Abstract
Background
Breast cancer stem cells with a CD44+CD24− phenotype are the origin of breast tumors. Strong CD44 expression in this population indicates its important role in maintaining the stem cell phenotype. Previous studies show that CD44 down-regulation causes CD44+CD24− breast cancer stem cells to differentiate into non-stem cells that are sensitive to antitumor drugs and lose many characteristics of the original cells. In this study, we determined tumor suppression in non-obese severe combined immunodeficiency mice using CD44 shRNA therapy combined with doxorubicin treatment.
Methods
Tumor-bearing non-obese severe combined immunodeficiency mice were established by injection of CD44+CD24− cells. To track CD44+CD24− cells, green fluorescence protein was stably transduced using a lentiviral vector prior to injection into mice. The amount of CD44 shRNA lentiviral vector used for transduction was based on CD44 down-regulation by in vitro CD44 shRNA transduction. Mice were treated with direct injection of CD44 shRNA lentiviral vector into tumors followed by doxorubicin administration after 48 hours. The effect was evaluated by changes in the size and weight of tumors compared with that of the control.
Results
The combination of CD44 down-regulation and doxorubicin strongly suppressed tumor growth with significant differences in tumor sizes and weights compared with that of CD44 down-regulation or doxorubicin treatment alone. In the combination of CD44 down-regulation and doxorubicin group, the tumor weight was significantly decreased by 4.38-fold compared with that of the control group.
Conclusion
These results support a new strategy for breast cancer treatment by combining gene therapy with chemotherapy.
Introduction
CD44+CD24− cells have been identified as a breast cancer stem cell population and the origin of tumors, metastasis, and relapse in breast cancer patients.Citation1–Citation3 Breast cancer stem cell targeting is considered a promising therapy. Thus far, various drugs that are specific to receptors such as Her2/neu and epidermal growth factor receptors have been used to target breast cancer stem cells.Citation4–Citation11 However, more than 50% of tumors do not express these receptors and are drug resistant.Citation12–Citation16 A recent report has shown that triple-negative breast carcinoma contains CD44+CD24− breast cancer stem cells.Citation17 Therefore, it is essential for treatment that new targets be discovered on breast cancer stem cells.
CD44 plays an important role in the phenotype of breast cancer stem cells and is responsible for cancer stem cell-specific characteristics, such as antitumor drug resistance in various cancers like colon cancer,Citation18 salivary gland cancer,Citation19 and metastasis from the breast to the liver.Citation20 In addition, CD44 has been used to isolate and enrich cells that are capable of forming breast cancer tumorsCitation21 and numerous other tumors, including head and neck squamous cell carcinoma,Citation22–Citation24 esophageal squamous cell carcinoma,Citation25 nasopharyngeal carcinoma,Citation26 and gastricCitation27 and colon cancer stem cells.Citation28
Down-regulation of CD44 using siRNA or shRNA results in metastasis suppression,Citation29 sensitizes cancer stem cells to drugs,Citation30 and causes differentiation of breast cancer stem cells.Citation31 Antibodies against survivin also show similar effects.Citation32 CD44 also plays an important role in other cancers. CD44 inhibition suppresses the development of colon tumors in miceCitation33 and inhibits the proliferation and metastasis of ovarianCitation34 and liver cancer cells.Citation35 This study evaluates breast cancer treatment in mouse models using a CD44 shRNA lentiviral vector to inhibit CD44 expression in combination with doxorubicin chemotherapy.
Materials and methods
Cell culture and establishment of green fluorescent protein (GFP)-expressing breast cancer stem cells
Breast cancer stem cells were isolated and purified as described elsewhere.Citation30 Briefly, tumor biopsies from consenting patients were obtained at hospitals and then transferred to our laboratory. Biopsy samples were washed 3–4 times with phosphate-buffered saline containing 1 × antibiotic-antimycotic (Sigma, St Louis, MO), and then homogenized into small pieces (approximately 1–2 mm3). Homogenized samples were resuspended in M171 medium (Invitrogen, Carlsbad, CA) containing mammary epithelial growth supplement (Invitrogen) and then seeded in 35 mm culture dishes (Nunc, Germany). Cells were incubated at 37°C with 5% CO2, and medium was replaced every third day. CD44+CD24− cells were isolated from the primary cell population by magnetic sorting using a commercial kit (Miltenyi Biotec, Germany). These CD44+CD24− cells were named BCSC1. For tracking, we established CD44+CD24− cells that stably expressed the gfp gene. We used a gfp lentiviral vector (Santa Cruz Biotechnology, CA) to transduce isolated CD44+CD24− cells. To select and establish GFP-expressing BCSC1, cells were cultured in medium containing 10 μg/mL puromycin dihydrochloride (Sigma-Aldrich, St Louis, MO) for 1 week.
