
Abstract
Background
Although the relationship between several single nucleotide polymorphisms (SNPs) of the oncogene EZH2 and cancer risk has been assessed by some case–control studies, results of subsequent studies are controversial. Sample sizes from single-center studies are also limited, thereby providing unreliable findings. Hence, we conducted a comprehensive search and meta-analysis to evaluate the associations between EZH2 SNPs and cancer risk.
Materials and methods
A comprehensive literature search for studies focusing on EZH2 SNPs and cancer risk was conducted on PubMed, Web of Science, Embase, and China National Knowledge Infrastructure online databases. Genotype data were extracted and examined through a meta-analysis, and pooled odds ratios (ORs) with 95% CIs were used to assess the corresponding associations. Sensitivity analysis, publication bias assessment, and heterogeneity test were performed using STATA 12.0.
Results
Twelve eligible studies were included in this meta-analysis. The association of 4 SNPs, namely, rs887569, rs2302427, rs3757441, and rs41277434, in the EZH2 locus with cancer risk was evaluated. Five studies (1,794 cases and 1,878 controls) indicated that rs887569 was related to a decreased cancer risk (CTTT/CC: OR =0.849, 95% CI: [0.740 to 0.973], P=0.019; TT/CCCT: OR =0.793, 95% CI: [0.654 to 0.962], P=0.019). Seven studies (2,408 cases and 2,910 controls) showed that rs2302427 was linked to a decreased cancer risk (GG/CC: OR =0.562, 95% CI: [0.400 to 0.792], P=0.001; CGGG/CC: OR =0.856, 95% CI: [0.748 to 0.980], P=0.024; GG/CCCG: OR =0.733, 95% CI: [0.571 to 0.940], P=0.015). No relationships were observed between rs3757441 or rs41277434 and cancer risk.
Conclusion
rs887569 and rs2302427 in EZH2 may be correlated with a decreased cancer risk. Although rs3757441 and rs41277434 are independent risk factors of cancer, further large-scale and functional studies are warranted to validate our findings.
Introduction
Approximately 1,688,780 new cancer cases and 600,920 cancer deaths are projected to occur in the USA in 2017.Citation1 Cancer is caused by uncontrolled cell division or inappropriate survival of a cell with DNA damage, which is critical for tumor initiation and progression.
Thousands of genes that are either transcriptionally upregulated or downregulated in tumor samples have been identified through microarray analysis, indicating that cancer is a disease with extreme heterogeneity. These deregulations act as the main drivers that enable tumors to invade cellular barriers, proliferate, and metastasize.Citation2 The dynamic regulation of histone modifications in promoters and enhancers plays a vital role in the control of gene expression and consequently affects disease susceptibility. EZH2 has been widely investigated because it serves as a master regulator of cancer epigenetics.Citation3 It is also a core component of Polycomb repressive complex 2, which mainly methylates lysine 27 of histone H3 (H3K27) to induce transcriptional gene silencing.Citation4 EZH2 overexpression causes epigenetic alterations in tumor suppressor genes, and such changes are required for cancer proliferation, migration, invasion, and metastasis.Citation5–Citation7 Therefore, aberrant EZH2 activities may participate in increasing the risk of tumorigenesis.
