
Abstract
Background
Brahma-related gene 1 (BRG1) has been implicated in a variety of biological processes, and it has been found to be mutated or silenced in numerous cancers, including lung cancer. Recent reports have proposed BRG1 as a tumor suppressor, but its roles in cell proliferation and metastasis remain unknown. miR-148b functions as a tumor suppressor in non-small-cell lung cancer. However, the mechanism responsible for the downregulation of miR-148b in lung cancer is still elusive.
Methods
The expression of BRG1 and miR-148b was evaluated in lung cancer tissues and cells using quantitative real-time polymerase chain reaction. The effect of BRG1 on proliferation of lung cancer cells was investigated using MTT assay. Transwell and Western blot assays were used to analyze the effect of BRG1 on invasion and epithelial–mesenchymal transition (EMT), respectively. The target of miR-148b was ascertained using luciferase reporter assay. Chromatin immunoprecipitation (ChIP) assay was performed to analyze the relation of BRG1 and the promoter region of miR-148b.
Results
Restoration of BRG1 was demonstrated to inhibit cell proliferation, metastasis, and EMT in lung cancer cell lines. Furthermore, we found that miR-148b was positively regulated by BRG1. Additionally, we suggested that miR-148b suppressed cell proliferation, metastasis, and EMT in lung cancer cells by directly binging to 3′-untranslated region of WNT1, blocking the WNT1/β-catenin signaling pathway. ChIP assay showed that BRG1 bound to the promoter of miR-148b in A549 cells.
Conclusion
BRG1 positively regulated the expression of miR-148b, leading to inhibition of cell proliferation, metastasis, restraint of EMT, and inactivation of the WNT/β-catenin signaling pathway, which highlights potential therapeutic possibilities for the treatment of lung cancer.
Introduction
Mammalian switch/sucrose nonfermentable (SWI/SNF) chromatin remodeling complexes contain either BRG1 or BRM exclusively as the catalytic ATPase subunit that control the alteration of DNA–nucleosome structure and thus modulate target gene transcription.Citation1,Citation2 The loss of BRG1 subunit disrupts the complex and inactivates its function, which is associated with a variety of malignancies, including ovarian, renal, liver, and pediatric cancers.Citation3,Citation4 Brahma-related gene 1 (BRG1) was the fourth most frequently altered gene in non-small-cell lung cancer (NSCLC) cell lines and low BRG1 protein level have also been observed in clinical lung primary tumors.Citation5–Citation7 Restoration of wild-type BRG1 expression activated massive transcription in lung cancer cells and significantly inhibited invasion and growth of lung cancer cells orthotopically implanted in nude mice.Citation6,Citation8 However, the regulatory mechanism continues to be obscure.
MicroRNAs (miRNAs) are a class of single-stranded, small noncoding RNAs measuring 18–25 nucleotides in length that guide the cleavage of their target messenger RNAs (mRNAs) or bind to 3′-untranslated region (3′-UTR) of their recognition sequences of mRNA, causing either mRNA degradation or translational repression, thus effectively silencing their mRNA targets. Each miRNA potentially regulates numerous target mRNAs, and it is predicted that more than one-third of protein-coding genes are regulated by miRNAs.Citation9,Citation10 Recently, miRNAs have emerged as posttranscriptional regulators that are implicated in multiple cell biological processes, including cell differentiation, proliferation, metastasis, apoptosis, metabolism, tumorigenesis, and cancer progression.Citation11,Citation12 There is accumulating evidence that miRNAs show aberrant expressions in cancers and modulate cancer cell behaviors.Citation13,Citation14
miR-148b is frequently downregulated in several types of cancers, such as pancreatic cancer,Citation15 gastric cancer,Citation16 breast cancer,Citation17 esophageal squamous cell carcinoma,Citation18 and colorectal cancer.Citation19 miR-148b functions as a new tumor suppressor via directly targeting specific protein-coding genes in these cancers. Moreover, miR-148b increases the radiosensitivity of non-Hodgkin’s lymphoma cells probably by enhancing ionizing radiation-induced apoptosis.Citation20 In pancreatic cancer cell lines, upregulation of miR-148b returns DNA methylation to normal patterns and promotes re-expression of tumor suppressor genes, like BNIP3 and SPARC, through suppression of DNA methyltransferase-1 (DNMT-1) expression.Citation21 miR-148b also plays a vital role in maintenance of a side population cells with cancer stem cell properties through regulating Neuropilin-1 in hepatocellular cancer.Citation22 The latest report showed that miR-148b was downregulated in NSCLC, and aberrant miR-148b expression was significantly associated with the clinicopathological characteristics, including TNM stage, lymph node metastasis, distant metastasis, and worse overall survival rates.Citation23 Liu et alCitation24 found that miR-148b was downregulated in NSCLC cells, which resulted in carcinoembryonic antigen overexpression and disease progression in patients with NSCLC. These data show that miR-148b plays a critical role in regulating these physiological events, but the underlying mechanism accounting for the downregulation of miR-148b in human cancers, especially in lung cancer, remains unclear.
