
Abstract
Introduction
The perioperative pain experience shows great interindividual variability and is difficult to predict. The mu-1 opioid receptor gene (OPRM1) is known to play an important role in opioid-pain pathways. Since deoxyribonucleic acid (DNA) methylation is a potent repressor of gene expression, DNA methylation was evaluated at the OPRM1 promoter, as a predictor of preoperative, acute, and chronic postsurgical pain (CPSP).
Methods
A prospective observational cohort study was conducted in 133 adolescents with idiopathic scoliosis undergoing spine fusion under standard protocols. Data regarding pain, opioid consumption, anxiety, and catastrophizing (using validated questionnaires) were collected before and 2–3 months postsurgery. Outcomes evaluated were preoperative pain, acute postoperative pain (area under curve [AUC] for pain scores over 48 hours), and CPSP (numerical rating scale >3/10 at 2–3 months postsurgery). Blood samples collected preoperatively were analyzed for DNA methylation by pyrosequencing of 22 CpG sites at the OPRM1 gene promoter. The association of each pain outcome with the methylation percentage of each CpG site was assessed using multivariable regression, adjusting for significant (P<0.05) nongenetic variables.
Results
Majority (83%) of the patients reported no pain preoperatively, while CPSP occurred in 36% of the subjects (44/121). Regression on dichotomized preoperative pain outcome showed association with methylation at six CpG sites (1, 3, 4, 9, 11, and 17) (P<0.05). Methylation at CpG sites 4, 17, and 18 was associated with higher AUC after adjusting for opioid consumption and preoperative pain score (P<0.05). After adjusting for postoperative opioid consumption and preoperative pain score, methylation at CpG sites 13 and 22 was associated with CPSP (P<0.05).
Discussion
Novel CPSP biomarkers were identified in an active regulatory region of the OPRM1 gene that binds multiple transcription factors. Inhibition of binding by DNA methylation potentially decreases the OPRM1 gene expression, leading to a decreased response to endogenous and exogenous opioids, and an increased pain experience.
Introduction
Inadequately controlled pain remains a significant problem after surgery, as it negatively affects quality of life and function and increases the risk of persistent postsurgical pain.Citation1 Approximately 50%–75% of patients undergoing surgery experience moderate or severe pain, and this is true for the 6 million children who undergo surgery every year in the US.Citation2,Citation3 Chronic postsurgical pain (CPSP) is the pain that lasts beyond 2 months postsurgery.Citation4 In children, the incidence of CPSP ranges from 13% to 68.8%.Citation5,Citation6 This implies that of the 1.5 million children who undergo major surgery every year in the US, ~225,000–1,000,000 children develop CPSP.Citation7 Interindividual variability in pain sensitivity,Citation8 acute postsurgical pain,Citation9–Citation11 and CPSP have been partly explained by genetic markers.Citation12 The mu-1 opioid receptor gene (OPRM1) that codes for the mu opioid receptor (MOR) plays an important role in opioid-pain pathways.Citation13 The OPRM1 variant that has been most commonly studied is the A118G variant; however, the results of association studies with pain and β-endorphin-binding activity between this variant and the wild-type receptors are controversial.Citation14–Citation17
Beyond genetics, chronic pain is a classic example of gene–environment interaction,Citation18 and hence recently interest has been directed toward the role of epigenetics in pain. Epigenetics is the study of changes in chromosomes that do not alter the sequence of deoxyribonucleic acid (DNA),Citation19 but may still lead to alterations in gene expression. Genetics and epigenetics together are important factors in the transition of acute postsurgical pain to CPSP.Citation20,Citation21 DNA methylation is a common epigenetic mechanism, which involves the addition of a methyl group to the 5′ position of a cytosine residue followed by a guanine residue (a CpG dinucleotide), which are often clustered (CpG islands) in the promoter regions of genes.Citation22 DNA methylation at the OPRM1 promoter region regulates DNA binding of transcription factors and is a potent epigenetic repressor of gene transcription.Citation23 An increase in the OPRM1 promoter methylation has been found to be associated with a decrease in protein expression of OPRM1.Citation24 Because DNA methylation is reversible, there is much interest in understanding its association with pain, as a potential target for intervention. The OPRM1 DNA methylation levels have been found to be elevated in opioid and heroin addicts.Citation25,Citation26 However, this has not been studied in relation to perioperative pain and CPSP. It has been hypothesized that DNA methylation at the promoter region of OPRM1 will be associated with pain before and after surgery. Understanding the contribution of OPRM1 genetic–epigenetic interactions to pain outcomes will allow prediction of susceptibility to poor pain control and CPSP and will enable target identification for modification of risk studies in the future.
