
Abstract
In recent years, the PD-1/PD-L1 axis blockade has become a very promising therapy with significant clinical benefits for multiple tumor types. However, some patients still do not respond sufficiently to PD-1/PD-L1 targeted monotherapy. Therefore, investigating the mechanism of PD-1 blockade resistance will assist in exploring new immunotherapy strategies, controlling the progress of the disease, and thus bringing more sustainable survival benefits to patients. The tumor-immune cycle is divided into the following seven steps: the release of cancer antigens, cancer antigen presentation, priming and activation, trafficking of T cells to tumors, infiltration of T cells into tumors, recognition of cancer cells by T cells, and killing of cancer cells. Given that PD-1/PD-L1 blockade is primarily involved in step 7, any abnormalities in the previous steps may affect the efficacy of PD-1/PD-L1 inhibitors and lead to drug resistance. This review discussed the resistance mechanisms of PD-1/PD-L1 blockade in each cancer-immunity step to finding a more suitable treatment population and an optimized combination therapy to exert immunotherapy in tumor treatment to a greater extent.
Keywords:
Introduction
Immunotherapy is considered as a promising method of treating cancers.Citation1 The 2018 Nobel Prize in Physiology or Medicine was awarded to James P. Allison and Tasuku Honjo, who discovered CTLA-4, and PD-1 separately. Immunotherapy has brought new directions to cancer treatment. PD-1/PD-L1 blockade has been used in the treatment of melanoma, non-small cell lung cancer (NSCLC),Citation2 bladder carcinoma,Citation3,Citation4 Hodgkin’s lymphoma,Citation5,Citation6 and Merkel cell carcinoma.Citation7,Citation8 However, in the actual clinical practice, the presence of drug resistance reduces the efficacy of PD-1/PD-L1 blockade.
Chen and Mellman (2013) proposed the concept of “the Cancer-Immunity Cycle”, helping us to understand the multi-step complexity of cancer-immunity clearly.Citation9 Tumor antigen is captured and processed into an immunogenic polypeptide by antigen-presenting cells (APCs) and is present on the APCs surface in the form of an antigen peptide-MHC molecular complex. The antigenic peptide-MHC molecules on the surface of the APCs bind to the TCR on the surface of the T cells. At the same time, the costimulatory molecules expressed by APC bind to the corresponding ligands on the surface of T cells and then activate antigen-specific T cell immune responses. PD-1/PD-L1 blockade targets the final step. It can be affected by abnormalities in any of the previous steps to produce resistance. This review discussed the resistance mechanisms of PD-1/PD-L1 blockade at each step ().
Figure 1 Resistance mechanism of PD-1/PD-L1 blockade in the cancer-immunity cycle. Each step of the cancer-immunity cycle requires the coordination of numerous factors. PD-1/PD-L1 blockade targets the final step. It can be affected by abnormalities in any of the previous steps to produce resistance. Examples of such abnormalities and the primary steps at which they can act are shown.
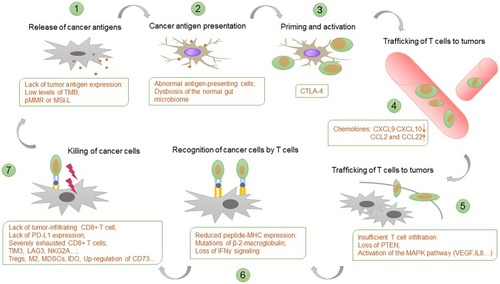
Lack of Tumor Antigen Expression
Tumor Mutation Burden (TMB) is defined as the total number of somatic gene coding errors, base substitutions, gene insertions, or deletion errors detected per million bases. Mutations of somatic cells can be transcribed or expressed into RNA and protein levels to produce neoantigen, protein fragments, or polypeptide segments, which can be recognized by the immune system as “non-self,” activating T cells and resulting in an immune response.Citation10,Citation11 Therefore, when the number of gene mutations accumulated per megabase increases, many neoantigens are produced. This outcome caused the tumor to be attacked by tumor-specific T cells, increasing its sensitivity to PD-1 treatment.Citation12–Citation14 High levels of TMB (>20 mut/Mb) results in high immunogenicities, which are sensitive to PD-1 blockade, as in melanomas, renal cell carcinomas, and non-small cell carcinomas.Citation11 Low levels of TMB (<10 mut/Mb) results in poor immunogenicities, as in pancreatic cancers and prostate cancers.Citation14
Tumors lacking DNA Damage Response (DDR) pathway are less efficient at correcting genetic damage and are therefore associated with increased TMB, new antigen loading, and a better response to Immune-Checkpoint Blockers (ICBs).Citation15 The human mismatch repair genes (MMR genes), such as MLH1, MSH2, MSH6, and PMS2, are responsible for repairing DNA mismatches in the genome.Citation16 MMR defects lead to the accumulation of base mismatch in the DNA replication process, resulting in the occurrence of microsatellite instability (MSI). About 15% of colorectal cancers are related to the MSI pathway. MSI is classified into high-level instability (MSI-H), low-level instability (MSI-L) and microsatellite-stable subtype (MS-S). MMR is classified into defective DNA mismatch repair (dMMR) and proficient mismatch repair (pMMR). dMMR is equivalent to MSI-H, and pMMR is equivalent to MSI-L or MSS. Patients with colorectal cancer experiencing dMMR or MSI-H are more likely to benefit from immunotherapy.Citation17 However, PD-1/PD-L1 drug therapy is unsatisfactory in patients with pMMR or MSI-L. The antigen expression is low, and the immunogenicity is deficient in these patients, failing T cell activation.
