
Abstract
Background
Checkpoint kinase 1 (CHK1) plays an important role in DNA damage response and cell cycle progression. Thus, targeting CHK1 is an efficient strategy for cancer therapy.
Purpose
The present study aimed to investigate the potential therapeutic effects of GDC-0575, a CHK1-specific inhibitor, in colitis-associated cancer (CAC) and colitis.
Methods
We established a DSS-induced acute colitis model and an azoxymethane/dextran sodium sulfate (DSS)-induced CAC model using mice and tested the effect of GDC-0575 on them. Flow cytometry and immunofluorescence were employed to investigate the infiltration of immune cells, and inflammatory cytokine expression in the colon of mice with CAC or colitis was investigated using ELISA and qPCR. We also investigated the correlation between CHK1 and CCL2/CCR2 in human colorectal cancer (CRC) tissues.
Results
Administration of GDC-0575 significantly inhibited CHK1 expression in the colon and dramatically impaired the development of CAC and colitis in mice. Moreover, the inhibition of CHK1 expression resulted in efficient inhibition of infiltration by iNOS-positive macrophages, but had no significant effect on CD4 T cells, CD8 T cells, and myeloid-derived suppressor cells (MDSCs). Significant downregulation of TNF-α, IL-6, and IL-1β and dramatic upregulation of IL-10 were observed in the colons of both mice with CAC and colitis treated with GDC-0575. CCL2 expression was also downregulated by GDC-0575 in both mice with CAC and colitis; this was followed by the inhibition of CCR2+ macrophage infiltration in the colon. Furthermore, we report a positive correlation between CHK1 expression and CCL2/CCR2 expression in the malignant tissues of patients with CRC.
Conclusion
Taken together, we infer that GDC-0575 impairs the development of CAC and colitis by regulating cytokine expression and inhibiting CCR2+ macrophage infiltration in mice colon.
Introduction
Colorectal cancer (CRC), one of the most prevalent cancers, is a leading cause of cancer-related deaths worldwide.Citation1,Citation2 CRC development is influenced by multiple risk factors, such as age, genetics, and the environment, and is often associated with an inflammatory response.Citation3–Citation5 Chronic inflammatory diseases of the intestinal tract, such as ulcerative colitis (UC) and Crohn’s disease, significantly increase the risk of developing CRC.Citation6–Citation8 The activation of STAT3 and NF-κB signaling, release of cytokines, and immune modulation have been demonstrated to be involved in colitis-induced CRC progression by increasing the risk of gene mutations and epigenetic changes.Citation9–Citation11 It is seen that 18% of the patients with UC develop CRC.Citation12
Initiation, progression, and resolution, and restoration of tissue integrity are the different stages of inflammation.Citation13,Citation14 Inflammatory monocytes and macrophages play a regulatory role in the initiation and resolution stages.Citation13,Citation14 In response to inflammatory mediators, macrophages express inducible nitric oxide synthase (iNOS) and release high levels of TNF-α, IL-6, IL-1β, and other pro-inflammatory cytokines.Citation15,Citation16 During the development of colitis and CAC, macrophages are activated and recruited by the DSS-induced inflammatory environment and proinflammatory cytokines, and release inflammatory cytokines that aggravate this development.Citation17,Citation18 Therefore, targeting macrophages and inflammatory cytokines is a potential therapeutic strategy for colitis and CAC.Citation19
Checkpoint kinase 1 (CHK1) functions as a Ser/Thr protein kinase that regulates cell cycle in response to DNA damage.Citation20 Various studies have demonstrated the multifunctional role played by CHK1 in regulating replication origin firing, stabilizing stalled replication forks, and creating a time window for resolving DNA lesions.Citation21 Owing to the role of CHK in cancer progression, CHK1 inhibitors, like GDC-0575 and prexasertib, have been used as monotherapies and in combination with chemotherapy for multiple preclinical models of solid tumors.Citation22–Citation24 To date, more than 15 trials on various CHK1 inhibitors, primarily in combination with gemcitabine, have been investigated.Citation20,Citation25 Combination of GDC-0575 with G-CSF overrides cytarabine resistance in human acute myeloid leukemia.Citation26
The present study aimed to investigate the potential function of CHK1 in CAC and colitis in mice. GDC-0575, a inhibitor of CHK1, was used to treat mice with CAC or colitis. Immune cell infiltration and inflammatory cytokine expression in response to treatment of the mouse models with GDC-0575 were measured using flow cytometry, immunofluorescence, and ELISA. The correlation between CHK1 expression and CCL2/CCR2 expression was investigated in CRC tissues. Our results serve as evidence to clarify the function and mechanism of CHK1 in the context of CAC and colitis development and present a potential therapeutic target for CAC.