Methods
A prospective observational cohort study was conducted in 133 adolescents with idiopathic scoliosis undergoing posterior spine fusion under standard intraoperative anesthesia (propofol–remifentanil total intravenous anesthesia, guided by electroencephalography monitoring for depth of anesthesia) and postoperative analgesia with morphine patient-controlled analgesia along with adjuvants (scheduled intravenous acetaminophen, ketorolac, and diazepam as needed and methocarbamol) managed by perioperative pain team. The study was approved by the Cincinnati Children’s Hospital Institutional Review Board. This study is registered with ClinicalTrials.gov identifiers NCT01839461 and NCT01731873. Written informed consent was obtained from parents, and assent was obtained from children before enrollment.
Participants
Healthy nonobese subjects with an American Society of Anesthesiologists (ASA) physical status ≤2 (mild systemic disease), aged 10–18 years, with a diagnosis of idiopathic scoliosis and/or kyphosis, and undergoing elective spinal fusion were recruited. The exclusion criteria included pregnant or breastfeeding females, presence of chronic pain defined as use of opioids in the past 6 months, liver or renal diseases, and developmental delays.
Data collection
Preoperatively, data regarding demographic factors (sex, age, and race), weight, pain scores (numerical rating scale [NRS]/0–10)Citation27 on the day of surgery (P0), and pain medications used were obtained. Anxiety scores for both child and a parent were assessed using the 0–10 visual analog scale (VAS), a simple validated scale that has been used previously in children.Citation28 Questionnaires were administered as described in the following sections. The intraoperative data collected included propofol and remifentanil doses, duration of surgery, and number of vertebral levels fused. In the immediate postoperative period (postoperative days [PODs] 1 and 2), pain scores (every 4 hours) and morphine and diazepam doses administered were noted. After hospital discharge, the questionnaires were administered per schedule presented in to obtain psychosocial and pain measures in a standard fashion.
Table 1 Data collection schema
Outcomes
Pain outcomes evaluated were 1) preoperative pain, 2) acute postoperative pain (defined as area under curve [AUC] for pain scores over time on POD 1 and 2 [AUCPOD1–2] calculated using trapezoidal rule), and 3) chronic pain/CPSP (NRS >3/10 at 2–3 months postsurgery) as defined by the International Association for the Study of Pain.Citation29 These cut offs for NRS were used because NRS pain scores >3 (moderate/severe pain) at 3 months have been described as a predictor for persistence of pain and associated with functional disability.Citation30
Measurement of DNA methylation
Blood was drawn upon intravenous line placement before surgery, from which the DNA was isolated on the same day and frozen at −20°C. To study DNA methylation, the focus was on a 251 bp region, including 22 CpG sites reaching from position −93 to position +159, whose methylation levels were previously shown to be associated with the OPRM1 expressionCitation24 and opioid/heroin addiction ().Citation25,Citation26 The CpG sites are numbered according to those studiesCitation25,Citation26 to allow for an easy comparison. This included a CpG site at +117, reported by Oertel et al.Citation24 The pyrosequencing assays utilize 50–500 ng of genomic DNA of acceptable quality (measured by Thermo Scientific NanoDrop spectrophotometer [Waltham, MA, USA] and with a 260/280 ratio ranging from 1.6 to 2.0). The extracted DNA was treated with bisulfite using Zymo EZ DNA Methylation Gold Kit (Zymo Research, Orange, CA, USA), according to the manufacturer’s instructions. Two sets of primers, long and nested, were designed using ZymoTaq Premix (Zymo Research) for two rounds of polymerase chain reaction (PCR). Methylation of the PCR products was quantified using PyroMark MD 1.0 software (Qiagen, Valencia, CA, USA). Primers used in the assays are listed in . Samples were repeated if the pyrosequencing runs did not pass quality checks of the pyrosequencer or if the internal bisulfite conversion controls failed. The sample runs were monitored through methylated and nonmethylated DNA as well as template-free negative controls.
Figure 1 Depiction of the OPRM1 promoter region (HG19; Chr 6: 154360587 to 154360838) and the location of the CpG sites. The knobs represent each CpG site, and the primers are indicated in brackets below. The red-colored knob at +117 indicates the CpG site (CpG17) associated with the variant A118G. The arrows indicate sites that have been described as Sp1 transcription factor-binding sites in previous studies (CpG sites 9, 10, 12, 16, 21, and 23 at −18, −14, 12, +84, +145, +150, and +159 from ATG site).