Lack of Effective Antigen Presentation and T Cell Initiation
Abnormal Antigen-Presenting Cells
Antigen-presenting cells (APCs) process antigens and present them to T-cells and are therefore vital for an effective adaptive immune response. Dendritic cells (DCs) are one of the essential APCs which can activate unsensitized naive T cells. Most of the DCs in the human body are in an immature state, expressing low levels of costimulatory factors and adhesion factors. Immature DCs have the powerful antigenic phagocytic ability. When stimulated by antigens, immature DCs differentiate into mature DCs, which express high levels of costimulatory and adhesion factors. During the maturation process, DCs migrate from the peripheral tissues into the secondary lymphoid organs, contacting T cells and stimulating an immune response. DCs, as the most potent APCs currently discovered, are capable of inducing specific cytotoxic T lymphocytes (CTLs) production.Citation18
However, DCs often do not function efficiently in the tumor environment. The tumor microenvironment can affect the distribution, differentiation, and maturation of DCs. (1) The tumor microenvironment affects DCs distribution. Studies have shown that the number of DCs in peripheral blood of patients with head and neck squamous cell carcinoma is different from that of healthy people. The bone marrow-derived DCs precursor cells in peripheral blood of patients with tumors were significantly lower than normal. In a series of studies on tumor prognosis, it was found that there was a significant correlation between abnormalities in the number of DCs in peripheral blood and poor prognosis. DCs infiltrated in tumor tissues are mostly immature phenotypes, while DCs in adjacent tissues are mostly mature DCs. Tumor-derived cytokines and chemokines can recruit immature DCs to migrate to tumor tissue, subsequently limit their maturation and function. (2) The tumor microenvironment affects DCs differentiation. DCs are antigen-presenting cell populations derived from CD34 hematopoietic stem cells. Studies showed that tumor cells and related factors in the tumor microenvironment could interfere with the differentiation of DC precursor cells, resulting in decreased or even loss of DCs’ activities. It was confirmed that IL-6 and granulocyte colony-stimulating factor (G-CSF) secreted by tumor cells blocked the differentiation of CD34 cells into DCs. IL-6 inhibits the differentiation of precursor cells into DCs by activating the STAT3 signaling pathway. VEGF secreted by tumor cells, which promotes tumor blood vessel growth, is directly involved in local immunosuppression. VEGF can block the differentiation of CD34 cells into DCs by inhibiting the activity of NF-kB in cells. Recent studies showed that prostaglandin E2 (PGE2) produced by tumor cells inhibited the differentiation of bone marrow precursor cells and monocytes into DCs.Citation19 (3) The tumor microenvironment affects DCs maturation. IL-10 is an essential regulator of DCs maturation and function. Studies showed that IL-10 mainly plays a role in blocking DCs maturation and activation of T cells, but does not affect the migration of mature DCs to regional lymph nodes. IL-10 negatively regulates the expression of DCs co-stimulatory molecules and the secretion of pro-inflammatory factors such as IL-12 and TNF by activating the STAT3 signaling pathway in immature DCs, making it unable to provide a second signal for T cells activation. Tumor cells, stromal cells, and various infiltrating immune cells in the tumor microenvironment can highly express PGE2. Functions of PGE2 are different due to different maturity degree and distribution of DCs. In peripheral tissues, PGE2 allows DCs to overexpress CCR7 and promote DCs migration to secondary lymphoid organs. When DCs migrate to the secondary lymphoid organs, PGE2 increases the secretion of IL-10 by DCs themselves, inhibits DCs maturation, significantly decreases the expression of MHC molecules, and decreases the ability to activate T cells.Citation20–Citation22
Dysbiosis of the Normal Gut Microbiome
An increasing number of studies have shown that the human gut microbiome can affect the development of cancer and the effect of immunotherapy.Citation23 Patients who respond well to treatment with PD-1 inhibitors show a notable difference in gut microbiome structure compared to patients who do not respond well to the same treatment.Citation24 Through large-scale tests, it was found that bifidobacteria may enhance the body’s immune response by stimulating DCs directly, inducing DCs maturation and cytokine secretion, contributing to the activation of tumor-killing T cells and the enhancement of PD-1/PD-L1 blockade.Citation25,Citation26 Akkermansiacea muciniphila can enhance the response of PD-1/PD-L1 blockade by increasing CD4+ central memory T cell (TCM), CD4/Foxp3 ratio, and IL-12 production in the tumor bed and increasing IFN-γ production.Citation23 TCM expresses chemokine receptor CXCR3 or CCR9. CXCR3 is related to the recruitment of Th1 cells to inflamed lesions, while the CCR9/CCL25 axis is associated with chemotactic migration of T cells, especially in intestine and colon.Citation27 Besides, Enterococcus hirae combined with Akkermansia muciniphila may elevate TCM and increase the production of IFN-γ so that it had a synergistic effect with PD-1 blockade combined with Akkermansia muciniphila.Citation23 Collinsella aerofaciens and Enterococcus faecium were related to enhanced efficacy of PD-1 blockade by leading to decreased peripherally derived Tregs.Citation28 Significant differences were observed in the diversity and composition of the patient gut microbiome of responders (R) versus non-responders (NR) in melanoma patients receiving PD-1 blockade therapy. Immune profiling suggested enhanced systemic and anti-tumor immunity in responding patients with a favorable gut microbiome.Citation27 In conclusion, the gut microbiome can affect at a critical point in the anti-tumor response, and the stable gut microbiome can enhance the immune surveillance of tumor cells.
Migration Disorders of T Cells
Chemokines are low molecular weight proteins that are capable of attracting immune cells to the site of infection. They play a two-way regulatory role in tumor immune microenvironment. On the one hand, stromal cells in tumors and tumor tissues recruit CXCL9, CXCL10, CXCL11, CCL3, CCL4, CCL5, and other chemokines to recruit NK cells and CTLs, causing anti-tumor immune response.Citation29,Citation30 On the other hand, high levels of chemokines such as CCL2 and CCL22 in the tumor microenvironment, inhibit the immune response by recruiting immunosuppressive cells such as MDSCs, Tregs, and M2 cells. Therefore, the level of chemokines in the tumor tissue determines the immune status of the tumor. For example, CXCL9 and CXCL10 silencing caused by DNA methylation inhibit T cells homing and thus cause resistance to immune checkpoint drugs.Citation30,Citation31 The use of Hypomethylating agents (HMAs) reduces the inhibition of chemokines and increases TILs, thereby increasing the therapeutic effect of PD-1/PD-L1 blockade.Citation32
Insufficient T Cell Infiltration
Loss of PTEN
The PI3K-AKT-mTOR pathway is a crucial signal transduction pathway encompassing several oncogenes. This pathway is involved in essential cellular processes such as cell survival, proliferation, and differentiation. In patients, the loss of PTEN is associated with a reduction of T cells infiltration in the tumor microenvironment and a weaker effect of PD-1 blockade treatment. In the mouse model of melanoma, Peng et al (2016) observed that the loss of PTEN in tumor cells increased the expression of immunosuppressive cytokines, which reduced the infiltration of T cells and inhibited autophagy. Treatment with a selective PI3Kβ inhibitor improved the efficacy of anti-PD-1 antibodies.Citation33 In a mouse model of prostate tumor, Garcia et al (2014) found that PTEN-null mice exhibited high levels of infiltration of MDSCs, which excluded CD8+ and NK cells from TME and reduced their cytotoxic activity.Citation34 Besides, PTEN-loss reduced autophagic activities in melanoma, reducing T cell-mediated tumor apoptosis.Citation35 Overexpressing MAP1LC3B, which is critical in autophagy initiation, improved T-cells killing, while the resistance to T cells killing was observed in MAP1LC3B-silenced tumor cells. Studies showed that overexpression of MAP1LC3B in PTEN-silenced tumor cells completely restored sensitivity to T-cells killing. Furthermore, pretreatment of melanoma cells with the autophagy inhibitor hydroxychloroquine (HCQ) reduced the apoptosis induced by autologous TILs.Citation33
Activation of the MAPK Pathway
The MAPK pathway involves many different cellular processes such as proliferation, differentiation, movement, apoptosis, and survival. The pathway’s carcinogenic effects have been confirmed.Citation36 Emerging evidence established the role of the MAPK pathway in regulating the immune response in TME. Activation of the MAPK pathway leads to the production of VEGF and IL-8, both of which are angiogenic factors that inhibit the recruitment and infiltration of T cells.Citation37,Citation38 In human melanoma samples, inhibition of the MAPK pathway promotes CD8+ T cells activation and infiltration. Experiments showed that a combination of PD-1 antibody and MEK blockade treatment strategy enhanced immunocyte infiltration in the tumor bed and can improve treatment outcomes.Citation39
Incompetent Recognition of Tumor Cells by T Cells
Mutations of β-2-Microglobulin
Two signals are required for T cells activation. The first signal is initiated by TCR recognition of the antigenic peptide-MHC complex (p-MHC) on the surface of the APCs; the second signal is initiated by the interaction of co-stimulatory molecules between the T cells and the APCs. Both are required to produce an effective immune response. Tumors become targets of specific cytotoxic T lymphocytes (CTLs) by expressing many different antigen peptide MHC molecule complexes. However, in some cases, tumors lose their antigen-presenting ability by genetic mutation or epigenetic modification, thereby escaping immune recognition.
