OC patients and cell lines and promoted OC cell proliferation and invasion.Citation15
By far, the detailed molecular mechanism by which NEAT1 exerts its oncogenic activity in OC remains to be clarified. Recently, a new regulatory mechanism about ceRNAs hypothesis has been proposed.Citation16 In this case, lncRNAs may function as molecular sponges or decoys for miRNAs to negatively regulate the expression and activities of miRNAs, thereby resulting in the derepression of miRNA targets at the level of posttranscriptional regulation.Citation17,Citation18 Any perturbation of ceRNA–miRNA–mRNA regulatory system may lead to pathologic processes such as malignant tumors.Citation19 According to a previous study, miR-34a was downregulated in OC tissues and functioned as a tumor suppressor in OC.Citation20 However, whether NEAT1 could directly interact with miR-34a-5p to regulate OC development remains largely unknown.
In this study, the expression of NEAT1 in OC cell lines and the biologic role of NEAT1 in OC cell proliferation and apoptosis were analyzed. Moreover, mechanistic analysis demonstrated that NEAT1 may function as a ceRNA to upregulate B-cell lymphoma-2 (BCL2) by sponging miR-34a-5p, thus promoting cell proliferation and inhibiting apoptosis in OC. Here, these findings provided a potential application of NEAT1-miR-34a-5p-BCL2 regulatory system in OC, shedding a new light on the treatment of OC.
Materials and methods
Cell lines and culture
Human OC cell lines (OVCAR3, SKOV3, HO8910, and OV90) and normal ovarian epithelial cell line HOSEpiC were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). All cells were maintained in RPMI 1640 medium (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (Thermo Fisher Scientific), 100 U/mL penicillin, and 100 μg/mL streptomycin (Thermo Fisher Scientific) in humidified incubator containing 5% CO2 at 37°C.
Cell transfection
To overexpress NEAT1, NEAT1 genomic fragment was cloned by polymerase chain reaction (PCR) and inserted into the pCMV6 empty vector (Vector; OriGene, Rockville, MD, USA), named as pCMV6-NEAT1 (NEAT1). pcDNA-BCL2 plasmid was established by cloning BCL2 sequence into the empty pcDNA Vector (Thermo Fisher Scientific). siRNA against NEAT1 (si-NEAT1), scrambled siRNA negative control (si-NC), miR-34a-5p mimic (miR-34a-5p), scrambled miRNA negative control (miR-NC), miR-34a-5p inhibitor (anti-miR-34a-5p), and anti-miRNA negative control (anti-miR-NC) were obtained from Genepharma (Shanghai, China). SKOV3 and OVCAR3 cells were transfected with oligonucleotide or plasmids using Lipofectamine 2000 (Thermo Fisher Scientific).
Quantitative real-time PCR (qRT-PCR)
Total RNA was extracted from cells using TRIzol reagent (Thermo Fisher Scientific). After RNase-free DNAse I (Takara, Tokyo, Japan) treatment, DNA-free RNA was reverse-transcribed into cDNA using a High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific). The expression level of NEAT1 was determined using the SYBR Green PCR kit (Thermo Fisher Scientific) and GAPDH served as an endogenous control. miR-34a-5p expression was examined using TaqMan miRNA assay (Thermo Fisher Scientific) and U6 snRNA was used as the internal control. All PCR steps were performed on the ABI 7300 system (Thermo Fisher Scientific) and the relative gene expression was calculated using the 2−∆∆CT method. The primers were as follows: NEAT1, 5′-CTTCCTCCCTTTA ACTTATCCATTCAC-3′ (forward) and 5′-CTCTTCCTC CACCATTACCAACAATAC-3′ (reverse); miR-34a-5p, 5′-TGGCAGTGTCTTAGCTGGTTGT-3′ (forward) and 5′-GCGAGCACAGAATTAATACGAC-3′ (reverse); GAPDH, 5′-ACATCAAGAAGGTGGTGAAGCAGG-3′ (forward) and 5′-CTCTTGCTCTCAGATCCTTGCTGG-3′ (reverse); U6, 5′-CTCGCTTCGGCAGCACA-3′ (forward) and 5′-AACGCTTCACGAATTTGCGT-3′ (reverse).
