
Abstract
Background
Delta-tocotrienol (δT), an isomer of vitamin E, exhibits anticancer properties in different cancer types including non-small-cell lung cancer (NSCLC). Yet, anti-invasive effects of δT and its underlying cellular mechanism in NSCLC have not been fully explored. Matrix metalloproteinase 9 (MMP-9)-based cell migration and invasion are critical cellular mechanisms in cancer development. The current evidence indicates that MMP-9 is upregulated in most patients, and the inhibition of MMPs is involved in decreasing invasion and metastasis in NSCLC. Therefore, its suppression is a promising strategy for attenuating cell invasion and metastasis processes in NSCLC.
Purpose
The aim of this study was to evaluate the possibility of MMP-9 inhibition as the underlying mechanism behind the antimetastatic properties of δT on NSCLC cells.
Methods
The effects of δT on cell proliferation, migration, invasion, adhesion, and aggregation capabilities were investigated using different cell-based assays. An inhibitory effect of MMP-9 enzyme activity with δT was also identified using gel zymography. Using real-time PCR and Western blot analysis, a number of cellular proteins, regulatory genes, and miRNA involved in the Notch-1 and urokinase-type plasminogen activator (uPA)-mediated MMP-9 pathways were examined.
Results
The study found that δT inhibited cell proliferation, cell migration, invasion, aggregation, and adhesion in a concentration-dependent manner and reduced MMP-9 activities. Real-time PCR and Western blot analysis data revealed that δT increased miR-451 expressions and downregulated Notch-1-mediated nuclear factor-κB (NF-κB), which led to the repressed expression of MMP-9 and uPA proteins.
Conclusion
δT attenuated tumor invasion and metastasis by the repression of MMP-9/uPA via downregulation of Notch-1 and NF-κB pathways and upregulation of miR-451. The data suggest that δT may have potential therapeutic benefit against NSCLC metastasis.
Introduction
Lung cancer is the leading cause of estimated cancer deaths in the USA.Citation1 Non-small-cell lung cancer (NSCLC) accounts for 85% of all lung cancer cases and can be classified into three subtypes: squamous cell carcinoma, large cell carcinoma, and adenocarcinoma. The initial stage of NSCLC has a 5-year survival rate of 55%, but this rate reduces to <4% for cases diagnosed with distant metastasis.Citation1 With current advances in the understanding of mechanisms of cancer invasion and metastasis, it is becoming clear that matrix metalloproteinases (MMPs), an enzyme with 21 subtypes in humans,Citation2,Citation3 have a strong association with local invasion or distant metastasis.Citation2 Several studies ranging from cell cultureCitation4 to clinical investigationsCitation5–Citation7 have reported the inhibition of MMPs in conditions of decreasing invasion and metastasis in NSCLC.
Matrix metalloproteinase 9 (MMP-9), a subtype of MMPs, regulates cell migration, angiogenesis, adhesion, aggregation, and immune response in cancer.Citation8–Citation10 In this process, MMP-9 is mainly responsible for degrading collagen type IV and elastin in basal membranes, facilitating lung cancer metastasis. High levels of MMP-9 have also been reported in the serum of lung carcinoma patients.Citation11 Therefore, the modulation of MMP-9 protein expressions and their activities would be excellent therapeutic targets for the inhibition of invasion and metastasis processes in NSCLC. Urokinase-type plasminogen activator (uPA), a serine proteinase, binds to the urokinase-type plasminogen activator receptor (uPAR) and transforms inactive plasmin and other proteases, including MMP-9, into their active forms. Regulating uPA is one of the major approaches that can directly modulate MMP-9 activities in cancer.Citation12 The uPA pathway includes several proteins such as serine protease, uPAR, and the endogenous inhibitors, plasminogen activator inhibitors 1 and 2.Citation13 The uPA system enables transformation of zymogen plasminogen into plasmin in the process of extracellular matrix (ECM) degradation.Citation14 The plasmin, then, facilitates the conversion of inactive pro-MMP-9 into active MMP-9. Increased expression of the uPA system has been reported in NSCLC tissue as compared to normal lung tissue.Citation15 Using antisense technology, Rao et alCitation16 showed that the inhibition of uPA and MMP-9 might be an excellent anti-invasion and antimetastatic approach for cancer gene therapy in lung cancer. Although the inhibition of uPA and/or MMP-9 is a possible therapeutic target for preventing local invasion or distant metastases in lung cancer, uPA and MMP-9 pathways have shown cross talks with external factors, namely transcription factors (TFs) and miRNA. These cross talks have made it more complex to modulate MMP-9 directly.
Tong et alCitation17 showed that nuclear factor-κB (NF-κB), a TF involved in cancer initiation and progression, directly binds with the uPA promoter in vitro. The same study showed that the inhibition of NF-κB activities decreased cell invasion and uPA synthesis in NSCLC cells. The MMP-9 promoter has binding sites for NF-κB.Citation18 Inability of NF-κB to bind with the MMP-9 promoter has been shown to decrease MMP-9 synthesis.Citation18–Citation21 Moreover, it has been suggested that the NF-κB signaling pathway contributes to the progression of metastasis by regulating MMP-9 in colorectal cancer,Citation22 prostate cancer,Citation23 renal cancer,Citation24 ovarian cancer,Citation25 and head and neck cancer.Citation26
In addition, elevated miR-451, a small noncoding RNA that controls gene expression through sequence-specific binding to target mRNA, was found to decrease cell invasion and metastasis, with corresponding decrease in MMP-9 expression levels in primary liver cancer.Citation27 Elevated miR-451 expressions were also found to suppress cell proliferation and metastasis in A549 lung cancer cell lines.Citation28 However, the role of NF-κB and/or uPA in mediating MMP-9 function and their interaction with miR-451 in cell invasion and metastasis in lung cancer remains generally unclear.
Previous studies from our laboratory showed that delta-tocotrienol (δT), an isomer of vitamin E found naturally in food sources, inhibits NF-κB signaling pathways via the downregulation of Notch-1, thereby decreasing the proliferation and metastatic/invasion potential, while inducing apoptosis of NSCLC cells in a concentration- and time-dependent manner.Citation28–Citation33 This study investigated the antimetastatic effect of δT on NSCLC cells with the hypothesis that the MMP-9-dependent invasion and metastasis of NSCLC cells are inhibited by the suppression of Notch-1-mediated NF-κB and uPA pathways and induction of miR-451.
Materials and methods
Reagents and antibodies
American River Nutrition (Delta Gold, MA, USA) donated δT for this experiment. Dimethyl sulfoxide (DMSO), tumor necrosis factor-alpha (TNF-α), bovine serum albumin (BSA), and bicinchoninic acid (BCA) assay were purchased from Thermo Scientific Pierce (St Louis, MO, USA). Two NSCLC cell lines, A549 and H1299, were purchased from American Type Culture Collection (Manassas, VA, USA). Roswell Park Memorial Institute (RPMI) medium (Mediatech, Manassas, VA, USA) was used as a growth medium for cells. Antibodies β-actin, MMP-9, Hairy and Enhancer of Split-1 (HES-1), Notch-1 (Cell Signaling Technology, Danvers, MA, USA), and uPA (Santa Cruz Biotechnology Inc., Dallas, TX, USA) were used for Western blot analysis.