The lack of MHC class I molecules is one of the leading causes of PD-1 blockade resistance. The lack can be caused by the decrease of mRNA transcription level of MHC molecules, the loss of genome, and the mutation of B microglobulin gene.Citation40 The B2M gene encodes B2M protein, which is an indispensable component of MHC class I molecules.Citation41 Thus, mutation of the B2M gene is a mechanism of anti-PD-1 antibody treatment for resistance.Citation42
Loss of IFNγ Signaling
IFNγ produced by tumor-specific T cells can recognize tumor cells or corresponding receptors on APC, thereby exerting a powerful anti-tumor immune effect. IFNγ up-regulates the expression of MHC molecules and PD-L1, thus enhancing tumor antigen presentation and response to PD-1 treatment.Citation41 IFNγ can also recruit other immune cells or directly inhibit tumor cell proliferation and promote apoptosis. Therefore, mutations and deletions of tumor cells IFNγ pathway-related proteins (such as IFNγ receptor IFNGR1 and IFNGR2, IFNγ receptor chain JAK1 and JAK2, STATs, and IRF1) can lead to resistance to ICIs.Citation43,Citation44
Abnormalities of T Cell Killing Tumors
Lack of Tumor-Infiltrating CD8+ T Cells
A certain amount of TILs in TME is the basis for judging the efficacy of checkpoint blockade. Chen DS and Mellman I (2017) published a review proposing a new classification of immune tumors with three phenotypes; immune-desert, immune-excluded, and inflamed. Among these phenotypes, the immune-desert and immune-excluded phenotypes are cold tumors (non-inflamed), and the density of CD8+ T cells in the tumors is low.Citation45
Lack of PD-L1 Expression
Of course, it is not enough to focus on CD8+ T cells, but also on PD-L1 expression.Citation46 A threshold of 5% PD-L1 positive expression is used to determine PD-L1 positivity. Subsequent studies showed a higher objective response rate for anti-PD-1 therapy in a population of patients with PD-L1 positive disease. Progression-free survival (PFS) and overall survival (OS) were also improved in PD-L1-positive advanced melanoma and NSCLC patients when PD-L1 expression subgroup analysis was performed.Citation47,Citation48 However, there are still some problems with PD-L1 detection. The most important thing is that the expression of PD-L1 is very dynamic and unevenly distributed in tumor tissues, which is likely to cause false-negative results. Besides, antibodies used in some tests may be insufficiently sensitive and may also cause false negatives. Now some researchers are optimizing the method of detecting PD-L1 and hoping to bring better prediction results by then.
Based on TILs and PD-L1 expression, Teng et al (2015) divided the tumor microenvironment into four categories. These include type I, (PD-L1+ with TILs), type II (PD-L1− with no TILs), type III (PD-L1+ with no TILs), and type IV (PD-L1− with TILs).Citation49 Among these four types, type I is the optimal candidate for PD-1/PD-L1 treatment.
Severely Exhausted CD8+ T Cells
In vitro studies have shown that the intensity of the PD-1 signal, namely its relative expression level or ligand abundance, can determine the severity of T cells exhaustion and thus affect the sensitivity to anti-PD-1 therapy. In some mouse models, PD-1 expression is high, and T cells are severely anergic, which is associated with resistance to anti-PD-1 therapy.Citation50 After PD-1 treatment in patients with low or moderate expression of PD-1, exhausted T cells can be reactivated to exert their immune effects. Therefore, whether PD-1 blockade intervention can effectively reverse T cells anergy may depend on the ratio of low-medium expression to high expression of T cells before treatment.
Activation of Other Inhibitory Immune Checkpoints
In the anti-tumor immunity process, there are some other immune checkpoints, including CTLA-4, TIM3, LAG3, NKG2A, and so on, which are gradually recognized and participate in co-regulating immune responses with PD-1/PD-L1 treatment. However, which immunological checkpoints are dominant and whether the intensity of different immune checkpoints is different in different tumors or tumor development stages remains to be studied.
CTLA-4
CTLA-4 is also an inhibitory immune checkpoint molecule whose ligands are CD80 and CD86 on APCs. CTLA-4 is primarily expressed on regulatory T cells (Tregs), Tregs are negative immunoregulatory cells which have inhibitory effects on tumor immunity.Citation51,Citation52 The combination of anti-PD-1 and anti-CTLA-4 synergistically reduces inhibitory signals and enhances anti-tumor response.Citation53 The efficacy of the clinical combination of CTLA-4 and PD-1/PD-L1 mAbs has been confirmed.Citation54 Therefore, screening of the best combination of immune checkpoints to achieve optimal clinical efficacy will be the direction of future research.Citation55
LAG-3
Lymphocyte activating gene 3 (LAG-3) is an inhibitory immune checkpoint molecule expressed by T cells, NK cells, and B cells. LAG-3 blockade can increase the number of activated CD8+ T cellsCitation56 and inhibit the activity of TregsCitation57 in the tumor microenvironment, thereby restoring anti-tumor immunity.Citation58 Additionally, anti-LAG-3 checkpoint blockade, in combination with tumor-specific CD4+ T cells and irradiation, overcame PD-L1 mAbs primary resistance and treated established tumors resulting in fewer recurrences.Citation59 LAG-3 expression on TILs was significantly correlated with that of PD-1 expression and was also significantly associated with negative prognostic factors.Citation60,Citation61 Surprisingly, the combination of the LAG-3 blockade and PD-1 blockade was more effective than the PD-1 blockade alone. Studies in other multiple models have shown significant inhibition of tumors by the combination of LAG-3 blockade and PD-1 blockade.Citation62
TIM-3
T cell immunoglobulin mucin-3 (TIM-3) plays a vital role in regulating TH1 cell-mediated immune responses. Inhibition of the TIM-3 pathway reduced the infiltration of Tregs and thus slowed down the growth of mouse Lewis lung cancer.Citation63 Koyama et al (2016) observed an increase in TIM-3 expression in TILs following PD-1 blockade acquired resistance. The expression degree of TIM-3 was also significantly correlated with the stage of PD-1 treatment. The positive expression was lower before treatment and during the sensitive period and was significantly increased after drug resistance. Moreover, the positive expression of TIM-3 was also significantly correlated with the degree of PD-1 antibody binding in T cells. The higher the degree of T cells binding to PD-1 blocking, the stronger the TIM-3 positive expression.Citation64
Limagne et al (2019) observed that the accumulation of lymphocytes and monocytes MDSCs (mMDSCs) expressing Tim-3 and galectin-9 after nivolumab treatment of a cohort of NSCLC was associated with primary or acquired resistance.Citation65 This result indicated that the Galectin-9/Tim-3 pathway might be a vital mechanism for resistance to PD-1 treatment and maybe a new target for immunotherapy drug combinations.
NKG2A
NKG2A is an inhibitory receptor-expressing both on CD8+ T cells and NK cells, whose main ligand is a non-canonical MHC-I molecule: HLA-E (human) or Qa-1b (murine). Blocking NKG2A can simultaneously abolish the immunosuppression of CD8+ T cells and NK cells.Citation66 Mingari et al (2019) used a kind of humanized anti-NKG2A antibody (Monalizumab), which enhanced the anti-tumor activity of NK cells. The effects of CTLs was restored when it was combined with PD-1/PD-L1 blockade. Therefore, the NKG2A blockade can adequately compensate for the deficiency of PD-1/PD-L1 treatment by improving the effect of NK cells.Citation67
Other Tumor-Suppressing Microenvironments
In the tumor microenvironment, there are also some potentially drug-related components, including immunosuppressive cells such as regulatory T cells (Tregs), bone marrow-derived suppressor cells (MDSCs) and M2 macrophages and some other inhibitory enzymes and cytokines.