Western blot
Cell proteins were extracted by RIPA buffer (Sigma, St Louis, MO, USA) with Complete Protease Inhibitor Cocktail (Beyotime Biotechnology, Beijing, China) and quantified using bicinchoninic acid protein assay kit (Pierce, Rockford, IL, USA). Equal amounts of protein (50 μg protein per lane) were loaded on 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis for separating them and then transferred to polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA, USA). After blocking with 5% nonfat milk for 1 h, the membranes were probed with primary antibodies against BCL2 and GAPDH (Santa Cruz Biotech, Santa Cruz, CA, USA) at 4°C overnight. Finally, the membranes were extensively washed and incubated with horseradish peroxidase-labelled goat anti-rabbit IgG antibody (Santa Cruz Biotech) at room temperature for 2 h. The protein bands were visualized using an enhanced chemiluminescence Western blotting detection system (Bio-Rad Laboratories Inc., Hercules, CA, USA).
Cell proliferation assay
Transfected OVCAR3 or SKOV3 cells (2,000 cells/well) were seeded in 96-well plates in normal growth media and cell viability was determined after incubating for 0, 24, 48, or 72 h. In brief, MTT solution (20 μL, 5 mg/mL; Sigma) was added to each well, followed by incubation at 37°C for another 4 h. The generated formazan crystals were dissolved using dimethyl sulfoxide (150 μL; Sigma). The optical density was measured at 490 nm using a microplate spectrophotometer (Thermo Fisher Scientific).
Luciferase reporter assay
NEAT1 fragment or 3′-untranslated region (3′-UTR) of BCL2 containing the putative miR-34a-5p binding sites was synthesized and then cloned into the KpnI and BglII sites of pGL3-Basic (Promega, Madison, WI, USA) to generate the wild reporter plasmids (WT-NEAT1 or WT-BCL2–3′-UTR). The mutant reporter plasmids (MUT-NEAT1 or MUT-BCL2–3′-UTR) containing the mutated putative binding sequences of miR-34a-5p were obtained through site-specific mutagenesis. OVCAR3 or SKOV3 cells were cotransfected with miR-34-a-5p or miR-NC and the constructed luciferase reporter vectors by using Lipofectamine 2000 (Thermo Fisher Scientific). At 48 h posttransfection, the dual luciferase reporter assay system (Promega) was used to detect luciferase activity, which was normalized against renilla luciferase activity.
Caspase-3 activity assay
Caspase-3 activity was assessed using the caspase-3 activity assay kit (Beyotime Biotechnology). Transfected OVCAR3 or SKOV3 cells were cultured in 96-well plates for 48 h and then treated with iced lysis buffer for 10 min. After centrifugation at 10,000× g for 1 min, the supernatant was incubated with 100 μL reaction buffer containing caspase-3 substrate (Ac-DEVD-pNA) at 37°C for 2 h. The caspase-3 activity was detected at 405 nm wavelength using VICTOR-X3 Multi-label Plate Reader (Perkin Elmer, Santa Clara, CA, USA).
Flow cytometry analysis
To detect apoptosis, transfected OVCAR3 or SKOV3 cells were harvested, washed twice, and resuspended in binding buffer. Then, the cells were stained using an Annexin V-fluorescein isothiocyanate Apoptosis Detection Kit (BD Biosciences, Franklin Lakes, NJ, USA), following which they were subjected to FACSan flow cytometry (BD Biosciences) to analyze apoptotic distribution.
To determine the distribution of cells in the different phases of the cell cycle, the treated OVCAR3 or SKOV3 cells were washed with PBS and fixed in 70% ice-cold ethanol at 4°C for 24 h. After treating with 0.5 mg/mL RNase A for 30 min at 37°C, cells were stained with propidium iodide for 30 min. The percentage of the cells in G0/G1, S, and G2/M phases was analyzed by FACSan flow cytometry (BD Biosciences) with FlowJo software (Tree Star Corp., Ashland, OR, USA).
Statistical analysis
All results were expressed as mean ± SD. All statistical analyses were carried out using Student’s t-test or one-way analysis of variance by SPSS 16.0 software (SPSS Inc., Chicago, IL, USA). P-values <0.05 were considered statistically significant.