Materials and Methods
Animal Study
C57 mice (7–8 weeks old, 20 g, male) were purchased from HFKbio (Beijing, China) and housed under specific pathogen-free conditions with free access to sterile, distilled water, and food. A colitis-associated cancer model was established as described in previous studies.Citation27,Citation28 Azoxymethane (AOM, Sigma, MA, USA) was intraperitoneally injected once (10 mg/kg body weight), followed by three cycles of dextran sulfate sodium (2% DSS prepared in drinking water for one week, and normal drinking water for 2 weeks in one cycle). At 15, 17, 19, and 21 days post AOM injection, GDC-0575 was administered orally at a final concentration of 7.5 mg/kg. The control group was administered DMSO. At 10 weeks post AOM injection, the mice were sacrificed, and the colons were collected. Macroscopic tumors in each colon were counted and measured using digital calipers. In case of the acute colitis model, 3% DSS dissolved in drinking water was used to feed the mice for 7 days, followed by 7 days of normal drinking-water feeding. GDC-0575 was orally administered on days 8 and 10 after 3% DSS treatment. The body weight and survival of the mice were recorded daily. Stool consistency and occult blood were scored as described previously.Citation29 Bleeding scores: 3 = gross rectal bleeding, 2 = blood traces in stool visible, 1 = positive hemoccult, 0 = no blood hemoccult. Stool scores: 3 = liquid stools that adhered to the anus, 2 = semi-formed stools that adhered to the anus, 1 = semi-formed stools that did not adhere to the anus, 0 = well-formed pellets. Ten mice were included in each group. All the animal experiments were approved by the Ethics Committee of Jishou University following the guidelines of Laboratory Animal Welfare and Ethics Committee of Jishou University.
Histological Staining
Whole colon tissues were fixed with 4% paraformaldehyde at room temperature (22–25 °C) for 48 hours. After dehydration, the colon tissues were embedded in paraffin. Histological examination was performed in accordance with the manufacturer’s instructions in the H&E staining kit (Beyotime, Beijing, China). Colitis gradation was scored according to a previous report ().Citation29
Table 1 Colitis Gradation Was Scored According to a Previous ReportCitation29
Immunohistochemistry (IHC) and Immunofluorescence
The IHC and immunofluorescence were performed on 4-micrometer sections of paraffin-embedded tissues. After deparaffinization and rehydration in water, the samples were subjected to antigen retrieval using euro neurite repair solution under high temperature/high pressure for 3 min. After blocking with goat serum for 15 min at room temperature, the sections were incubated with antibodies against CHK1 (Origene Technologies), CCL2 (Abcam), CCR2 (Abcam), F4/80 (Proteintech), and iNOS (Proteintech) at 4 °C overnight. For IHC, a detection system from Zsbio (SP9001, SP9002, Beijing, China) was used as per the manufacturer’s instructions. Positive cells were detected using a DAB kit (Maixin, Fuzhou, China). For immunofluorescence, sections were incubated with fluorescein isothiocyanate-conjugated secondary antibody and Texas red-conjugated secondary antibody at 37 °C for 1 hour. The nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI; Beyotime, Beijing). The positive cells and total cells in each frame were counted to calculate the percentage of positive cells. All images were acquired on an Olympus BX51 microscope (Tokyo, Japan). The positive cells and total cells in each frame were counted and the percent of positive cells were calculated by positive cells/total cells.
Quantitative PCR
Total RNA was extracted from whole colon tissues using TRIzol (Invitrogen, CA, USA), following as per the manufacturer’s instructions. RNA purity and concentration were monitored using a NanoDrop ND-2000 spectrophotometer (Thermo Fisher Scientific, CA, USA). One microgram of total RNA was used as template to synthesize cDNA using the SuperScript III Reverse transcriptase assay (Invitrogen, CA, USA). TaqMan Gene Expression Master Mix (Thermo Fisher Scientific, CA, USA) was used for real-time qPCRs with a StepOne Real-Time PCR System (Applied Biosystems, CA, USA). GAPDH was used as the loading control. The results of qPCR were analyzed using the conventional ΔΔCt method.
Flow Cytometry
Colonic lamina propria immunocytes were isolated as previously described.Citation30 Briefly, the whole colon was collected and washed three times with ice-cold PBS (Invitrogen, CA, USA). After cutting into small pieces, the colons were digested in 20 mL RPMI 1640 containing 5% fetal calf serum, 5 mM ethylenediaminetetraacetic acid (EDTA; pH 8.0, Sigma, MA, USA), and 1 mM dithiothreitol (DTT, Sigma, MA, USA)) at 37 °C for 25 min. The remaining tissues were digested in 20 mL RPMI 1640 containing 5% fetal calf serum, 0.2% type IV collagenase (Invitrogen, CA, USA), and 0.025% DNase I (Invitrogen)) at 37 °C for 50 min. The supernatant was collected and resuspended in ice-cold PBS containing 3% fetal bovine serum (FBS; Gibco) and 0.2% EDTA. Then, blocking was done with Fc-block (BD Biosciences, USA) and stained with the following antibodies: CD45 (cat. no. 103,116), F4/80 (cat. no. 123,108), CD11b (cat. no. 101,228), Ly6G (cat. no. 127,623), which were obtained from BioLegend, CD4 (cat. no. 553,030), and CD8 (cat. no. 100,722), obtained from BD Bioscience. Data were acquired and analyzed using a NovoCyte flow cytometer (ACEA Biosciences, USA).