Tregs
Tregs inhibit the function of effector T cells (Teffs) by secreting certain inhibitory cytokines, including IL-10 and TGF-βCitation68 In vivo studies showed that removal of Tregs from the tumor microenvironment enhanced anti-tumor immunity. Besides, the efficacy of the PD-1/PD-L1 antibody was associated with an increased Teff to Tregs ratio.Citation69 The acquired resistance of PD-1 blockade alone or PD-1 blockade combined radiotherapy was associated with the increased Tregs infiltration. Furthermore, this resistance can be restored after the use of anti-CD25 treatment due to the depletion of Tregs.Citation70
MDSCs
Myeloid-derived suppressor cells (MDSCs) are a heterogeneous population of cells, including myeloid cell precursors, immature granulocytes, monocytes, and dendritic cells. Tumors can be seen as a long-term unhealed inflammatory process. There are many inflammation-related factors in the tumor microenvironment. Inflammatory factors IL-6, VEGF, proinflammatory factor COX2, PGE2, GM-CSF, M-SCF, and stem cell factor 1 (SCF-1) all can induce the expansion and activation of MDSCs. MDSCs not only inhibit the innate immune response by inhibiting the anti-tumor effect of NK cells and macrophages but also inhibit the adaptive immune response by blocking the activation of CD4+/CD8+ T cells and increasing the infiltration of Tregs.Citation71
Furthermore, the presence of MDSCs in the tumor microenvironment reduces the effectiveness of immunotherapy.Citation72,Citation73 A series of mouse models studies indicated that PI3K inhibitors could be used as potential therapeutic targets in combination with PD-1/PD-L1 blockade.Citation74 The STING/type I interferon pathway enhanced suppressive inflammation in tumors by recruiting myeloid cells in part via the CCR2 pathway. Knockout of CCR2 or treatment with anti-CCR2 blockade resulted in a blockade of radiation-induced MDSCs infiltration, thereby increasing the anti-tumor effect of radiotherapy combined with PD-1Citation75 treatment.
M2 Macrophages
Tumor-associated macrophages (TAMs) affect the efficacy of immunotherapy. TAMs include M1 macrophages promoting anti-tumor immunity and M2 macrophages suppressing anti-tumor immunity.Citation76 M2 is involved in the whole process of tumorigenesis, growth, invasion, and metastasis. In the early stage of tumor formation, M2 can promote tumor neovascularization and enhance the invasion ability of tumor cells. In the process of tumor metastasis, M2 promotes the formation of tumor “microenvironment before metastasis,” and promotes extravasation, survival and sustained the growth of tumor cells at the metastatic site. Besides, TAMs can also play a role in immunosuppressive function by releasing IL-10, TGF-β, and some other cytokines and protecting tumor cells from NK cells and T cells. In a mouse model of lung adenocarcinoma, reducing the recruitment of M2 macrophages inhibits the tumor growth, which may be related to CCL2 and CCR2 signal inactivation.Citation77 To overcome macrophage-associated potential drug resistance, blocking the colony-stimulating growth factor receptor (CSF-1R) in a mouse model of pancreatic cancer reduced the frequency of TAMs, increased IFN production, and enhanced the response of PD-1 treatment.Citation78 Rodell et al (2019) designed TLR7/8 agonist-loaded nanoparticles to promote the polarization of tumor-associated macrophages to M1 rather than M2 to enhance PD-1 blockade efficacy.Citation79
IDO
Indoleamine 2,3-dioxygenase 1 (IDO) produced by tumors or immune cells can increase the production and activity of Tregs and MDSCs. IDO can lower tryptophan levels, cause GCN2 activation and mTOR inhibition, inhibit the proliferation of effector T cells, and promote its apoptosis.Citation80 On the other hand, IDO up-regulates kynurenine, activates Aryl hydrocarbon receptor (AhR), promotes differentiation of Tregs, and ultimately suppresses immune responses.Citation81,Citation82 Studies showed that when B16 melanoma was treated with PD-1 blockade, the tumor growth of IDO knockout mice was significantly delayed, and the overall survival rate was improved compared with wild type mice.Citation83 Based on these studies, the combination of IDO inhibitors with PD-1/PD-L1 antibodies may be more effective than single agents.Citation84 However, on April 6, 2018, Incyte and Merck announced that an external Data Monitoring Committee (eDMC) review of the pivotal Phase 3 ECHO-301/KEYNOTE-252 study results evaluating Incyte’s epacadostat in combination with Merck’s KEYTRUDA® in patients with unresectable or metastatic melanoma determined that the study did not meet the primary endpoint of improving PFS in the overall population compared to KEYTRUDA monotherapy. The study’s second primary endpoint of OS also was not expected to reach statistical significance. Based on these results, and at the recommendation of the eDMC, the study was stopped.Citation85 This proves that immunotherapy has a long way to go. We should continue to explore other novel mechanisms that can potentially improve outcomes for patients in need.
Up-Regulation of CD73
CD39 is capable of hydrolyzing ATP to AMP and further processing AMP into immunosuppressive molecule adenosine by extracellular nucleosidase CD73. Adenosine can inhibit T cells proliferation and activity through the A2A receptor of T cells and can promote tumor metastasis through the A2B receptor of tumor cells.Citation86 Besides, the co-expression of CD39 and CD73 is considered a key mechanism for Tregs-mediated immunosuppression. In many types of tumors, the high expression of CD73 is accompanied by a poor prognosis and efficiency of PD-1 blockade.Citation87 Antisense oligonucleotide targeting CD39 can improve T cell anti-tumor immunity.Citation88
Expression of Specific Driver Genes
EGFR Mutation
NSCLC patients with an EGFR mutation have limited benefits.Citation89 Firstly, a patient with an EGFR mutation was found to have a low tumor mutation load (TMB). Patients with a dominant gene had lower overall TMB, resulting in unsatisfactory benefits for patients from immunotherapy. Secondly, PD-L1 expression levels were lower for EGFR+ NSCLC. The global multicenter EXPRESS study analyzed global data on PD-L1 expression in patients with late NSCLC and found that PD-L1 expression was lower in EGFR+ NSCLC patients than in wild-type patients.Citation90 Finally, the tumor microenvironment of EGFR+ NSCLC patients was mostly immune-desert or immune-excluded. A retrospective study found that patients with EGFR+ NSCLC had lower levels of PD-L1 expression and less CD8+ TILs infiltration.Citation91
KRAS Mutation
In tumor cells, KRAS mutation (KRAS*) can significantly inhibit the expression of interferon regulatory factor 2 (IRF2), thereby suppressing the inhibition of CXCL3 transcription by IRF2, namely up-regulating the expression and secretion of CXCL3. When CXCL3 binds to CXCR2, it can recruit MDSCs into TME and inhibit the effect of PD-1 treatment. Enhancing the expression of IRF2 or a combination of CXR2 inhibitors can relieve PD-1 resistance in CRC patients.Citation92
Previous studies showed that different subgroups of KRAS mutant LUAC could be distinguished based on whether there was STK11/LKB1 or TP53 co-mutation. STK11/LKB1-type lung adenocarcinoma exhibited shorter PFS in the KRAS mutant lung adenocarcinoma cohort. Skoulidis et al (2018) reported that STK11/LKB1 alteration was a major driver of primary resistance to PD-1 inhibitors in KRAS mutant lung adenocarcinoma patients. In the KRAS mutant mouse lung adenocarcinoma model, STK11/LKB1 deletion promoted PD-1/PD-L1 inhibitor resistance.Citation93
KIR Mutation
Killer cell immunoglobulin-like receptor (KIR) interacts with human leukocyte antigen (HLA) to affect NK cells’ reactivity and the total number of mature NK cells. Therefore, blocking KIR can induce anti-tumor effects by activating NK cells. Trefny et al (2019) identified a genetic variant of KIR (KIR3DS1) that was associated with primary resistance to PD-1 blockade in patients with NSCLC.Citation94
Conclusion
Based on the current limitations of PD-1/PD-L1 blockade, the combination of PD-1/PD-L1 blockade with other therapies has gradually become another significant development direction.Citation95 Radiotherapy can lead to the release of tumor antigens and increase the immunogenicity of tumors. Combining PD-1/PD-L1 blocking therapy with radiotherapy has been proposed.Citation96,Citation97 Vaccines, CD40 agonists, and TLR agonists can primarily promote cross-presentation of DCs to enhance T cell anti-tumor responses. CTLA-4 blockade, CD137 agonists, and inflammatory cytokines such as IL-2 and IL-12 can enhance priming and activation of T cells. Bevacizumab, an anti-VEGF antibody, can disrupt immunosuppressive states and neutralize inhibitory cytokines and enzymes such as IL-10, TGF-βCitation98 and IDO. Besides, by blocking TIM-3, Lag-3, and other inhibitory immune checkpoint molecules, it is also expected to enhance anti-tumor immunity by assisting PD-1 treatment.