In this study, we confirmed that BRG1 and miR-148b were expressed at low levels in lung cancer tissues and cells and observed the correlation between BRG1 and miR-148b. Furthermore, we investigated the effects of restoration of BRG1 on the proliferation and metastasis of lung cancer cells and the potential mechanisms for lung cancer behavior affected by BRG1/miR-148b. These findings will help to better understand the roles of BRG1 and miR-148b in the progression of lung cancer.
Materials and methods
Clinical specimens
Surgically resected lung cancer tissues and matched adjacent normal lung tissues were collected from 29 lung cancer patients during operation at the Second Affiliated Hospital of Zhengzhou University. Exclusion criteria were presence of chemotherapy or radiotherapy prior to lung resection. All specimens were stored at −80°C until analysis. Written informed consent for the studies was obtained from all participants and their guardians. This study was approved by the Ethics Committee of the Second Affiliated Hospital of Zhengzhou University.
Cell culture and treatment
Human lung cancer cell lines and lung cell lines were obtained from the American Type Culture Collection (ATCC, Rock-ville, MD, USA). A549, H522, and H1299 were grown in Roswell Park Memorial Institute (RPMI) 1640 with 10% fetal calf serum, 5 mM L-glutamine, 100 U/mL penicillin, and 100 µg/mL streptomycin, and maintained in a humidified incubator at 37°C containing 5% CO2. The normal lung cell lines (WI38 and MRC5) and the human bronchial epithelial cell line (Beas-2B) were cultured as described previously.Citation25 For evaluation of epithelial–mesenchymal transition (EMT), cells were treated with 4 ng/mL of TGFβ1 (R&D Systems, Minneapolis, MN, USA) for 48 hours. Then the relative expression levels of E-cadherin, N-cadherin, and vimentin in cells were detected by using Western blot analysis.
Quantitative real-time PCR
Total mRNA from tissues and cells was extracted using Unizol Reagent (Biostar, Shanghai, People’s Republic of China) according to the manufacturer’s instructions. Total RNAs were reverse transcribed into cDNA using a High Capacity cDNA Archive Kit (Applied Biosystems, Foster City, CA, USA). Expression level of miR-148b was quantified using a miRNA-specific TaqMan MiRNA Assay Kit (Applied Biosystems) according to the manufacturer’s instructions. For the amplification of BRG1 and GAPDH (glyceraldehyde 3-phosphate dehydrogenase) mRNA, the primers were synthesized as follows: BRG1 (forward: 5′-CAT CAT CGT GCC TCT CTC AAC-3′; reverse: 5′-ACA CGC ACC TCG TTC TGC TG-3′) and GAPDH (forward: 5′-CAC GAT GGA GGG GCC GGA CTC ATC-3′; reverse: 5′-TAA AGA CCT CTA TGC CAA CAC AGT-3′). Relative levels of mRNA (amount of target gene normalized to GAPDH) were calculated using the comparative Ct method formula 2−∆∆Ct.