Enzyme-Linked Immunosorbent Assay
The colons were collected and washed three times with ice-cold PBS (Invitrogen, CA, USA). After cutting into pieces, the colons were lysed with ice-cold lysis buffer (RIPA lysis buffer with 1% protease inhibitor cocktail (Beyotime, Beijing, China). After lysing for 30 min on ice and centrifuging at 12,000 g (4 °C) for 15 min, the supernatant was collected and subjected to the BCA assay for protein concentration determination (Beyotime, Beijing, China). The expression of TNF-α, CCL-2, IL-10, IL-1β, cyclooxygenase-2 (COX-2) and IL-6 was determined using an ELISA kit (NeoBioscience, Shenzhen, China) according to the manufacturer’s instructions.
Statistical Analysis
All statistical analyses were performed using GraphPad Prism 7.0. Results are expressed as mean ± standard error (mean ± SE). Student’s t-test was used to calculate the P values for the comparison of the two groups. Log-rank (Mantel–Cox) test was used to calculate the P values by comparing the mouse survival curves. A level of P<0.05 was considered to be significant.
Results
GDC-0575 Impairs the Development of Colitis-Associated Cancer in Mice
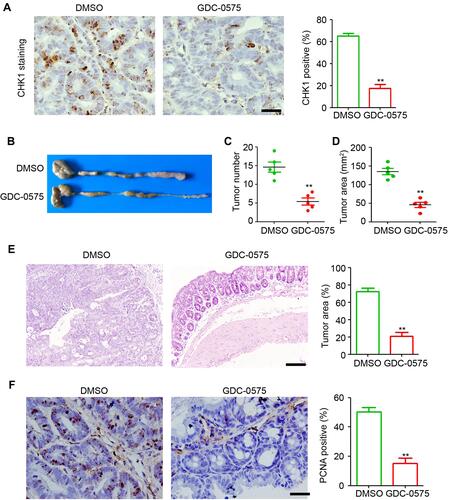
To investigate the function of CHK1 in CAC, GDC-0575 was used to treat mice with CAC. After euthanizing the mice, the colons were collected, and the effects of GDC-0575 were examined using immunohistochemical staining. The results indicated that GDC-0575 significantly inhibited the expression of colonic CHK1 in CAC mice (). Further results indicated that the inhibition of CHK1 by GDC-0575 dramatically impaired the development of CAC in mice, as evidenced by the lower tumor count and smaller tumoral area in the colons of CAC mice compared to those of the control mice (–). H&E staining also indicated that the tumors formed specifically in the colons of the CAC-DMSO group, and decreased tumor sizes were observed in the colons of the GDC-0575-treated mice with CAC (). The PCNA staining results indicated that GDC-0575 efficiently attenuated the proliferative ability of colonic cells in CAC mice (). Altogether, these results demonstrate that inhibition of CHK1 by GDC-0575 significantly impaired CAC development in mice.
Figure 1 GDC-0575 impairs the development of colitis-associated cancer in mice. (A) IHC staining of CHK1 expression in colons of CAC mice treated with DMSO or GDC-0575. The percentage of CHK1 positive cells in each frame was analyzed. Scale bar = 100 μm. (B) Image of colons of CAC mice treated with DMSO or GDC-0575. (C) The tumor number in colons of CAC mice treated with DMSO or GDC-0575 (n=5, **p<0.01). (D) The tumor area in colons of CAC mice treated with DMSO or GDC-0575 (n=5, **p<0.01). (E) H&E staining of colons from colons from CAC mice treated with DMSO or GDC-0575. The tumor area in each frame was analyzed (n=4, **p<0.01). Scale bar = 200 μm. (F) IHC staining of PCNA expression in colons of CAC mice treated with DMSO or GDC-0575. The percentage of PCNA positive cells in each frame was analyzed. Scale bar = 100 μm, **p<0.01.
GDC-0575 Impairs the Development of Colitis in Mice
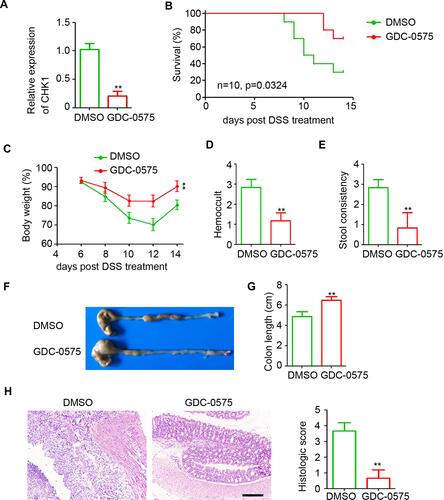
Colitis plays an important role in the development of CAC. Thus, we aimed to determine the role of CHK1 in colitis by administering GDC-0575 to mice with colitis for treatment. Quantitative PCR results indicated that GDC-0575 efficiently downregulated the expression of colonic CHK1 in mice with colitis (). Mice with low colonic CHK1 expression exhibited long survival times and low mortality during colitis development (). Body weight analysis indicated that GDC-0575 markedly attenuated colitis-induced weight loss (), hemoccult (), and stool consistency () in mice. As shown in , the colons collected from the GDC-0575 group were longer than those collected from the DMSO group, thereby indicating that the inhibition of CHK1 significantly attenuated colitis-induced colon shortening in mice (). The H&E staining results indicated that the goblet cells were disrupted and inflammatory cells infiltrated the colons of the control mice (). However, less inflammatory injury was detected in the colons of mice with colitis that received GDC-0575 treatment (). These results indicate that GDC-0575 impairs the development of colitis in mice.