Abbreviations
AhR: Aryl hydrocarbon receptor; APCs, Antigen-presenting cells; B2M, β2 microglobulin; CSF-1R, Colony-stimulating growth factor receptor; CTLs, Cytotoxic T lymphocytes; CTLA-4, Cytotoxic T-lymphocyte-associated antigen-4; DCs, Dendritic cells; DDR, DNA damage repair; EGFR, Epidermal growth factor receptor; ICBs, Immune-Checkpoint Blockers; IDO, Indolamine-2,3-dioxygenase; IFN-γ, Interferon-γ; IL-2, Interleukin-2; IL-8, Interleukin-8; IL-10, Interleukin-10; IL-12, Interleukin-12; IRF2, interferon regulatory factor 2; JAK, Janus Kinase; LAG-3, Lymphocyte activating gene 3; KIR, Killer cell immunoglobulin-like receptor; MAPK, Mitogen-activated protein kinase; MDSC, Marrow-derived inhibitory cells; MHC, Major histocompatibility complex; MMR, Mismatch repair; MSS, microsatellite stabilization; NSCLC, Non-small cell lung cancer; OS, Overall survival; PD-1, Programmed death-1; PD-L1, Programmed cell death 1 ligand 1; PFS, Progression-free survival; PTEN, Phosphatase and tensin homolog deleted on chromosome ten; SCF-1, stem cell factor 1; STAT, Signal transducers and activators of transcription; TAMs, Tumor-associated macrophages; Tcm, memory T cell; TCR, T cell receptors; Teff, effector T cells; TIL, Tumor infiltrating lymphocyte; TIM-3, T cell immunoglobulin mucin-3; TMB, Tumor mutation burden; TME, Tumor microenvironment; VEGF, Vascular endothelial growth factor.
Statement of Human and Animal Rights
No experiments have been performed, including patients or animals.
Statement of Informed Consent
Patients have not been involved in the study.
Author Contributions
All authors contributed to data analysis, drafting or revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Acknowledgments
The Chinese Society of Clinical Oncology supported this work.
Funding
This study was supported by funding from the CSCO-QiLu Oncology Research Fund (No. Y-G201801-055).
Disclosure
The authors declare that they have no conflicts of interest.
References
- Rotte A, D’Orazi G, Bhandaru M. Nobel committee honors tumor immunologists. J Exp Clin Cancer Res. 2018;37(1):262. doi:10.1186/s13046-018-0937-630376854
- Wang S, Hao J, Wang H, Fang Y, Tan L. Efficacy and safety of immune checkpoint inhibitors in non-small cell lung cancer. Oncoimmunology. 2018;7(8):e1457600. doi:10.1080/2162402X.2018.145760030221052
- Bellmunt J, Powles T, Vogelzang NJ. A review on the evolution of PD-1/PD-L1 immunotherapy for bladder cancer: the future is now. Cancer Treat Rev. 2017;54:58–67. doi:10.1016/j.ctrv.2017.01.00728214651
- Suzman DL, Agrawal S, Ning YM, et al. FDA approval summary: atezolizumab or pembrolizumab for the treatment of patients with advanced urothelial carcinoma ineligible for cisplatin-containing chemotherapy. Oncologist. 2019;24(4):563–569. doi:10.1634/theoncologist.2018-008430541754
- Ansell SM, Lesokhin AM, Borrello I, et al. PD-1 blockade with nivolumab in relapsed or refractory Hodgkin’s lymphoma. N Engl J Med. 2015;372(4):311–319. doi:10.1056/NEJMoa141108725482239
- Kasamon YL, de Claro RA, Wang Y, Shen YL, Farrell AT, Pazdur R. FDA approval summary: nivolumab for the treatment of relapsed or progressive classical hodgkin lymphoma. Oncologist. 2017;22(5):585–591. doi:10.1634/theoncologist.2017-000428438889
- Nghiem PT, Bhatia S, Lipson EJ, et al. PD-1 blockade with pembrolizumab in advanced merkel-cell carcinoma. N Engl J Med. 2016;374(26):2542–2552. doi:10.1056/NEJMoa160370227093365
- Mitteldorf C, Berisha A, Tronnier M, Pfaltz MC, Kempf W. PD-1 and PD-L1 in neoplastic cells and the tumor microenvironment of merkel cell carcinoma. J Cutan Pathol. 2017;44(9):740–746. doi:10.1111/cup.2017.44.issue-928569410
- Chen DS, Mellman I. Oncology meets immunology: the cancer-immunity cycle. Immunity. 2013;39(1):1–10. doi:10.1016/j.immuni.2013.07.01223890059
- Chalmers ZR, Connelly CF, Fabrizio D, et al. Analysis of 100,000 human cancer genomes reveals the landscape of tumor mutational burden. Genome Med. 2017;9(1):34. doi:10.1186/s13073-017-0424-228420421
- Goodman AM, Kato S, Bazhenova L, et al. Tumor mutational burden as an independent predictor of response to immunotherapy in diverse cancers. Mol Cancer Ther. 2017;16(11):2598–2608. doi:10.1158/1535-7163.MCT-17-038628835386
- Riaz N, Morris L, Havel JJ, Makarov V, Desrichard A, Chan TA. The role of neoantigens in response to immune checkpoint blockade. Int Immunol. 2016;28(8):411–419. doi:10.1093/intimm/dxw01927048318
- Furness AJ, Quezada SA, Peggs KS. Neoantigen heterogeneity: a key driver of immune response and sensitivity to immune checkpoint blockade? Immunotherapy. 2016;8(7):763–766. doi:10.2217/imt-2016-006427349975
- Schumacher TN, Schreiber RD. Neoantigens in cancer immunotherapy. Science. 2015;348(6230):69–74. doi:10.1126/science.aaa497125838375
- Le DT, Durham JN, Smith KN, et al. Mismatch repair deficiency predicts response of solid tumors to PD-1 blockade. Science. 2017;357(6349):409–413. doi:10.1126/science.aan673328596308
- AACR Project GENIE. Powering precision medicine through an international consortium. Cancer Discov. 2017;7(8):818–831. doi:10.1158/2159-8290.CD-17-015128572459
- Sclafani F. PD-1 inhibition in metastatic dMMR/MSI-H colorectal cancer. Lancet Oncol. 2017;18(9):1141–1142. doi:10.1016/S1470-2045(17)30512-028734760
- Xu A, Zhang L, Yuan J, et al. TLR9 agonist enhances radiofrequency ablation-induced CTL responses, leading to the potent inhibition of primary tumor growth and lung metastasis. Cell Mol Immunol. 2018;16(10):820–832.30467420
- Ye F, Chen HZ, Xie X, Ye DF. [Effects of vascular endothelial growth factor on differentiation and function of dendritic cells generated from CD34+ hematopoietic progenitor cells in vitro]. Zhonghua Xue Ye Xue Za Zhi. 2004;25(9):532–535.15569531