Chromatin immunoprecipitation assay
For the chromatin immunoprecipitation (ChIP) assay, chromatin was prepared from A549 cells. A549 cells were treated with 10% formaldehyde for 10 minutes at room temperature, washed with phosphate buffered saline (PBS), and resuspended in lysis buffer (50 mM Tris-HCl pH 8.0, 40 mM ethylenediaminetetraacetic acid (EDTA), 0.5% SDS). Chromatin was prepared according to standard protocols and sheared to fragments in a Bioruptor (Diagenode, Philadelphia, PA, USA) by three sonication cycles of 5 minutes. The sheared chromatin was then immunoprecipitated by using anti-BRG1 antibody (Abcam, Cambridge, UK) or control IgG (Sigma, St. Louis, MO, USA). After extensive washing and elution, crosslinks were reverted by adjusting NaCl concentrations and heat treatment. Precipitated DNA fragments were purified by using PCR purification kit (Qiagen, Valencia, CA, USA). Real-time PCR analysis was performed for BRG1 binding sites in miR-148b promoter by using the following primers: 5′-AGC GCC AGT GTT AAA GGC TA-3′ (forward) and 5′-TCC ATG GGG AAC AGA AGA AG-3′ (reverse).
Western blot analysis
Protein extraction and Western blot analysis were performed. Briefly, proteins were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to nitrocellulose membranes. After blocking, the filters were incubated with BRG1 (Abcam), Ki67 (Abcam), WNT1 (Cell Signaling Technology, Beverly, MA, USA), β-catenin (Abcam), c-myc (Cell Signaling Technology), E-cadherin (Santa Cruz Biotech, Santa Cruz, CA, USA), N-cadherin (Santa Cruz Biotech), vimentin (Santa Cruz Biotech), and GAPDH (Sigma, USA) antibodies at 4°C overnight. After washing and incubation with horseradish peroxidase-conjugated secondary antibodies for 1 hour, blots were visualized by using the Odyssey Infrared Imaging System (LI-COR Biosciences, Lincoln, NE, USA). Detection of GAPDH on the same membrane was used as a loading control.
Transfection
A549 and H522 cells (1×105 per well) were maintained in six-well plates and incubated for 24 hours. Cells were transfected with 50 nM of miR-148b mimics or control mimics (miR-control), 100 nM miR-148b inhibitors (anti-148b, with sequence of 5′-ACA AAG UUC UGU GAU GCA CUG A-3′) or control inhibitors (anti-ctrl), and 100 nM specific siRNA of the human BRG1 gene or control siRNA (si-control) with lipofectamine 2000 Reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s protocol. Transfection efficiency was evaluated by qRT-PCR or Western blot analysis. A549 cells were transfected with a retrovirus encoding shRNA sequence specific for WNT1 (shRNA-WNT1) or negative control sequence (shRNA-control). Approximately 10 µg of BRG1 expression plasmids (pcDNA3.1-3× Flag-BRG1) or empty vectors were transfected into A549 and H522 cells with Lipofectamine Plus (Invitrogen), according to the manufacturer’s instructions.
MTT assay
Following transfection with BRG1 vector (or cotransfection with BRG1 or control vector and anti-miR-148b mimics or anti-miR-control), MTT assay was used to assess relative cell viability of A549 and H522 cells, as described previously.Citation26 The absorbance values were determined at a wavelength of 490 nm using a microplate spectrophotometer (Molecular Devices, Sunnyvale, CA, USA).
Cell migration assay
Cell migration was measured by using the Transwell assay. Briefly, A549 and H522 cells (2×104) transfected with vector containing BRG1 or control (or cotransfected with vector containing BRG1 or control and anti-miR-148b mimics or anti-miR-control) were seeded in serum-free medium in top chambers (BD Bioscience, Franklin Lakes, NJ, USA), whereas in the lower compartments Dulbecco’s Modified Eagle’s Medium containing 5% fetal bovine serum was added. After 20 hours of incubation, the nonmigrated cells in the top chambers were removed by a cotton swab. The number of A549 cells adhering to the lower membranes was counted after cells were fixed and stained with crystal violet.
Luciferase activity assay
A549 cells (2×104) in a 48-well plate were cotransfected with 10 nM miR-148b mimics or mimic control, 2 ng pRL-TK (Promega, Madison, WI, USA). Approximately 10 ng firefly luciferase reporter containing the wild-type or mutant 3′-UTR of WNT1 pRL-TK, which expresses Renilla luciferase, was used as the internal control in the dual-luciferase assay. Forty-eight hours after transfection, the luciferase activities were measured using a Dual-Luciferase Reporter Assay System (Promega) according to the manufacturer’s protocol.
Statistics analysis
The data were expressed as the mean ± standard deviation from at least three independent experiments. Statistical significance between two groups was evaluated using Student’s t-test. All statistical tests were two sided, and a P-value <0.05 was considered to be statistically significant.