CD44 knockdown of CD44+CD24− cells with shRNA using lentivirus particles
In the first assay, we determined a suitable dose of lentiviral particle vector infectious units (IFUs) to apply in the next experiment. CD44 shRNA lentivirus particles (Santa Cruz Biotechnology, Inc, Santa Cruz, CA) were stably transfected according to the manufacturer’s instructions. Briefly, BCSC1 cells were seeded on day 1 in a twelve-well plate with complete medium (Dulbecco’s modif ied Eagle’s medium/F12 supplemented with 10% fetal bovine serum and 1 × antibiotic-mycotic) and incubated overnight.
Medium was replaced on day 2 with fresh complete medium containing 5 μg/mL polybrene (Sigma-Aldrich, St Louis, MO) for 6 hours, then 20 μL of modified Eagle’s medium with 25 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid containing 1 × 105 IFUs of virus was added to the culture. The culture plate was shaken to mix the virus particles and was then incubated overnight at 37°C with 5% CO2. On day 3, medium was replaced with fresh complete medium without polybrene. Half of the transduced cells were confirmed by CD44 detection using flow cytometry. Half of the transduced cells were selected by culturing in complete medium containing 10 μg/mL puromycin dihydrochloride for 12 hours, followed by 5 μg/mL puromycin dihydrochloride for 1 week.
Flow cytometry
Cells were washed twice in phosphate-buffered saline containing 1% bovine serum albumin (Sigma-Aldrich, St Louis, MO). Fc receptors were blocked by incubation with immunoglobulin G (Santa Cruz Biotechnology, CA) on ice for 15 minutes. Cells were stained with anti-CD44-PE monoclonal antibodies (BD Biosciences, Franklin Lakes, NJ) at 4°C for 30 minutes. After washing, cells were analyzed using a FAC SCalibur flow cytometer (BD Biosciences) and CellQuest Pro software (BD Biosciences) with 10,000 events collected.
CD44 shRNA gene therapy
Female (5–6 weeks old) NOD/severe combined immunodeficiency (SCID) mice (NOD.CB17-Prkdcscid/J; Charles River Laboratories, Wilmington, MA) were subcutaneously injected with BCSC1 cells (2 × 106 cells/mouse). After 2 weeks, tumors were formed and mice were divided into four groups: Group 1 (control) mice (n = 4) were used as untreated controls; they received biweekly intratumoral phosphate-buffered saline injections for 6 weeks. Group 2 (doxorubicin [Dox]) mice received intratumoral Dox injections (2 mg/kg) weekly for 4 weeks. Group 3 (shRNA) mice received intratumoral CD44 shRNA lentiviral vector injections with a dose of IFUs that was doubled compared with that of the tumor cell number. Group 4 (CD44 shRNA in combination with Dox treatment [shRNA + Dox]) mice received intratumoral injections of CD44 shRNA lentiviral vector with IFUs similar to that of Group 3 and, after 48 hours, received intratumoral injections of Dox (2 mg/kg) weekly for 4 weeks. Tumor size was measured as described below. Animals were killed after 7 weeks, and tumors were excised and weighed to record the wet tumor weight. All animal experiments were approved by the Institutional Animal Care and Use Committee of Stem Cell Research and Application Laboratory, University of Science, VNU-HCM.
Tumor size measurement
Tumor size was measured with calipers in two dimensions, and size was calculated using the following formula: a × b2/2, where “a” is the tumor length and “b” is the diameter.Citation36
Statistical analysis
All experimental procedures were performed in triplicate, except for mouse experiments. The significance of differences between mean values was assessed by a Student’s t-test and analysis of variance. P < 0.05 was considered significant.