Results
NEAT1 promoted cell proliferation in OC
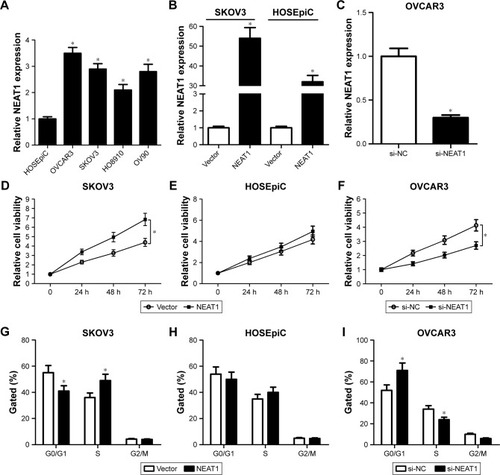
To confirm the biologic role of NEAT1 in the development and progression of OC, the expression level of NEAT1 in OC cells was evaluated by qRT-PCR. As shown in , NEAT1 expression was exceptionally higher in OC cell lines (OVCAR3, SKOV3, HO8910, and OV90) than that in normal ovarian epithelial cell line HOSEpiC. Next, gain-of-function or loss-of function experiments were performed by transfecting NEAT1 into SKOV3 and HOSEpiC cells or by introducing si-NEAT1 into OVCAR3 cells. The transfection efficiency was verified by qRT-PCR. The results showed that introduction of NEAT1 significantly improved NEAT1 expression () in SKOV3 and HOSEpiC cells, and transfection of si-NEAT1 dramatically reduced NEAT1 expression in OVCAR3 cells (). MTT assay showed that ectopic expression of NEAT1 effectively promoted cell proliferation in SKOV3 () and had no effect on HOSEpiC () cells, while NEAT1 knockdown strikingly restrained OVCAR3 cell proliferation (). Cell cycle analysis revealed that NEAT1 overexpression led to a marked decrease of G0/G1 phase and a significant increase of S phase in SKOV3 () and HOSEpiC () cells, suggesting that NEAT1 overexpression promoted OC cell proliferation by increasing the proportion of cells in S phase. In contrast, NEAT1 knockdown resulted in a substantial proportion of cells being arrested in the G0/G1 phase and an obvious reduction in cell number in the S phase in OVCAR3 (), indicating that NEAT1 knockdown blocked OC cells proliferation by blocking their progression from the G0/G1 to S phase. Collectively, these data implied that upregulation of NEAT1 promoted cell proliferation in OC.
Figure 1 Effects of alteration of NEAT1 expression on OC cell proliferation.
Abbreviations: NEAT1, nuclear enriched abundant transcript 1; OC, ovarian cancer; qRT-PCR, quantitative real-time polymerase chain reaction; si-NC, scrambled siRNA negative control.
NETA1 inhibited apoptosis of OC cells
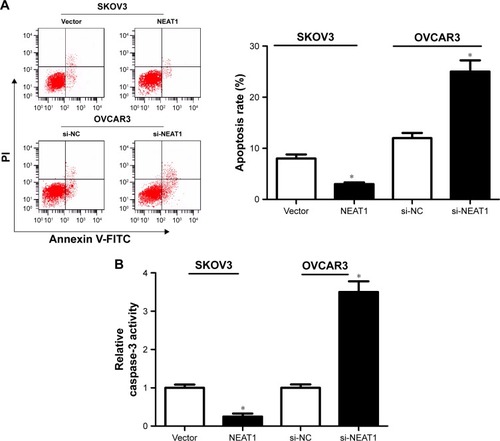
The effect of NEAT1 on apoptosis of OC cells was further analyzed by flow cytometry analysis and caspase-3 activity assay. Flow cytometry analysis implicated that apoptosis in si-NEAT1-transfected OVCAR3 cells was distinctively induced relative to si-NC group, while apoptosis in NEAT1-transfected SKOV3 cells was markedly impeded versus the Vector group (). Caspase-3 activity assay uncovered that NEAT1 knockdown significantly increased caspase-3 activity in OVCAR3 cells compared with si-NC-transfected cells, while exogenous expression of NEAT1 remarkably decreased caspase-3 activity in SKOV3 cells in comparison with Vector-transfected cells (). Together, these results suggested that NEAT1 suppressed apoptosis of OC cells.
Figure 2 Effects of alteration in NEAT1 expression on OC cell apoptosis.