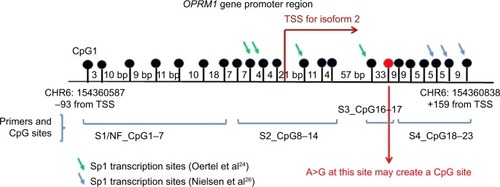
Table 2 Primers used in the pyrosequencing assay
Data analysis
Prior to the analyses, the data quality was assessed. Demographics and patients’ clinical characteristics were summarized as mean (standard deviation [SD]), median, and frequency (percentage) according to the distribution of the data. Prior to evaluation of the association between pain outcome and DNA methylation, the effects of covariables were tested, which included age, sex, race, morphine dose in mg/kg on POD 1 and 2, preoperative anxiety score (VAS), preoperative pain score, duration of surgery, vertebral levels fused, propofol and remifentanil doses used during surgery (per kg), use of intravenous acetaminophen/ketorolac (yes/no), diazepam doses (mg/kg), and pain catastrophizing scale (parent version) and parent pain history scores and sequential scores for Childhood Anxiety Sensitivity Index and pain catastrophizing scale (child version).Citation31 Three pain outcomes were examined. Preoperative pain was dichotomized as yes versus no, because, although it was measured using NRS as a 0–10 scale, 83% of the patients reported no preoperative pain. It was then associated with covariables using logistic regression. Similarly, acute postoperative pain was analyzed using simple linear regression models, and CPSP using logistic regression models. Covariables associated at P<0.10 were entered into multivariable models, and stepwise selection was used to derive a final nongenetic model for each outcome where only variables with P<0.05 were retained.Citation32 DNA methylation levels were then added to the final nongenetic model to assess their association with the pain outcomes. Statistical analyses were performed using Statistical Analysis System, version 9.3 (SAS Institute Inc., Cary, NC, USA). P≤0.05 was used as the threshold for statistical significance. No multiple testing correction was done, thus it is possible that some findings occurred by chance.
Functional genomics analysis
To identify potential regulatory mechanisms in the OPRM1 promoter, a large collection of functional genomics data sets from various sources, including ENCODE,Citation33 Road-map Epigenomics,Citation34 Cistrome,Citation35 and ReMap-ChIP, was compiled.Citation36 The genomic coordinates of the OPRM1 promoter were intersected against the genomic coordinates contained in each data set. In total, this database contains 4,953 data sets performed in 1,706 different cell types and conditions; 1,911 data sets monitor binding interactions of transcription factors with the human genome using ChIP-seq; 1,214 measure the presence of a particular histone mark using ChIP-seq; 226 measure open chromatin through DNase-seq; 57 measure expression quantitative trait loci (eQTLs); and 558 predict “ActiveChromatin” states using combinations of histone marks.Citation37 Collectively, 240 of these experiments were performed in brain-related cell lines and cell types.
Results
The final cohort comprised 133 participants; the mean age was 14.49 years (SD 1.91), and they were mostly white (83%) and female (74%) (). The recruitment timeline is described in . Descriptions of variables that were evaluated for inclusion in the multiple regression model are presented in .
Figure 2 Recruitment timeline for the spine surgery study cohort is delineated. Of the 261 eligible patients who satisfied the inclusion/exclusion criteria, reasons for not enrolling and derivation of final cohorts included in the study with preoperative, acute, and chronic pain outcomes are described.
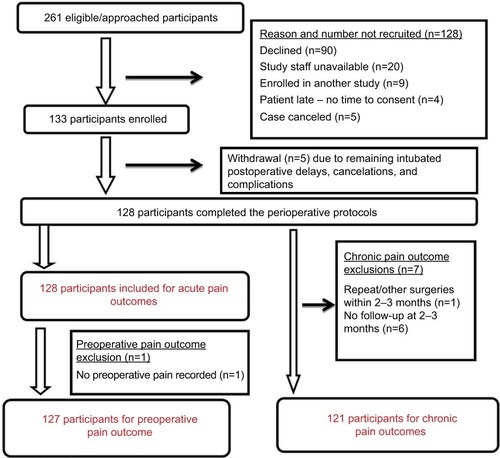
Table 3 Demographics of the cohorts and description of the covariates used in the regression model
Pain descriptives
Acute and chronic postoperative pain data were collected for 128 and 121 patients, respectively. The overall incidence of CPSP was found to be 36.4% (44/121). The preoperative pain score was 0 in 83% (106/127) of the cohort.