- Wolfle SJ, Strebovsky J, Bartz H, et al. PD-L1 expression on tolerogenic APCs is controlled by STAT-3. Eur J Immunol. 2011;41(2):413–424. doi:10.1002/eji.20104097921268011
- Koga MM, Bizzarro B, Sa-Nunes A, Rios FJ, Jancar S. Activation of PAF-receptor induces regulatory dendritic cells through PGE2 and IL-10. Prostaglandins Leukot Essent Fatty Acids. 2013;89(5):319–326. doi:10.1016/j.plefa.2013.09.00324120121
- Youlin K, Weiyang H, Simin L, Xin G. Prostaglandin E2 inhibits prostate cancer progression by countervailing tumor microenvironment-induced impairment of dendritic cell migration through LXRalpha/CCR7 pathway. J Immunol Res. 2018;2018:5808962. doi:10.1155/2018/580896229850633
- Routy B, Le Chatelier E, Derosa L, et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science. 2018;359(6371):91–97. doi:10.1126/science.aan370629097494
- Burki TK. Gut microbiome and immunotherapy response. Lancet Oncol. 2017;18(12):e717. doi:10.1016/S1470-2045(17)30841-029129446
- Ruiz L, Delgado S, Ruas-Madiedo P, Sanchez B, Margolles A. Bifidobacteria and their molecular communication with the immune system. Front Microbiol. 2017;8:2345. doi:10.3389/fmicb.2017.0234529255450
- Sivan A, Corrales L, Hubert N, et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science. 2015;350(6264):1084–1089. doi:10.1126/science.aac425526541606
- Gopalakrishnan V, Spencer CN, Nezi L, et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science. 2018;359(6371):97–103.29097493
- Matson V, Fessler J, Bao R, et al. The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science. 2018;359(6371):104–108. doi:10.1126/science.aao329029302014
- Tokunaga R, Zhang W, Naseem M, et al. CXCL9, CXCL10, CXCL11/CXCR3 axis for immune activation - A target for novel cancer therapy. Cancer Treat Rev. 2018;63:40–47. doi:10.1016/j.ctrv.2017.11.00729207310
- Herbst RS, Soria JC, Kowanetz M, et al. Predictive correlates of response to the anti-PD-L1 antibody MPDL3280A in cancer patients. Nature. 2014;515(7528):563–567. doi:10.1038/nature1401125428504
- Hu J, Sun C, Bernatchez C, et al. T-cell homing therapy for reducing regulatory T cells and preserving effector T-cell function in large solid tumors. Clin Cancer Res. 2018;24(12):2920–2934. doi:10.1158/1078-0432.CCR-17-136529391351
- Peng D, Kryczek I, Nagarsheth N, et al. Epigenetic silencing of TH1-type chemokines shapes tumour immunity and immunotherapy. Nature. 2015;527(7577):249–253. doi:10.1038/nature1552026503055
- Peng W, Chen JQ, Liu C, et al. Loss of PTEN promotes resistance to T cell-mediated immunotherapy. Cancer Discov. 2016;6(2):202–216. doi:10.1158/2159-8290.CD-15-028326645196
- Garcia AJ, Ruscetti M, Arenzana TL, et al. Pten null prostate epithelium promotes localized myeloid-derived suppressor cell expansion and immune suppression during tumor initiation and progression. Mol Cell Biol. 2014;34(11):2017–2028. doi:10.1128/MCB.00090-1424662052
- Lorin S, Hamai A, Mehrpour M, Codogno P. Autophagy regulation and its role in cancer. Semin Cancer Biol. 2013;23(5):361–379. doi:10.1016/j.semcancer.2013.06.00723811268
- Dhillon AS, Hagan S, Rath O, Kolch W. MAP kinase signalling pathways in cancer. Oncogene. 2007;26(22):3279–3290. doi:10.1038/sj.onc.121042117496922
- Whipple CA, Boni A, Fisher JL, et al. The mitogen-activated protein kinase pathway plays a critical role in regulating immunological properties of BRAF mutant cutaneous melanoma cells. Melanoma Res. 2016;26(3):223–235. doi:10.1097/CMR.000000000000024426974965
- Hugo W, Zaretsky JM, Sun L, et al. Genomic and transcriptomic features of response to anti-PD-1 therapy in metastatic melanoma. Cell. 2016;165(1):35–44. doi:10.1016/j.cell.2016.02.06526997480
- Deken MA, Gadiot J, Jordanova ES, et al. Targeting the MAPK and PI3K pathways in combination with PD1 blockade in melanoma. Oncoimmunology. 2016;5(12):e1238557. doi:10.1080/2162402X.2016.123855728123875
- Gettinger S, Choi J, Hastings K, et al. Impaired HLA class I antigen processing and presentation as a mechanism of acquired resistance to immune checkpoint inhibitors in lung cancer. Cancer Discov. 2017;7(12):1420–1435. doi:10.1158/2159-8290.CD-17-059329025772
- Zaretsky JM, Garcia-Diaz A, Shin DS, et al. mutations associated with acquired resistance to PD-1 blockade in melanoma. N Engl J Med. 2016;375(9):819–829. doi:10.1056/NEJMoa160495827433843
- Sucker A, Zhao F, Real B, et al. Genetic evolution of T-cell resistance in the course of melanoma progression. Clin Cancer Res. 2014;20(24):6593–6604. doi:10.1158/1078-0432.CCR-14-056725294904
- Shin DS, Zaretsky JM, Escuin-Ordinas H, et al. Primary resistance to PD-1 blockade mediated by JAK1/2 mutations. Cancer Discov. 2017;7(2):188–201. doi:10.1158/2159-8290.CD-16-122327903500
- Sucker A, Zhao F, Pieper N, et al. Acquired IFNgamma resistance impairs anti-tumor immunity and gives rise to T-cell-resistant melanoma lesions. Nat Commun. 2017;8:15440. doi:10.1038/ncomms1544028561041
- Chen DS, Mellman I. Elements of cancer immunity and the cancer-immune set point. Nature. 2017;541(7637):321–330. doi:10.1038/nature2134928102259
- Bocanegra A, Fernandez-Hinojal G, Zuazo-Ibarra M, et al. PD-L1 expression in systemic immune cell populations as a potential predictive biomarker of responses to PD-L1/PD-1 blockade therapy in lung cancer. Int J Mol Sci. 2019;20:7. doi:10.3390/ijms20071631
- Rizk EM, Gartrell RD, Barker LW, et al. Prognostic and predictive immunohistochemistry-based biomarkers in cancer and immunotherapy. Hematol Oncol Clin North Am. 2019;33(2):291–299. doi:10.1016/j.hoc.2018.12.00530833001