Results
Isolation and establishment of breast cancer stem cells expressing green fluorescent protein
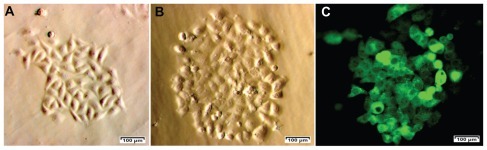
We primary-cultured 31 tumor samples from patients; 23 of these samples showed numerous single cells surrounding the tumor tissue. Cells from the 23 samples were allowed to propagate to 80% confluence (). CD44 and CD24 were analyzed and all 23 primary cell samples showed a small population of cells that were positive for CD44 and negative or weakly positive for CD24. This population constituted 3.96% ± 1.72% of the total cells derived from primary culture. We isolated two populations of CD44+CD24− cells from the 23 primary-culture samples. One cell population, termed “BCSC1,” was used for subsequent experiments (). The BCSC1 cell line was transduced with the gfp gene using a lentiviral vector, resulting in 43.12% and 99.9% of BCSC1 cells expressing GFP before and after selection with puromycin, respectively ().
Figure 1 Breast cancer cells from breast tumors (A) were used to isolate CD44+CD24− breast cancer stem cell populations (B) for green fluorescent protein expression after transduction with green fluorescent protein using a lentiviral vector and selection with puromycin (C).
Tumor-bearing mouse models
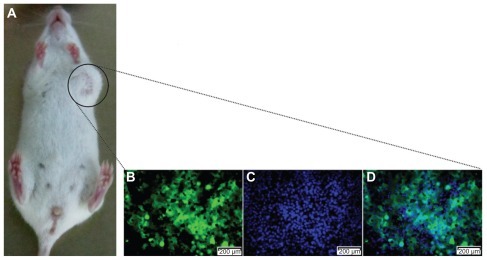
To establish the tumor-bearing mouse models, we used 5–6-week-old NOD/SCID mice. GFP-expressing BCSC1 cells (2.106 cells/mouse) were injected into mammary fat using an insulin needle. This resulted in 100% of mice forming tumors that were apparent after 3 weeks. All tumors contained GFP-expressing cells ().
Figure 2 A tumor produced in the mouse model. The tumor (A) was excised and observed by monochromatic fluorescence microscopy (Carl Zeiss AG, Oberkochen, Germany) using fluorescein isothiocyanate (B) and Hoechst 33342 filters (Carl Zeiss AG, Oberkochen, Germany) (C) for a merged image (D).
In vitro CD44 down-regulation by the CD44 shRNA lentiviral vector
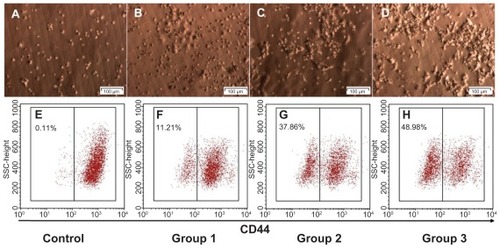
Next, we evaluated in vitro CD44 down-regulation with CD44 shRNA using a lentiviral vector to determine a suitable dose for in vivo transduction. CD44 down-regulation was dependent on the ratio of IFUs to BCSC1 cells, with a higher ratio of lentiviral vector to BCSC1 cells resulting in higher transduction efficiency. The percentages of CD44 down-regulated BCSC1 cells in the control (1:0), Dox (2:1), CD44 shRNA (1:1), and CD44 shRNA + Dox (1:2) groups were 0.14% ± 0.08%, 12.21% ± 3.30%, 37.87% ± 5.34%, and 47.41% ± 3.90%, respectively (P < 0.05) (). Based on these results, the suitable dose of lentiviral vector IFUs was double that of the number of tumor cells. This dose was applied in further experimentation.
Figure 3 In vitro CD44 down-regulation using the CD44 shRNA lentiviral vector with doses of infectious units to breast cancer stem cells at ratios 1:0 (A and E), 2:1 (B and F), 1:1 (C and G) and 1:2 (D and H).