Results
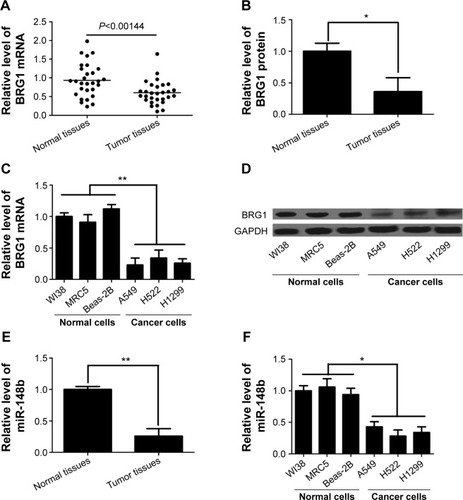
BRG1 and miR-148b levels are significantly lower in lung cancer tissues and cell lines
It has been reported that dysregulated expression of BRG1 is frequently present in lung cancer and that tumor progression and poor prognosis are greatly affected by downregulation of BRG1.Citation27,Citation28 Here, we confirmed the levels of BRG1 in lung cancer tissues and cell lines by using qRT-PCR and Western blot analysis. As shown in , the expression levels of BRG1 mRNA and protein in cancer lung tissues were obviously lower than those in matched adjacent normal lung tissues. In addition, lung cancer cell lines (A549, H522, and H1299) showed reduced BRG1 mRNA and protein levels compared with normal lung cells (WI38, MRC5, and Beas-2B; ). Low expression of miR-148b has been found in NSCLC, and miR-148b functions as a tumor suppressor by directly targeting specific oncogenes.Citation24 Here, we confirmed that the expression of miR-148b was decreased in lung cancer tissues and cell lines (). Collectively, these cumulative data verify that lower BRG1 and miR-148b expression levels frequently occur in human lung cancer.
Figure 1 BRG1 and miR-148b are expressed at low levels in lung cancer tissues and cell lines.
Abbreviations: BRG1, Brahma-related gene 1; mRNA, messenger RNA; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
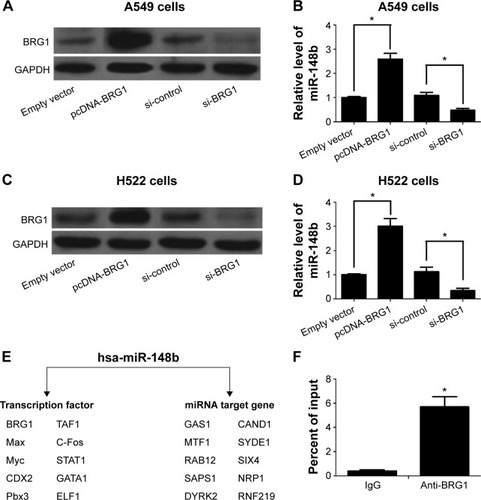
BRG1 binds to miR-148b target gene promoter in lung cancer cells
We further found that the expression of miR-148b correlates with the levels of BRG1 in A549 and H522 cells. As shown in , overexpression of BRG1 elevated the expression of miR-148b in A549 and H522 cells. In contrast, the expression levels of miR-148b were decreased when BRG1 was knocked down. The online platform ChIPBase shows that BRG1 is a predicted transcription factor of miR-148b (). To determine whether BRG1 binds to miR-148b promoter, ChIP assay was conducted using anti-BRG1 antibody in A549 cells. As shown in , BRG1 showed a high capacity to bind to the promoter region of miR-148b compared with control IgG.
Figure 2 BRG1 binds to miR-148b target gene promoter in lung cancer cells.
Abbreviations: BRG1, Brahma-related gene 1; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; si-control, control siRNA; ChIP, chromatin immunoprecipitation; TF, Transcription factor.