Cell culture and treatments
A549 and H1299 were cultured in RPMI medium and supplemented with 10% fetal bovine serum (FBS) and 1% penicillin and streptomycin in 5% CO2 at 37°C. The cell growth medium was changed every 2–3 days. Adherent cells were detached using trypsin–EDTA and centrifuged at 1,500 rpm. The treatment media were prepared by mixing δT (10, 20, or 30 μM <0.01% DMSO as a vector) in the RPMI medium, whereas the control was grown in RPMI media containing <0.01% DMSO.
Cell viability assay
A 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) assay was conducted to analyze the antiproliferative effects of δT on the above NSCLC cell lines; 5×104 of A549 and H1299 cells were grown in 96-well plates overnight. Afterward, the media were changed, and the cells were treated with fresh cell growth medium containing <0.01% DMSO (control) or treatment medium containing 10, 20, 30, and 40 μM of δT. After 72 hours of treatment, 20 μL of CellTiter 96® Aqueous Reagent from Promega (Madison, WI, USA) was mixed into each well and incubated for 2 hours. The absorbance at 490 nm was then measured using the Bio-Tek ELx800™ plate reader (Winooski, VT, USA). Each variant of the experiment was performed with six replicates.
Cell invasion assay
The ability of A549 and H1299 lung cancer cells to pass through filters coated with Matrigel® was assessed by a BD Biocoat™ invasion kit (BD Biosciences, San Jose, CA, USA); 2.5×105 of both cell lines were prepared in an aqueous environment with PBS and resuspended into each upper chamber of 6-well plates with a serum-free medium in the presence or absence of treatment; 3 mL of growth medium with 15% FBS was added to lower chambers and incubated for 20 hours. After incubation, the attached cells on the upper side of the Matrigel surface (in the upper chamber) were wiped with a cotton swab, and 50 μL of MTS reagent was added to the lower chamber. The absorbance of the media contained in the lower chamber was measured at 450 nm using the Bio-Tek ELx800 plate reader. Each experiment was carried out in triplicate.
Again, the same procedure was continued up to 20 hours of incubation in a BD Biocoat invasion kit. The cells attached to the Matrigel upper surface were then wiped using a cotton swab, and the cells invading through the Matrigel to the lower surface were fixed with 4% paraformaldehyde and stained with 2% crystal violet. The invading cells on the Matrigel lower surface were counted and photographed with the camera attached to the light microscope. The data were shown in pictures as stained cells attached to the bottom of Matrigel surface from randomly selected areas.
Cell migration assay
To detect the migration rate of cells using a wound healing assay, A549 and H1299 cells were grown in a 6-well dish at a concentration of 1×106 cells per well. After the cells had been incubated for 30 hours, the growth media were replaced with or without treatment media. Scratches were made using a pipette tip (100 μL) and photographed immediately (time 0). After a 20-hour incubation with treatment media at 0, 10, 20, and 30 μM, all the wound areas were washed with PBS three times to ensure that no loosely held cells were attached. The plate was imaged, and the width of the scratch was measured by using a Nikon H 600L microscope (Nikon Corporation, Tokyo, Japan) at 10× objective.
Cell aggregation assay
To investigate the antiaggregation capabilities of δT, a single-cell suspension was created using cell trypsinization technique; 2×105 cells in 1 mL of RPMI growth medium with or without δT (10, 20, and 30 μM) concentration were placed in polystyrene microtubes and were gently shaken every 5 minutes for 1 hour at 37°C. After 1 hour, glutaraldehyde was added at a final concentration of 2% (v/v) to halt the cell aggregation. Homotypic aggregation was assessed and photographed by a LUNA™ automated cell counter (Logos Biosystems, Gyeonggi-do, South Korea). The aggregates were counted per cm2, and significant differences between control and treatments were evaluated using ezANOVA software.
Cell adhesion assay
The cell lines A549 and H1299 (1,000,000) were pretreated with or without δT concentrations (control, 10, 20, and 30 μM) for 72 hours. The cell lines were suspended in cell growth medium to form a single-cell suspension, and 2×106 cells/mL (2×105 cells/well) were seeded into a 96-well cell culture plate precoated with Matrigel. After 1 hour of incubation, the wells were washed three times with PBS to eliminate nonadherent cells, and 100 mL of RPMI medium with 20 μM MTS reagent was added into each well for an additional 2 hours. As a measure of a number of cells, the absorbance at 570 nm was taken on an ELx800 plate reader. Significant differences between control and treatments were evaluated using ezANOVA software.
Zymogram gel assay
To investigate the activity of MMP-9, A549 and H1299 cells were plated in a six-well plate with or without treatment by δT in FBS-free RPMI media. The media of all groups were removed and concentrated into 10× media by using 3-KDa Amicon® centrifugal filters (MilliporeSigma, Burlington, MA, USA). Next, the total protein concentrations of the concentrated media were estimated by a Pierce BCA protein assay kit (Bio-Rad Laboratories, Hercules, CA, USA) and normalized to 2,000 mg in the sample. Zymogram precast gel (Bio-Rad Laboratories) with gelatin coating was used as a separation gel. One part of sample was mixed with two parts of zymogram sample buffer. Samples were loaded in the gel, and the gel was run with 1× Tris–glycine–sodium dodecyl sulfate (SDS) buffer at 200 V. When the tracking dye (bromophenol blue) touched the end of the gel, the gel was removed from the apparatus. Then, the gel was incubated in the renaturing buffer (100 mL for one or two mini-gels) with gentle shaking for 30 minutes at 25°C. Afterward, the zymogram renaturing buffer was exchanged with 1× developing buffer, and the gel was equilibrated for 30 minutes at room temperature with gentle agitation, followed by incubation with fresh 1× zymogram developing buffer at 37°C overnight for maximum sensitivity. The gel was stained with Coomassie blue R-250 for 30 minutes to 1 hour the next day. Stain concentration of 0.5% (w/v in 40% methanol and 10% acetic acid) was used, and the gels were destained with Coomassie R-250 destaining solution (methanol/acetic acid/water, 40:10:50). Where the MMP-9 had digested the substrate, zones of MMP-9 activity appeared as clear bands against a dark blue background.
