
Abstract
Background
Papillary thyroid carcinoma (PTC) is the most prevalent type of thyroid cancer. Herein, we purposed to explore the expression patterns of circRNAs in PTC with the overarching goal of improving early diagnosis rates for individuals with PTC.
Methods
We used RNA deep sequencing to determine the expression patterns of circRNAs in PTC. Besides, RT-qPCR was employed to confirm circRNAs. The diagnostic potential of the circRNAs was explored by constructing ROC curves. GO along with KEGG pathway analyses were utilized to elucidate the potential biological roles of differentially expressed circRNAs. Moreover, we predicted cross talks among circRNAs, miRNAs, and mRNAs, followed by establishment of a ceRNA network.
Results
Deep sequencing of four PTC pairs and neighboring nontumor tissues identified 16569 circRNAs, of which, 301 were upregulated and 419 were downregulated. The RT-qPCR data demonstrated that the expression of chr5: 38481299–38530666-, chr2: 159932176–159945082-, chr10: 179994–249088+, chr3: 121378716–121381532+, and chr1: 237423092–237445522+ was downregulated, while the expression of chr4: 25665378–25667298+, chr5: 161330883–161336769-, chr1: 12578718–12579412-, chr7: 116695750–116700284+, and chr7: 116699071–116700284+ was upregulated. The stability test exhibited that circRNAs were more tolerant to temperature, RNase R, and time. On the other hand, ROC curves illustrated that chr4: 25665378–25667298+, chr1: 12578718–12579412-, chr7: 116699071–116700284+, chr7: 116695750–116700284+, chr5: 161330883–161336769-, and chr10: 179994–249088+ were effective as diagnostic indicators. However, a logistic regression model combining the six indicators achieved a better combined prediction index, with 97.7% sensitivity and 95.3% specificity. Moreover, GO along with KEGG pathway analyses illustrated that differentially expressed circRNAs were linked to tumorigenesis. Furthermore, bioinformatics analyses established a promising ceRNAs network among mRNAs, circRNAs, and miRNAs.
Conclusion
Herein, we demonstrated that several circRNAs are promising PTC diagnostic biomarkers. Further study on the functions and mechanisms of these circRNAs may contribute to the understanding of PTC.
Introduction
The incidence rate of thyroid cancer is increasing annually, with the major histopathologic type still being papillary thyroid carcinoma (PTC). Most PTCs have an excellent prognosis, with 10-year survival rates that can reach 95%.Citation1 However, survival is mainly affected by extrathyroid infiltration, lymph node metastasis, and distant metastasis due to advanced disease, which are the main causes of death.Citation2 Currently, the proportion of microcarcinoma detected using ultrasound is rising. Follow-up studies have shown that some thyroid cancer nodules progress slowly and can be observed with dynamic follow-up.Citation3 However, the tumorigenesis of PTC has not yet been fully elucidated, alongside other microcarcinomas which also lead to severe symptoms.Citation4 At the molecular biology level, these microcarcinomas may still have similar characteristics with the large ones. Therefore, this calls for the identification of specific indicators, and design of specific diagnostic and therapeutic regimens that target these features.
Previous studies have identified several genetic indicators associated with thyroid cancer diagnosis and prognosis including BRAF and TERT mutations.Citation5,Citation6 However, it is possible that there are many other ways through which tumor development is regulated at the RNA level in addition to DNA mutations.Citation7 Non coding RNAs belong to an important family of RNAs, and recent investigations have shown that miRNAs and lncRNAs have many modulatory functions in cancer development.
Recently, circRNAs (circular RNAs) have been rediscovered as a novel class of RNA, which initially were recognized as “scrambled” exons in the early 1990s.Citation8 The special circular structure of circRNAs, closed circular sequences, is more stable than linear RNAs, which have 5ʹ and 3ʹ ends.Citation9 Moreover, longer circRNAs may also locally appear as double stranded RNAs in order to increase their stability.Citation10 This stable structure implies a more stable regulatory role in tumorigenesis. Current functional studies on circRNAs are more focused on adsorbing miRNAs as sponges which influence expression of mRNAs.Citation11,Citation12 circRNAs resistance to exonucleases makes them relatively stable in cells, and thus they may constitute ideal biomarkers for cancer diagnosis and can even act as liquid biopsy biomarkers for human diseases.Citation13 Researchers should identify novel early diagnostic biomarkers and elucidate the responsible molecular mechanism of PTC with the overarching goal of improving early detection rates and reducing mortality in advanced PTC patients.