Figure 2 GDC-0575 impairs the development of colitis in mice. (A) qPCR analysis of CHK1 expression in colons of colitis mice treated with DMSO or GDC-0575 (n=3, **p<0.01). (B) The survival analysis of colitis mice treated with DMSO or GDC-0575 (n=10, Log-rank (Mantel-Cox) test). (C) The body weight analysis of colitis mice treated with DMSO or GDC-0575 (n=5, **p<0.01). (D) Hemoccult analysis of colitis mice treated with DMSO or GDC-0575 (n=5, **p<0.01). (E) Stool consistency analysis of colitis mice treated with DMSO or GDC-0575 (n=5, **p<0.01). (F) Image of colons of colitis mice treated with DMSO or GDC-0575. (G) Colon length of colitis mice treated with DMSO or GDC-0575 (n=5, **p<0.01). (H) H&E staining of colons from colons from colitis mice treated with DMSO or GDC-0575. The histologic score was analyzed (n=5, **p<0.01; Scale bar = 200 μm).
GDC-0575 Inhibits Macrophage Infiltration in Colon
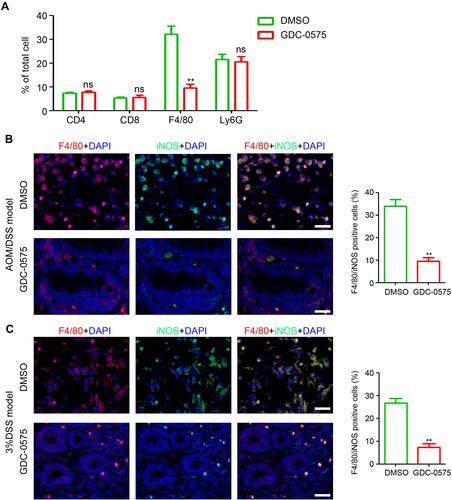
Immune cells, including T cells, macrophages, and MDSCs, participate in the pathogenesis of colitis and CAC. Flow cytometry was performed to determine the infiltration of immune cells in the colons of DMSO- or GDC-0575 treated CAC mice. As shown in , no significant differences were found in the infiltration of T cells (CD4 and CD8) and MDSCs among the colons from the DMSO and GDC-0575 groups. However, a significant reduction in macrophage infiltration was observed in the colons of the GDC-0575-treated CAC mice compared to that in DMSO-treated mice with CAC (). Further, immunofluorescence staining results indicated that the number of iNOS-positive macrophages in the colons of the GDC-0575-treated mice with CAC was less than that in the colons of the DMSO-treated mice with CAC (). The number of iNOS-positive macrophages in the colons of the GDC-0575-treated mice with colitis was also lower than that in the colons of the DMSO-treated mice with colitis (). Altogether, inhibition of CHK1 impaired the infiltration of macrophages in the colon of mice with CAC and colitis.
Figure 3 GDC-0575 inhibits the infiltration of macrophages in colons. (A) Flow cytometry analysis of immune cells infiltration in colons of CAC mice treated with DMSO or GDC-0575 (n=3, **p<0.01). (B) Immunofluorescence staining of F4/80 and iNOS expression in colons of CAC mice treated with DMSO or GDC-0575. The percentage of F4/80 and iNOS positive cells in each frame was analyzed (n=3, **p<0.01; Scale bar = 200 μm). (C) Immunofluorescence staining of F4/80 and iNOS expression in colons of colitis mice treated with DMSO or GDC-0575. The percentage of F4/80 and iNOS positive cells in each frame was analyzed (n=3, **p<0.01; Scale bar = 200 μm).
GDC-0575 Regulates Inflammatory Cytokine Expression in Colon
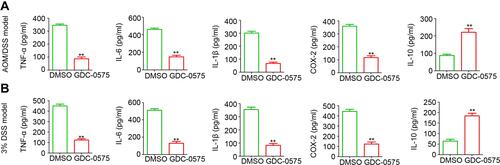
To determine the potential role of GDC-0575 in inflammatory cytokine expression, colonic tissues were collected for protein extraction and ELISA analysis. As shown in , the inhibition of CHK1 significantly inhibited the expression of TNF-α, IL-6, IL-1β, and COX-2 and markedly increased the expression of IL-10 in mice with CAC. Our results indicated a significant reduction in TNF-α, IL-6, IL-1β, and COX-2 levels and a significant increase in IL-10 expression in mice with colitis as well (). The above results confirmed the regulatory role of CHK1 in inflammatory cytokine expression in the colon of mice with CAC and colitis.
Figure 4 GDC-0575 regulates inflammatory cytokines expression in colons. (A) ELISA analysis of TNF-α, IL-6, IL-1β, COX-1, and IL-10 expression in colons of CAC mice treated with DMSO or GDC-0575 (n=3, **p<0.01). (B) ELISA analysis of TNF-α, IL-6, IL-1β, COX-2 and IL-10 expression in colons of colitis mice treated with DMSO or GDC-0575 (n=3, **p<0.01).