Tumor size and weight
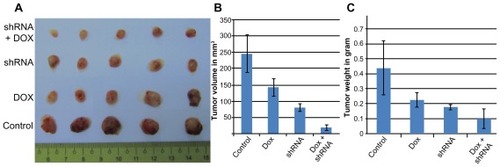
As shown in , the size and weight of tumors were significantly different between the four groups (P < 0.05). The average tumor sizes were 246.39 ± 56.80 mm3, 142 ± 25.98 mm3, 80.89 ± 11.11 mm3, and 19.75 ± 8.50 mm3 in the control, Dox, CD44 shRNA, and CD44 shRNA + Dox groups, respectively. In comparison with the control group, the tumor sizes were significantly decreased by 1.74-, 3.04-, and 12.47-fold in the Dox, CD44 shRNA, and CD44 shRNA + Dox groups, respectively. Tumor weights also gradually decreased (0.44 ± 0.18 g, 0.23 ± 0.05 g, 0.18 ± 0.02 g, and 0.1 ± 0.07 g). In CD44 shRNA + Dox, the tumor weight was significantly decreased by 4.38-fold compared with that of the control group. These changes in tumor size and weight confirmed the beneficial effects of CD44 down-regulation, Dox treatment, and particularly, the combination of CD44 down-regulation and Dox treatment. Thus, combinatorial therapy of CD44 down-regulation and Dox efficiently suppressed tumor growth in the mouse model.
Figure 4 Tumor size and weight in experimental groups (A). Graphs of the differences in the size (B) and weight (C) of tumors in control, Dox, CD44 shRNA, and CD44 shRNA + Dox groups.
Discussion
Cancer stem cells are considered the origin of malignant tissues. The existence of cancer stem cells has been recently confirmed in solid tumors of the brain, prostate, pancreas, liver, colon, head and neck, lung, and skin.Citation36–Citation42 Moreover, CD44+CD24− cells have been identified as breast cancer stem cells.Citation21
Since the discovery of cancer stem cells, the study of cancer treatment in general, and breast cancer in particular, has gradually focused on targeting cancer stem cells. Thus far, targeting of breast cancer stem cells has been performed using various approaches, but has mainly targeted self-renewal and differentiation of breast cancer stem cells. To influence self-renewal and differentiation, signaling pathways that are important in breast cancer stem cells, such as Wnt, Notch, and Hedgehog, can be targeted.Citation43–Citation46 There are numerous methods to target signaling pathways, including gene therapy, immunotherapy, and targeting the cell environment. In our previous study, we found that CD44 down-regulation reduces the drug resistance of breast cancer stem cells to Dox.Citation30 In previous research, we also confirmed that CD44 shRNA lentiviral particles reduced CD44 expression and caused breast cancer stem cell differentiation.Citation31 In this study, we used an experimental treatment to target breast cancer stem cells by combining gene therapy targeting CD44 and Dox treatment.
First, we established a breast cancer stem cell line that stably expressed GFP to monitor the xenografted breast cancer tumor in mice. To establish this cell line, breast cancer stem cells were transduced with a lentiviral vector carrying gfp and a puromycin resistance gene for selection. Because random insertion of lentiviral DNA into the genome can cause detrimental mutations, we isolated CD44+CD24− cells from GFP-breast cancer stem cells using a magnetic cell separation method, and re-analyzed with flow cytometry. Indeed, a study showed that lentiviral vectors demonstrate a low tendency to integrate into genes that cause cancer,Citation47 and another study found no increase in tumor incidence and no earlier onset of tumors in a mouse strain following the use of lentiviral vectors.Citation48
These BSCS1 cells were used to evaluate the potential to form tumors in NOD/SCID mice and CD44 knockdown mice using a CD44 shRNA lentiviral vector as well as determination of the optimal dose of lentiviral particles for in vivo analyses. GFP-expressing BCSC1 maintained a tumorigenic capacity and formed malignant tumors in NOD/SCID mice with numerous poorly differentiated and abnormal cells.