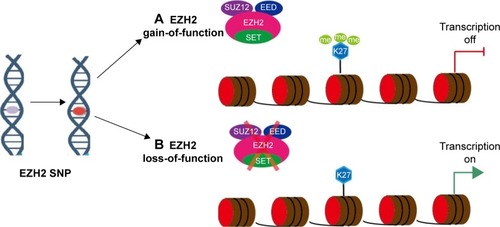
The oncogenic role of EZH2 has been observed in numerous cancers, including prostate cancer, bladder cancer, breast cancer, and melanoma, whose high EZH2 expression levels are positively correlated with poor survival rate and aggressiveness.Citation8–Citation11 The function of EZH2 in cancer progression may also be affected by mutations. For example, the mutation of tyrosine 641 (Y641) within the C-terminal catalytic SET domain of EZH2 increases the levels of trim-ethylated H3K27 (H3K27me3) and thus represses the expression of Polycomb targets.Citation12 The loss-of-function mutations of EZH2 may occur during cancer development. The frequency of missense mutations of EZH2 in the pediatric subtype of human T-cell acute lymphoblastic leukemia (T-ALL) and early T-cell precursor (ETP) ALL is higher than that in non-ETP pediatric T-ALL.Citation13,Citation14 Similarly, single nucleotide polymorphisms (SNPs) of EZH2 may have different effects on disease susceptibility through the transcriptional regulation of genes involved in cancer initiation and progression (). Although several studies have investigated the relationship of 4 SNPs (rs887569 C>T, rs2302427 C>G, rs3757441 T>C, and rs41277434 A>C) of EZH2 and cancer risk, results are inconsistent. This relationship has yet to be systematically investigated, and definitive conclusions have yet to be presented. Hence, comprehensive reviews and meta-analyses should be performed. Here, we conducted a meta-analysis to precisely assess and provide a comprehensive conclusion about the associations between EZH2 variations and cancer risk from all eligible case–control studies published to date.
Figure 1 EZH2 polymorphism affects transcription of downstream targets.
Materials and methods
Search strategy and identification of eligible studies
Two reviewers (Ling and You) searched the online databases PubMed, Google Scholar, Web of Science, Embase, China National Knowledge Infrastructure (CNKI), and Wangfang Data to identify relevant articles published until September 2017. The following search terms were used either separately or in combination: “EZH2, enhancer of zeste homolog 2,” “rs887569, rs2302427, rs3757441, rs41277434,” “cancer, carcinoma, neoplasm,” “tumor, tumour,” and “SNP, polymorphism, allele, variation.” Studies were limited to articles published in Chinese or English, and the references of pertinent articles were manually screened and checked. Articles that satisfied the following criteria were included: 1) studies that assessed the association between a SNP from EZH2 (rs887569, rs2302427, rs3757441, and rs41277434) and cancer risk; 2) case–control or population-based studies; and 3) studies with available genotype frequencies. Studies were excluded according to the following criteria: 1) articles that were presented as a systematic review or focusing on animals; 2) studies that involved DNA extracted from cancer tissues rather than blood samples, or studies that did not provide usable data for meta-analysis; and 3) studies that reported data overlapping with those described in the included studies.
Data extraction
Two reviewers (Ling and You) independently extracted the following information from each study: first author, year of publication, cancer types, country or region, ethnicity, genotype detection method, control source of each study, number of cases and controls, polymorphism site included in each study, and results of Hardy–Weinberg equilibrium (HWE). Inconsistencies were resolved by discussion until a consensus was obtained. Newcastle–Ottawa Quality Assessment Scale was used to examine the quality of the articles included in this study.Citation15
Statistical analysis
The strength of the association between SNPs and cancer risk was evaluated by determining the odds ratio (OR) with 95% CI, which was calculated by Z-test, and the result of the pooled OR was considered significant when P<0.05. This association was also examined by using homozygote, heterozygote, dominant genetic, and recessive genetic models. Subgroup analyses were conducted according to cancer types and ethnic groups. Heterogeneity between articles was identified with Q-test and I2 index.Citation16 When heterogeneity was observed (P<0.05 or I2>50%), a random-effect model (DerSimonian–Laird method) was applied; otherwise, a fixed-effect model (Mantel–Haenszel method) was utilized.Citation17,Citation18 Publication bias was evaluated by Egger’s test and Begg’s test, with a P-value >0.05 considered evidence for no potential publication bias. Begg’s or Egger’s test was performed only for SNPs involved in 5 or more studies. Statistical tests were 2-sided, and analyses were carried out with Stata 12.0 at least twice.