DNA methylation and pain
Three pain outcomes were examined. For the association with dichotomized preoperative pain scores, logistic regression was used. After adjusting for age and sex, the methylation level of six CpG sites (sites 1, 3, 4, 9, 11, and 17, ) was significantly associated with whether a patient reported preoperative pain. For all the six CpG sites, higher methylation was associated with higher odds of reporting preoperative pain, except for site 17. No impact of the methylation on the actual pain scores was detected (data not shown). For acute postoperative pain (AUCPOD1–2), significant impact of methylation was detected in CpG sites 4, 17, and 18 () when preoperative pain and morphine consumption were adjusted. For all three CpG sites, higher methylation was associated with higher preoperative pain. For chronic postoperative pain, significant association was detected for the nonpromoter site, CpG13 and 22 (), with preoperative pain and morphine consumption being controlled. For all the CpG sites, higher methylation was associated with higher odds of having chronic pain. Methylation at two CpG sites was associated with both preoperative pain and AUCPOD1–2 (CpG 4 and 17); methylation at these sites also showed a trend toward CPSP risk, although not statistically significant. depicts estimated probabilities of developing CPSP using median preoperative pain score (0), median morphine doses (1.7 mg/kg), and 2.5th, 25th, 50th, 75th and 97.5th percentiles of the methylation data of each of the two sites CpG13 and CpG22.
Figure 3 The probability of developing CPSP based on DNA methylation at CpG 13 and 22, derived from the regression model, is depicted. The probabilities were estimated using median preoperative pain scores (0), median morphine consumption (1.7 mg/kg), and 2.5%, 25%, 50%, 75%, and 97.5% of the methylation data of each of the two sites. The 97.5% values for DNA methylation in the data are 40% for CpG13 and 57% for CpG22. The nongenetic covariates are already adjusted for in the regression model. Hence, the probability of CPSP holding other variables constant increases with increased methylation at these sites.
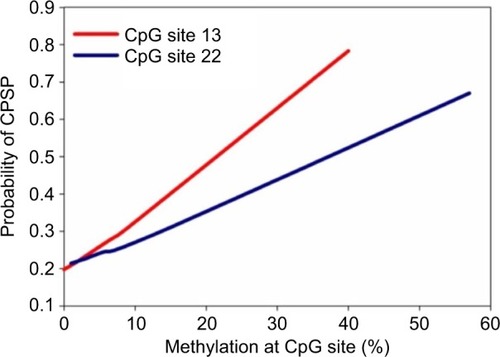
Table 4 Association of DNA methylation of CpG sites at the OPRM1 promoter with pain outcomes
Functional genomics analysis
Query of the region evaluated using a large collection of ChIP-seq, DNase-seq, and eQTL data, as described in the “Methods” section, showed that this region is located in open chromatin and is marked by H3K27ac, H3K4me1, and H3K4me3 (indicative of active regulatory regions)Citation38–Citation40 in noncancer brain cells from the caudate–putamen, temporal, frontal lobes, and angular gyri. Moreover, this region contains ChIP-seq peaks for binding of multiple transcription factors, including REST, RAD21, SP1, YY1, and ZNF263 in various tissues. In particular, REST and RAD21 bind the OPRM1 promoter region in three cell lines (SK-N-SH, PFSK-1, and SK-N-SH) derived from brain tissue where the opioid receptors responsible for analgesia are found (). Collectively, these results suggest that the differential DNA methylation patterns observed in the OPRM1 promoter might functionally act by modulating the expression of OPRM1 via alteration of the binding of REST and other neuron-expressed transcription factors.
Table 5 Findings from evaluation of OPRM1 promoter region using functional genomics datasets in neuronal cell-type
Discussion
It has been previously shown that psychological and clinical variables contribute to CPSP.Citation31 This study finds associations between epigenetics and CPSP in children for the first time and adds to the emerging evidence linking epigenetic mechanisms to the development of chronic pain states.Citation41 Specifically, it was found that novel biomarkers (DNA methylation of certain CpG sites in the OPRM1 promoter region) associated with preoperative pain, acute postoperative pain, and CPSP following posterior spine fusion in healthy adolescents. DNA methylation was also associated with pain before surgical stimulus and exposure to opioids. These findings allow for prediction of risk for the pain response to surgery and possible novel mechanisms that could be targeted for prevention and treatment of CPSP.