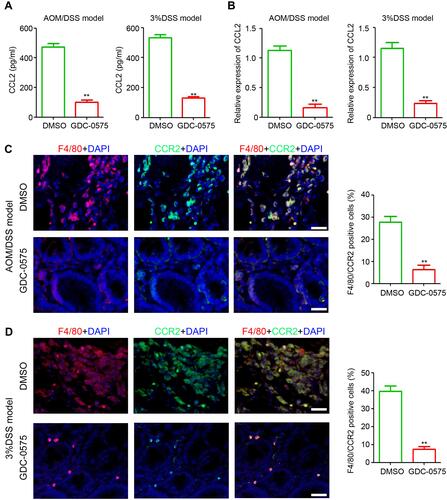
GDC-0575 Inhibits CCL2 Expression and CCR2+ Macrophage Infiltration in the Colon
CCL2 is an important cytokine involved in macrophage infiltration. CCL2 expression in the colons of mice with CAC and colitis was determined using ELISA and qPCR. The results indicated that GDC-0575 markedly inhibited the expression of colonic CCL2 in both the CAC and colitis mice at the protein () and mRNA level (). Immunofluorescence indicated that various CCR2+ macrophages infiltrated the colons of mice with CAC, and that the number of CCR2+ macrophages in the colons of CAC mice treated with GDC-0575 was decreased (). Similar inhibition of CCR2+ macrophage infiltration was also observed in the colons of colitis mice treated with GDC-0575 (). Altogether, these results indicate that CCL2 expression and CCR2+ macrophage infiltration were inhibited by GDC-0575 in CAC and colitis mice.
Figure 5 GDC-0575 inhibits CCL-2 expression and CCR2+ macrophages infiltration in colons. (A) ELISA analysis of CCL2 expression in colons of CAC mice and colitis mice treated with DMSO or GDC-0575 (n=3, **p<0.01). (B) ELISA analysis of CCL2 expression in colons of CAC mice and colitis mice treated with DMSO or GDC-0575 (n=3, **p<0.01). (C) Immunofluorescence staining of F4/80 and CCR2 expression in colons of CAC mice treated with DMSO or GDC-0575. The percentage of F4/80 and CCR2 positive cells in each frame was analyzed (n=3, **p<0.01; Scale bar = 200 μm). (D) Immunofluorescence staining of F4/80 and CCR2 expression in colons of colitis mice treated with DMSO or GDC-0575. The percentage of F4/80 and CCR2 positive cells in each frame was analyzed (n=3, **p<0.01; Scale bar = 200 μm).
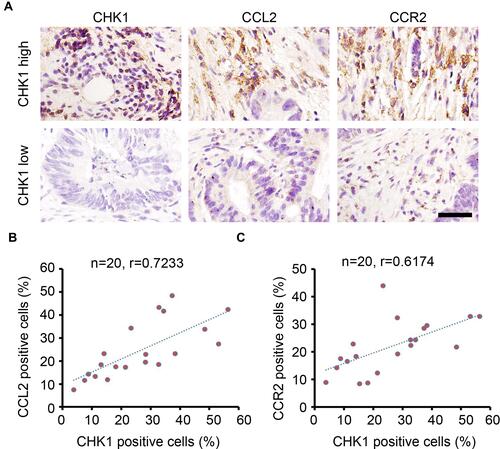
CHK1 is Positively Correlated with CCL2 and CCR2 Expression in Colorectal Cancer Tissues
Based on the analysis of colonic tissues in mice, our results indicate the regulatory role of CHK1 on CCL2 expression and CCR2+ macrophage infiltration. We confirmed the regulatory role of CHK1 by performing IHC staining of malignant tissues from patients with CRC. As shown in , high expression of CCL2 and CCR2 was observed in malignant tissues with a high expression of CHK1. The number of CHK1-, CCL2-, and CCR2-positive cells was calculated for each sample. Pearson correlation analysis indicated that CHK1 expression was positively correlated with the expression of CCL2 and CCR2 in CRC tumors ( and ). These results provide further evidence for the regulatory role of CHK1 in CCL2/CCR2 expression in CRC tumors.
Figure 6 CHK1 correlates with CCL2 and CCR2 expression in colorectal cancer tissues. (A) IHC staining of CHK1, CCL2 and CCR2 expression in colorectal cancer tissues. Scale bar = 100 μm. (B) Correlation analysis of the percentage of CHK1 and CCL2 positive cells in each sample (n=20, r = 0.7233). (C) Correlation analysis of the percentage of CHK1 and CCR2 positive cells in each sample (n=20, r = 0.6174).
Discussion
Previous studies have demonstrated a direct relationship between chronic pancreatitis and pancreatic cancer,Citation31,Citation32 Barrett’s esophagus and esophageal cancer,Citation33 and Helicobacter pylori and gastric cancer.Citation34 Patients with inflammatory bowel disease have also been reported to have a high probability of developing CAC.Citation12 In the present study, we demonstrated that GDC-0575 impairs the development of CAC and colitis in mice, and that GDC-0575 might have contributed to the inhibition of macrophage infiltration and activation in the colon of mice. Administration of GDC-0575 significantly reduced TNF-α, IL-6, and IL-1β expression and increased IL-10 expression. CCL2 expression was also downregulated by GDC-0575 in CAC and colitis mice; this downregulation was followed by the inhibition of CCR2+ macrophage infiltration in the colon. Further results confirmed a positive correlation between CHK1 expression and CCL2 and CCR2 expression in the malignant tissues of patients with CRC. The present study suggests that GDC-0575 has an inhibitory role in the development of CAC and colitis by regulating cytokine expression and inhibiting CCR2+ macrophage infiltration. Taken together, we infer that the administration of GDC-0575 is a potential therapeutic strategy for CAC and colitis.