Results
Characteristics of the included studies
After PubMed, Google Scholar, Web of Science, Embase, CNKI, and Wangfang Data online databases were extensively screened, 216 relevant articles were identified. As shown in the flowchart in , 12 case–control studies involving the 4 EZH2 SNPs were finally included for further meta-analysis after ineligible articles were excluded according to our inclusion and exclusion criteria.Citation19–Citation30 The characteristics of the included studies are summarized in . Of the 12 included studies, 6 focused on digestive system cancers (DSCs; gastric cancer, hepatocellular carcinoma, colorectal cancer [CRC], and esophageal squamous cell carcinoma), 4 examined urogenital system cancers (USCs; prostate cancer, urothelial cell carcinoma, and bladder cancer), and 2 investigated other types of cancers. The detailed information of the analyzed articles for each SNP is shown in .
Table 1 Characteristics of studies included in the meta-analysis
Figure 2 Studies identified with criteria of inclusion and exclusion.

Quantitative synthesis
The associations between EZH2 SNPs and human cancer risks were evaluated (; and ). Overall, the EZH2 rs887569 C>T polymorphism was significantly associated with a decreased cancer risk in the dominant and recessive models (CTTT/CC: OR =0.849, 95% CI: [0.740 to 0.973], P=0.019; TT/CCCT: OR =0.793, 95% CI: [0.654 to 0.962], P=0.019). EZH2 rs2302427 C>G polymorphism was also related to the decreased overall cancer risk in the homozygote dominant genetic and recessive genetic models (GG/CC: OR =0.562, 95% CI: [0.400 to 0.792], P=0.001; CGGG/CC: OR =0.856, 95% CI: [0.748 to 0.980], P=0.024; GG/CCCG: OR =0.733, 95% CI: [0.571 to 0.940], P=0.015). In other genotype models, such a relationship remains controversial.
Table 2 Analysis of associations between SNPs of EZH2 and cancer risk
Figure 3 Forest plot for the relationship between rs887569 and cancer risk: (A) CT/CC; (B) TT/CC; (C) CTTT/CC; (D) TT/CCCT.
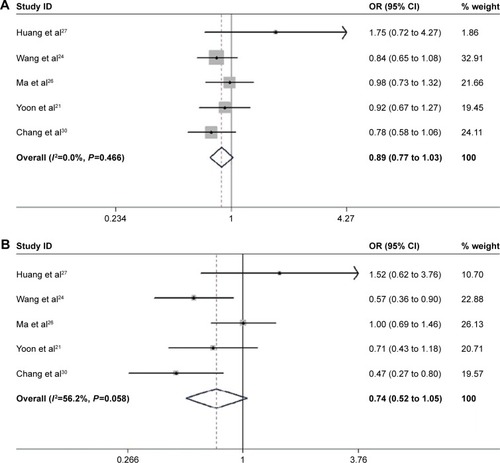
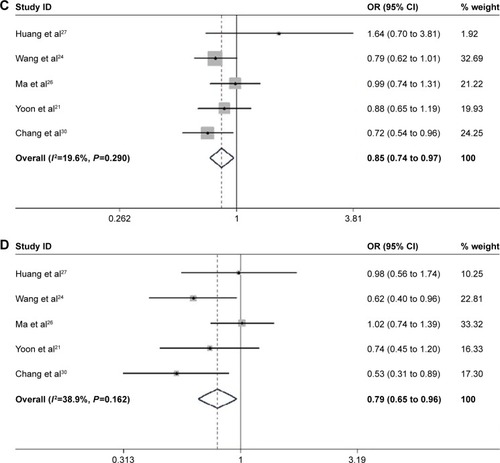
Figure 4 Forest plot for the relationship between rs2302427 and cancer risk: (A) CG/CC; (B) GG/CC; (C) CGGG/CC; (D) GG/CCCG.