Next, we determined the appropriate dose of virus particles to infect tumors, which was considered to be the IFUs that down-regulated CD44 at the highest rate. To determine the appropriate dose, we conducted serial assays with ratios between cells and IFUs at 2:1, 1:1, and 1:2. CD44 down-regulation was highest using double the IFUs compared with that of the cell number. To determine the number of cells in a tumor, we measured the tumor size at the time of treatment. The number of tumor cells is calculated as 1 cm3 tumor contains ~1 × 109 cells.Citation49 Although recent studies have supported this claim,Citation50–Citation52 experiments using the same mouse breed under the same conditions are necessary to apply this rule in calculation and comparison among the mice.
Lentiviral vector-injected mice were treated with Dox after 48 hours. This period was chosen because previous study has shown that viruses infect target cells and inhibit CD44 expression after 24 hours.Citation30 The Dox dose used was 2 mg/kg body weight and this was chosen based on a previous study.Citation53
The results showed significant differences in the size and weight of tumors of treated mice compared with those of the controls. Dox treatment and CD44 siRNA therapy alone or in combination inhibited tumor growth. Tumor inhibition with Dox treatment and CD44 shRNA therapy alone was identical, while a significant difference (P < 0.05) was demonstrated between combinatorial therapy with Dox and CD44 shRNA compared with that of single treatments.
CD44 down-regulation also effects adhesion, invasion, and metastasis,Citation54–Citation59 and the inhibition of CD44 is also considered as treatment therapy in many cancer targets.Citation57–Citation59 In addition, CD44 down-regulation that suppresses the development of tumors has also been shown in in vivo colon cancer tumors,Citation33 ovarian cancer cells,Citation34 and nasopharyngeal carcinoma cells.Citation60–Citation61 In recent research, we recognized that CD44 maintains the stemness of breast cancer stem cells. CD44 knocked-down breast cancer stem cells by CD44 shRNA lentiviral particles can cause differentiation of breast cancer stem cells or loss of stemness can change the tumor formation and metastasis related genes, and can reduce tumor formations in NOD/SCID mice.Citation31
CD44 down-regulation using shRNA suppressed xenografted breast tumor growth in a mouse model with or without Dox treatment. However, there are limitations for the clinical application of this therapy. The two most significant issues are the host’s immune response to the lentiviral vector and random insertion mutagenesis. The immune response to the lentiviral vector is very low because viral proteins are not translated. Therefore, an immune response occurs only as a primary response to the virus or products of transgenes. In this study, the lentiviral vector was only transcribed into shRNA. Moreover, an immune response occurs only in response to adenoviral vectors or in the nature of the mechanism of adeno-associated viral production of antibodies against them,Citation62 while lentiviral vectors possess many traits that enable avoidance of the immune response. As mentioned, insertion mutations caused by lentiviral vectors are fewer and less serious compared with those caused by other vectors. Insertion mutations have been detected in three out of eleven cross-linked SCID children after applying ex vivo therapy using a murine leukemia virus vector.Citation63 Murine leukemia virus vectors are often inserted into promoters and CpG islands that affect transcriptionally active genes.Citation64,Citation65 Integrations near transcription start sites may increase oncogenesis, either by influencing the activity of host promoters or producing new full-length transcripts. In contrast, lentiviral vectors that integrate into the entire transcribed region are less likely to disturb the regulation and expression of host genes.Citation66 This claim is supported by a Montini et al,Citation48 which showed that lentiviral vectors cause insertion mutations related to cancer less often compared with murine leukemia virus vectors in a mouse model. However, these problems can be solved by using site-specific gene transfer. With the structural advantages of this vector system, cassettes that contain numerous genes can be expressed in the same vector, such as a Cre recombinase in combination with loxP sites or a zinc finger nuclease. However, there are some limitations in applying these results in clinical trials. First, the high dose of lentiviral vector can cause some side effects; in particular, lentiviral vectors can migrate into bone marrow to suppress the mesenchymal stem cells and other cells that strongly express CD44. Second, in practice, intratumoral delivery is not generally carried out. However, as many kinds of cells as well as stem cells strongly express CD44, we cannot apply systemic therapy in this case.
Conclusion
Strong CD44 expression in a breast cancer stem cell population with a CD44+CD24− phenotype plays a pivotal role in the proliferation and drug resistance of malignant cells. Our data suggest that CD44 down-regulation suppresses tumor growth in a mouse model. Combinatorial therapy of CD44 down-regulation using a CD44 shRNA lentiviral vector and Dox treatment strongly inhibits tumor growth. These results support a new targeted therapy using gene therapy and chemotherapy to eradicate breast cancer stem cells. If this therapy is found to be safe, it may be a promising therapy for breast cancer through the targeting of breast cancer stem cells.