Endogenous opioid peptides and exogenously administered opioid analgesics bind to MOR to regulate pain responses. The MOR activity is regulated at different levels, including epigenetic mechanisms.Citation23 Many of the CpG sites found to be associated with pain outcomes (1, 3, 4, 9, 11, 13, 17, 18, and 22) have been previously described as putative Sp1 transcription factor-binding sites. These include CpG sites 9, 10, 12, 16, 21, and 23 at −18, −14, 12, +84, +145, and +159 from ATG site, respectively.Citation26 Methylation levels at some of these sites were previously found to be associated with opioid addiction/dependence. Nielsen et al found that CpG sites at −18 and +84 (9 and 16) were more strongly methylated in heroin addicts than controls. Similarly, seven CpG sites showed significant hypermethylation of blood DNA taken from male opioid addicts when compared to blood DNA from controls (CpGs 5, 9, 10, 11, 18, and 23).Citation26 Increased methylation within the OPRM1 promoter (at −14, −10 [sites 10, 11]) was also found to be associated with worse neonatal abstinence syndrome outcomes in infants exposed to opioids in utero.Citation42 It has been shown in mouse brain tissues that DNA methylation of the OPRM1 promoter decreases expression of the gene; through interaction with chromatin-remodeling factors, remodeling occurs, thus allowing access for Sp1 binding,Citation43 which results in the MOR upregulation. Thus, methylation at these sites can prevent the OPRM1 activation, leading to decreased endogenous and exogenous opioid effects, manifested in the experience of increased perioperative pain. In addition, Chorbov et al also observed increased DNA methylation in the sperm of addicts, which may suggest a means of epigenetic heritability of opioid phenotypes.Citation25
Animal studies have shown that OPRM1 promoter methylation reduced OPRM1 expression.Citation23 Knothe et al confirmed methylation effects on OPRM1 expression through experiments in human cell lines (neuronal SHSY5Y and Kelly, and nonneuronal HEK-293) with and without the demethylating agent 5′-aza-2′-deoxycytidine.Citation44 But they did not find that methylation profiles across the OPRM1 gene from CpG position −93 to +159 in brain tissues collected postmortem from heroin addicts correlated with differences in OPRM1 mRNA expression in the corresponding brain regions.Citation44 Opioid exposure in all their subjects may have played a role in minimizing methylation differences between the specimens and the individuals. Hence, the lack of correlation in their study does not rule out the differences that may exist between opioid-exposed and nonopioid-exposed (or pain vs no pain) subjects.
The findings of this study corroborate a growing body of evidence that OPRM1 DNA methylation affects pain behaviors and contributes to the limited efficacy of opiates in certain cases,Citation45 which is improved upon blockade of methylation.Citation46 In a neuropathic animal model, it was shown that increased methylation of the MOR gene proximal promoter in dorsal root ganglion decreased morphine analgesia.Citation47 Administration of demethylating agents reversed the hypermethylation of the OPRM1 gene and improved the analgesic effect of morphine in mice pain models.Citation47,Citation48 It is not clear if pain is the trigger or the consequence of increased methylation of the OPRM1 promoter. In fact, chronic opioid use in methadone-substituted former opiate addicts was found to be associated with increased DNA methylation at the OPRM1 position +126 (CpG 18), correlating with increased pain, compared to controls with pain not treated with opioids.Citation49 The authors note that causal relationships with opioid use could not be established, as there was no opioid dose dependency. In their discussion, they mentioned that the trigger for increased methylation may actually be the baseline increased pain.Citation50 This might partly explain the association of OPRM1 methylation with baseline pain identified in this study, as MOR function is essential for endogenous opioid action even in the absence of opioids. Useful information would be gained by evaluating DNA methylation changes over time in those who do and do not develop CPSP, in the presence/absence of opioids.
Although the exact mechanisms by which DNA methylation of OPRM1 promoter region contributes to pain are not known, the computational analysis revealed a DNA region bound by the repressor element 1 silencing transcription factor (REST) in multiple neuronal cell lines at the OPRM1 promoter region (). Intriguingly, REST has previously been implicated in OPRM1 gene silencing via epigenetic modifications.Citation51 REST, a member of the zinc finger transcription factor family, represses transcription of neural genesCitation52 in nonneuronal cells by binding a DNA sequence element called the neuron-restrictive silencer elementCitation53 and recruiting several chromatin-modifying enzymes.Citation54 There is also evidence for the involvement of REST in chronic neuropathy.Citation52 Nerve injury results in a long-lasting increase in REST expression in mouse dorsal root ganglia.Citation55 Hence, it is hypothesized that, in patients with already suppressed OPRM1 expression due to DNA methylation of the OPRM1 promoter, increased REST expression after injury possibly leads to further OPRM1 gene silencing and worsens the pain experience. If true, this mechanism could potentially create new avenues for pain therapy. Besides the sequence-specific REST, RAD21 was another transcription factor that was found to be relevant in neuronal cells for the OPRM1 promoter region. REST is involved in three-dimensional DNA organization. Recent research, using ChIP-seq and RNA-seq data from matching cell types from the human ENCODE resource, showed that RAD21 mostly functions as an activator of transcription.Citation56 Although not known to play a role in OPRM1 regulation or pain, DNA methylation preventing binding of an activating transcription factor may also be responsible for decreased OPRM1 function and hence pain in these patients.