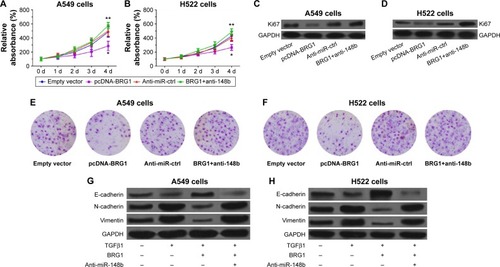
Restoration of BRG1 inhibits proliferation and metastasis of lung cancer cells
To demonstrate that restoration of BRG1 suppresses lung cancer development, we observed the effects of overexpression of BRG1 on lung cancer cells. As shown in , BRG1-overexpressing lung cancer cells had lower relative viability than the control group, but anti-miR-148b enhanced the cell viability. To confirm that BRG1 has an inhibitory effect on proliferation of cancer cells, we detected the expression of Ki67 in A549 and H522 cells by using Western blot analysis. The results showed that the Ki67 levels in BRG1-overexpressing lung cancer cells were significantly lower than those in the control groups, but downregulation of miR-148b elevated its levels (). We assessed the effects of BRG1 on metastasis of lung cancer in vitro by Transwell assay. As shown in , the number of cells transfected vector with BRG1 that migrated was reduced compared with the control, while knockdown of miR-148b could increased it. As EMT has been reported to facilitate tumor cell metastasis, we analyzed whether BRG1 functioned as an inhibitor of TGFβ1-induced EMT in A549 and H522 cells by detecting the expressions of E-cadherin, N-cadherin, and vimentin using Western blot analysis. The results indicated that after a 48-hour treatment of TGFβ1, the level of E-cadherin was decreased, while the levels of N-cadherin and vimentin were elevated, suggesting that TGFβ1 could induce EMT in lung cancer cells. Interestingly, we observed that overexpression of BRG1 could inhibit TGFβ1-induced EMT, but downregulation of miR-148b showed an opposite effect in A549 and H522 cells (). These data showed that restoration of BRG1 suppresses proliferation and metastasis of lung cancer cells and that miR-148b also functions as a tumor inhibitor.
Figure 3 Restoration of BRG1 inhibits proliferation and metastasis of lung cancer cells but downregulation of miR-148b resists the effects.
Abbreviations: BRG1, Brahma-related gene 1; EMT, epithelial–mesenchymal transition; d, days; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; Anti-miR-ctrl, Anti-miR-control.
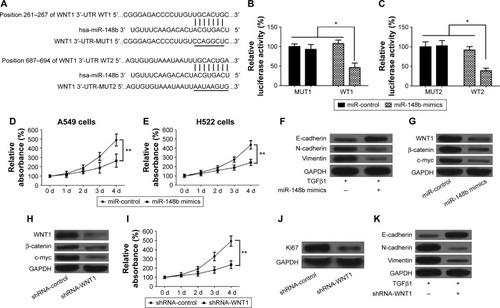
miR-148b suppresses lung cancer via regulating the WNT/β-catenin signaling pathway
Increasing evidence showed that the WNT/β-catenin signaling pathway plays a vital role in the promotion of the EMT and the progression of cancer.Citation29,Citation30 We used the bioinformatic analysis (MiRanda and TargetScan) for prediction of miR-148b-binding sites. Two putative miR-148b complementary regions in the 3′-UTR of WNT1 mRNA were observed (). To confirm whether miR-148b regulates the expression of WNT1 via directly binding to WNT1 sequences, the reporter plasmids containing the wild-type or mutant 3′-UTR of WNT1 were constructed and then transfected into A549 cells along with pre-miR-148b or a precursor control, and then luciferase activity was detected. High expression of miR-148b significantly decreased reporter activity of wild-type WNT1 constructs but not mutant WNT1 (), suggesting that the 3′-UTR of WNT1 is responsible for miR-148b function. To confirm that miR-148b regulated the progress of lung cancer through inactivating the WNT/β-catenin signaling pathway, we first assessed the effects of miR-148b on lung cancer cells and the expression levels of WNT1, β-catenin, and c-myc. The results showed that upregulation of miR-148b suppressed proliferation of A549 and H522 cells compared with the miR-control group (). We also evaluated the effects of miR-148b on the EMT of lung cancer cells by detecting the expression levels of E-cadherin, N-cadherin, and vimentin. Our results showed that overexpression of miR-148b increased the level of E-cadherin but decreased the levels of N-cadherin and vimentin compared with the miR-control group (), which suggested that miR-148b could inhibit EMT of lung cancer. Furthermore, upregulation of miR-148b decreased the levels of WNT1, β-catenin, and c-myc in A549 and H522 cells (), which suggested that miR-148b has an inhibitory effect to the WNT/β-catenin signaling. In addition, we assessed the roles of the WNT/β-catenin signaling pathway in EMT and lung cancer. As shown in , knockdown of WNT1 significantly decreased the levels of WNT1, β-catenin, and c-myc. In addition, knockdown of WNT1 inhibited proliferation and the expression level of Ki67 (). Compared to the control group, knockdown of WNT1 elevated the level of E-cadherin but reduced the levels of N-cadherin and vimentin (), displaying a similar role to miR-148b. Taken together, these data revealed that miR-148b suppresses the promotion of EMT and the progression of lung cancer via inactivating the WNT/β-catenin signaling pathway.