Western blot analysis
To investigate the protein expression in signaling pathways of A549 and H1299 cells with δT exposure, 1,000,000 cells were plated per 100-mm dish and incubated for 24 hours. The cells were then grown for 72 hours with treatment and control medium. Next, the cells were lysed in the cold 1× cell lysis buffer (Cell Signaling Technology) for 30 minutes on ice with 1× protease inhibitor (Cell Signaling Technology), and protein concentrations were calculated using a Pierce BCA protein assay kit. Subsequently, 50 mg of total cell lysates was mixed with same amounts of 4× Laemmli buffer (Bio-Rad Laboratories), and the samples were loaded onto a 10% SDS–polyacrylamide gel. The gel was electrophoretically transferred to a polyvinylidene difluoride (PVDF) transfer membrane (Whatman, Clifton, NJ, USA) using a transfer buffer (25 mM Tris, 190 mM glycine, and 20% methanol) after gel electrophoresis. The PVDF membranes were incubated for 2 hours at room temperature with 5% BSA in 1× Tris-buffered saline buffer containing 0.1% Tween (TBS-T) and then incubated overnight at 4°C with primary antibodies (1:1,000). The membranes were washed three times with TBS-T and incubated with the secondary antibodies (1:5,000) containing 2% BSA at room temperature for 2 hours. The signal strength was then measured in an image with Chemi-Doc™ XRS system (Bio-Rad Laboratories).
Quantitative real-time PCR (qRT-PCR)
For this process, 1,000,000 A549 and H520 cells were seeded per 100-mm dish per plate and incubated for 24 hours. Subsequently, the culturing media were replaced with treatment or control medium and incubated for another 72 hours. The RNA was then extracted using RNeasy® Mini Kit from Qiagen (Valencia, CA, USA); 1,000 ng of total RNA from each sample was subjected to first-strand cDNA synthesis using high-capacity RNA to cDNA master mix (Applied Biosystems, Foster City, CA, USA) in a total volume of 50 mL. qRT-PCR was performed as part of gene expression analysis. The appropriate primers – sense of MMP-9, 5′-CAACATCACCTATTGGATCC-3′, antisense of MMP-9, 5′-CGGGTGTAGAGTCTCTCGCT-3′, sense of β-actin, 5′-TGGTCCTCTGGGCATCTCAGGC-3′, antisense of 5′-GGTGAACCTGCTGTTGCCCTCA-3′ – were used for PCR amplification. Diluted cDNA (2 μL) and 2 μL of each reverse and forward primer and 12.5 μL master mix (SYBR GREEN PCR Master Mix; Applied Biosystems, Warrington, UK) were used in each 25 μL PCR performed in Eppendorf MasterCycler realplex 4 (Eppendorf, Hauppauge, NY, USA) at 25°C for 10 minutes, followed by 48°C for 30 minutes, and 95°C for 5 minutes. Expression values were normalized with β-actin. Each gene expression was tested in triplicate.
miRNA-451 expression analysis
To examine the miRNA-451 expression, the total RNA was extracted from the cultured cells using RNeasy Mini Kit from Qiagen according to the manufacturer’s protocols. The concentration and purity of the RNA were determined using the NanoDrop 2000 system (Thermo Fisher Scientific, Tokyo, Japan). SYBR GREEN PCR Master Mix (Applied Biosystems, Warrington, UK) was used for the qRT-PCR according to the manufacturer’s instructions. U-6 primers from miScript miRNA PCR (Qiagen) were used as an endogenous reference, and its levels were used for normalization.
NF-κB-binding activity
To determine the NF-κB TF-binding activity, A549 and H1299 cells were seeded in Petri dishes and incubated for 24 hours. The cells were then treated with or without δT. After 72 hours of treatment, they were collected and washed with the PBS. The nuclear protein extraction was performed with an NE-PER Nuclear and Cytoplasmic Extraction Reagent kit (Pierce Biotechnology, Rockford, IL, USA) based on the manufacturer’s protocols. Nuclear protein concentrations were determined using the Pierce BCA protein assay kit (Pierce Biotechnology), and an NF-κB filter plate assay kit (Signosis, Sunnyvale, CA, USA) was used to determine the NF-κB DNA-binding ability. The assay was performed according to the manufacturer’s protocol. The biotin-labeled DNA sequence of NF-κB was mixed with 3 μg of nuclear extract to make an NF-κB–DNA-binding complex. Then, 10 μL TF-binding buffer mix, 2 μL NF-κB probe, and 3 μg of distilled water were added to each sample, bringing the total volume to 20 μL. The filter plate was used to retain the bound NF-κB probe, while the unbound NF-κB probe was filtered out. The bound, prelabelled NF-κB probe was then eluted from the filter, collected, and transferred to a hybridization plate for quantitative analysis. The NF-κB probe was further detected using streptavidin–horseradish peroxidase, and the luminescence of the probe was measured using Bio-Tek Synergy Microplate Reader.
Statistical analysis
The data were expressed as the mean ± SD of the mean. Differences were evaluated by using the one-way analysis of variance followed by Tukey honestly significant difference multiple comparison tests using the ezANOVA software. P<0.05 was considered to specify a statistically significant change.
Results
Antiproliferative effect of δT
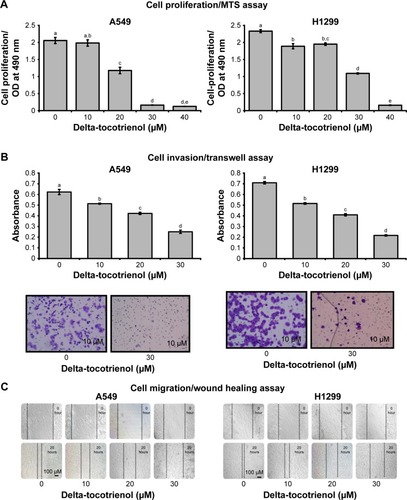
The MTS assay showed a concentration-dependent reduction in cell proliferation in both A549 and H1299 cells with the addition of δT. A reduction of ~5%, 40%, 91%, and 92% of cell growth was observed after 72 hours of incubations with 10, 20, 30, and 40 μM of δT, respectively, compared with control A549 cells. Similarly, the H1299 cell line showed ~20%, 16%, 53%, and 93% cell growth inhibition, respectively, under the same conditions and same doses of δT and ().
Figure 1 δT inhibits cell proliferation, invasion, and cell migration in NSCLC cells.
Abbreviations: ANOVA, analysis of variance; δT, delta-tocotrienol; HSD, honestly significant difference; MTS, (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium); NSCLC, non-small-cell lung cancer.
Inhibition of cell invasion and migration by δT
We observed dose-dependent anti-invasive effects of δT in both A549 and H1299 cells at concentration levels ranging from 0 to 30 μM in the Matrigel invasion assay (). At 10, 20, and 30 μM concentrations of δT, cell invasion was reduced by ~10%, 35%, and 65% in A549 cell lines, respectively, after 72-hour incubation. Similarly, H1299 cells showed ~35%, 45%, and 70% inhibition of cell invasion under the same conditions. For further confirmation, microscopic images of the migrated cells through the membrane were taken, stained, and analyzed. In A549, as compared to the untreated cells, a small number of treated cells were observed to migrate through the membrane. Hardly any invaded cells could be observed upon the treatment of H1299 cells at 30 μM of δT (P<0.05 at 30 μM treatment; ). The wound healing assay was completed to explore the migration of A549 and H1299 cells after δT treatment. As shown in , δT significantly inhibited the migration of cells after 20 hours of treatment with 30 μM δT.