Disclosure
The authors report no conflicts of interest in this work.
References
- MeyerMJFlemingJMAliMAPeseskyMWGinsburgEVonderhaarBKDynamic regulation of CD24 and the invasive, CD44posCD24neg phenotype in breast cancer cell linesBreast Cancer Res200911R8219906290
- SheridanCKishimotoHFuchsRKCD44+/CD24− breast cancer cells exhibit enhanced invasive properties: an early step necessary for metastasisBreast Cancer Res20068R5917062128
- TiezziDGValejoFAMaranaHRCD44(+)/CD24(−) cells and lymph node metastasis in stage I and II invasive ductal carcinoma of the breastMed Oncol Epub June 29, 2011
- JohnstonSRTargeting downstream effectors of epidermal growth factor receptor/HER2 in breast cancer with either farnesyltransferase inhibitors or mTOR antagonistsInt J Gynecol Cancer20061654354817010069
- NandaRTargeting the human epidermal growth factor receptor 2 (HER2) in the treatment of breast cancer: recent advances and future directionsRev Recent Clin Trials2007211111618473995
- PiechockiMPYooGHDibbleySKLonardoFBreast cancer expressing the activated HER2/neu is sensitive to gefitinib in vitro and in vivo and acquires resistance through a novel point mutation in the HER2/neuCancer Res2007676825684317638894
- ReidAVidalLShawHde BonoJDual inhibition of ErbB1 (EGFR/HER1) and ErbB2 (HER2/neu)Eur J Cancer20074348148917208435
- SantinADBelloneSRomanJJMcKenneyJKPecorelliSTrastuzumab treatment in patients with advanced or recurrent endometrial carcinoma overexpressing HER2/neuInt J Gynaecol Obstet200810212813118555254
- FreudenbergJAWangQKatsumataMDrebinJNagatomoIGreeneMIThe role of HER2 in early breast cancer metastasis and the origins of resistance to HER2-targeted therapiesExp Mol Pathol20098711119450579
- Diermeier-DaucherSBreindlSBuchholzSOrtmannOBrockhoffGModular anti-EGFR and anti-Her2 targeting of SK-BR-3 and BT474 breast cancer cell lines in the presence of ErbB receptor-specific growth factorsCytometry A201179A68469321786419
- LiuTYacoubRTaliaferro-SmithLDCombinatorial effects of lapatinib and rapamycin in triple-negative breast cancer cellsMol Cancer Ther2011101460146921690228
- MassarwehSOsborneCKCreightonCJTamoxifen resistance in breast tumors is driven by growth factor receptor signaling with repression of classic estrogen receptor genomic functionCancer Res20086882683318245484
- YonesakaKZejnullahuKOkamotoIActivation of ERBB2 signaling causes resistance to the EGFR-directed therapeutic antibody cetuximabSci Transl Med2011399ra8699ra86
- ChenYJHuangWCWeiYLElevated BCRP/ABCG2 expression confers acquired resistance to gefitinib in wild-type EGFR-expressing cellsPLoS One20116e2142821731744
- JinKKongXShahTPenetMFThe HOXB7 protein renders breast cancer cells resistant to tamoxifen through activation of the EGFR pathwayProc Natl Acad Sci U S A Epub June 20, 2011
- BrandTMIidaMWheelerDLMolecular mechanisms of resistance to the EGFR monoclonal antibody cetuximabCancer Biol Ther20111177779221293176
- IdowuMOKmieciakMDumurCCD44(+)/CD24(−/low) cancer stem/progenitor cells are more abundant in triple-negative invasive breast carcinoma phenotype and are associated with poor outcomeHum Pathol Epub August 10, 2011
- BatesRCEdwardsNSBurnsGFFisherDEA CD44 survival pathway triggers chemoresistance via lyn kinase and phosphoinositide 3-kinase/Akt in colon carcinoma cellsCancer Res2001615275528311431370
- ShenSYangWWangZTumor-initiating cells are enriched in CD44 population in murine salivary gland tumorPLoS One20116e2328221858056