Currently, RNA deep sequencing is progressively replacing microarrays because it has a greater dynamic range relative to microarrays.Citation14 Accumulating research evidence has opined that circRNAs are usually aberrantly expressed in human cancers, and participate in oncogenesis via multiple approaches.Citation15 Herein, we used RNA deep sequencing (RNA-seq) to identify the circRNAs in PTC and verified differentially expressed circRNAs in PTC samples using RT-qPCR. Furthermore, we employed bioinformatics analyses to predict the potential role of circRNAs in PTC. Our data will provide relevant reference for future studies.
Materials and Methods
Patient Tissue Samples and Cell Lines
Here, 99 fresh PTC and neighboring non-tumor tissues were collected from patients treated with thyroid surgery at the First Hospital of China Medical University between 2018 and 2019. All tissue samples were washed with saline and immediately frozen in liquid nitrogen until later use. PTC diagnosis was confirmed by two pathologists postoperative. The TNM (tumor‐node‐metastasis) staging system (8th ed.) was employed to classify tumors. High-throughput sequencing was performed on first four pairs of tissue samples (). The remaining 95 patients were randomly divided into the discovery cohort and the validation cohort according to the ratio of 1:2. ()
Table 1 Clinic Pathological Features of Patients Whose Samples Were Obtained for Sequencing
Table 2 Clinic Pathological Features of Patients Whose Samples Were Obtained for qRT-PCR
The TPC1 cell line was generously provided by Professor Meiping Shen (Department of General Surgery, The First Affiliated Hospital of Nanjing Medical University, Nanjing, Jiangsu).
RNA Extraction and Sequencing
The TriZol reagent (Cat No. 15596018, Life Technologies, Carlsbad, CA, USA) was employed to isolate total RNA from the tissues as described by the manufacturer. RNA quality was checked through determination of 260/280 OD values on a NanoDrop ND-2000 instrument (Thermo Fisher Scientific, Waltham, MA, USA). Total RNAs of four paired PTC and neighboring non-malignant tissues were treated with VAHTS Total RNA-seq (H/M/R) Library Prep Kit for Illumina as described in the manufacturer provided manual. Subsequently, sequencing of the products was run on the Illumina HiSeqTM 2500 platform by Gene Denovo Biotechnology Co. (Guangzhou, China). The processed clean reads were then aligned to the human genome (version: hg38_GRCh38).
RT-qPCR Validation
The remaining 95 sample pairs were employed for confirmation via RT-qPCR. Concisely, the TriZol reagent (Cat No. 15596018, Life Technologies, Carlsbad, CA, USA) was employed to isolate total RNA from the tissues as described by the manufacturer. After that, cDNA was regenerated from the total RNA by using PrimeScript RT Master Mix (Cat No. RR036A, Perfect Real Time, TaKaRa) with random primers. Thereafter, SYBR Premix Ex Taq II (Cat No. RR820A, Tli RHaseH Plus, TaKaRa) was employed to prepare a qPCR reaction mixture and the amplification was run on the LightCycler 480 system (Roche, Basel, Switzerland). We used the following RT-qPCR protocol: denaturation at 95°C for 30 seconds, followed by 50 cycles of: 95°C for 5 seconds and 60°C for 30 seconds. GAPDH served as the normalization standard. The ΔCt method was employed to assess relative RNA expression. Primers for RT-qPCR analysis were designed to target the circRNA specificity junction site, and the specificity of primers was tested using CircPrimer v1.2.0.5.Citation16 The sequences of all primers utilized in RT-qPCR assays as . The diagnostic potential of the circRNAs for PTC patients was explored through the construction of ROC (Receiver operating characteristic) curves.