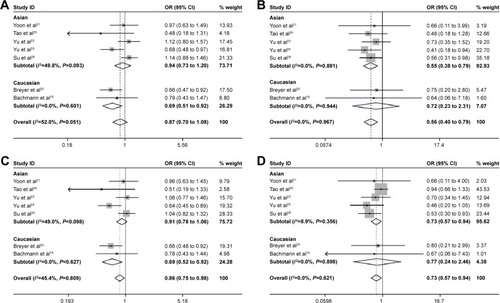
Subgroup analysis revealed that the variant CG (OR =0.686, 95% CI: [0.511 to 0.921], P=0.012) and CG/GG (OR =0.688, 95% CI: [0.515 to 0.917], P=0.01) genotypes of rs2302427 C>G polymorphism were associated with a decreased cancer risk compared with the wild-type CC genotype in individuals of Caucasian descent. rs2302427 C>G polymorphism in Asian descent was linked to the decreased overall cancer risk in the homozygote and recessive genetic models (GG/CC: OR =0.550, 95% CI: [0.384 to 0.787], P=0.001; GG/CCCG: OR =0.731, 95% CI: [0.566 to 0.944], P=0.016).
We also conducted a stratified analysis of the data in terms of cancer types, namely, USCs and DSCs. With regard to subgroup analysis of USCs, our results did not show any association of rs887569 C>T polymorphism with cancer risk in any genotype model. However, rs2302427 C>G polymorphism was correlated with a decreased cancer risk in homozygote and recessive genetic models for DSCs. As for USCs, similar results were observed in homozygote, heterozygote, and dominant genetic models.
For rs3757441 T>C and rs41277434 A>C polymorphisms, 9 and 7 studies were included, respectively. No evidence suggested that these 2 SNPs might be associated with cancer risk either in overall or subgroup analysis (P>0.05; ; and ).
Sensitivity analysis and publication bias assessment
Sensitivity analyses were conducted by omitting each individual article to measure its specific effect on the pooled ORs (). The sensitivity analysis forest plot indicated that no single study significantly affected the pooled ORs for any genetic models of the 4 SNPs. A random-effect model was used when obvious heterogeneity was observed (P<0.05 or I2>50%); otherwise, a fixed-effect model was applied. Considering the small number of studies included in the meta-analysis, we conducted Begg’s and Egger’s tests to assess the publication bias for each genetic model of the 4 SNPs. No evidence of publication bias was detected in any of the homozygote, heterozygote, and dominant and recessive models of each SNP except rs3757441 and rs41277434 ().
Table 3 Publication bias in meta-analysis for each inheritance model
Discussion
EZH2 overexpression is a marker of advanced and metastatic diseases in many solid tumors, including prostate,Citation8 bladder,Citation31 gastric,Citation32 lung,Citation33 and breast cancer.Citation34 EZH2 has also been implicated in cancer initiation, promotion, and progression.Citation35 Therefore, genetic mutations may significantly influence the function of EZH2 in cancer initiation and risk.Citation36 Cumulative studies have suggested that recurrent heterozygous point mutations affecting tyrosine 641 (Y641) in germinal center B-cell and point mutations at alanine 687 or 677 in non-Hodgkin’s lymphomas can increase H3K27me3 levels, thereby repressing the expression of Polycomb targets.Citation37–Citation39
SNPs, as the most common genetic sequence variation, can affect the function of EZH2 and its downstream targets by altering EZH2 transcription and H3K27 trimethylation. For example, the rs3757441 polymorphism C/C genotype is associated with strong EZH2 and H3K27me3 immunoreactivity in primary CRC, indicating that this genotype can be a promising biomarker for EZH2-targeting agents.Citation27 The rs887569 TT genotype is correlated with a significantly increased overall survival and a reduced risk of mortality in patients with cholangiocarcinoma.Citation40 Zhou et alCitation22 found that the haplotypes of EZH2 genes with minor alleles of rs12670401 and rs6464926 or major alleles of rs2072407, rs734005, and rs734004 significantly increase the risk of gastric cancer, whereas the haplotypes of EZH2 genes with major alleles of rs12670401 and rs6464926 or minor alleles of rs2072407, rs734005, and rs734004 can reduce the risk of gastric cancer. These studies have demonstrated that the SNPs of EZH2 are closely related to cancer risk and prognosis. Although studies have revealed that EZH2 polymorphisms are associated with cancer risk, results are inconsistent. Therefore, we systematically reviewed the literature through a meta-analysis of the association between EZH2 gene polymorphisms and cancer risk. To the best of our knowledge, this study is the first meta-analysis to investigate the relationship between EZH2 SNPs and cancer risk.