Abbreviations: NEAT1, nuclear enriched abundant transcript 1; OC, ovarian cancer; PI, propidium iodide; si-NC, scrambled siRNA negative control.
miR-34a-5p was a target of NEAT1
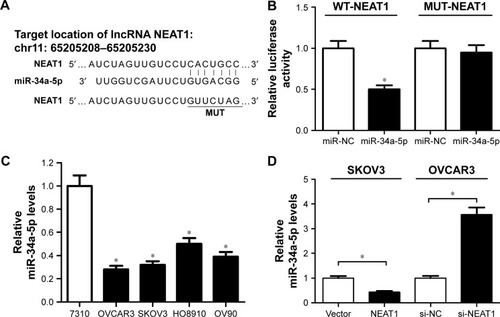
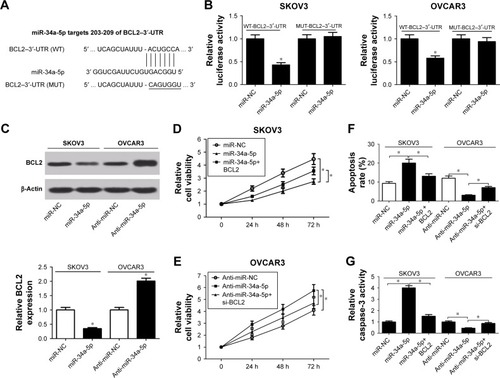
Previous studies reported that NEAT1 served as ceRNAs or molecular sponges for miRNAs to negatively regulate the expression and activities of miRNAs.Citation21 While searching for potential miRNA-containing binding sites within NEAT1 by Starbase 2.0, miR-34a-5p was found to have complementary base pairing with NEAT1 (). To confirm the prediction, luciferase reporter vectors containing the wild-type or mutated miR-34a-5p binding sites on NEAT1 were constructed and cotransfected with miR-34a-5p or miR-NC into OVCAR3 cells. Luciferase reporter assay indicated that miR-34a-5p overexpression effectively reduced luciferase activity of WT-NEAT reporter, but not that of the MUT-NEAT1 reporter (). The expression of miR-34-35p in human OC cell lines was further analyzed by qRT-PCR. As shown in , miR-34a-5p expression was markedly decreased in human OC cell lines (OVCAR3, SKOV3, HO8910, and OV90) compared with normal ovarian epithelial cell line HOSEpiC. To explore the regulatory effect of NEAT1 on miR-34a-5p expression, NEAT1 or si-NEAT1 was transfected into SKOV3 or OVCAR3 cells to overexpress or knock down NEAT1. As presented in , ectopic expression of NEAT1 significantly decreased miR-34a-5p expression in SKOV3 cells, while NEAT1 knockdown markedly increased miR-34a-5p expression in OVCAR3 cells.
Figure 3 Relationship between NEAT1 and miR-34a-5p.
Abbreviations: miR-34a-5p, miR-34a-5p mimic; miR-NC, miRNA negative control; MUT-NEAT1, mutant reporter plasmid-NEAT1; NEAT1, nuclear enriched abundant transcript 1; OC, ovarian cancer; qRT-PCR, quantitative real-time polymerase chain reaction; WT-NEAT1, wild reporter plasmid-NEAT1.
