
Abstract
The ultimate goal of regenerative medicine is to replace damaged tissues with new functioning ones. This can potentially be accomplished by stem cell transplantation. While stem cell transplantation for blood diseases has been increasingly successful, widespread application of stem cell therapy in the clinic has shown limited results. Despite successful efforts to refine existing methodologies and to develop better ones for reprogramming, clinical application of stem cell therapy suffers from issues related to the safety of the transplanted cells, as well as the low efficiency of reprogramming technology. Better understanding of the underlying mechanism(s) involved in pluripotency should accelerate the clinical application of stem cell transplantation for regenerative purposes. This review outlines the main decision-making factors involved in pluripotency, focusing on the role of microRNAs, epigenetic modification, signaling pathways, and toll-like receptors. Of special interest is the role of toll-like receptors in pluripotency, where emerging data indicate that the innate immune system plays a vital role in reprogramming. Based on these data, we propose that nongenetic mechanisms for reprogramming provide a novel and perhaps an essential strategy to accelerate application of regenerative medicine in the clinic.
Current status of regenerative medicine
Humans have a constrained ability to regenerate and restore their tissues and organs. Some organs have a higher regenerative capacity, such as the blood and the liver, while other organs have very limited ability to self-renew, such as the heart and the brain. Some tissues, such as the liver, regenerate by proliferation, while others undergo self-renewal, such as hematopoietic stem cells. In contrast with humans, some vertebrates have far-reaching regenerative capacities that, in specific cases, extend as far as replacing complete limbs.Citation1
The field of regenerative medicine aims to cure intractable illness by replacing damaged tissues and failing organs, and/or fortifying the body’s own repair mechanisms. Regenerative medicine also incorporates tissue engineering, through which scaffolds, cells, and biologically active molecules are combined into functional tissues. This powerful capacity can conceivably overcome the hurdles of organ transplantation, including the shortage of organs available for donation and the severe problems associated with graft rejection or graft versus host disease.Citation2
Stem cell therapy aims to regenerate malfunctioning tissues via several mechanisms.Citation3 Injectable stem cells may replace defective cells and differentiate into functioning ones. Alternatively, stem and progenitor cells can induce regeneration by secreting biologically active molecules. Furthermore, stem cells can be differentiated in vitro into functioning cells and grown on scaffolds into functioning organs prior to transplantation.Citation4
By taking advantage of biotechnological advances in scaffolding material, tissue engineering appears to be today’s promising treatment for replacing damaged tissues. The principle is straightforward: cells are gathered and introduced with or without alteration of their biological properties directly into the harmed tissue or into a permeable three-dimensional matrix. In these carefully designed scaffolds, stem cells are manipulated in a controlled environment in which physicochemical and mechanical parameters are carefully monitored. After reaching the desired differentiation, these cells or tissues could be grafted.
Dedifferentiation, transdifferentiation, and reprogramming; the three processes compared
Dedifferentiation is one of the mechanisms linked to natural regeneration, in which a terminally differentiated cell returns back to a less differentiated stage from within its own particular genealogy. This procedure permits the cell to proliferate again before redifferentiating, prompting the substitution of those cells that have been lost. A few nonmammalian vertebrates have a remarkable ability to regenerate. As a rule, this process includes the dedifferentiation of mature cells followed by redifferentiation. In nonmammalian vertebrates, there are several illustrations of dedifferentiation, such as heart regeneration in zebrafish. Zebrafish can completely regenerate their heart following amputation of up to 20% of the ventricle.Citation5 Throughout this process, differentiated cardiomyocytes that are still present in the heart dedifferentiate and proliferate to recover the missing tissue.Citation6 As the cardiomyocytes dismantle their contractile apparatus, they additionally detach from each other and begin to express the positive cell cycle regulators monopolar spindle 1, polo-like kinase 1, and cdc2 ().Citation6 At present, little is known about the signaling pathways involved, despite the fact that fibroblast growth factor and platelet-derived growth factor signaling have been specifically linked to this regeneration. Hindrance of fibroblast growth factor or platelet-derived growth factor signaling prompts incomplete regeneration and formation of scar tissue.Citation7 In this case, dedifferentiation seems to be unpredictably linked to the cell cycle, as re-entry into the cell cycle does not appear to be vital following dedifferentiation.Citation8 Recent evidence suggests that the tumor suppressor retinoblastoma protein not only controls cell cycle arrest but might also play an active role in maintaining the differentiated status of a cell. Interestingly, blocking proliferation by blocking retinoblastoma protein does not prevent dedifferentiation from happening. It appears that dedifferentiation is independent of the cell cycle, and that retinoblastoma protein likewise has an important role in keeping up the differentiated status of a cell.Citation9