Figure 4 miR-148b inhibits proliferation and metastasis of lung cancer by directly regulating the WNT1/β-catenin signaling.
Abbreviations: EMT, epithelial–mesenchymal transition; 3′-UTR, 3′-untranslated region; d, days; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; MUT, mutation; WT, wild type.
Discussion
Recent sequencing efforts have shown silence of BRG1 in multiple primary NSCLC clinical specimens and human lung tumor cell lines.Citation31 In this study, BRG1 was confirmed to be significantly expressed at low levels in lung cancer tissues and cells. Furthermore, we validated that overexpression of BRG1 inhibited proliferation and metastasis of lung cancer cells and suppressed growth of xenografts in nude mice. Romero et alCitation8 reported that BRG1-lacking cell lines from several cancer types were not sensitive to retinoic acid or glucocorticoid, while upregulation of BRG1 restored sensitivity. Additionally, restoration of BRG1 significantly suppressed invasion and progression of lung cancer cells orthotopically transplanted in nude mice. Another study showed that decreased BRG1 enhanced EMT and metastasis of colorectal cancer by regulating the miR-550a-5p/ring finger 43/WNT/β-catenin signaling pathway in vitro and in vivo.Citation32 However, the effects of BRG1 on EMT and metastasis of lung cancer are unknown. In this study, we displayed that restoration of BRG1 suppressed EMT and metastasis of lung cancer cells by upregulation of miR-148b and inhibition of the WNT/β-catenin signaling pathway.
Overexpression of BRG1 promoted the transcriptional activity of miR-148b and silence of BRG1 by siRNA led to decreased miR-148b expression in this study, accounting for the low expression level of miR-148b, a new cancer suppressor, in lung cancer cells. BRG1 acts as a master regulator of the genome, for it transcriptionally regulates approximately 4.8% of the human genome, including genes that participate in plenty of signaling pathways.Citation33 For example, BRG1 inhibits the transcriptional activity of miR-550a-5p promoter, and decreased BRG1 expression elevates miR-550a-5p expression level in colorectal cancer.Citation33 BRG1 accelerates COUP-TFII expression via binding conserved regulatory regions to the COUP-TFII promoter and remodeling chromatin to make the promoter accessible to transcriptional machinery in vascular endothelial cells.Citation34 In this study, we showed that BRG1 is a transcription factor of miR-148b using ChIP assay. Therefore, we deduced that BRG1 restoration transcriptionally regulated miR-148b, leading to inhibition of proliferation and metastasis of human lung cancer.
The WNT/β-catenin signaling pathway has been documented to play a critical role in the progression and promotion of EMT and cancer metastasis.Citation35 Many studies have showed that WNT1, a mammary oncogene, is associated with mammary oncogenesis and that β-catenin is overexpressed in human cancers, which in turn promotes the expression of target genes such as E-cadherin and c-myc that are characterized by cancer development and poor clinical prognosis.Citation36,Citation37 Most recently, miR-148b was demonstrated to suppress cell proliferation and invasion by downregulating the expression of WNT1, β-catenin, and c-myc and upregulating E-cadherin expression in hepatocellular carcinoma.Citation37 In breast cancer cell lines, miR-374a directly targets and suppresses numerous negative regulators of the WNT/β-catenin signaling cascade, including WIF1, PTEN, and WNT5A, promoting EMT and metastasis both in vitro and in vivo.Citation35 In this study, we identified that miR-148b, regulated by BRG1, bound to two putative complementary regions in the 3′-UTR of WNT1 mRNA, inactivating the WNT/β-catenin signaling, resulting in inhibition of cell proliferation and metastasis in lung cancer cells. We also found that inactivation of the WNT/β-catenin signaling suppressed EMT of lung cancer cells. Previous studies have showed that EMT allows benign tumor cells to infiltrate surrounding tissues and metastasize to distant sites, facilitating the early stage of metastatic progression.Citation38,Citation39 Hence, our study revealed that inhibition of EMT by BRG1-depended miR-148b overexpression may contribute to decrease the metastatic capacity of lung cancer cells.