NEAT1 overexpression promoted proliferation and inhibited apoptosis of OC cells by negatively regulating miR-34a-5p expression
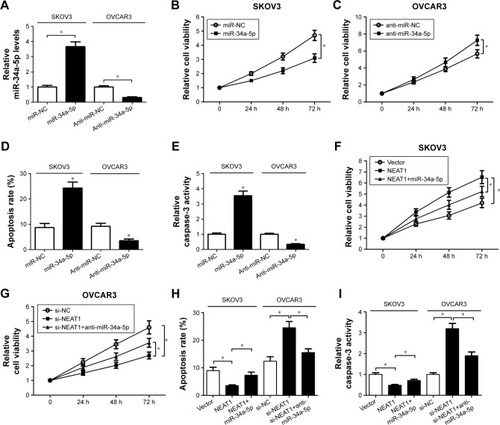
The role of miR-34a-5p in OC cell proliferation and apoptosis was explored. miR-34a-5p mimic was transfected into SKOV3 cells or anti-miR-34a-5p was transfected into OVCAR3 cells. As shown in , miR-34a-5p expression was successfully upregulated in SKOV3 cells following miR-34a-5p mimic transfection; conversely, miR-34a-5p level was substantially downregulated in OVCAR3 cells after anti-miR-34a-5p treatment. The results of MTT assay demonstrated that miR-34a-5p overexpression significantly suppressed proliferation in SKOV3 cells (), while miR-34a-5p inhibition led to an obvious improvement in OVCAR3 cell proliferation (). Flow cytometry results indicated that the apoptotic rate of SKOV3 cells was enhanced by enforced expression of miR-34a-5p; in contrast, anti-miR-34a-5p-transfected OVCAR3 cells showed a lower apoptosis, compared to the corresponding control group (). As expected, caspase-3 activity assay implied that forced expression of miR-34a-5p markedly improved caspase-3 activity of SKOV3 cells (), while down-regulation of miR-34a-5p repressed caspase-3 activity of OVCAR3 cells when compared with the respective control group. To further investigate whether the effects of NEAT1 on OC cell proliferation and apoptosis were mediated by miR-34a-5p, we conducted a rescue experiment by transfecting SKOV3 cells with NEAT1 or NEAT1+miR-34a-5p or by transfecting OVCAR3 cells with si-NEAT1 or si-NEAT1+anti-miR-34a-5p. As illustrated in , cell proliferation was remarkably improved in SKOV3 cells treated with NEAT1, whereas ectopic expression of miR-34a-5p strikingly attenuated the effect. Inversely, NEAT1 knockdown blocked OVCAR3 cell proliferation, which reduced after cotransfection with anti-miR-34a-5p (). In addition, overexpression of miR-34a-5p partly reversed NEAT1-mediated decline in the apoptotic rate of SKOV3 cells, and reintroduction of anti-miR-34a-5p also overturned si-NEAT1-induced apoptosis in OVCAR3 cells (). As might be expected, caspase-3 activity assay showed similar results with apoptosis in SKOV3 and OVCAR3 cells (). Collectively, these results demonstrated that NEAT1 overexpression contributed to proliferation and inhibited apoptosis of OC cells by targeting miR-34a-5p.
Figure 4 Effect of NEAT1 on proliferation and apoptosis of OC cells was mediated by miR-34a-5p.
Abbreviations: miR-34a-5p, miR-34a-5p mimic; NEAT1, nuclear enriched abundant transcript 1; OC, ovarian cancer; qRT-PCR, quantitative real-time polymerase chain reaction.
miR-34a-5p overexpression suppressed proliferation and triggered apoptosis of OC cells by targeting BCL2
Since miRNAs exerted their functions by binding to the 3′-UTR of specific genes, the potential targets of miR-34a-5p were predicted using the TargetScan tool (www.targetscan.org). As shown in , BCL2 was predicted to contain complementary sites for miR-34a-5p targeted binding. To further confirm the authentic binding between miR-34a-5p and BCL2, luciferase reporter plasmids containing the wild-type or mutated miR-34a-5p binding region in the 3′-UTR of BCL2 were established, and then luciferase reporter assay was used to detect luciferase activity. The results showed that overexpression of miR-34a-5p significantly reduced luciferase activity of WT-BCL2–3′-UTR reporter in both SKOV3 and OVCAR3 cells, but had no inhibitory effect on the luciferase activity of MUT-BCL2–3′-UTR reporter (). To analyze the actual effect of miR-34a-5p on BCL2 expression, Western blot was performed to determine the protein level of BCL2 in SKOV3 treated with miR-34a-5p and OVCAR3 cells transfected with anti-miR-34a-5p. As shown in , the protein level of BCL2 was significantly decreased in miR-34-a-5p-transfected SKOV3 cells and obviously increased in anti-miR-34a-5p-transfected OVCAR3 cells, suggesting that miR-34a-5p had a negative effect on regulating BCL2 level. Subsequently, the effects of interaction of miR-34a-5p and BCL2 on OC cell proliferation and apoptosis were investigated by MTT assay, flow cytometry, and caspase-3 activity assay in SKOV3 cells transfected with miR-NC, miR-34a-5p, or miR-34a-5p+BCL2, as well as in OVCAR3 cells treated with anti-miR-NC, anti-miR-34a-5p, or anti-miR-34a-5p+si-BCL2. MTT assay showed that ectopic expression of miR-34a-5p significantly suppressed proliferation in SKOV3 cells, while overexpression of BCL2 overturned miR-34a-5p–induced inhibition of cell proliferation (). Meanwhile, knockdown of miR-34a-5p greatly promoted OVCAR3 cell proliferation, which was attenuated by cotransfection of ani-miR-34a-5p and si-BCL2 (). As demonstrated by flow cytometry in , apoptotic rate in miR-34a-5p-transfected SKOV3 cells was substantially enhanced, which was reduced by BCL2 overexpression, while apoptosis in anti-miR-34a-5p-transfected OVCAR3 cells was repressed, which was partly reversed by BCL2 knockdown. Moreover, as expected, caspase-3 activity assay further validated the promoting effect of miR-34a-5p-BCL2 axis on OC cell apoptosis (). Taken together, these data suggested that miR-34a-5p overexpression hindered proliferation and induced apoptosis of OC cells by targeting BCL2.