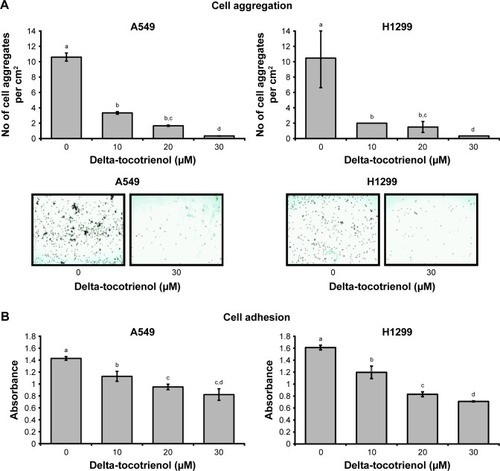
Inhibition of cell aggregation and adhesion capabilities by δT
As shown in , δT inhibited cell aggregation at 30 μM of δT in both A549 and H1299 cells. To test the effects of δT on cell adhesion to the Matrigel surface, A549 and H1299 cells were exposed with δT for 72 hours, and adhesion capacities to Matrigel surface were measured. We observed a dose-dependent inhibition of cell adhesion in both A549 and H1299 cells ().
Figure 2 δT inhibits cell aggregation and adhesion in A549 and H1299 cells.
Abbreviations: ANOVA, analysis of variance; δT, delta-tocotrienol; HSD, honestly significant difference; SEM, standard error of the mean.
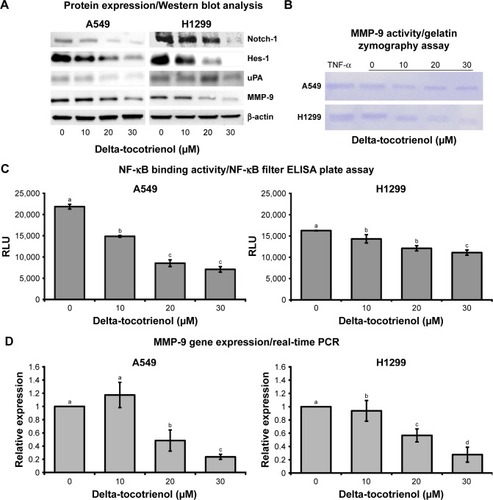
δT suppressed expression of MMP-9
Since metalloproteinase plays a central function in cell invasion, we then performed the zymography assay to compare the action of MMPs in treated and control cells. A dose-dependent decrease in the MMP-9 activity resulted in the zymography assay with δT () at concentrations of 0, 10, 20, and 30 μM. TNF-α-treated cells served as a positive control. The results from the Western blot analysis indicate that δT inhibits MMP-9 protein expressions in a concentration- and time-dependent manner compared with the control in both A549 and H1299 cells (). In order to verify whether the suppression of MMP-9 protein expression resulted from decreasing m-RNA expression, real-time PCR analysis was performed. PCR is a gold standard for gene expression analysis, and it has been widely used for gene expression analysis in a variety of study models including cells, microboes,Citation34–Citation36 and animals.Citation37 We observed that the mRNA level of MMP-9 was reduced significantly in a dose-dependent manner, compared with the control group (), in both A549 and H1299 cells after the treatment with δT for 72 hours. The δT-mediated change in the mRNA levels of MMP-9 was correlated with the protein level expression of MMP-9, as indicated by the results from the Western blot analysis, suggesting that δT may regulate MMP-9 expression at transcription levels from TFs, upstream signaling pathways, or other factors such as miRNA.
Figure 3 δT inhibits MMP-9 activity and expression via the inhibition of Notch-1, NF-κB, and uPA.
Abbreviations: ANOVA, analysis of variance; δT, delta-tocotrienol; HSD, honestly significant difference; MMP-9, matrix metallopeptidase 9; qRT-PCR, quantitative real-time PCR; TNF-α, tumor necrosis factor-alpha; NF-κB, nuclear factor-κB; uPA, urokinase-type plasminogen activator.
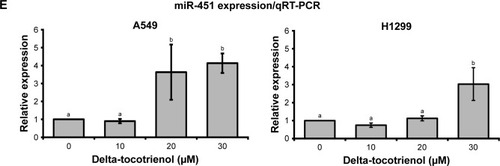
δT increases expression of miR-451
Accumulating evidence showed that miR-451 plays a key role in different aspects of carcinogenesis in lung cancer including tumor growth, invasion, radioresistance, and chemoresistance.Citation38–Citation41 A recent case analysis with 370 NSCLC incidents revealed that miR-451 expression was lower in carcinoma tissues than in paired background lung tissues.Citation42 The same study also revealed that the miR-451 expression was inversely correlated with macrophage migration inhibitory factor expression, a factor that induces the expression of MMP-9.Citation42 In another study, A549 cells transfected with miR-451 showed reduced cell invasion and metastasis compared with the control by MTT transwell invasion assay and wound healing assay, respectively. Cancer metastasis factors (MMP-9, VEGF, and CXCR4) showed corresponding reduction in protein expression by Western blot analysis.Citation43 This evidence suggests the influence of miR-451 on MMP-9 expression. Therefore, we evaluated the differences in miR-451 expression upon δT treatments in A549 and H1299 cells. Indeed, we found that miR-451 expression increased by 2- and 3.5-fold upon treatment with δT in A549 and H1299 cells, respectively, as compared to control cells (). These results suggest that δT possibly inhibits the cell invasion of NSCLS via increased expression of miR-451, which may further mediate the expression of MMP-9 in A549 and H1299 cells.
δT inhibits expression uPA/Notch-1 pathway proteins
In the cell metastasis process, MMP-9 and/or their activators (uPA) play a major role in making the interaction between cells and the ECM.Citation44 Elevated expression levels of the uPA and MMP-9 were reported in NSCLC tissue as compared to normal lung tissue,Citation15 indicating that they may be facilitators for the metastasis process. Infection with Ad-uPAR-MMP-9 caused a decrease in the expression of uPA and MMP-9 by >90% with a concomitant reduction in cell invasion by H1299 cells.Citation16 In breast cancer cells, silencing of uPA and MMP-9 reduced their wound healing, migratory, invasive, and adhesive capacity.Citation45 We observed similar effects upon treatment of NSCLC cells with δT. This evidence strongly suggests the possible relationship between MMP-9 and uPA expressions in cancer cells. Therefore, we explored the protein expressions of uPA and MMP-9 using Western blot analysis. Our data showed a dose-dependent inhibition of both MMP-9 and uPA expressions in A549 and H1299 cells upon δT treatment, suggesting that δT inhibits cell invasion and metastasis via the inhibition of uPA and MMP-9 in A549 and H1299 cells ().