This study has a few limitations, one of which is the use of blood samples for DNA methylation, instead of target tissue like brain, which are understandably inaccessible in clinical human studies. However, this approach has been employed successfully before.Citation25,Citation26,Citation42 Fan and Zhang compared methylation profiles of human chromosome 6 (OPRM1 gene location), derived from 12 tissues, and reported that CpG island methylation profiles were highly correlated between somatic tissues.Citation57 Davies et al found that some interindividual variation in DNA methylation was reflected across brain and blood, indicating that peripheral tissues may have utility in studies of complex neurobiological phenotypes.Citation58
In summary, we report novel associations of DNA methylation in the OPRM1 promoter with preoperative, acute, and CPSP in children undergoing spine surgery. Since DNA methylation is influenced by multiple modifiable factors (diet, exercise, parental upbringing, and stress), understanding the role of epigenetic regulation of OPRM1 in pain opens new avenues of pain research. Understanding susceptibility may act as a guide for targeted use of aggressive multimodal analgesia, use of calcium-channel modulators (like gabapentin) and preemptive analgesics like N-methyl-D-aspartate receptor antagonists (like ketamine), regional anesthesia, and behavioral therapies in patients with higher risk.Citation59 Addressing this higher risk using alternative strategies may be especially important as the MOR protein encoded by OPRM1 plays a key role in mediating not only pain responses but also the development of tolerance and physical dependence.Citation60,Citation61 Future studies need to map methylation changes in the OPRM1 promoter over the time course of CPSP, and the effect of preventive and therapeutic strategies on transition of acute pain to CPSP, and development of tolerance/opioid hyperalgesia.
Summary
Surgery is a stressful and painful experience; however, some individuals continue to experience pain, even months or years after surgery. This is because pain experience is unique. It has been shown that psychological factors play a role in shaping pain responses after surgery. Genes also play an important role in determining pain. One such gene is the mu-1 opioid receptor gene (OPRM1) that codes for the mu opioid receptor (MOR), where potent pain medications (opioids) bind. In this study, we evaluated whether nonstructural changes in the deoxyribonucleic acid (DNA) (DNA methylation) of the regulatory region of the OPRM1 gene could predict the individual pain experience. In 133 adolescents undergoing spine fusion, after controlling for important nongenetic covariates identified from a previous analysis, it was found that methylation at several regulatory sites was associated with preoperative, immediate postoperative, and chronic postoperative pain. These sites were identified as binding regions for important transcription factors; methylation likely affects factor binding and decreases the OPRM1 expression, leading to increased pain sensitivity. These serve as novel biomarkers for pain and, since DNA methylation is modifiable, might provide a basis for future preventive and therapeutic strategies.
Acknowledgments
The authors would like to acknowledge Ashley Ulm and Veda Yadagiri (Pyrosequencing Core, Cincinnati Children’s Hospital Medical Center [CCHMC]) and Diane Kissell for their role in analyzing the DNA extraction and pyrosequencing, under the supervision of Hong Ji (Director, Pyrosequencing Core) and Kejian Zhang (Director of Molecular Genetics Lab, CCHMC). They would also like to acknowledge Kayla Stallworth and Hope Esslinger, CCRC IV, previous research coordinators for the Department of Anesthesia, CCHMC, for their help with patient recruitment in the earlier stages of the study. This study was supported by 5K23HD082782 through the Eunice Kennedy Shriver National Institute of Child Health and Human Development, National Institutes of Health (PI: Chidambaran), Center for Pediatric Genomics, and Shared Facility Discovery Award from CCHMC (PI: Chidambaran). The authors are solely responsible for the content, and it does not necessarily represent the official views of the National Institutes of Health.
Disclosure
The authors report no conflicts of interest in this work.