In conclusion, the gene encoding BRG1 was frequently downregulated in human lung cancer. Restoration of BRG1 suppressed cell proliferation and metastasis in lung cancer cells. Moreover, we identified that BRG1 positively regulated the expression of miR-148b and that miR-148b suppressed EMT of lung cancer cells by regulating the WNT/β-catenin signaling via directly binding to WNT1 sequences. The BRG1/miR-148b/WNT/β-catenin pathway provides potential therapeutic value for the treatment of lung cancer.
Disclosure
The authors report no conflicts of interest in this work.
References
- WileyMMMuthukumarVGriffinTMGriffinCTSWI/SNF chromatin-remodeling enzymes Brahma-related gene 1 (BRG1) and Brahma (BRM) are dispensable in multiple models of postnatal angiogenesis but are required for vascular integrity in infant miceJ Am Heart Assoc201544e00197225904594
- ZhangXLiBLiWTranscriptional repression by the BRG1-SWI/SNF complex affects the pluripotency of human embryonic stem cellsStem Cell Reports20143346047425241744
- OrvisTHepperlaAWalterVBRG1/SMARCA4 inactivation promotes non-small cell lung cancer aggressiveness by altering chromatin organizationCancer Res201474226486649825115300
- ReismanDNSciarrottaJWangWFunkhouserWKWeissmanBELoss of BRG1/BRM in human lung cancer cell lines and primary lung cancers: correlation with poor prognosisCancer Res200363356056612566296
- FukuokaJFujiiTShihJHChromatin remodeling factors and BRM/BRG1 expression as prognostic indicators in non-small cell lung cancerClin Cancer Res200410134314432415240517
- MedinaPPCarreteroJBallestarETranscriptional targets of the chromatin-remodelling factor SMARCA4/BRG1 in lung cancer cellsHum Mol Genet200514797398215731117
- MedinaPPRomeroOAKohnoTFrequent BRG1/SMARCA4–inactivating mutations in human lung cancer cell linesHum Mutat200829561762218386774
- RomeroOASetienFJohnSThe tumour suppressor and chromatin-remodelling factor BRG1 antagonizes Myc activity and promotes cell differentiation in human cancerEMBO Mol Med20124760361622407764
- ChenLWangXWangHmiR-137 is frequently down-regulated in glioblastoma and is a negative regulator of Cox-2Eur J Cancer201248163104311122406049
- ZhuXLiYShenHmiR-137 inhibits the proliferation of lung cancer cells by targeting Cdc42 and Cdk6FEBS Lett20135871738123178712
- LiuMLangNQiuMmiR-137 targets Cdc42 expression, induces cell cycle G1 arrest and inhibits invasion in colorectal cancer cellsInt J Cancer201112861269127920473940
- KimHSLeeKSBaeHJMicroRNA-31 functions as a tumor suppressor by regulating cell cycle and epithelial-mesenchymal transition regulatory proteins in liver cancerOncotarget20156108089810225797269
- ChrestaCMSchlickerABeranGCerilloGWesselsLFOrphanidesGReciprocal expression of miR-125b and miR-192/miR-200 families define clinically relevant sub-types of colorectal cancer with differential sensitivity to EGFR and MEK targeted agentsCancer Res20147419 Suppl5559
- HuangYCHungLYTumor suppressive miR-137 targets Aurora-A and leads to apoptosis in colorectal cancer cellsCancer Res20147419 Suppl4344
- ZhaoGZhangJGLiuYmiR-148b functions as a tumor suppressor in pancreatic cancer by targeting AMPKα1Mol Cancer Ther2013121839323171948
- SongYXYueZYWangZNMicroRNA-148b is frequently down-regulated in gastric cancer and acts as a tumor suppressor by inhibiting cell proliferationMol Cancer2011101121205300
- CiminoDDe PittàCOrsoFmiR148b is a major coordinator of breast cancer progression in a relapse-associated microRNA signature by targeting ITGA5, ROCK1, PIK3CA, NRAS, and CSF1FASEB J20132731223123523233531