While searching for eligible studies, we found 11 EZH2 SNPs that were reported to be associated with cancer risk: rs887569, rs2302427, rs375441, rs41277434, rs6950683, rs2072407, rs734005, rs734004, rs6464926, rs12670401, and rs1880357. However, only the first 4 SNPs were examined in at least 5 individual studies. We then performed 4 genotype distributions between cases and controls. Our study included 5 articles, with a pooled total of 1,794 cases and 1,878 controls, which were relevant to the relationship between the rs887569 SNP and cancer risk. The cancer risk was significantly reduced in CT/TT genotype relative to CC genotype (CTTT/CC: OR =0.849, 95% CI: [0.740 to 0.973], P=0.019). This association was also detected in the recessive genetic model (TT/CCCT: OR =0.793, 95% CI: [0.654 to 0.962], P=0.019). Z-scores and P-values were calculated to evaluate the reliability of our results, and the P-values of the dominant and recessive genetic models of rs887569 were 0.019, which might strengthen our findings. We also found a significant link between rs2302427 polymorphism and cancer susceptibility in the homozygote genotype, dominant genetic, and recessive genetic models (GG/CC: OR =0.562, 95% CI: [0.400 to 0.792], P=0.001; CGGG/CC: OR =0.856, 95% CI: [0.748 to 0.980], P=0.024; GG/CCCG: OR =0.733, 95% CI: [0.571 to 0.940], P=0.015). In the subgroup analysis of ethnicity, rs2302427 CG or CG/GG genotype was significantly related to a decreased prostate cancer risk in the Caucasian population, whereas the GG genotype was closely linked to a decreased overall cancer risk in the Asian population. However, the reliability of our data would have improved had we enrolled more eligible studies and a larger sample size than the obtained data.
We subsequently examined the effect of EZH2 SNP rs3757441, which is a key indicator of poor prognosis in metastatic CRC, on overall cancer risk by analyzing 9 eligible studies.Citation27 However, in our current meta-analysis, the association between rs3757441 and cancer risk is controversial. We also performed a stratified analysis by cancer types, but no association was observed between rs3757441 and USC or DSC. These inconsistent results might be due to the heterogeneity of cancer type, ethnicity, and sample size, considering that rs3757441 plays a protective role in lung cancer in a Korean populationCitation21 but acts as a risk factor in CRC in a Han Chinese population.Citation24 Furthermore, we searched for articles related to EZH2 rs41277434, and our results indicated that no significant association was found between rs41277343 and overall cancer risk or DSC risk.
Sensitivity analysis revealed that the results of our study were robust. Egger’s and Begg’s tests indicated a publication bias in homozygote and recessive models of rs3757441 and rs41277434. Future large-scale well-designed studies should be conducted to confirm the publication bias of the genetic models of rs375441 and rs41277434.
Several limitations of our meta-analysis should be considered. First, most of the eligible studies mainly focused on East Asian populations, whereas 2 studies involved Caucasians. Studies on other ethnicities were not included in this meta-analysis. Thus, our results were incomplete. The number of eligible studies and the sample size were relatively small and might consequently cause a type II error. Second, our results were based on unadjusted estimates because of the lack of original data on age, gender, and smoking status. Potential bias caused by these factors might also persist. Third, differences among various cancers might lead to heterogeneity when all cancer types were pooled. Stratified analysis by specific cancer type was not conducted because of the insufficient number of studies on single cancer type. Finally, we only searched for publications in Chinese and English. As such, language restriction would limit our sample size.