References
- KehletHJensenTSWoolfCJPersistent postsurgical pain: risk factors and preventionLancet200636795221618162516698416
- GanTJHabibASMillerTEWhiteWApfelbaumJLIncidence, patient satisfaction, and perceptions of post-surgical pain: results from a US national surveyCurr Med Res Opin201430114916024237004
- PolkkiTPietilaAMVehvilainen-JulkunenKHospitalized children’s descriptions of their experiences with postsurgical pain relieving methodsInt J Nurs Stud2003401334412550148
- MacraeWAChronic post-surgical pain: 10 years onBr J Anaesth20081011778618434337
- LandmanZOswaldTSandersJDiabMSpinal Deformity Study GroupPrevalence and predictors of pain in surgical treatment of adolescent idiopathic scoliosisSpine (Phila Pa 1976)2011361082582921192302
- PageMGStinsonJCampbellFIsaacLKatzJIdentification of pain-related psychological risk factors for the development and maintenance of pediatric chronic postsurgical painJ Pain Res2013616718023503375
- KainZNMayesLCO’ConnorTZCicchettiDVPreoperative anxiety in children. Predictors and outcomesArch Pediatr Adolesc Med199615012123812458953995
- LaCroix-FralishMLAustinJSZhengFYLevitinDJMogilJSPatterns of pain: meta-analysis of microarray studies of painPain201115281888189821561713
- De GregoriMDiatchenkoLIngelmoPMHuman genetic variability contributes to postoperative morphine consumptionJ Pain201617562863626902643
- SadhasivamSChidambaranVPharmacogenomics of opioids and perioperative pain managementPharmacogenomics201213151719174023171337
- SadhasivamSChidambaranVOlbrechtVAGenetics of pain perception, COMT and postoperative pain management in childrenPharmacogenomics201415327728424533707
- ClarkeHKatzJFlorHRietschelMDiehlSRSeltzerZGenetics of chronic post-surgical pain: a crucial step toward personal pain medicineCan J Anaesth201562329430325471684
- UhlGRSoraIWangZThe mu opiate receptor as a candidate gene for pain: polymorphisms, variations in expression, nociception, and opiate responsesProc Natl Acad Sci U S A199996147752775510393893
- BranfordRDroneyJRossJROpioid genetics: the key to personalized pain control?Clin Genet201282430131022780883
- LeeMGKimHJLeeKHChoiYSThe influence of genotype polymorphism on morphine analgesic effect for postoperative pain in childrenKorean J Pain2016291343926839669
- WalterCLotschJMeta-analysis of the relevance of the OPRM1 118A>G genetic variant for pain treatmentPain2009146327027519683391
- BeyerAKochTSchroderHSchulzSHolltVEffect of the A118G polymorphism on binding affinity, potency and agonist-mediated endocytosis, desensitization, and resensitization of the human mu-opioid receptorJ Neurochem200489355356015086512
- MogilJSPain genetics: past, present and futureTrends Genet201228625826622464640
- BirdAPerceptions of epigeneticsNature2007447714339639817522671
- BuchheitTVan de VenTShawAEpigenetics and the transition from acute to chronic painPain Med201213111474149022978429
- CrowMDenkFMcMahonSBGenes and epigenetic processes as prospective pain targetsGenome Med2013521223409739
- UshijimaTNakajimaTMaekitaTDNA methylation as a marker for the past and futureJ Gastroenterol200641540140716799880
- HwangCKSongKYKimCSEvidence of endogenous mu opioid receptor regulation by epigenetic control of the promotersMol Cell Biol200727134720473617452465
- OertelBGDoehringARoskamBGenetic-epigenetic interaction modulates μ-opioid receptor regulationHum Mol Genet201221214751476022875838
- ChorbovVMTodorovAALynskeyMTCiceroTJElevated levels of DNA methylation at the OPRM1 promoter in blood and sperm from male opioid addictsJ Opioid Manag20117425826421957825
- NielsenDAYuferovVHamonSIncreased OPRM1 DNA methylation in lymphocytes of methadone-maintained former heroin addictsNeuropsychopharmacology200934486787318650805
- von BaeyerCLNumerical rating scale for self-report of pain intensity in children and adolescents: recent progress and further questionsEur J Pain200913101005100719766028
- BringuierSDadureCRauxODuboisAPicotMCCapdevilaXThe perioperative validity of the visual analog anxiety scale in children: a discriminant and useful instrument in routine clinical practice to optimize postoperative pain managementAnesth Analg2009109373774419690240
- MacraeWADaviesHTOChronic Postsurgical PainSeattleIASP Press1999
- GerbershagenHJRothaugJKalkmanCJMeissnerWDetermination of moderate-to-severe postoperative pain on the numeric rating scale: a cut-off point analysis applying four different methodsBr J Anaesth2011107461962621724620
- ChidambaranVMooreDSpruanceKPredicting the pain continuum after adolescent idiopathic scoliosis surgery – a prospective cohort studyEur J Pain2017