- ZhangCWangCChenXExpression profile of microRNAs in serum: a fingerprint for esophageal squamous cell carcinomaClin Chem201056121871187920943850
- SongYXuYWangZMicroRNA-148b suppresses cell growth by targeting cholecystokinin-2 receptor in colorectal cancerInt J Cancer201213151042105122020560
- WuYLiuGLLiuSHMicroRNA-148b enhances the radio-sensitivity of non-Hodgkin’s Lymphoma cells by promoting radiation-induced apoptosisJ Radiat Res201253451652522843616
- AziziMTeimoori-ToolabiLArzananiMKAzadmaneshKFard-EsfahaniPZeinaliSMicroRNA-148b and microRNA-152 reactivate tumor suppressor genes through suppression of DNA methyltransferase-1 gene in pancreatic cancer cell linesCancer Biol Ther201415441942724448385
- LiuQXuYWeiSmicroRNA-148b suppresses hepatic cancer stem cell by targeting neuropilin-1Bios Rep2015354e00229
- GeHLiBHuWXMicroRNA-148b is down-regulated in non-small cell lung cancer and associated with poor survivalInt J Clin Exp Pathol20158180080525755777
- LiuGLLiuXLvXBWangXPFangXSSangYmiR-148b functions as a tumor suppressor in non-small cell lung cancer by targeting carcinoembryonic antigen (CEA)Int J Clin Exp Med2014781990199925232379
- TangSCSheuGTWongRHExpression of glutathione S-transferase M2 in stage I/II non-small cell lung cancer and alleviation of DNA damage exposure to benzo [a] pyreneToxicol Lett2010192331632319900515
- XiaJWuZYuCmiR-124 inhibits cell proliferation in gastric cancer through down-regulation of SPHK1J Pathol2012227447048022450659
- GlarosSCirrincioneGMPalancaAMetzgerDReismanDTargeted knockout of BRG1 potentiates lung cancer developmentCancer Res200868103689369618483251
- WatanabeHMizutaniTHaraguchiTSWI/SNF complex is essential for NRSF-mediated suppression of neuronal genes in human nons-mall cell lung carcinoma cell linesOncogene200625347047916247481
- ThuKRadulovichNBecker-SantosDSOX15 is a candidate tumor suppressor in pancreatic cancer with a potential role in Wnt/β-catenin signalingOncogene201433327928823318427
- KimWKOhJBaeSYAntitumor activity of 2-hydroxycinnam-aldehyde against human colorectal cancer cells is mediated by the Wnt/β-catenin signaling pathwayCancer Res20147419 Suppl4237
- SongSWalterVKaracaMGene silencing associated with SWI/SNF complex loss during NSCLC developmentMol Cancer Res201412456057024445599
- WangGFuYYangXBrg-1 targeting of novel miR550a-5p/RNF43/Wnt signaling axis regulates colorectal cancer metastasisOncogene Epub511201510.1038/onc.2015.124
- ZhangLNemzowLChenHHuJJGongFWhole genome expression profiling shows that BRG1 transcriptionally regulates UV inducible genes and other novel targets in human cellsPloS One201498e10576425157878
- DavisRBCurtisCDGriffinCTBRG1 promotes COUP-TFII expression and venous specification during embryonic vascular developmentDevelopment201314061272128123406903
- CaiJGuanHFangLMicroRNA-374a activates Wnt/β-catenin signaling to promote breast cancer metastasisJ Clin Invest2013123256657923321667
- ChenGShukeirNPottiAUp-regulation of Wnt-1 and β-catenin production in patients with advanced metastatic prostate carcinomaCancer200410161345135615316903
- ZhangJShiYHongDMiR-148b suppresses cell proliferation and invasion in hepatocellular carcinoma by targeting Wnt/β-catenin pathwaySci Rep20155808725627001
- GujralTSChanMPeshkinLSorgerPKKirschnerMWMacBeathGA noncanonical frizzled2 pathway regulates epithelial-mesenchymal transition and metastasisCell2014159484485625417160
- SleemanJPThieryJPSnapShot: the epithelial-mesenchymal transitionCell20111451162.e121458675