- Naidoo J, Iyriboz T, Cunningham J, Hellmann MD. Reply to M. Nishino et al. J Clin Oncol. 2017;35(14):1629–1630. doi:10.1200/JCO.2016.71.663928165907
- Teng MW, Ngiow SF, Ribas A, Smyth MJ. Classifying cancers based on T-cell infiltration and PD-L1. Cancer Res. 2015;75(11):2139–2145. doi:10.1158/0008-5472.CAN-15-025525977340
- Ngiow SF, Young A, Jacquelot N, et al. A threshold level of intratumor CD8+ T-cell PD1 expression dictates therapeutic response to anti-PD1. Cancer Res. 2015;75(18):3800–3811. doi:10.1158/0008-5472.CAN-15-108226208901
- Buchbinder EI, Desai A. CTLA-4 and PD-1 pathways: similarities, differences, and implications of their inhibition. Am J Clin Oncol. 2016;39(1):98–106. doi:10.1097/COC.000000000000023926558876
- Son CH, Bae J, Lee HR, Yang K, Park YS. Enhancement of antitumor immunity by combination of anti-CTLA-4 antibody and radioimmunotherapy through the suppression of Tregs. Oncol Lett. 2017;13(5):3781–3786. doi:10.3892/ol.2017.593328521478
- Beavis PA, Henderson MA, Giuffrida L, et al. Dual PD-1 and CTLA-4 checkpoint blockade promotes antitumor immune responses through CD4(+)Foxp3(-) cell-mediated modulation of CD103(+) dendritic cells. Cancer Immunol Res. 2018;6(9):1069–1081. doi:10.1158/2326-6066.CIR-18-029130018045
- Swanson MS, Sinha UK. Rationale for combined blockade of PD-1 and CTLA-4 in advanced head and neck squamous cell cancer-review of current data. Oral Oncol. 2015;51(1):12–15. doi:10.1016/j.oraloncology.2014.10.01025459157
- Tanvetyanon T, Gray JE, Antonia SJ. PD-1 checkpoint blockade alone or combined PD-1 and CTLA-4 blockade as immunotherapy for lung cancer? Expert Opin Biol Ther. 2017;17(3):305–312. doi:10.1080/14712598.2017.128045428064556
- Grosso JF, Kelleher CC, Harris TJ, et al. LAG-3 regulates CD8+ T cell accumulation and effector function in murine self- and tumor-tolerance systems. J Clin Invest. 2007;117(11):3383–3392. doi:10.1172/JCI3118417932562
- Huang CT, Workman CJ, Flies D, et al. Role of LAG-3 in regulatory T cells. Immunity. 2004;21(4):503–513. doi:10.1016/j.immuni.2004.08.01015485628
- Goding SR, Wilson KA, Xie Y, et al. Restoring immune function of tumor-specific CD4+ T cells during recurrence of melanoma. J Immunol. 2013;190(9):4899–4909. doi:10.4049/jimmunol.130027123536636
- Goding SR, Wilson KA, Rosinsky C, Antony PA. PD-L1-independent mechanisms control the resistance of melanoma to CD4(+) T cell adoptive immunotherapy. J Immunol. 2018;200(9):3304–3311. doi:10.4049/jimmunol.170161729602773
- He Y, Yu H, Rozeboom L, et al. LAG-3 protein expression in non-small cell lung cancer and its relationship with PD-1/PD-L1 and tumor-infiltrating lymphocytes. J Thorac Oncol. 2017;12(5):814–823. doi:10.1016/j.jtho.2017.01.01928132868
- Lee WJ, Lee YJ, Choi ME, et al. LAG-3 and TIGIT protein expressions in cutaneous melanoma and their relationship with PD-1 tumor-infiltrating lymphocytes. J Am Acad Dermatol. 2019;70(2):394.
- Burugu S, Gao D, Leung S, Chia SK, Nielsen TO. LAG-3+ tumor infiltrating lymphocytes in breast cancer: clinical correlates and association with PD-1/PD-L1+ tumors. Ann Oncol. 2017;28(12):2977–2984. doi:10.1093/annonc/mdx55729045526
- Sakuishi K, Apetoh L, Sullivan JM, Blazar BR, Kuchroo VK, Anderson AC. Targeting Tim-3 and PD-1 pathways to reverse T cell exhaustion and restore anti-tumor immunity. J Exp Med. 2010;207(10):2187–2194. doi:10.1084/jem.2010064320819927
- Koyama S, Akbay EA, Li YY, et al. Adaptive resistance to therapeutic PD-1 blockade is associated with upregulation of alternative immune checkpoints. Nat Commun. 2016;7:10501. doi:10.1038/ncomms1050126883990
- Limagne E, Richard C, Thibaudin M, et al. Tim-3/galectin-9 pathway and mMDSC control primary and secondary resistances to PD-1 blockade in lung cancer patients. Oncoimmunology. 2019;8(4):e1564505. doi:10.1080/2162402X.2018.156450530906658
- Andre P, Denis C, Soulas C, et al. Anti-NKG2A mAb is a checkpoint inhibitor that promotes anti-tumor immunity by unleashing both T and NK cells. Cell. 2018;175(7):1731–1743.e1713. doi:10.1016/j.cell.2018.10.01430503213
- Mingari MC, Pietra G, Moretta L. Immune checkpoint inhibitors: anti-NKG2A antibodies on board. Trends Immunol. 2019;40(2):83–85. doi:10.1016/j.it.2018.12.00930609967
- Li M, Eckl J, Geiger C, Schendel DJ, Pohla H. A novel and effective method to generate human porcine-specific regulatory T cells with high expression of IL-10, TGF-beta1 and IL-35. Sci Rep. 2017;7(1):3974. doi:10.1038/s41598-017-04322-328638110
- Arce Vargas F, Furness AJS, Solomon I, et al. Fc-optimized anti-CD25 depletes tumor-infiltrating regulatory T cells and synergizes with PD-1 blockade to eradicate established tumors. Immunity. 2017;46(4):577–586. doi:10.1016/j.immuni.2017.03.01328410988
- Oweida A, Hararah MK, Phan A, et al. Resistance to radiotherapy and PD-L1 blockade is mediated by TIM-3 upregulation and regulatory T-cell infiltration. Clin Cancer Res. 2018;24(21):5368–5380. doi:10.1158/1078-0432.CCR-18-103830042205
- Lu LC, Chang CJ, Hsu CH. Targeting myeloid-derived suppressor cells in the treatment of hepatocellular carcinoma: current state and future perspectives. J Hepatocell Carcinoma. 2019;6:71–84. doi:10.2147/JHC.S15969331123667
- Srivastava MK, Andersson A, Zhu L, et al. Myeloid suppressor cells and immune modulation in lung cancer. Immunotherapy. 2012;4(3):291–304. doi:10.2217/imt.11.17822401635
- Meyer C, Cagnon L, Costa-Nunes CM, et al. Frequencies of circulating MDSC correlate with clinical outcome of melanoma patients treated with ipilimumab. Cancer Immunol Immunother. 2014;63(3):247–257.24357148