Aberrant Notch-1 expressions have been reported in a variety of human malignancies. It appears to regulate many basic processes essential for cancer development and progression, including cell growth, survival, apoptosis, migration, and invasion along with implicated MMP-9 expressions. Inhibiting Notch signaling using gamma-secretase inhibitor resulted in reduced expression of MMP-9 induced by DLK1 in H1299 and A549 cell lines.Citation46 Moreover, silencing of Notch-1 protein led to downregulation and inactivation of VEGF and MMP-9, thereby inhibiting invasion and metastasis of pancreatic cancer cells.Citation47 Since these studies indicate a direct relationship between Notch-1 and MMP-9 proteins, we tested Notch-1 expression in whole-cell lysates by Western blotting to determine whether δT downregulates MMP-9 expression in a Notch-signaling-dependent manner in NSCLC. Our results showed that δT did downregulate Notch-1 expression in both A549 and H1299 cells () along with the expression of MMP-9. Furthermore, the expression of HES-1, a Notch-1 downstream target gene,Citation48 was also examined by Western blot in A549 and H1299 cells to validate downstream involvement of Notch-1 signaling in the regulation of MMP-9 expression. Our results showed that δT downregulated HES-1 expression along with Notch-1 expression in both A549 and H1299 cells (). The simultaneous inhibition of Notch-1/HES-1 and MMP-9 with δT suggests a mechanistic interplay between MMP-9 and uPA/Notch-1 in metastasis.
Inhibition of NF-κB DNA-binding activity with δT
MMP-9 promoter is one of the most intensively studied promotersCitation49 containing several TF-binding elements, including binding sites for NF-κB.Citation50 Since NF-κB plays important roles in the regulation of MMP-9 expression, we evaluated whether δT affected the DNA-binding activity of the nuclear translocated NF-κB in these cells. Our data showed a dose-dependent inhibition of NF-κB-binding activities in A549 and H1299 upon δT treatment, suggesting that the inhibition of MMP-9 expression lowers DNA-binding activity of translocated NF-κB ().
Discussion
The metastasis process is a multistep process, involving the migration/invasion of cells through the ECM/basement membrane into the circulatory system, survival without contact with the ECM, adhesion to the wall of lymphatic or blood vessels, and detachment from the vessels (extravasation), followed by survival and proliferation in target tissues.Citation51 The effect of δT on the cascade of steps involved in metastasis was tested in NSCLC cells using in vitro assays: wound healing, cell invasion, cell adhesion assay, aggregation assay, and the MTS for cell proliferation. Based on our findings, δT inhibited all the metastasis functions in the above in vitro assays, suggesting its potential as an antimetastasis facilitator in A549 and H1299 cells. Furthermore, our previous studies, using histone enzyme-linked immunosorbent assay and Annexin-V stain-based flow cytometry analysis, demonstrated that the 0–30 μM range of δT was not necrotic to A549 or H1299 cells and instead induced apoptosis in a dose-dependent manner.Citation30 Therefore, antimetastasis functions of δT in in vitro assays are not due to necrotic effects.
The ECM is proteolytically degraded, facilitating the migration and invasion of cancer cells. Single cells detached from a tumor and released to the intercellular junctions during metastasisCitation52 are capable of making cell aggregates. Inherently, nontumor single cells are prone to anoikis, a form of apoptosis which is encouraged by insufficient or inappropriate cell–ECM interactions or attachments.Citation53 However, cancer cells showed resistance to anoikis, and as a result, they can survive after detachment from their main location.Citation51 Cancer cell aggregates exhibit lower levels of anoikis compared with regular apoptosis. Therefore, the formation of cell aggregates may induce regular cell apoptosis in cancer cells. However, we observed a dose-dependent decrease in cell aggregate formation as well as a reduction in cell adhesion capabilities with δT treatment. This may have occurred due to the stimulation of anoikis in nonadhesive cancer cells/single migrating cells and an induction of regular apoptosis in cell aggregates. Future studies are needed to explain δT-based apoptosis approaches in A549 and H1299 cells.
The Notch-1 signaling pathway plays a significant function in regulating cancer cell invasion and metastasis. Notch-1 expression was shown to increase in some NSCLC patients and is found to be associated with a TNM stage in histological grading,Citation54 suggesting that Notch-1 may play a fundamental role in the metastasis of NSCLC. Notch-1 inhibition decreases the development of brain metastases from breast cancer.Citation55 Abnormal Notch-1 expression is also strongly correlated with metastatic tumor disease in hepatocellular carcinoma patients.Citation56 Silencing of Notch-1 inhibited the invasion of human pancreatic cancer cells by inhibiting the expression of MMP-9 and uPA.Citation57 Interestingly, our study found that δT dose dependently inhibited the Notch-1 expressions compared with the control. To further validate its activity, we investigated the expression of Notch-1 downstream protein HES-1, as it is well documented that the activated Notch-1 pathway can increase the expression of its downstream HES-1 activities. Indeed, we observed that δT downregulated HES-1 protein expression dose dependently along with Notch-1 expression in A549 and H1299 cells, which suggests that δT is a potential inhibitor of Notch-1 signaling pathway.
Many studies have established that miR-451 is widely dysregulated in human malignancies, including lung cancer,Citation38,Citation58 gastric cancer,Citation59 breast cancer,Citation60 glioma,Citation61 and leukemia,Citation62 indicating the important role of miR-451 in oncogenesis. The downregulation of miR-451 has been reported in gastric and colon cancers and childhood B-cell precursor acute lymphoblastic leukemia.Citation63 miR-451 functions as a tumor suppressor and regulates survival in NSCLC cells.Citation55 It is also one of the most downregulated miRs in lung cancer patients.Citation55 Our results demonstrated that δT treatment induced the expression of miR-451 in a dose-dependent manner in both A549 and H1299 NSCLC cell lines. Moreover, the induction of miR-451 expression by δT occurred with a suppression of the Notch-1 signaling pathway. These findings reveal possible cross talk between Notch-1 signaling and miR-451 for regulation of cell migration and invasion in A549 and H1299 cell lines. This corroborates previous findings where the suppression of miR-451 was shown to induce Notch-1-induced oncogenesis in T-cell acute lymphoblastic leukemia.Citation62
Although a variety of stimuli from different signaling pathways are known to activate NF-κB, Notch-1 is considered as one of the key pathways in the activation of NF-κB via its ligands.Citation64 The downregulation of Notch-1 has been shown to inhibit invasion by the inactivation of NF-κB and MMP-9 in pancreatic cancer cells,Citation47 lung cancer,Citation29 and prostate cancer.Citation65 In this study, the data showed a dose-dependent decrease in NF-κB DNA-binding activity in A549 and H1299 cells with different concentrations of δT. NF-κB has multiple binding targets including the MMP-9 promoter in cancer cells.Citation66 Regulatory elements in the 670-bp promoter region of MMP-9 include activator protein (AP)-1, AP-2, specificity of protein 1, and NF-κB.Citation18 Mutation of the NF-κB binding site of MMP-9 promoter leads to a dramatic reduction in MMP-9 promoter activation and MMP-9 expression.Citation18–Citation21 Therefore, NF-κB DNA-binding activities are very crucial for the expression of MMP-9 genes. Modulating NF-κB activation using several natural and artificial agents was shown to alter MMP-9 expression in lung cancer cell lines. For instance, stromal cell-derived factor 1 was reported to increase the invasiveness of A549 cells through activation of ERK/NF-κB signaling, which is responsible for the increase of MMP-9 expression of the cells.Citation67 Nifedipine exerts its NF-κB inhibiting activity by suppressing IKK-mediated Ikα degradation, subsequently suppressing MMP-9 expression and activity.Citation68,Citation69 In lung cancer cells, radiotherapy-induced MMP-9 expression and lung metastasis were reduced after blocking NF-κB using arsenic trioxide.Citation70 In this study, we observed a dose-dependent inhibition of MMP-9 expression at the protein and mRNA levels along with reduced NF-κB DNA-binding activities upon the δT treatment, suggesting that the binding of TF NF-κB was reduced at the MMP-9 promoter upon δT treatment. Further, several other studies have shown that δT inhibits the expression of NF-κB in different cells lines.Citation71–Citation73 Therefore, it is reasonable to suggest that the lower binding activities of NF-κB are due to the lower expression of NF-κB upon δT treatment.