Conclusion
Despite the limitations, our meta-analysis revealed that EZH2 rs887569 and rs2302427 might be correlated with a decreased cancer risk in specific genetic models, whereas the association of EZH2 rs3757441 and rs41277434 polymorphisms with overall cancer risk was not observed. To confirm our results and provide highly reliable evidence supporting these associations, we recommend future large-scale and well-designed studies on diverse ethnic populations and cancer types.
Author contributions
ZL performed the experiments and wrote the paper. ZY performed the experiments, prepared figures, and/or tables. LH analyzed the data, prepared figures, and/or tables. LZ ana-lyzed the data. YW reviewed drafts of the paper. MZ analyzed the data, contributed reagents/materials/analysis tools. GZ contributed reagents/materials/analysis tools and reviewed drafts of the paper. SC contributed reagents/materials/analysis tools. BX and MC conceived and designed the experiments, and reviewed drafts of the paper. All authors contributed toward data analysis, drafting and revising the paper and agree to be accountable for all aspects of the work.
Acknowledgments
This work was supported by grants from National Natural Science Foundation of China (NO 81672551, 81572517, 81370849, 81300472), Natural Science Foundation of Jiangsu Province (BK20161434, BL2013032, BK20150642, and BK2012336), Six Talent Peaks Project in Jiangsu Province, Jiangsu Provincial Medical Innovation Team (CXTDA2017025), Jiangsu Provincial Medical Talent (ZDRCA2016080), Jiangsu Provincial Medical Youth Talent (QNRC2016821, QRNC2016820), Graduate Research Innovation Program (KYCX17_0180).
Supplementary materials
Figure S1 Forest plot for the relationship between rs3757441 and cancer risk: (A) CT/TT; (B) CC/TT; (C) CCCT/TT; (D) CC/CTTT.

Figure S2 Forest plot for the relationship between rs41277434 and cancer risk: (A) AC/AA; (B) CC/AA; (C) ACCC/AA; (D) CC/AAAC.
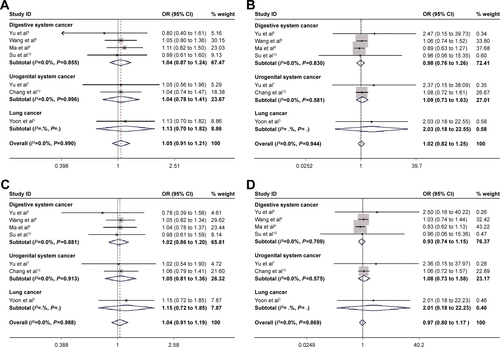
Figure S3 Forest plot of sensitivity analysis for EZH2 SNPs.

Table S1 Characteristics of eligible studies for each SNP in the meta-analysis
References
- BachmannNHoegelJHaeuslerJMutation screen and association study of EZH2 as a susceptibility gene for aggressive prostate cancerProstate200565325225916015586
- BreyerJPMcReynoldsKMYaspanBLBradleyKMDupontWDSmithJRGenetic variants and prostate cancer risk: candidate replication and exploration of viral restriction genesCancer Epidemiol Biomarkers Prev20091872137214419567509
- YoonKAGilHJHanJParkJLeeJSGenetic polymorphisms in the polycomb group gene EZH2 and the risk of lung cancerJ Thorac Oncol201051101619901851
- ZhouYDuWDWuQEZH2 genetic variants affect risk of gastric cancer in the Chinese Han populationMol Carcinog201453858959722228224
- YuYLSuKJHsiehYHEffects of EZH2 polymorphisms on susceptibility to and pathological development of hepatocellular carcinomaPLoS One201389e7487024040354
- WangJMaZBLiKGuoGHAssociation between EZH2 polymorphisms and colorectal cancer risk in Han Chinese populationMed Oncol201431387424488618
- YuYLSuKJHsiehMJImpact of EZH2 polymorphisms on urothelial cell carcinoma susceptibility and clinicopathologic featuresPLoS One201494e9363524691023
- MaZBGuoGHNiuQShiNRole of EZH2 polymorphisms in esophageal squamous cell carcinoma risk in Han Chinese populationInt J Mol Sci2014157126881269725036033
- HuangYWangXZhangLCorrelations of single nucleotide polymorphisms of EZH2 gene with genetic susceptibility of colorectal cancerChin J Clin Lab Sci2015334262265
- SuKJLinCWChenMKYangSFYuYLEffects of EZH2 promoter polymorphisms and methylation status on oral squamous cell carcinoma susceptibility and pathologyAm J Cancer Res20155113475348426807327
- TaoRChenZWuPThe possible role of EZH2 and DNMT1 polymorphisms in sporadic triple-negative breast carcinoma in southern Chinese femalesTumour Biol201536129849985526162541
- ChangWSLiaoCHTsaiCWAssociation of Enhancer of Zeste 2 (EZH2) Genotypes with Bladder Cancer Risk in TaiwanAnticancer Res20163694509451427630289
Disclosure
The authors report no conflicts of interest in this work.