- Kim K, Skora AD, Li Z, et al. Eradication of metastatic mouse cancers resistant to immune checkpoint blockade by suppression of myeloid-derived cells. Proc Natl Acad Sci U S A. 2014;111(32):11774–11779. doi:10.1073/pnas.141062611125071169
- Liang H, Deng L, Hou Y, et al. Host STING-dependent MDSC mobilization drives extrinsic radiation resistance. Nat Commun. 2017;8(1):1736. doi:10.1038/s41467-017-01566-529170400
- Santoni M, Romagnoli E, Saladino T, et al. Triple negative breast cancer: key role of tumor-associated macrophages in regulating the activity of anti-PD-1/PD-L1 agents. Biochim Biophys Acta Rev Cancer. 2018;1869(1):78–84. doi:10.1016/j.bbcan.2017.10.00729126881
- Lehmann B, Biburger M, Bruckner C, et al. Tumor location determines tissue-specific recruitment of tumor-associated macrophages and antibody-dependent immunotherapy response. Sci Immunol. 2017;2:7. doi:10.1126/sciimmunol.aah6413
- Dammeijer F, Lievense LA, Kaijen-Lambers ME, et al. Depletion of tumor-associated macrophages with a CSF-1R kinase inhibitor enhances antitumor immunity and survival induced by DC immunotherapy. Cancer Immunol Res. 2017;5(7):535–546. doi:10.1158/2326-6066.CIR-16-030928536100
- Rodell CB, Arlauckas SP, Cuccarese MF, et al. TLR7/8-agonist-loaded nanoparticles promote the polarization of tumour-associated macrophages to enhance cancer immunotherapy. Nat Biomed Eng. 2018;2:578–588. doi:10.1038/s41551-018-0236-8
- Brochez L, Chevolet I, Kruse V. The rationale of indoleamine 2,3-dioxygenase inhibition for cancer therapy. Eur J Cancer. 2017;76:167–182. doi:10.1016/j.ejca.2017.01.01128324751
- Labadie BW, Bao R, Luke JJ. Reimagining IDO pathway inhibition in cancer immunotherapy via downstream focus on the tryptophan-kynurenine-aryl hydrocarbon axis. Clin Cancer Res. 2019;25(5):1462–1471. doi:10.1158/1078-0432.CCR-18-288230377198
- Eleftheriadis T, Pissas G, Liakopoulos V, Stefanidis I. IDO decreases glycolysis and glutaminolysis by activating GCN2K, while it increases fatty acid oxidation by activating AhR, thus preserving CD4+ Tcell survival and proliferation. Int J Mol Med. 2018;42(1):557–568. doi:10.3892/ijmm.2018.362429693118
- Holmgaard RB, Zamarin D, Munn DH, Wolchok JD, Allison JP. Indoleamine 2,3-dioxygenase is a critical resistance mechanism in antitumor T cell immunotherapy targeting CTLA-4. J Exp Med. 2013;210(7):1389–1402. doi:10.1084/jem.2013006623752227
- Khair DO, Bax HJ, Mele S, et al. Combining immune checkpoint inhibitors: established and emerging targets and strategies to improve outcomes in melanoma. Front Immunol. 2019;10:453. doi:10.3389/fimmu.2019.0045330941125
- Long GV, Dummer R, Hamid O, et al. Epacadostat plus pembrolizumab versus placebo plus pembrolizumab in patients with unresectable or metastatic melanoma (ECHO-301/KEYNOTE-252): a phase 3, randomised, double-blind study. Lancet Oncol. 2019;20(8):1083–1097. doi:10.1016/S1470-2045(19)30274-831221619
- Allard B, Beavis PA, Darcy PK, Stagg J. Immunosuppressive activities of adenosine in cancer. Curr Opin Pharmacol. 2016;29:7–16. doi:10.1016/j.coph.2016.04.00127209048
- Deng WW, Li YC, Ma SR, et al. Specific blockade CD73 alters the “exhausted” phenotype of T cells in head and neck squamous cell carcinoma. Int J Cancer. 2018;143(6):1494–1504. doi:10.1002/ijc.3153429663369
- Kashyap AS, Thelemann T, Klar R, et al. Antisense oligonucleotide targeting CD39 improves anti-tumor T cell immunity. J Immunother Cancer. 2019;7(1):67. doi:10.1186/s40425-019-0545-930871609
- Gainor JF, Shaw AT, Sequist LV, et al. EGFR mutations and ALK rearrangements are associated with low response rates to PD-1 pathway blockade in non-small cell lung cancer: a retrospective analysis. Clin Cancer Res. 2016;22(18):4585–4593. doi:10.1158/1078-0432.CCR-15-310127225694
- Ji M, Liu Y, Li Q, et al. PD-1/PD-L1 expression in non-small-cell lung cancer and its correlation with EGFR/KRAS mutations. Cancer Biol Ther. 2016;17(4):407–413. doi:10.1080/15384047.2016.115625626954523
- Dong ZY, Zhang JT, Liu SY, et al. EGFR mutation correlates with uninflamed phenotype and weak immunogenicity, causing impaired response to PD-1 blockade in non-small cell lung cancer. Oncoimmunology. 2017;6(11):e1356145. doi:10.1080/2162402X.2017.135614529147605
- Liao W, Overman MJ, Boutin AT, et al. KRAS-IRF2 axis drives immune suppression and immune therapy resistance in colorectal cancer. Cancer Cell. 2019;35(4):559–572.e557. doi:10.1016/j.ccell.2019.02.00830905761
- Skoulidis F, Goldberg ME, Greenawalt DM, et al. STK11/LKB1 mutations and PD-1 inhibitor resistance in KRAS-mutant lung adenocarcinoma. Cancer Discov. 2018;8(7):822–835. doi:10.1158/2159-8290.CD-18-009929773717
- Trefny MP, Rothschild SI, Uhlenbrock F, et al. A variant of a killer cell immunoglobulin-like receptor is associated with resistance to PD-1 blockade in lung cancer. Clin Cancer Res. 2019. doi:10.1158/1078-0432.CCR-18-3041
- Fan L, Li Y, Chen JY, Zheng YF, Xu XM. Immune checkpoint modulators in cancer immunotherapy: recent advances and combination rationales. Cancer Lett. 2019. doi:10.1016/j.canlet.2019.03.050
- Twyman-Saint Victor C, Rech AJ, Maity A, et al. Radiation and dual checkpoint blockade activate non-redundant immune mechanisms in cancer. Nature. 2015;520(7547):373–377. doi:10.1038/nature1429225754329
- Takamori S, Toyokawa G, Takada K, Shoji F, Okamoto T, Maehara Y. Combination therapy of radiotherapy and anti-PD-1/PD-L1 treatment in non-small-cell lung cancer: a mini-review. Clin Lung Cancer. 2018;19(1):12–16. doi:10.1016/j.cllc.2017.06.01528739315
- Mariathasan S, Turley SJ, Nickles D, et al. TGFbeta attenuates tumour response to PD-L1 blockade by contributing to exclusion of T cells. Nature. 2018;554(7693):544–548. doi:10.1038/nature2550129443960