Table 3 PCR Primer Sequences for circRNAs
Gene Ontology and Kyoto Encyclopedia of Genes and Genomes Pathway Analysis
Paired-end reads were harvested from an Illumina HiSeqTM2500 sequencer and Bowtie2 v2.4.2Citation17 was employed to map the reads to the ribosome RNA (rRNA) data resource. Next, high-quality reads were aligned to the reference genome using Hisat2 v2.2.1.Citation18 We then collected the unmapped reads for circRNA identification using Find_circ v1,Citation19 followed by blasting against circBaseCitation20 for annotation. The circRNAs that could not be annotated were defined as novel circRNAs. Moreover, circRNAs that exhibited fold changes ≥ 2 along with adjusted P< 0.05 were considered to be remarkably expressed differentially.
GO along with KEGG pathway analyses were then employed to identify the source genes for the differentially expressed circRNAs.
circRNA-miRNA-mRNA Network Construction
We predicted the miRNAs that might target cirRNAs. For circRNAs that have been annotated in circBase, we used StarBase (v2.0) to determine the target relationship with miRNAs. On the other hand, we used three softwares; Mireap, Miranda (v3.3a), as well as TargetScan (Version:7.0) for novel circRNAs. miRTarBase v8.0Citation21 was employed to predict circRNAs/mRNAs targeted by the miRNAs sponge and the resulting correlation of circRNAs-miRNAs-mRNAs was visualized using Cytoscape v3.7.2.Citation22
Statistical Analyses
Statistical analyses were implemented in the GraphPad Prism 6 software (La Jolla, CA) and SPSS 19.0 (IBM, Chicago, IL, USA). P<0.05 signified statistical significance. The Student’s t-test was employed to explore the differences between groups where appropriate.
Compliance with Ethical Standards
We obtained written informed consent for the experimental use of surgical tissue samples from all participants, and the study was approved by the ethics committee of the First Hospital of China Medical University, Shenyang, China.
Results
Overview of circRNA Profiles in PTC
High-throughput RNA sequencing was performed in four pairs of PTC and neighboring nontumor tissues. In total, 16569 circRNAs were identified (), with most of them coming from exons (). For consistency, we named these circRNAs according to the location of their source chromosome.
Figure 1 (A) Distribution of identified circRNAs on the chromosome; (B) Type of circRNAs; (C) Hierarchical cluster analysis of differentially expressed circRNAs; (D) Volcano plot of differential expressed circRNAs in PTC.
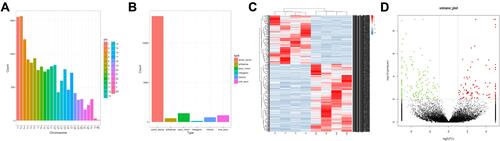
On the basis of the screening criteria (adjusted P values ≤ 0.05 along with fold change ≥ 2), we screened out 301 upregulated and 419 downregulated circRNAs. shows the heat-map of all differentially expressed circRNAs, while summarizes the genetic information of the top 20 differentially expressed circRNAs. Moreover, the volcano plot exhibits the statistical significance of differentially expressed circRNAs between PTC and neighboring nontumor tissues ().
Table 4 Top 20 Up-Regulated and Down-Regulated Expressed circRNAs
Verification of circRNA Expression
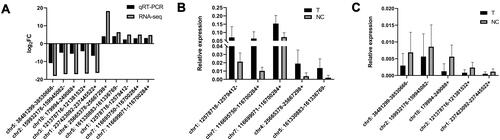
The ten most differentially expressed circRNAs were selected to verify the sequencing quality in the discovery data set using qRT-PCR. Data illustrated that the expression of chr5: 38481299–38530666-, chr2: 159932176–159945082-, chr10: 179994–249088+, chr3: 121378716–121381532+, and chr1: 237423092–237445522+ was downregulated, while the expression of chr4: 25665378–25667298+, chr5: 161330883–161336769-, chr1: 12578718–12579412-, chr7: 116695750–116700284+, and chr7: 116699071–116700284+ was upregulated (–).