δT has been shown to have a very broad range of therapeutic potentials; it is used as an antioxidant, analgesic, anti-inflammatory, antibacterial, antipyretic, antithrombotic, anticancer agent and has cardioprotective, hepatoprotective, hypoglycemic, and nephroprotective properties.Citation74,Citation75 Multiple downstream targets of δT have been summarized in several reviews.Citation74–Citation76 In our previous studies, we reported that δT has multiple downstream targets, namely HES-1, VEGF, BCL-2, BCL-XL, Cyclin D1, Survivin, P-mTOR, mTOR, P-AKT, AKT, S6K, LAT-1, ASCT2, and c-MYC.Citation28–Citation33 However, in this study, we targeted MMP-9 as a key metastasis mediator in NSCLC.Citation77
Other studies have reported that the antimetastatic effect of nutraceuticals is individually linked to different metastatic pathways. For example, the nutraceuticals carnosic acid, silibinin, curcumin, epigallocatechin gallate, quercetin, and baicalein inhibited cell migration and invasion by downregulating uPA and/or MMP-9 expression in cervical cancer cells, melanoma cells, human breast epithelial cells, oral cancer cells, and prostate and liver cancer cells, respectively.Citation78–Citation82 Liu et alCitation83 showed that γ-tocotrienol inhibits cell invasion and metastasis of the human gastric system by downregulating MMP-9 and MMP-2. Interestingly, our results demonstrated that the anti-metastatic functions of δT on the inhibition of invasion and migration in lung cancer cells are due to the suppression of Notch-1, NF-κB, MMP-9, or uPA along with induced miR-451. Thus, our data suggested that δT is an effective and efficient antimetastatic compound with multiple therapeutic targets.
Conclusion
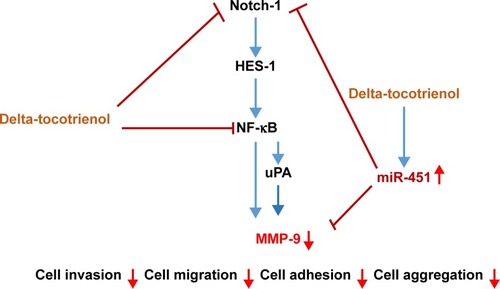
Our results provide mechanistic insight into the modulation of in vitro invasion and metastasis in lung cancer cells by δT. The data showed that δT suppressed cell migration, invasion, and adhesion in NSCLC cells by significantly lowering protease activity of MMP-9, inhibiting Notch-1-mediated NF-κB and uPA pathways, which led to the downregulated expression of MMP-9 with simultaneous increased expression of miR-451. Based on our findings, we propose that δT attenuates tumor aggressiveness, invasion, and metastasis by the downregulation of MMP-9 via Notch-1 and uPA pathways (). This study suggested consideration of δT as an effective potential natural therapeutic approach against lung cancer invasion and metastasis. Further studies to establish the efficacy of δT in preventing lung cancer migration and invasion at the animal and clinical levels are highly recommended.
Figure 4 Proposed mechanism by which delta-tocotrienol suppresses cell migration and invasion in A549 and H1299 cells.
Abbreviations: MMP-9, matrix metallopeptidase 9; NF-κB, nuclear factor-κB; uPA, urokinase-type plasminogen activator.
Acknowledgments
We thank American River Nutrition, Inc. for their kind gift of δT.
Disclosure
The authors report no conflicts of interest in this work.
References
- SiegelRLMillerKDJemalACancer statistics, 2017CA Cancer J Clin201767173028055103
- EgebladMWerbZNew functions for the matrix metalloproteinases in cancer progressionNat Rev Cancer20022316117411990853
- FingletonBMMPs 53Cancer Therapeutic Targets2017591
- HiratsukaSNakamuraKIwaiSMMP9 induction by vascular endothelial growth factor receptor-1 is involved in lung-specific metastasisCancer Cell20022428930012398893
- LinderRRönmarkEPourazarJBehndigABlombergALindbergASerum metalloproteinase-9 is related to COPD severity and symptoms-cross-sectional data from a population based cohort-studyRespir Res2015162825849664
- ZhuLJChengLSunNHouPFInvasion and metastasis significance of serum MMP-9 in lung cancer patientsGlobal Advances in Pharmaceutical Research2017411
- Blanco-PrietoSBarcia-CastroLPaez de la CadenaMRelevance of matrix metalloproteases in non-small cell lung cancer diagnosisBMC Cancer201717182329207990
- OsmanBAkoolel-SDollerAMüllerRPfeilschifterJEberhardtWDifferential modulation of the cytokine-induced MMP-9/TIMP-1 protease–antiprotease system by the mTOR inhibitor rapamycinBiochem Pharmacol201181113414320854798
- KangHJangSWRoles of matrix metalloproteinase-9 in cancer metastasisBiomed Sci Lett2014203103110
- PiperigkouZManouDKaramanouKTheocharisADStrategies to target matrix metalloproteinases as therapeutic approach in cancerMethods Mol Biol2018173132534829318564
- KopczynskaEDancewiczMKowalewskiJKardymowiczHTyrakowskiTThe serum concentration of metalloproteinase 9 and 2 in non-small cell lung cancer patientsPol Merkur Lekarski200722132539541 Polish17874625
- Ramos-DeSimoneNHahn-DantonaESipleyJNagaseHFrenchDLQuigleyJPActivation of matrix metalloproteinase-9 (MMP-9) via a converging plasmin/stromelysin-1 cascade enhances tumor cell invasionJ Biol Chem199927419130661307610224058
- CurranSMurrayGIMatrix metalloproteinases: molecular aspects of their roles in tumour invasion and metastasisEur J Cancer20003613 Spec1621163010959048
- DuffyMJUrokinase plasminogen activator and its inhibitor, PAI-1, as prognostic markers in breast cancer: from pilot to level 1 evidence studiesClin Chem20024881194119712142372
- SaldenMSplinterTAPetersHAThe urokinase-type plasminogen activator system in resected non-small-cell lung cancer. Rotterdam Oncology Thoracic Study GroupAnn Oncol200011332733210811500
- RaoJSGondiCChettyCChittiveluSJosephPALakkaSSInhibition of invasion, angiogenesis, tumor growth, and metastasis by adenovirus-mediated transfer of antisense uPAR and MMP-9 in non–small cell lung cancer cellsMol Cancer Ther2005491399140816170032
- TongYYueJMaoMLiuQZhouJYangJRecombinant nematode anticoagulant protein c2 inhibits cell invasion by decreasing uPA expression in NSCLC cellsOncol Rep20153341815182225672417