References
- SiegelRLMillerKDJemalACancer statistics, 2017CA Cancer J Clin201767173028055103
- Pedraza-FarinaLGMechanisms of oncogenic cooperation in cancer initiation and metastasisYale J Biol Med2006793–49510317940619
- VolkelPDupretBLe BourhisXAngrandPODiverse involvement of EZH2 in cancer epigeneticsAm J Transl Res20157217519325901190
- Di CroceLHelinKTranscriptional regulation by Polycomb group proteinsNat Struct Mol Biol201320101147115524096405
- YangYZhouLLuLA novel miR-193a-5p-YY1-APC regulatory axis in human endometrioid endometrial adenocarcinomaOncogene201332293432344222907428
- BoulayGDubuissezMVan RechemCHypermethylated in cancer 1 (HIC1) recruits polycomb repressive complex 2 (PRC2) to a subset of its target genes through interaction with human polycomb-like (hPCL) proteinsJ Biol Chem201228713105091052422315224
- SmitsMNilssonJMirSEmiR-101 is down-regulated in glioblastoma resulting in EZH2-induced proliferation, migration, and angiogenesisOncotarget20101871072021321380
- VaramballySDhanasekaranSMZhouMThe polycomb group protein EZH2 is involved in progression of prostate cancerNature2002419690762462912374981
- BrackenAPPasiniDCapraMProsperiniEColliEHelinKEZH2 is downstream of the pRB-E2F pathway, essential for proliferation and amplified in cancerEMBO J200322205323533514532106
- BachmannIMHalvorsenOJCollettKEZH2 expression is associated with high proliferation rate and aggressive tumor subgroups in cutaneous melanoma and cancers of the endometrium, prostate, and breastJ Clin Oncol200624226827316330673
- SauvageauMSauvageauGPolycomb group proteins: multi-faceted regulators of somatic stem cells and cancerCell Stem Cell20107329931320804967
- YapDBChuJBergTSomatic mutations at EZH2 Y641 act dominantly through a mechanism of selectively altered PRC2 catalytic activity, to increase H3K27 trimethylationBlood201111782451245921190999
- SimonCChagraouiJKroslJA key role for EZH2 and associated genes in mouse and human adult T-cell acute leukemiaGenes Dev201226765165622431509
- ZhangJDingLHolmfeldtLThe genetic basis of early T-cell precursor acute lymphoblastic leukaemiaNature2012481738015716322237106
- StangACritical evaluation of the Newcastle-Ottawa scale for the assessment of the quality of nonrandomized studies in meta-analysesEur J Epidemiol201025960360520652370
- HigginsJPThompsonSGDeeksJJAltmanDGMeasuring inconsistency in meta-analysesBMJ2003327741455756012958120
- DerSimonianRLairdNMeta-analysis in clinical trialsControl Clin Trials1986731771883802833
- MantelNHaenszelWStatistical aspects of the analysis of data from retrospective studies of diseaseJ Natl Cancer Inst195922471974813655060
- BachmannNHoegelJHaeuslerJMutation screen and association study of EZH2 as a susceptibility gene for aggressive prostate cancerProstate200565325225916015586
- BreyerJPMcReynoldsKMYaspanBLBradleyKMDupontWDSmithJRGenetic variants and prostate cancer risk: candidate replication and exploration of viral restriction genesCancer Epidemiol Biomarkers Prev20091872137214419567509