Figure 2 Ten circRNAs were selected to verify the sequencing quality by qRT-PCR. (A) Comparison of log2 fold changes in circRNAs between RNA-Seq and qRT-PCR results; (B and C) The relative expression levels of circRNAs in 32 PTC tissues and paired normal thyroid tissues.
Diagnostic Significance of RT-qPCR Verified circRNAs
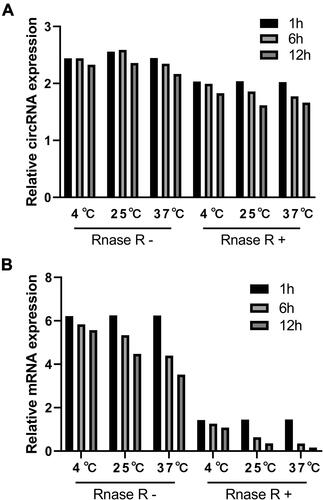
To further explore the diagnostic capacity of circRNAs, we chose chr7: 116699071–116700284+ which had the highest expression in PTC. After quantification, we extracted fresh RNA from TPC1 cell line and divided it into six samples. Next, three samples were inoculated with RNase R according to the manufacturer’s instructions, and placed in an enclosed environment without RNA enzyme pollution at 4°C, 25°C, and 37°C, respectively. After 1 h, 6 h, and 12 h, the same volume of sample RNA was taken for reverse transcription with random primers. The expression levels of circRNA and source gene mRNA were then determined using qRT-PCR. Results indicated that circRNAs were more stable than mRNAs under the influence of temperature, RNase R, and time ().
Figure 3 The stability of circRNAs (A) and its source mRNA (B) in cell line TPC1.
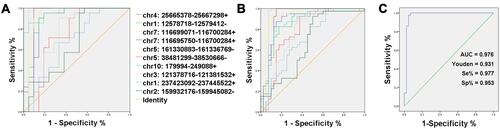
We then developed ROC curves for RT-qPCR verified circRNAs using discovery data set (n = 32, ) and verification data set (n = 63, ). Our results indicated that the ten circRNAs all showed potential diagnostic capabilities for diagnosing PTC, with consistent areas under the curve (AUC) in both the verification and discovery data sets ().
Table 5 Area Under the Curve (AUC) of qRT-PCR Verified circRNAs in the Discovery and Validation Cohort
Figure 4 ROC curves qRT-PCR verified circRNAs: (A) Discovery cohort (n = 32); (B) Validation cohort (n = 63); (C) The ROC of Combined Prediction Index.
Furthermore, a combined prediction index was established based on a logistic regression model which combined the expression levels of the most efficient circRNAs (AUC > 0.8, sensitivity > 80%, and specificity > 80%). The coefficients in the prediction equation were as follows: Combined Prediction Index =−0.321× (chr4: 25665378–25667298+)+0.23×(chr1: 12578718–12579412-) −2.818× (chr7: 116699071–116700284+)-1.078× (chr7: 116695750–116700284+)+0.689× (chr5: 161330883–161336769-)−0.723× (chr10: 179994–249088+)+15.627.
The results showed that the performance of AUC, sensitivity, and specificity was greater when the Combined Prediction Index was used as a diagnostic marker in PTC. Moreover, the Combined Prediction Index showed the best diagnosis efficiency compared to the single markers ().
GO Enrichment Analysis and Pathway Enrichment Analysis
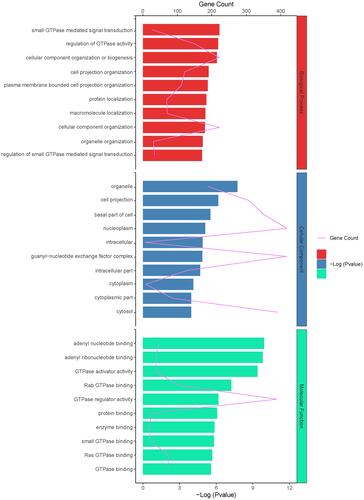
GO annotation reveals the source genes of the differentially expressed circRNAs. The most dramatically enriched GO terms with regard to the biological process, cellular component, and molecular function categories were modulation of small GTPase mediated signal transduction (GO: 0051056, gene count = 27, P = 5.75E-07), organelle (GO: 0043226, gene count = 392, P = 1.14E-04), and adenyl nucleotide-binding (GO: 0030554, gene count = 73, P = 2.72E-06), respectively ().