Several studies have demonstrated the oncogenic function of CHK1 in various cancers. In solid tumor cells, CHK1 has been reported to be involved in controlling cell cycle progression by inducing CDC25 degradation.Citation35,Citation36 Furthermore, inhibition of CHK1 activity in Ewing sarcoma cells has been reported to result in DNA damage and apoptosis by promoting CDK2-mediated degradation of RRM2.Citation37 The activation of CHK1 also is also a step in the development of resistance in several solid tumors against irradiation and chemotherapy.Citation38,Citation39 The specific inhibitor GDC-0575, which targets CHK1, has been in development for two decades.Citation40,Citation41 Treatment with GDC-0575 has been reported to delay tumor growth in xenograft models.Citation40 A Phase I study has indicated that GDC-0575 can be safely administered as a monotherapy and in combination with gemcitabine, and that it has preliminary antitumor activity in several solid tumors, including those of breast cancer.Citation41 In our study, we investigated the potential role of CHK1 in CAC, and demonstrated that GDC-0575 has efficient antitumor activity in mice. Furthermore, the inhibition of GDC-0575 attenuated DSS-induced colonic damage in mice and the expression of pro-inflammatory cytokines in mice with colitis. These results indicate that GDC-0575 has anti-tumor and anti-inflammatory functions in mice; however, studies with large sample sizes and human patients are needed to further investigate the potential therapeutic application of GDC-0575.
Macrophages are a class of monocytes that play a crucial role in inflammation and immune responses.Citation42 In the development of colitis and CAC, macrophages infiltrate the colon and release a series of cytokines to remodel the affected microenvironment and promote the initiation and progression of colon cancer.Citation43–Citation45 The inhibition of macrophage infiltration or activation induces an efficient antitumor and anti-colitis effect, respectively, in mice and also in clinical use for humans.Citation43,Citation46,Citation47 Our study demonstrated that the number of activated macrophages that infiltrated the colons of GDC-0575-treated CAC and colitis mice decreased compared with that of the DMSO-treated control mice; this decrease was accompanied by a reduction in the expression of pro-inflammatory cytokines that were released by the infiltrating macrophages. The expression of CCL2, which is required for the expression of CCR2 in macrophages,Citation48,Citation49 was downregulated in the GDC-0575-treated colitis and CAC mice; following the reduction of CCL2, the infiltration of CCR2-positive macrophages was inhibited. Thus, we speculated that GDC-0575 inhibits CCL2 expression and CCR2+ macrophage infiltration in mice colon. However, the molecular mechanism underlying the regulatory cascade of CHK1 upon CCL2 expression remains unclear. The results of CCL2 mRNA expression in AOM/DSS model and 3%DSS model shown that GDC-0575 inhibits the transcription of CCL2, which indicated that CHK1 regulates the CCL2 expression at transcriptional level. Further studies are required to elucidate the details of this mechanism. Altogether, the present study provides evidence regarding the function and mechanism of CHK1 in regulating the development of CAC and colitis. The use of GDC-0575, a specific inhibitor of CHK1, is a potential therapeutic strategy for CAC and colitis. Further studies are necessary to determine the adverse effects of GDC-0575 administration in mice.
Ethics Approval and Consent to Participate
This animal study was approved by the Ethics Committee of Jishou University and complied with the animal guidelines.
Disclosure
The authors declare that they have no competing interests.