Figure 5 Effect of miR-34a-5p on proliferation and apoptosis of OC cells was modulated by BCL2.
Abbreviations: BCL2, B-cell lymphoma-2; miR-34a-5p, miR-34a-5p mimic; miR-NC, miRNA negative control; NEAT1, nuclear enriched abundant transcript 1; OC, ovarian cancer; UTR, untranslated region.
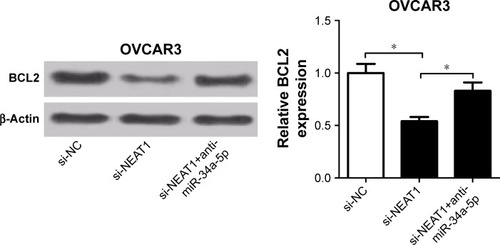
NEAT1 upregulated BCL2 expression via miR-34a-5p in OC cells
To investigate whether NEAT1 influenced BCL2 expression by regulating miR-34a-5p, Western blot was used to determine the protein level of BCL2 in OVCAR3 cells trans-fected with si-NEAT1, or along with anti-miR-34a-5p. The results showed that NEAT1 knockdown significantly reduced the protein level of BCL2 in OVCAR3 cells, whereas anti-miR-34a-5p transfection dramatically abrogated the inhibitory effect of si-NEAT1 on BCL2 expression (), indicating that NEAT1 upregulated BCL2 expression via miR-34a-5p in OC cells.
Figure 6 si-NEAT1-induced decrease in BCL2 expression was abated after introduction of miR-34a-5p in OVCAR3 cells.
Abbreviations: BCL2, B-cell lymphoma-2; miR-34a-5p, miR-34a-5p mimic; NEAT1, nuclear enriched abundant transcript 1; si-NC, scrambled siRNA negative control; si-NEAT1, siRNA against NEAT1.
Discussion
lncRNAs play a pivotal role in many biologic processes, and abnormal expression of lncRNAs is associated with the carcinogenesis and development of malignant tumors.Citation22 This study focused on the involvement of NEAT1 in OC and its possible molecular mechanism. Here, it is demonstrated that NEAT1 was highly expressed in OC cells and promoted proliferation and suppressed apoptosis in OC cells. Mechanistic analysis revealed that NEAT1’s function as an oncogenic lncRNA was partially attributed to its ability to serve as a molecular sponge of miR-34a-5p, leading to upregulation of BCL2. Thus, this study not only uncovers the important role of NEAT1/miR-34a-5p/BCL2 axis in OC tumorigenesis, but also makes a contribution to elucidating the significance of nonannotated lncRNA species in the field of cancer research.