Figure 5 GO terms analysis for differential expressed host gene of circRNAs.
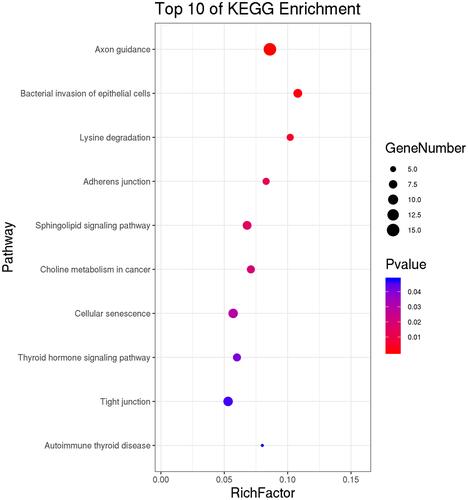
On the other hand, KEGG pathway analysis demonstrated that the differentially expressed circRNA genes were primarily associated with axon guidance, lysine degradation, bacterial invasion of epithelial cells, adherens junction, sphingolipid signaling cascade, choline metabolism in cancer, cellular senescence, thyroid hormone signaling pathway, tight junction, and autoimmune thyroid disease ().
Figure 6 KEGG pathways analysis for differential expressed host gene of circRNAs.
circRNA-miRNA-mRNA Network Construction
In addition, miRNAs sponge has been widely studied in order to determine the function of circRNAs. The top five miRNAs were selected for each circRNAs, the top ten mRNAs were selected for each miRNAs. We constructed the predicted circRNA-miRNA-mRNA network for the top three upregulated and downregulated circRNAs in PTC ().
Figure 7 The potential competing endogenous RNA (ceRNA) relationships in PTC. Yellow represents circRNAs, green represents miRNAs, and blue represents mRNAs.

Discussion
Circular RNA is a class of endogenous RNA that widely exists in eukaryotes,Citation23 and was once considered to be the product of wrong splicing. However, with the development of research, the characteristics and functions of circular RNA are becoming more and more clear.
CircRNAs may arise from exons or introns of pre‐mRNA after back‐splicing.Citation24 Interestingly, they regulate gene expression and biological processes, serve as miRNA sponge, regulate transcription, are involved in rolling circle translation, generate pseudogenes, and affect alternative splicing.Citation25 The progress of high-throughput sequencing technology in recent years has resulted in identification of several circRNAs. Accumulating evidence has opined that circRNAs play a core role in the onset of cancers and may be applied as novel biomarkers.Citation26–Citation29 It is worth noting that they are few studies on thyroid cancer, and thus it is essential to explore the expression of circRNA in PTC.
Herein, we chose ten differentially expressed circRNAs for RT-qPCR verification. The obtained results were consistent with the reliability of RNA-Seq. chr4: 25665378–25667298+ was the most up-regulated circRNA, while chr5: 38481299–38530666 was the most down-regulated circRNA in PTC. We then tested the stability of circRNA and its source gene mRNA in PTC cell line. Results indicated that circRNAs can tolerate the effects of RNase R treatment and temperature stimulation, and time extension demonstrated that the stability of circRNAs was stronger than that of linear RNAs. Next, we plotted ROC curves using the data verified by RT-qPCR as the discovery data set. ROC curves illustrated that the verified circRNAs are promising PTC diagnostic biomarkers. Moreover, we validated the ROC curves using 63 newly paired samples of cancer and neighboring nontumor tissues. The data obtained from the verification data set also showed that the circRNAs could stably predict PTC. To achieve better diagnostic ability, we constructed a Combined Prediction Index using a logistic regression model which combined the six most efficient circRNAs. Results indicated that the Combined Prediction Index achieved the highest AUC, sensitivity, and specificity compared to the original six indicators. With regard to verification of stability, we believe that the above-mentioned circRNAs can all be used as new biomarkers for PTC identification. However, combined use of multiple circRNAs has the highest diagnostic efficiency.