- OuhtitAAbd ElmageedZYAbdrabohMELioeTFRajMHIn vivo evidence for the role of CD44s in promoting breast cancer metastasis to the liverAm J Pathol20071712033203917991717
- Al-HajjMWichaMSBenito-HernandezAMorrisonSJClarkeMFProspective identification of tumorigenic breast cancer cellsProc Natl Acad Sci U S A20031003983398812629218
- FaberABarthCHörmannKCD44 as a stem cell marker in head and neck squamous cell carcinomaOncol Rep20112632163221617876
- ChikamatsuKIshiiHTakahashiGResistance to apoptosis-inducing stimuli in CD44+ head and neck squamous cell carcinoma cellsHead Neck Epub April 5, 2011
- KokkoLLHurmeSMaulaSMSignificance of site-specific prognosis of cancer stem cell marker CD44 in head and neck squamous-cell carcinomaOral Oncol20114751051621514878
- ZhaoJSLiWJGeDTumor initiating cells in esophageal squamous cell carcinomas express high levels of CD44PLoS One20116e2141921731740
- SuJXuXHHuangQIdentification of cancer stem-like CD44+ cells in human nasopharyngeal carcinoma cell lineArch Med Res201142152121376257
- ZhangCLiCHeFCaiYYangHIdentification of CD44+CD24+ gastric cancer stem cellsJ Cancer Res Clin Oncol Epub September 1, 2011
- ChenKLPanFJiangHHighly enriched CD133(+)CD44(+) stem-like cells with CD133(+)CD44 (high) metastatic subset in HCT116 colon cancer cellsClin Exp Metastasis Epub July 13, 2011
- FangXJJiangHZhaoXPJiangWMThe role of a new CD44st in increasing the invasion capability of the human breast cancer cell line MCF-7BMC Cancer20111129021749678
- PhucPVNhanPLNhungTHDownregulation of CD44 reduces doxorubicin resistance of CD44CD24 breast cancer cellsOnco Targets Ther20114717821792314
- PhamPVPhanNLNguyenNTDifferentiation of breast cancer stem cells by knockdown of CD44: promising differentiation therapyJ Transl Med2011920922152097
- AbdrabohMEGaurRLHollenbachADSandquistDRajMHOuhtitASurvivin is a novel target of CD44-promoted breast tumor invasionAm J Pathol201117955556321718681
- SubramaniamVVincentIRGilakjanMJothySSuppression of human colon cancer tumors in nude mice by siRNA CD44 gene therapyExp Mol Pathol20078333234017945212
- LiCZLiuBWenZQLiHYInhibition of CD44 expression by small interfering RNA to suppress the growth and metastasis of ovarian cancer cells in vitro and in vivoFolia Biol (Praha)20085418018619393131
- HuangXShengYGuanMCo-expression of stem cell genes CD133 and CD44 in colorectal cancers with early liver metastasisSurg Oncol Epub July 14, 2011
- RakJMitsuhashiYBaykoLMutant ras oncogenes upregulate VEGF/VPF expression: implications for induction and inhibition of tumor angiogenesisCancer Res199555457545807553632
- Antón AparicioLMCassinello EspinosaJGarcía CampeloRGómez VeigaFDíaz PradoSAparicio GallegoGProstate carcinoma and stem cellsClin Transl Oncol20079667617329217
- EramoALottiFSetteGIdentification and expansion of the tumorigenic lung cancer stem cell populationCell Death Differ20081550451418049477
- GlinskyGVStem cell origin of death-from-cancer phenotypes of human prostate and breast cancersStem Cell Rev20073799317873385
- LiCHeidtDGDalerbaPIdentification of pancreatic cancer stem cellsCancer Res20076710301037200717283135
- PrinceMESivanandanRKaczorowskiAIdentification of a subpopulation of cells with cancer stem cell properties in head and neck squamous cell carcinomaProc Natl Acad Sci U S A200710497397817210912
- SeoDCSungJMChoHJGene expression profiling of cancer stem cell in human lung adenocarcinoma A549 cellsMol Cancer200767518034892
- VorechovskýIBenediktssonKPToftgårdRThe patched/hedgehog/smoothened signalling pathway in human breast cancer: no evidence for H133Y SHH, PTCH, and SMO mutationsEur J Cancer19993571171310505029