NEAT1, an important cancer-associated lncRNA, was found to be upregulated and to play an oncogenic role in most types of solid tumors, including lung cancer, esophageal cancer, laryngeal cancer, colorectal cancer, hepatocellular cancer, prostate cancer, breast cancer, and glioma.Citation23 For instance, NEAT1 expression was significantly elevated in NSCLC tissues and contributed to oncogenesis and progression of NSCLC by activating Wnt/β-catenin signaling pathway.Citation24 Highly expressed NEAT1 promoted cell growth, invasion, and migration and attenuated apoptosis in endometrial endometrioid adenocarcinoma cells.Citation25 Elevated expression of NEAT1 was correlated with poor prognosis, stimulated cell proliferation, and promoted migration and invasion in esophageal squamous cell carcinoma cells.Citation26 These findings corroborated with our study findings, which revealed that NEAT1 was upregulated in OC cells, and overexpression of NEAT1 promoted cell proliferation and suppressed apoptosis of OC cells, while knockdown of NEAT1 exerted the opposite effect. In accordance with our results, NEAT1 was overexpressed in OC tissues, and upregulated NEAT1 expression could serve as a promising marker for tumor progression and prognosis in OC.Citation27 Moreover, enforced expression of NEAT1 was demonstrated to promote OC cell proliferation and invasion, consistent with our finding.Citation15
It is well known that lncRNAs are involved in ceRNA regulatory system, wherein lncRNAs function as miRNA sponges to negatively modulate miRNAs, thus derepressing miRNA-specific targets at the posttranscriptional regulation.Citation17 To further explore the mechanism of NEAT1 in OC progression, potential miRNAs of NEAT1 were searched by Starbase 2.0. As a result, NEAT1 was confirmed to directly interact with miR-34a-5p. Overexpression of miR-34a-5p impeded OC cell proliferation and increased apoptosis, while knockdown of miR-34a-5p improved OC cell growth and inhibited apoptosis. Furthermore, rescue experiments revealed that NEAT1 regulated OC cell proliferation and apoptosis by sponging miR-34a-5p. In agreement with our data, the tumor-suppressive role of miR-34a has been reported in several cancers, such as cervical cancer,Citation28 hepatocellular carcinoma,Citation29 and breast cancer.Citation30 Also, miR-34a was documented to suppress OC cell proliferation and motility by targeting receptor tyrosine kinase.Citation20 Next, BCL2, the most important antiapoptotic oncogene, was identified as a direct target of miR-34a-5p, and miR-34a-5p overexpression suppressed cell proliferation and induced apoptosis in OC cells by binding to BCL2. Similarly, in meningioma, miR-34a-3p was indicated to modulate cell proliferation and apoptosis by targeting BCL2.Citation31 Finally, the relationship among NEAT1, miR-34a-5p, and BCL2 was explored, and it was found that downregulation of NEAT1 suppressed BCL2 expression; however, this inhibitory effect was abrogated by miR-34a-5p knockdown, suggesting that NEAT1 regulated BCL2 expression by sponging miR-34a-5p.
Conclusion
This study provided the first evidence that NEAT1 regulated the proliferation and apoptosis of OC cells through miR-34-a-5p/BCL2 axis, elucidating the underlying mechanism of NEAT1 as a carcinogenic factor in OC. Therefore, NEAT1/miR-34a-5p/BCL2 axis played a crucial role in the pathogenesis and development of OC, providing a potential therapeutic approach for OC patients.
Disclosure
The authors report no conflicts of interest in this work.
References
- ZhongpingCJingGLiCNingLWeihongYXiaoyanQA long noncoding RNA AB0,73,614 promotes tumorigenesis and predicts poor prognosis in ovarian cancerOncotarget2015628253812538926299803
- JemalABrayFCenterMMFerlayJWardEFormanDGlobal cancer statisticsCA Cancer J Clin2011612699021296855
- SiegelRNaishadhamDJemalACancer statistics, 2013CA Cancer J Clin2013631113023335087
- ShapiraIOswaldMLovecchioJCirculating biomarkers for detection of ovarian cancer and predicting cancer outcomesBr J Cancer2014110497698324366298