CircRNAs are derived from alternative splicing of RNA. Most of the circRNAs sequences are similar to their parent genes and their functions may also be similar. Moreover, a previous research documented that circRNAs can modulate the transcription of parental genes.Citation30 To further explore the role of circRNAs, we identified differentially expressed circRNAs among groups and analyzed the function of source genes through GO along with KEGG pathway analyses. Our results illustrated that differentially expressed circRNAs were associated with tumorigenesis including adherens junction, cellular senescence, and tight junction. The most commonly reported function of circRNAs is their action as miRNA sponges.Citation31 One study reported that circRNAs are involved in the pathogenesis of PTC through modulating miRNAs and their target mRNA.Citation32 Herein, we predicted the possible miRNA targets of circRNAs and developed a circRNA-miRNA-mRNA network for determining the regulatory role of circRNAs in PTC. Our network indicated that some miRNAs have been identified in other tumors. For instance, hsa-miR-1299 regulates breast cancer,Citation33 prostate cancer,Citation34 hepatocellular carcinoma,Citation35 and cholangiocarcinoma.Citation36 In addition, Hsa-mir-22-3p regulates lung adenocarcinoma progression,Citation37 while Wnt9A activation is related to repression of human colorectal cancer cell proliferation.Citation38 Furthermore, FOXA2 suppresses the metastasis of hepatocellular carcinoma,Citation39 and ZEB1 activation induces epithelial-mesenchymal transition.Citation40 Therefore, future research should focus on these miRNAs and mRNAs identified in our network.
However, this study has some limitations. Firstly, we only selected four pairs of cancer tissues and nontumor tissues for RNA sequence analysis, which may lead to bias. Secondly, there is a long way to go before circRNAs can be used in the diagnosis of PTC.
In conclusion, this study has exhibited the circRNA expression profiles in PTC. To the best of our knowledge, this is the first study that has shown that chr4: 25665378–25667298+, chr1: 12578718–12579412-, chr7: 116699071–116700284+, chr7: 116695750–116700284+, chr5: 161330883–161336769-, and chr10: 179994–249088+ may be potential biomarkers for the diagnosis of PTC. Further studies on the functions and mechanisms of these circRNAs will contribute to our understanding of PTC tumorigenesis.
Data Sharing Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: the NCBI Gene Expression Omnibus (GSE171011).
Ethics Statement
The study protocol was approved by the Clinical Research Ethics Committee of The First Hospital of China Medical University. Written informed consent was obtained from all participants. The acquisition of tissue specimens was conducted in accordance with the Declaration of Helsinki.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Additional information
Funding
References
- Haugen BR, Alexander EK, Bible KC, et al. 2015 American thyroid association management guidelines for adult patients with thyroid nodules and differentiated thyroid cancer: the American thyroid association guidelines task force on thyroid nodules and differentiated thyroid cancer. Thyroid. 2016;26(1):1–133. doi:10.1089/thy.2015.0020
- Khan ZF, Kutlu O, Picado O, Lew JI. Margin positivity and survival outcomes: a review of 14,471 patients with 1-cm to 4-cm papillary thyroid carcinoma. J Am Coll Surg. 2021;232(4):545–550. doi:10.1016/j.jamcollsurg.2020.12.018
- Ito Y, Miyauchi A. Active surveillance as first-line management of papillary microcarcinoma. Annu Rev Med. 2019;70:369–379. doi:10.1146/annurev-med-051517-125510
- Sheng L, Shi J, Han B, et al. Predicting factors for central or lateral lymph node metastasis in conventional papillary thyroid microcarcinoma. Am J Surg. 2020;220(2):334–340. doi:10.1016/j.amjsurg.2019.11.032
- Xing M, Liu R, Liu X, et al. BRAF V600E and TERT promoter mutations cooperatively identify the most aggressive papillary thyroid cancer with highest recurrence. J Clin Oncol. 2014;32(25):2718–2726. doi:10.1200/JCO.2014.55.5094
- Liu R, Xing M. TERT promoter mutations in thyroid cancer. Endocr Relat Cancer. 2016;23(3):R143–R155. doi:10.1530/ERC-15-0533
- Anastasiadou E, Jacob LS, Slack FJ. Non-coding RNA networks in cancer. Nat Rev Cancer. 2018;18(1):5–18. doi:10.1038/nrc.2017.99
- Nigro JM, Cho KR, Fearon ER, et al. Scrambled exons. Cell. 1991;64(3):607–613. doi:10.1016/0092-8674(91)90244-s
- Suzuki H, Tsukahara T. A view of pre-mRNA splicing from RNase R resistant RNAs. Int J Mol Sci. 2014;15(6):9331–9342. doi:10.3390/ijms15069331
- Sun H, Wu Z, Liu M, et al. CircRNA may not be “circular”. bioRxiv. 2020.