Additional information
Funding
References
- Feng RM, Zong YN, Cao SM, Xu RH. Current cancer situation in China: good or bad news from the 2018 Global Cancer Statistics? Cancer Commun. 2019;39:22. doi:10.1186/s40880-019-0368-6
- Ferlay J, Colombet M, Soerjomataram I, et al. Cancer incidence and mortality patterns in Europe: estimates for 40 countries and 25 major cancers in 2018. Eur J Cancer. 2018;103:356–387. doi:10.1016/j.ejca.2018.07.005
- Azeem S, Gillani SW, Siddiqui A, Jandrajupalli SB, Poh V, Syed Sulaiman SA. Diet and colorectal cancer risk in Asia–a systematic review. Asian Pac J Cancer Prev. 2015;16:5389–5396. doi:10.7314/APJCP.2015.16.13.5389
- Gao R, Gao Z, Huang L, Qin H. Gut microbiota and colorectal cancer. Eur J Clin Microbiol Infect Dis. 2017;36:757–769. doi:10.1007/s10096-016-2881-8
- Thanikachalam K, Khan G. Colorectal cancer and nutrition. Nutrients. 2019;11(1):11. doi:10.3390/nu11010164
- Baek SJ, Kim SH. Colitis-associated colorectal cancer in patients with inflammatory bowel disease. Minerva Chir. 2017;72:520–529. doi:10.23736/S0026-4733.17.07437-5
- Bopanna S, Ananthakrishnan AN, Kedia S, Yajnik V, Ahuja V. Risk of colorectal cancer in Asian patients with ulcerative colitis: a systematic review and meta-analysis. Lancet Gastroenterol Hepatol. 2017;2(4):269–276. doi:10.1016/S2468-1253(17)30004-3
- Rogler G. Chronic ulcerative colitis and colorectal cancer. Cancer Lett. 2014;345(2):235–241. doi:10.1016/j.canlet.2013.07.032
- Cooks T, Pateras IS, Tarcic O, et al. Mutant p53 prolongs NF-κB activation and promotes chronic inflammation and inflammation-associated colorectal cancer. Cancer Cell. 2013;23:634–646. doi:10.1016/j.ccr.2013.03.022
- Porta C, Ippolito A, Consonni FM, et al. Protumor steering of cancer inflammation by p50 NF-κB enhances colorectal cancer progression. Cancer Immunol Res. 2018;6:578–593. doi:10.1158/2326-6066.CIR-17-0036
- Bromberg J, Wang TC. Inflammation and cancer: IL-6 and STAT3 complete the link. Cancer Cell. 2009;15:79–80. doi:10.1016/j.ccr.2009.01.009
- Yashiro M. Ulcerative colitis-associated colorectal cancer. World J Gastroenterol. 2014;20(44):16389–16397. doi:10.3748/wjg.v20.i44.16389
- Medzhitov R. Origin and physiological roles of inflammation. Nature. 2008;454(7203):428–435. doi:10.1038/nature07201
- Singh N, Baby D, Rajguru JP, Patil PB, Thakkannavar SS, Pujari VB. Inflammation and cancer. Ann Afr Med. 2019;18(3):121–126. doi:10.4103/aam.aam_56_18
- Wynn TA, Chawla A, Pollard JW. Macrophage biology in development, homeostasis and disease. Nature. 2013;496(7446):445–455. doi:10.1038/nature12034
- Wynn TA, Vannella KM. Macrophages in tissue repair, regeneration, and fibrosis. Immunity. 2016;44(3):450–462. doi:10.1016/j.immuni.2016.02.015
- Bain CC, Schridde A. Origin, differentiation, and function of intestinal macrophages. Front Immunol. 2018;9:2733. doi:10.3389/fimmu.2018.02733
- Benoit M, Desnues B, Mege JL. Macrophage polarization in bacterial infections. J Immunol. 2008;181:3733–3739. doi:10.4049/jimmunol.181.6.3733
- Ngambenjawong C, Gustafson HH, Pun SH. Progress in tumor-associated macrophage (TAM)-targeted therapeutics. Adv Drug Deliv Rev. 2017;114:206–221. doi:10.1016/j.addr.2017.04.010
- Qiu Z, Oleinick NL, Zhang J. ATR/CHK1 inhibitors and cancer therapy. Radiother Oncol. 2018;126(3):450–464. doi:10.1016/j.radonc.2017.09.043
- Zhang Y, Hunter T. Roles of Chk1 in cell biology and cancer therapy. Int J Cancer. 2014;134(5):1013–1023. doi:10.1002/ijc.28226
- Rundle S, Bradbury A, Drew Y, Curtin NJ. Targeting the ATR-CHK1 axis in cancer therapy. Cancers. 2017;9(12):41. doi:10.3390/cancers9050041
- Hwang BJ, Adhikary G, Eckert RL, Lu AL. Chk1 inhibition as a novel therapeutic strategy in melanoma. Oncotarget. 2018;9:30450–30464. doi:10.18632/oncotarget.25765
- Dent P. Investigational CHK1 inhibitors in early phase clinical trials for the treatment of cancer. Expert Opin Investig Drugs. 2019;28(12):1095–1100. doi:10.1080/13543784.2019.1694661
- González Besteiro MA, Gottifredi V. The fork and the kinase: a DNA replication tale from a CHK1 perspective. Mutat Res Rev Mutat Res. 2015;763:168–180. doi:10.1016/j.mrrev.2014.10.003
- Di Tullio A, Rouault-Pierre K, Abarrategi A, et al. The combination of CHK1 inhibitor with G-CSF overrides cytarabine resistance in human acute myeloid leukemia. Nat Commun. 2017;8(1):1679. doi:10.1038/s41467-017-01834-4
- Parang B, Barrett CW, Williams CS. AOM/DSS model of colitis-associated cancer. Methods Mol Biol. 2016;1422:297–307.