- EllisBCGrahamLDMolloyPLCRNDE, a long non-coding RNA responsive to insulin/IGF signaling, regulates genes involved in central metabolismBiochim Biophys Acta20141843237238624184209
- MaLBajicVBZhangZOn the classification of long non-coding RNAsRNA Biol201310692593323696037
- MartensuzunovaESBöttcherRCroceCMJensterGVisakorpiTCalinGALong noncoding RNA in prostate, bladder, and kidney cancerEur Urol20136561140115124373479
- GibbEABrownCJLamWLThe functional role of long non-coding RNA in human carcinomasMol Cancer20111013821489289
- ZhouSWangJZhangZAn emerging understanding of long noncoding RNAs in kidney cancerJ Cancer Res Clin Oncol2014140121989199524816785
- SunwooHDingerMEWiluszJEAmaralPPMattickJSSpectorDLMEN epsilon/beta nuclear-retained non-coding RNAs are up-regulated upon muscle differentiation and are essential components of paraspecklesGenome Res200919334735919106332
- ClemsonCMHutchinsonJNSaraSAAn architectural role for a nuclear non-coding RNA: NEAT1 RNA is essential for the structure of paraspecklesMole Cell2009336717726
- PanLJZhongTFTangRXUpregulation and clinicopathological significance of long non-coding NEAT1 RNA in NSCLC tissuesAsian Pac J Cancer Prev20151672851285525854373
- ZhenLYun-HuiLHong-YuDJunMYi-LongYLong noncoding RNA NEAT1 promotes glioma pathogenesis by regulating miR-449b-5p/c-Met axisTumor Biol2016371673683
- HaoKZhaoLXuFNEAT1 is required for survival of breast cancer cells through FUS and miR-548Gene Regul Syst Bio201610Suppl 11117
- ChaiYLiuJZhangZLiuLHuR-regulated lncRNA NEAT1 stability in tumorigenesis and progression of ovarian cancerCancer Med2016571588159827075229
- YoonJHAbdelmohsenKGorospeMFunctional interactions among microRNAs and long noncoding RNAsSemin Cell Dev Biol201434491424965208
- SalmenaLPolisenoLTayYKatsLPandolfiPPA ceRNA hypothesis: the Rosetta stone of a hidden RNA language?Cell2011146335335821802130
- WangYXuZJiangJEndogenous miRNA sponge lincRNA-RoR regulates Oct4, Nanog, and Sox2 in human embryonic stem cell self-renewalDev Cell2013251698023541921
- ErgunSOztuzcuSOncocers: ceRNA-mediated cross-talk by sponging miRNAs in oncogenic pathwaysTumor Biol201536531293136
- LiRShiXLingFMiR-34a suppresses ovarian cancer proliferation and motility by targeting AXLTumor Biol201536972777283
- TanJQiuKLiMLiangYDouble-negative feedback loop between long non-coding RNA TUG1 and miR-145 promotes epithelial to mesenchymal transition and radioresistance in human bladder cancer cellsFEBS Lett201558920 Pt B3175318126318860
- HaemmerleMGutschnerTLong non-coding RNAs in cancer and development: where do we go from here?Int J Mol Sci20151611395140525580533
- YuXLiZZhengHChanMTWuWKNEAT1: a novel cancer-related long non-coding RNACell Prolif2017502
- SunSJLinQMaJXShiWWYangBLiFLong non-coding RNA NEAT1 acts as oncogene in NSCLC by regulating the Wnt signaling pathwayEur Rev Med Pharmacol Sci201721350451028239820
- LiZWeiDYangCSunHLuTKuangDOverexpression of long noncoding RNA, NEAT1 promotes cell proliferation, invasion and migration in endometrial endometrioid adenocarcinomaBiomed Pharmacother20168424425127664948
- ChenXKongJMaZGaoSFengXUp regulation of the long non-coding RNA NEAT1 promotes esophageal squamous cell carcinoma cell progression and correlates with poor prognosisAm J Cancer Res2015592808281526609486
- ChenZJZhangZXieBBZhangHYClinical significance of up-regulated lncRNA NEAT1 in prognosis of ovarian cancerEur Rev Med Pharmacol Sci201620163373337727608895
- GengDSongXNingFSongQYinHMiR-34a inhibits viability and invasion of human papillomavirus-positive cervical cancer cells by targeting E2F3 and regulating survivinInt J Gynecol Cancer201525470771325675046
- SunTYXieHJZhenLmiR-34a regulates HDAC1 expression to affect the proliferation and apoptosis of hepatocellular carcinomaAm J Transl Res20179110311428123637
- SiWLiYShaoHMiR-34a inhibits breast cancer proliferation and progression by targeting Wnt1 in Wnt/β-catenin signaling pathwayAm J Med Sci2016352219119927524218
- WernerTVHartMNickelsRMiR-34a-3p alters proliferation and apoptosis of meningioma cells in vitro and is directly targeting SMAD4, FRAT1 and BCL2Aging (Albany NY)20179393295428340489