- Ye M, Hou H, Shen M, Dong S, Zhang T. Circular RNA circFOXM1 plays a role in papillary thyroid carcinoma by sponging miR-1179 and regulating HMGB1 expression. Mol Ther Nucleic Acids. 2020;19:741–750. doi:10.1016/j.omtn.2019.12.014
- Li S, Yang J, Liu X, Guo R, Zhang R. circITGA7 functions as an oncogene by sponging miR-198 and upregulating FGFR1 expression in thyroid cancer. Biomed Res Int. 2020;2020:8084028. doi:10.1155/2020/8084028
- Wen G, Zhou T, Gu W. The potential of using blood circular RNA as liquid biopsy biomarker for human diseases. Protein Cell. 2020. doi:10.1007/s13238-020-00799-3
- Hitzemann R, Bottomly D, Darakjian P, et al. Genes, behavior and next-generation RNA sequencing. Genes Brain Behav. 2013;12(1):1–12. doi:10.1111/gbb.12007
- Hansen TB, Jensen TI, Clausen BH, et al. Natural RNA circles function as efficient microRNA sponges. Nature. 2013;495(7441):384–388. doi:10.1038/nature11993
- Zhong S, Wang J, Zhang Q, Xu H, Feng J. CircPrimer: a software for annotating circRNAs and determining the specificity of circRNA primers. BMC Bioinform. 2018;19(1):292. doi:10.1186/s12859-018-2304-1
- Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nat Methods. 2012;9(4):357–359. doi:10.1038/nmeth.1923
- Kim D, Paggi JM, Park C, Bennett C, Salzberg SL. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat Biotechnol. 2019;37(8):907–915. doi:10.1038/s41587-019-0201-4
- Memczak S, Jens M, Elefsinioti A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013;495(7441):333–338. doi:10.1038/nature11928
- Glazar P, Papavasileiou P, Rajewsky N. circBase: a database for circular RNAs. RNA. 2014;20(11):1666–1670. doi:10.1261/rna.043687.113
- Huang HY, Lin YC, Li J, et al. miRTarBase 2020: updates to the experimentally validated microRNA-target interaction database. Nucleic Acids Res. 2020;48(D1):D148–D154. doi:10.1093/nar/gkz896
- Shannon P, Markiel A, Ozier O, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13(11):2498–2504. doi:10.1101/gr.1239303
- Broadbent KM, Broadbent JC, Ribacke U, Wirth D, Rinn JL, Sabeti PC. Strand-specific RNA sequencing in Plasmodium falciparum malaria identifies developmentally regulated long non-coding RNA and circular RNA. BMC Genomics. 2015;16:454. doi:10.1186/s12864-015-1603-4
- Fu L, Jiang Z, Li T, Hu Y, Guo J. Circular RNAs in hepatocellular carcinoma: functions and implications. Cancer Med. 2018;7(7):3101–3109. doi:10.1002/cam4.1574
- Yao T, Chen Q, Fu L, Guo J. Circular RNAs: biogenesis, properties, roles, and their relationships with liver diseases. Hepatol Res. 2017;47(6):497–504. doi:10.1111/hepr.12871
- Ahmed I, Karedath T, Andrews SS, et al. Altered expression pattern of circular RNAs in primary and metastatic sites of epithelial ovarian carcinoma. Oncotarget. 2016;7(24):36366–36381. doi:10.18632/oncotarget.8917
- Li F, Zhang L, Li W, et al. Circular RNA ITCH has inhibitory effect on ESCC by suppressing the Wnt/β-catenin pathway. Oncotarget. 2015;6(8):6001–6013. doi:10.18632/oncotarget.3469
- Wang X, Zhang Y, Huang L, et al. Decreased expression of hsa_circ_001988 in colorectal cancer and its clinical significances. Int J Clin Exp Pathol. 2015;8(12):16020–16025.