- YoonKAGilHJHanJParkJLeeJSGenetic polymorphisms in the polycomb group gene EZH2 and the risk of lung cancerJ Thorac Oncol201051101619901851
- ZhouYDuWDWuQEZH2 genetic variants affect risk of gastric cancer in the Chinese Han populationMol Carcinog201453858959722228224
- YuYLSuKJHsiehYHEffects of EZH2 polymorphisms on susceptibility to and pathological development of hepatocellular carcinomaPLoS One201389e7487024040354
- WangJMaZBLiKGuoGHAssociation between EZH2 polymorphisms and colorectal cancer risk in Han Chinese populationMed Oncol201431387424488618
- YuYLSuKJHsiehMJImpact of EZH2 polymorphisms on urothelial cell carcinoma susceptibility and clinicopathologic featuresPLoS One201494e9363524691023
- MaZBGuoGHNiuQShiNRole of EZH2 polymorphisms in esophageal squamous cell carcinoma risk in Han Chinese populationInt J Mol Sci2014157126881269725036033
- HuangYWangXZhangLCorrelations of single nucleotide polymorphisms of EZH2 gene with genetic susceptibility of colorectal cancerChin J Clin Lab Sci2015334262265
- SuKJLinCWChenMKYangSFYuYLEffects of EZH2 promoter polymorphisms and methylation status on oral squamous cell carcinoma susceptibility and pathologyAm J Cancer Res20155113475348426807327
- TaoRChenZWuPThe possible role of EZH2 and DNMT1 polymorphisms in sporadic triple-negative breast carcinoma in southern Chinese femalesTumour Biol201536129849985526162541
- ChangWSLiaoCHTsaiCWAssociation of Enhancer of Zeste 2 (EZH2) Genotypes with Bladder Cancer Risk in TaiwanAnticancer Res20163694509451427630289
- ArisanSBuyuktuncerEDPalavan-UnsalNCaskurluTCakirOOErgenekonEIncreased expression of EZH2, a polycomb group protein, in bladder carcinomaUrol Int200575325225716215315
- MatsukawaYSembaSKatoHItoAYanagiharaKYokozakiHExpression of the enhancer of zeste homolog 2 is correlated with poor prognosis in human gastric cancerCancer Sci200697648449116734726
- WatanabeHSoejimaKYasudaHDeregulation of histone lysine methyltransferases contributes to oncogenic transformation of human bronchoepithelial cellsCancer Cell Int200881518980680
- KleerCGCaoQVaramballySEZH2 is a marker of aggressive breast cancer and promotes neoplastic transformation of breast epithelial cellsProc Natl Acad Sci U S A200310020116061161114500907
- TsangDPChengASEpigenetic regulation of signaling pathways in cancer: role of the histone methyltransferase EZH2J Gastroenterol Hepatol20112611927
- KimKHRobertsCWTargeting EZH2 in cancerNat Med201622212813426845405
- MorinRDJohnsonNASeversonTMSomatic mutations altering EZH2 (Tyr641) in follicular and diffuse large B-cell lymphomas of germinal-center originNat Genet201042218118520081860
- MajerCRJinLScottMPA687V EZH2 is a gain-of-function mutation found in lymphoma patientsFEBS Lett2012586193448345122850114
- McCabeMTGravesAPGanjiGMutation of A677 in histone methyltransferase EZH2 in human B-cell lymphoma promotes hypertrimethylation of histone H3 on lysine 27 (H3K27)Proc Natl Acad Sci U S A201210982989299422323599
- PaolicchiEPacettiPGiovannettiEA single nucleotide polymorphism in EZH2 predicts overall survival rate in patients with cholangiocarcinomaOncol Lett2013651487149124179546