- SorianoJVUyttendaeleHKitajewskiJMontesanoRExpression of an activated Notch4(int-3) oncoprotein disrupts morphogenesis and induces an invasive phenotype in mammary epithelial cells in vitroInt J Cancer20008665265910797286
- KellyOGPinsonKISkarnesWCThe Wnt co-receptors Lrp5 and Lrp6 are essential for gastrulation in miceDevelopment20041312803281515142971
- HuelskenJVogelRBrinkmannVErdmannBBirchmeierCBirchmeierWRequirement for beta-catenin in anterior-posterior axis formation in miceJ Cell Biol200014856757810662781
- CattoglioCFacchiniGSartoriDHot spots of retroviral integration in human CD34+ hematopoietic cellsBlood20071101770177817507662
- MontiniECesanaDSchmidtMHematopoietic stem cell gene transfer in a tumor-prone mouse model uncovers low genotoxicity of lentiviral vector integrationNat Biotechnol20062468769616732270
- DeVitaVTJrYoungRCCanellosGPCombination versus single agent chemotherapy: a review of the basis for selection of drug treatment of cancerCancer19753598110162854
- EvansPMAnatomical imaging for radiotherapyPhys Med Biol200853R151R19118495981
- OlivottoMDello SbarbaPEnvironmental restrictions within tumor ecosystems select for a convergent, hypoxia-resistant phenotype of cancer stem cellsCell Cycle2008717618718256528
- BlagosklonnyMVResearch by retrieving experimentsCell Cycle200761277128317525526
- OttewellPDMönkkönenHJonesMLefleyDVColemanREHolenIAntitumor effects of doxorubicin followed by zoledronic acid in a mouse model of breast cancerJ Natl Cancer Inst20081001167117818695136
- XieZChoongPFPoonLFInhibition of CD44 expression in hepatocellular carcinoma cells enhances apoptosis, chemosensitivity, and reduces tumorigenesis and invasionCancer Chemother Pharmacol20086294995718259754
- HenryJCParkJKJiangJmiR-199a-3p targets CD44 and reduces proliferation of CD44 positive hepatocellular carcinoma cell linesBiochem Biophys Res Commun201040312012521055388
- YangKTangYHabermehlGKIczkowskiKAStable alterations of CD44 isoform expression in prostate cancer cells decrease invasion and growth and alter ligand binding and chemosensitivityBMC Cancer2010101620074368
- KrauseDSLazaridesKvon AndrianUHVan EttenRARequirement for CD44 in homing and engraftment of BCR-ABL-expressing leukemic stem cellsNat Med2006121175118016998483
- PontaHShermanLHerrlichPACD44: from adhesion molecules to signaling regulatorsNat Rev Mol Cell Biol20034334512511867
- ShipitsinMCampbellLLArganiPMolecular definition of breast tumor heterogeneityCancer Cell20071125927317349583
- ShiYTianYZhouYQInhibition of malignant activities of nasopharyngeal carcinoma cells with high expression of CD44 by siRNAOncol Rep20071839740317611662
- JodJYShiYZhouYQExperimental siCD44-targeted therapy of human nasopharyngeal carcinoma mediated by adenovirusZhongguo Yi Xue Ke Xue Yuan Xue Bao200729626630 Chinese18051718
- BessisNGarciaCozarFJBoissierMCImmune responses to gene therapy vectors: influence on vector function and effector mechanismsGene Ther200411S10S1715454952
- Hacein-Bey-AbinaSVon KalleCSchmidtMLMO2-associated clonal T cell proliferation in two patients after gene therapy for SCID-X1Science200330241541914564000
- SchroderARShinnPChenHBerryCEckerJRBushmanFHIV-1 integration in the human genome favors active genes and local hotspotsCell200211052152912202041
- MitchellRSBeitzelBFSchroderARRetroviral DNA integration: ASLV, HIV, and MLV show distinct target site preferencesPLoS Biol20042E23415314653
- JakobssonJLundbergCLentiviral vectors for use in the central nervous systemMol Ther20061348449316403676