- Qu S, Song W, Yang X, et al. Microarray expression profile of circular RNAs in human pancreatic ductal adenocarcinoma. Genom Data. 2015;5:385–387. doi:10.1016/j.gdata.2015.07.017
- Holdt LM, Kohlmaier A, Teupser D. Molecular functions and specific roles of circRNAs in the cardiovascular system. Noncoding RNA Res. 2018;3(2):75–98. doi:10.1016/j.ncrna.2018.05.002
- Zheng Q, Bao C, Guo W, et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat Commun. 2016;7:11215. doi:10.1038/ncomms11215
- Xin Z, Ma Q, Ren S, Wang G, Li F. The understanding of circular RNAs as special triggers in carcinogenesis. Brief Funct Genomics. 2017;16(2):80–86. doi:10.1093/bfgp/elw001
- Liu LH, Tian QQ, Liu J, Zhou Y, Yong H. Upregulation of hsa_circ_0136666 contributes to breast cancer progression by sponging miR-1299 and targeting CDK6. J Cell Biochem. 2019;120(8):12684–12693. doi:10.1002/jcb.28536
- Zhang FB, Du Y, Tian Y, Ji ZG, Yang PQ. MiR-1299 functions as a tumor suppressor to inhibit the proliferation and metastasis of prostate cancer by targeting NEK2. Eur Rev Med Pharmacol Sci. 2019;23(2):530–538. doi:10.26355/eurrev_201901_16865
- Zhu H, Wang G, Zhou X, et al. miR-1299 suppresses cell proliferation of hepatocellular carcinoma (HCC) by targeting CDK6. Biomed Pharmacother. 2016;83:792–797. doi:10.1016/j.biopha.2016.07.037
- Xu Y, Yao Y, Liu Y, et al. Elevation of circular RNA circ_0005230 facilitates cell growth and metastasis via sponging miR-1238 and miR-1299 in cholangiocarcinoma. Aging. 2019;11(7):1907–1917. doi:10.18632/aging.101872
- Dong HX, Wang R, Jin XY, Zeng J, Pan J. LncRNA DGCR5 promotes lung adenocarcinoma (LUAD) progression via inhibiting hsa-mir-22-3p. J Cell Physiol. 2018;233(5):4126–4136. doi:10.1002/jcp.26215
- Ali I, Medegan B, Braun DP. Wnt9A induction linked to suppression of human colorectal cancer cell proliferation. Int J Mol Sci. 2016;17(4):495. doi:10.3390/ijms17040495
- Wang J, Zhu CP, Hu PF, et al. FOXA2 suppresses the metastasis of hepatocellular carcinoma partially through matrix metalloproteinase-9 inhibition. Carcinogenesis. 2014;35(11):2576–2583. doi:10.1093/carcin/bgu180
- Meng X, Kong DH, Li N, et al. Knockdown of BAG3 induces epithelial-mesenchymal transition in thyroid cancer cells through ZEB1 activation. Cell Death Dis. 2014;5:e1092. doi:10.1038/cddis.2014.32