- Dai L, Cui X, Zhang X, et al. SARI inhibits angiogenesis and tumour growth of human colon cancer through directly targeting ceruloplasmin. Nat Commun. 2016;7(1):11996. doi:10.1038/ncomms11996
- Wirtz S, Popp V, Kindermann M, et al. Chemically induced mouse models of acute and chronic intestinal inflammation. Nat Protoc. 2017;12(7):1295–1309. doi:10.1038/nprot.2017.044
- Dai L, Liu Y, Cheng L, et al. SARI attenuates colon inflammation by promoting STAT1 degradation in intestinal epithelial cells. Mucosal Immunol. 2019;12(5):1130–1140. doi:10.1038/s41385-019-0178-9
- Kirkegård J, Mortensen FV, Cronin-Fenton D. Chronic pancreatitis and pancreatic cancer risk: a systematic review and meta-analysis. Am J Gastroenterol. 2017;112(9):1366–1372. doi:10.1038/ajg.2017.218
- Raimondi S, Lowenfels AB, Morselli-Labate AM, Maisonneuve P, Pezzilli R. Pancreatic cancer in chronic pancreatitis; aetiology, incidence, and early detection. Best Pract Res Clin Gastroenterol. 2010;24(3):349–358. doi:10.1016/j.bpg.2010.02.007
- Thrift AP. Barrett’s Esophagus and Esophageal adenocarcinoma: how common are they really? Dig Dis Sci. 2018;63(8):1988–1996. doi:10.1007/s10620-018-5068-6
- Wang F, Meng W, Wang B, Qiao L. Helicobacter pylori-induced gastric inflammation and gastric cancer. Cancer Lett. 2014;345(2):196–202. doi:10.1016/j.canlet.2013.08.016
- Goto H, Natsume T, Kanemaki MT, et al. Chk1-mediated Cdc25A degradation as a critical mechanism for normal cell cycle progression. J Cell Sci. 2019;132(2):jcs223123. doi:10.1242/jcs.223123
- Shen T, Zhou H, Shang C, Luo Y, Wu Y, Huang S. Ciclopirox activates ATR-Chk1 signaling pathway leading to Cdc25A protein degradation. Genes Cancer. 2018;9(1–2):39–52. doi:10.18632/genesandcancer.166
- Koppenhafer SL, Goss KL, Terry WW, Gordon DJ. Inhibition of the ATR-CHK1 pathway in ewing sarcoma cells causes DNA damage and apoptosis via the CDK2-mediated degradation of RRM2. Mol Cancer Res. 2020;18:91–104. doi:10.1158/1541-7786.MCR-19-0585
- Bai M, Song N, Che X, Wang X, Qu X, Liu Y. Chk1 activation attenuates sensitivity of lapatinib in HER2-positive gastric cancer. Cell Biol Int. 2018;42(7):781–793. doi:10.1002/cbin.10922
- Dent P, Tang Y, Yacoub A, Dai Y, Fisher PB, Grant S. CHK1 inhibitors in combination chemotherapy: thinking beyond the cell cycle. Mol Interv. 2011;11:133–140. doi:10.1124/mi.11.2.11
- Grellety T, Callens C, Richard E, et al. Enhancing abiraterone acetate efficacy in androgen receptor-positive triple-negative breast cancer: chk1 as a potential target. Clin Cancer Res. 2019;25:856–867. doi:10.1158/1078-0432.CCR-18-1469
- Italiano A, Infante JR, Shapiro GI, et al. Phase I study of the checkpoint kinase 1 inhibitor GDC-0575 in combination with gemcitabine in patients with refractory solid tumors. Ann Oncol. 2018;29:1304–1311. doi:10.1093/annonc/mdy076
- Gordon S, Martinez FO. Alternative activation of macrophages: mechanism and functions. Immunity. 2010;32:593–604. doi:10.1016/j.immuni.2010.05.007
- Gowrikumar S, Ahmad R, Uppada SB, et al. Upregulated claudin-1 expression promotes colitis-associated cancer by promoting β-catenin phosphorylation and activation in Notch/p-AKT-dependent manner. Oncogene. 2019;38(26):5321–5337. doi:10.1038/s41388-019-0795-5
- Katholnig K, Schütz B, Fritsch SD, et al. Inactivation of mTORC2 in macrophages is a signature of colorectal cancer that promotes tumorigenesis. JCI Insight. 2019;4(20). doi:10.1172/jci.insight.124164
- Yao D, Dong M, Dai C, Wu S. Inflammation and inflammatory cytokine contribute to the initiation and development of ulcerative colitis and its associated cancer. Inflamm Bowel Dis. 2019;25(10):1595–1602. doi:10.1093/ibd/izz149
- Hardbower DM, Coburn LA, Asim M, et al. EGFR-mediated macrophage activation promotes colitis-associated tumorigenesis. Oncogene. 2017;36(27):3807–3819. doi:10.1038/onc.2017.23
- Singh K, Coburn LA, Asim M, et al. Ornithine decarboxylase in macrophages exacerbates colitis and promotes colitis-associated colon carcinogenesis by impairing M1 immune responses. Cancer Res. 2018;78(15):4303–4315. doi:10.1158/0008-5472.CAN-18-0116
- Kawano Y, Nakae J, Watanabe N, et al. Colonic pro-inflammatory macrophages cause insulin resistance in an intestinal Ccl2/Ccr2-dependent manner. Cell Metab. 2016;24(2):295–310. doi:10.1016/j.cmet.2016.07.009
- Li X, Yao W, Yuan Y, et al. Targeting of tumour-infiltrating macrophages via CCL2/CCR2 signalling as a therapeutic strategy against hepatocellular carcinoma. Gut. 2017;66(1):157–167. doi:10.1136/gutjnl-2015-310514