- WuHTSieSSKuanTCLinCSIdentifying the regulative role of NF-κB binding sites within promoter region of human matrix metalloproteinase 9 (mmp-9) by TNF-α inductionAppl Biochem Biotechnol2013169243844923224948
- BondMChaseAJBakerAHNewbyACInhibition of transcription factor NF-kappaB reduces matrix metalloproteinase-1, -3 and -9 production by vascular smooth muscle cellsCardiovasc Res200150355656511376631
- RheeJWLeeKWKimDNF-kappaB-dependent regulation of matrix metalloproteinase-9 gene expression by lipopolysaccharide in a macrophage cell line RAW 264.7J Biochem Mol Biol2007401889417244487
- VincentiMPBrinckerhoffCESignal transduction and cell-type specific regulation of matrix metalloproteinase gene expression: can MMPs be good for you?J Cell Physiol2007213235536417654499
- WangSLiuZWangLZhangXNF-kappaB signaling pathway, inflammation and colorectal cancerCell Mol Immunol20096532733419887045
- SuhJRabsonABNF-kappaB activation in human prostate cancer: important mediator or epiphenomenon?J Cell Biochem200491110011714689584
- PeriSDevarajanKYangDHKnudsonAGBalachandranSMeta-analysis identifies NF-κB as a therapeutic target in renal cancerPLoS One2013810e7674624116146
- BelottiDCalcagnoCGarofaloAVascular endothelial growth factor stimulates organ-specific host matrix metalloproteinase-9 expression and ovarian cancer invasionMol Cancer Res20086452553418403633
- YanMXuQZhangPZhouXJZhangZYChenWTCorrelation of NF-κB signal pathway with tumor metastasis of human head and neck squamous cell carcinomaBMC Cancer20101043720716363
- LiuXZhangXXiangJLvYShiJmiR-451: potential role as tumor suppressor of human hepatoma cell growth and invasionInt J Oncol201445273974524841638
- RajasingheLDGuptaSVDelta tocotrienol inhibit mTOR pathway by modulating glutamine uptake and transporters in non-small cell lung cancerFASEB J201630Suppl 1688.61626481308
- RajasingheLGuptaSTocotrienols suppress non-small lung cancer cells via downregulation of the Notch-1 signaling pathway (644.1)FASEB J201428Suppl 1644.64124174427
- JiXWangZGeamanuASarkarFHGuptaSVInhibition of cell growth and induction of apoptosis in non-small cell lung cancer cells by delta-tocotrienol is associated with notch-1 down-regulationJ Cell Biochem2011112102773278321598300
- RajasingheLPindiproluRRazalliNWuYGuptaSDelta tocotrienol inhibits MMP-9 dependent invasion and metastasis of non-small cell lung cancer (NSCLC) cell by suppressing Notch-1 mediated NF-κb and uPA pathwaysFASEB J201529Suppl 1752.718
- RajasingheLDGuptaSVTocotrienol-rich mixture inhibits cell proliferation and induces apoptosis via down-regulation of the Notch-1/NF-κB pathways in NSCLC cellsNutr Diet Suppl20179103114
- RajasingheLDAnti-Cancer Effects of Tocotrienols in NSCLCDetroit, MIWayne State University2017
- GunathilakaGUTahlanVMafizAPolurMZhangYPhages in urban wastewater have the potential to disseminate antibiotic resistanceInt J Antimicrob Agents201750567868328803933
- GunathilakaGWickramasingheH18s rDNA based molecular characterization of locally isolated Aspergillus oryzaeProceedings of the Peradeniya University Research Sessions2009143
- GunathilakaGUCharacterization of Bacteriophages from Environmental Water Samples and the Potential of Bacteriophages Tailspike Proteins (tsp) in Bacteria Detection [Wayne State University Theses]Detroit, MIWayne State University2014
- LivakKJSchmittgenTDAnalysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT methodMethods200125440240811846609
- BianHBPanXYangJSWangZXDeWUpregulation of microRNA-451 increases cisplatin sensitivity of non-small cell lung cancer cell line (A549)J Exp Clin Cancer Res2011302021329503
- ChenDHuangJZhangKMicroRNA-451 induces epithelial-mesenchymal transition in docetaxel-resistant lung adenocarcinoma cells by targeting proto-oncogene c-MycEur J Cancer201450173050306725310895
- HuangJChenYLiJNotch-1 confers chemoresistance in lung adenocarcinoma to taxanes through AP-1/microRNA-451 mediated regulation of MDR-1Mol Ther Nucleic Acids2016510e37527727250
- TianFHanYYanXUpregulation of microrna-451 increases the sensitivity of A549 cells to radiotherapy through enhancement of apoptosisThorac Cancer20167222623127042226
- GotoATanakaMYoshidaMThe low expression of miR-451 predicts a worse prognosis in non-small cell lung cancer casesPLoS One2017127e018127028704499
- YinPPengRPengHMiR-451 suppresses cell proliferation and metastasis in A549 lung cancer cellsMol Biotechnol201557111125150396
- LevičarNNutallRKLahTTProteases in brain tumour progressionActa Neurochir (Wein)2003145982583814505115
- MoirangthemABondhopadhyayBMukherjeeMSimultaneous knockdown of uPA and MMP9 can reduce breast cancer progression by increasing cell-cell adhesion and modulating EMT genesSci Rep201662190326906973
- LiLTanJZhangYDLK1 promotes lung cancer cell invasion through upregulation of MMP9 expression depending on Notch signalingPLoS One201493e9150924621612
- WangZBanerjeeSLiYRahmanKMZhangYSarkarFHDown-regulation of notch-1 inhibits invasion by inactivation of nuclear factor-kappaB, vascular endothelial growth factor, and matrix metalloproteinase-9 in pancreatic cancer cellsCancer Res20066652778278416510599
- JouveCPalmeirimIHenriqueDNotch signalling is required for cyclic expression of the hairy-like gene HES1 in the presomitic mesodermDevelopment200012771421142910704388
- St-PierreYVan ThemscheCEstevePOEmerging features in the regulation of MMP-9 gene expression for the development of novel molecular targets and therapeutic strategiesCurr Drug Targets Inflamm Allergy20032320621514561155
- WestermarckJKähäriVMRegulation of matrix metalloproteinase expression in tumor invasionFASEB J199913878179210224222