- BursacZGaussCHWilliamsDKHosmerDWPurposeful selection of variables in logistic regressionSource Code Biol Med200831719087314
- ENCODE Project ConsortiumAn integrated encyclopedia of DNA elements in the human genomeNature20124897414577422955616
- BernsteinBEStamatoyannopoulosJACostelloJFThe NIH Roadmap Epigenomics Mapping ConsortiumNat Biotechnol201028101045104820944595
- LiuTOrtizJATaingLCistrome: an integrative platform for transcriptional regulation studiesGenome Biol2011128R8321859476
- GriffonABarbierQDalinoJvan HeldenJSpicugliaSBallesterBIntegrative analysis of public ChIP-seq experiments reveals a complex multi-cell regulatory landscapeNucleic Acids Res2015434e2725477382
- ErnstJKheradpourPMikkelsenTSMapping and analysis of chromatin state dynamics in nine human cell typesNature20114737345434921441907
- KochCMAndrewsRMFlicekPThe landscape of histone modifications across 1% of the human genome in five human cell linesGenome Res200717669170717567990
- BarskiACuddapahSCuiKHigh-resolution profiling of histone methylations in the human genomeCell2007129482383717512414
- CreyghtonMPChengAWWelsteadGGHistone H3K27ac separates active from poised enhancers and predicts developmental stateProc Natl Acad Sci U S A201010750219312193621106759
- DenkFMcMahonSBTraceyIPain vulnerability: a neurobiological perspectiveNat Neurosci201417219220024473267
- WachmanEMHayesMJLesterBMEpigenetic variation in the mu-opioid receptor gene in infants with neonatal abstinence syndromeJ Pediatr2014165347247824996986
- HwangCKSongKYKimCSEpigenetic programming of mu-opioid receptor gene in mouse brain is regulated by MeCP2 and Brg1 chromatin remodelling factorJ Cell Mol Med2009139B3591361519602036
- KnotheCOertelBGUltschAPharmacoepigenetics of the role of DNA methylation in μ-opioid receptor expression in different human brain regionsEpigenomics20168121583159927685027
- PorrecaFTangQBBianDRiedlMEldeRLaiJSpinal opioid mu receptor expression in lumbar spinal cord of rats following nerve injuryBrain Res19987951–21972039622629
- VietCTDangDYeYOnoKCampbellRRSchmidtBLDemethylating drugs as novel analgesics for cancer painClin Cancer Res201420184882489324963050
- ZhouXLYuLNWangYIncreased methylation of the MOR gene proximal promoter in primary sensory neurons plays a crucial role in the decreased analgesic effect of opioids in neuropathic painMol Pain2014105125118039
- SunYSahbaiePLiangDDNA methylation modulates nociceptive sensitization after incisionPLoS One20151011e014204626535894
- DoehringAOertelBGSittlRLotschJChronic opioid use is associated with increased DNA methylation correlating with increased clinical painPain20131541152323273101
- DenkFMcMahonSBChronic pain: emerging evidence for the involvement of epigeneticsNeuron201273343544422325197
- FormisanoLNohKMMiyawakiTMashikoTBennettMVZukinRSIschemic insults promote epigenetic reprogramming of mu opioid receptor expression in hippocampal neuronsProc Natl Acad Sci U S A2007104104170417517360495
- BruceAWDonaldsonIJWoodICGenome-wide analysis of repressor element 1 silencing transcription factor/neuron-restrictive silencing factor (REST/NRSF) target genesProc Natl Acad Sci U S A200410128104581046315240883
- SchoenherrCJAndersonDJThe neuron-restrictive silencer factor (NRSF): a coordinate repressor of multiple neuron-specific genesScience19952675202136013637871435
- BuckleyNJJohnsonRZuccatoCBithellACattaneoEThe role of REST in transcriptional and epigenetic dysregulation in Huntington’s diseaseNeurobiol Dis2010391283920170730
- UchidaHMaLUedaHEpigenetic gene silencing underlies C-fiber dysfunctions in neuropathic painJ Neurosci201030134806481420357131
- BanksCJJoshiAMichoelTFunctional transcription factor target discovery via compendia of binding and expression profilesSci Rep201662064926857150
- FanSZhangXCpG island methylation pattern in different human tissues and its correlation with gene expressionBiochem Biophys Res Commun2009383442142519364493
- DaviesMNVoltaMPidsleyRFunctional annotation of the human brain methylome identifies tissue-specific epigenetic variation across brain and bloodGenome Biol2012136R4322703893
- AndreaeMHAndreaeDARegional anaesthesia to prevent chronic pain after surgery: a Cochrane systematic review and meta-analysisBr J Anaesth2013111571172023811426
- KiefferBLEvansCJOpioid tolerance-in search of the holy grailCell2002108558759011893329
- LawPYLohHHWeiLNInsights into the receptor transcription and signaling: implications in opioid tolerance and dependenceNeuropharmacology200447Suppl 130031115464146