- SimpsonCDAnyiweKSchimmerADAnoikis resistance and tumor metastasisCancer Lett2008272217718518579285
- KwonGTChoHJChungWYParkKKMoonAParkJHIsoliquiritigenin inhibits migration and invasion of prostate cancer cells: possible mediation by decreased JNK/AP-1 signalingJ Nutr Biochem200920966367618824345
- FrischSMScreatonRAAnoikis mechanismsCurr Opin Cell Biol200113555556211544023
- JinMMYeYZQianZDZhangYBNotch signaling molecules as prognostic biomarkers for non-small cell lung cancerOncol Lett20151053252326026722321
- McGowanPMSimedreaCRibotEJNotch1 inhibition alters the CD44hi/CD24lo population and reduces the formation of brain metastases from breast cancerMol Cancer Res20119783484421665937
- WangXQZhangWLuiELNotch1-Snail1-E-cadherin pathway in metastatic hepatocellular carcinomaInt J Cancer20121313E163E17222052196
- Bin HafeezBAdhamiVMAsimMTargeted knockdown of Notch1 inhibits invasion of human prostate cancer cells concomitant with inhibition of MMP9 and uPAClin Cancer Res200915245245919147749
- WangRWangZXYangJSPanXDeWChenLBMicroRNA-451 functions as a tumor suppressor in human non-small cell lung cancer by targeting ras-related protein 14 (RAB14)Oncogene201130232644265821358675
- BandresEBitarteNAriasFmicroRNA-451 regulates macrophage migration inhibitory factor production and proliferation of gastrointestinal cancer cellsClin Cancer Res20091572281229019318487
- BergamaschiAKatzenellenbogenBSTamoxifen downregulation of miR-451 increases 14-3-3ζ and promotes breast cancer cell survival and endocrine resistanceOncogene2012311394721666713
- GalHPandiGKannerAAMIR-451 and Imatinib mesylate inhibit tumor growth of Glioblastoma stem cellsBiochem Biophys Res Commun20083761869018765229
- LiXSandaTLookATNovinaCDvon BoehmerHRepression of tumor suppressor miR-451 is essential for NOTCH1-induced oncogenesis in T-ALLJ Exp Med2011208466367521464222
- JuXLiDShiQHouHSunNShenBDifferential microRNA expression in childhood B-cell precursor acute lymphoblastic leukemiaPediatr Hematol Oncol200926111019206004
- OsipoCGoldeTEOsborneBAMieleLAOff the beaten pathway: the complex cross talk between Notch and NF-[kappa]BLab Invest2007881111718059366
- WangZLiYBanerjeeSDown-regulation of Notch-1 and Jagged-1 inhibits prostate cancer cell growth, migration and invasion, and induces apoptosis via inactivation of Akt, mTOR, and NF-kappaB signaling pathwaysJ Cell Biochem2010109472673620052673
- Boston University BiologyNF-κB Target Genes2018 Available from: https://www.bu.edu/nf-kb/gene-resources/target-genes/Accessed November 3, 2018
- TangCHTanTWFuWMYangRSInvolvement of matrix metalloproteinase-9 in stromal cell-derived factor-1/CXCR4 pathway of lung cancer metastasisCarcinogenesis2008291354317916907
- GaoXIwaiMInabaSAttenuation of monocyte chemoattractant protein-1 expression via inhibition of nuclear factor-kappaB activity in inflammatory vascular injuryAm J Hypertens200720111170117517954363
- TomitaNYamasakiKIzawaKInhibition of experimental abdominal aortic aneurysm progression by nifedipineInt J Mol Med200821223924418204791
- WeiLHLaiKPChenCAArsenic trioxide prevents radiation-enhanced tumor invasiveness and inhibits matrix metalloproteinase-9 through downregulation of nuclear factor κBOncogene2004243390398
- HusainKFrancoisRAYamauchiTPerezMSebtiSMMalafaMPVitamin E δ-tocotrienol augments the anti-tumor activity of gemcitabine and suppresses constitutive NF-κB activation in pancreatic cancerMol Cancer Ther201110122363237221971120
- YamasakiMNishimuraMSakakibaraYSuikoMMorishitaKNishiyamaKDelta-tocotrienol induces apoptotic cell death via depletion of intracellular squalene in ED40515 cellsFood Funct20145112842284925225850
- WuSJLiuPLNgLTTocotrienol-rich fraction of palm oil exhibits anti-inflammatory property by suppressing the expression of inflammatory mediators in human monocytic cellsMol Nutr Food Res200852892192918481320
- SailoBLBanikKPadmavathiGJavadiMBordoloiDKunnumak-karaABTocotrienols: the promising analogues of vitamin E for cancer therapeuticsPharmacol Res201813025927229496592
- AhsanHAhadAIqbalJSiddiquiWAPharmacological potential of tocotrienols: a reviewNutr Metabol (Lond)201411152
- ComitatoRAmbraRVirgiliFTocotrienols: a family of molecules with specific biological activitiesAntioxidants (Basel)201764 pii:E93
- GongLWuDZouJPrognostic impact of serum and tissue MMP-9 in non-small cell lung cancer: a systematic review and meta-analysisOncotarget2016714184581846826918342
- AgarwalASharmaVTewariRKoulNJosephCSenEEpigallocatechin-3-gallate exhibits anti-tumor effect by perturbing redox homeostasis, modulating the release of pro-inflammatory mediators and decreasing the invasiveness of glioblastoma cellsMol Med Rep20081451151521479441
- LinCMChenYHMaHPSilibinin inhibits the invasion of IL-6-stimulated colon cancer cells via selective JNK/AP-1/MMP-2 modulation in vitroJ Agric Food Chem20126051124511245723210512
- LeeKWKimJHLeeHJSurhYJCurcumin inhibits phorbol ester-induced up-regulation of cyclooxygenase-2 and matrix metalloproteinase-9 by blocking ERK1/2 phosphorylation and NF-κB transcriptional activity in MCF10A human breast epithelial cellsAntioxid Redox Signal2005711–121612162016356124
- ChouRHHsiehSCYuYLHuangMHHuangYCHsiehYHFisetin inhibits migration and invasion of human cervical cancer cells by down-regulating urokinase plasminogen activator expression through suppressing the p38 MAPK-dependent NF-κB signaling pathwayPLoS One201388e7198323940799
- BarniMCarliniMJCafferataEGPuricelliLMorenoSCarnosic acid inhibits the proliferation and migration capacity of human colorectal cancer cellsOncol Rep20122741041104822246562
- LiuHKWangQLiYInhibitory effects of gamma-tocotrienol on invasion and metastasis of human gastric adenocarcinoma SGC-7901 cellsJ Nutr Biochem201021320621319195866