Figure 1 Review of reprogramming, dedifferentiation, and transdifferentiation. Pluripotent cells have the potential to give rise to any lineage of cells. Fibroblasts can be reprogrammed back to pluripotent cells by inducing expression of Oct4, Sox2, Klf4, and c-Myc.Citation24 Zebrafish cardiomyocytes provide a good example of dedifferentiation by expressing mps1, plk1, and cdc2.Citation6 B-cells can be transdifferentiated into macrophages utilizing CEBPα and CEBPβ.Citation14 Also, fibroblasts can be transdifferentiated into cardiomyocytes utilizing GATA4, Tbx5, and Mef2c.Citation17
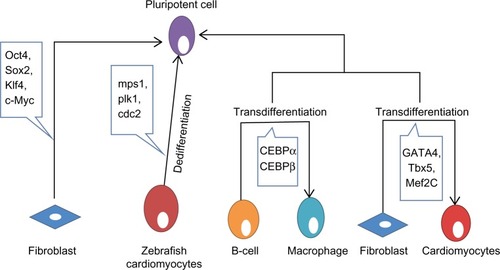
Transdifferentiation involves the irreversible change of cells from one differentiated cell type to another. Biological transdifferentiation takes place in two stages: first, the cell dedifferentiates, then the natural developmental program is stimulated, permitting the cell to differentiate into the new lineage.Citation10 This sequence, however, may not be mandatory in all cases. The regenerating lens of the newt and salamander perfectly depicts the naturally occurring transdifferentiation process.Citation11 In these species, after evacuation of the lens from the eye, new pigmented epithelial cells of the lens regenerate and further differentiate to structure a new lens.Citation11 This regeneration involves inactivation of retinoblastoma protein, permitting the cells to re-emerge the cell cycle.Citation12 Recently, Day and Beck showed that Wnt and bone morphogenetic protein signaling pathways are required for transdifferentiation from cornea to lens, a process in which both the PITX and Wnt genes are essential.Citation13 The experimental transdifferentiation of B-cells into macrophages utilizing the transcription factors CCAAT-enhancer-binding protein-α (CEBPα) and CEBPβ provides a good illustration of in vitro transdifferentiation ().Citation14 Forced expression of CEBPβ and CEBPα in differentiated B-cells prompts reprogramming them into macrophages without notable changes in DNA methylation.Citation15 Following a comparable strategy, it is conceivable to transdifferentiate murine fibroblasts into functional neurons using three transcription factors (Brn2, Ascl1, and Mytl1).Citation16 Fibroblasts have been transdifferentiated into cardiomyocytes utilizing GATA4, Tbx5, and Mef2c ().Citation17 GATA4 activates the process by opening chromatin, permitting access to the other transcription factors.Citation18 This offers a new strategy to replace the lost or damaged cardiomyocytes by inducing fibroblast transdifferentiation in vivo. Human dermal fibroblasts have additionally been transdifferentiated into multipotent progenitors that have the potential to differentiate into monocytic, granulocytic, erythroid, and megakaryocytic lineages with in vivo engraftment capacity utilizing the ectopic expression of Oct4 in the presence of particular cytokine treatment.Citation19 A therapeutic example of transdifferentiation has been shown by Sapir et al, when human liver cells were induced to transdifferentiate into functional insulin-producing β-cells by transduction with PDX-1 virus.Citation20 In an effort to determine the underlying mechanisms of transdifferentiation, a recent report showed that stepwise epigenetic modifications stabilize the changes caused by transcriptional factors to ensure transdifferentiation.Citation21 Zuryn et al have further shown that there is parallelism between epigenetic modifications underlying transdifferentiation in vivo and efficient cell reprogramming in vitro.Citation22
Reprogramming occurs naturally during fertilization to produce totipotent cells that can differentiate into any cell type.Citation23 Reprogramming can also be induced artificially, whereby a fully differentiated cell is induced to regress into its pluripotent origin, with the possibility of differentiating into almost any other cell type. In 2006, Takahashi and Yamanaka induced pluripotency of a differentiated somatic cell through overexpression of only four transcription factors (Oct4, Sox2, c-Myc, and Klf4, ).Citation24 These cells were called induced pluripotent stem cells (iPSCs). Oct4 has a fundamental role in the development and maintenance of pluripotency by activating or repressing specific genes,Citation25 such as Nanog,Citation26 Fgf4,Citation27 Utf1,Citation28 and Zfp206.Citation29 Indeed, cells lacking functional Oct4 can no longer support this key property.Citation30 In addition to activating pluripotency, Oct4 prevents the differentiation of pluripotent cells by acting as a depressor of lineage-specific transcription factors. Oct4 forms a repressive complex with Cdx2 (a transcription factor important for trophectoderm specification), and hinders its transcription.Citation31 Moreover, Oct4 has been shown to suppress the transcription of the epithelial mesenchymal transition mediator Snail to encourage reprogramming.Citation32 Recently, Oct4 was found to interact with nuclear β-catenin and to enhance its proteasomal degradation, thus maintaining the undifferentiated state of embryonic stem cells (ESCs).Citation33 Despite the fact that Sox2 is involved in the self-renewal of stem cells, one of its essential capacities is to maintain Oct4 expression at the levels required to maintain pluripotency.Citation34 It has been shown that Sox2 may act with Oct4 to activate Oct-Sox enhancers that in turn upregulate the expression of many pluripotency genes, including Nanog, Oct4, and Sox2 itself.Citation34 During development, Nanog is expressed by the pluripotent cells of the inner cell mass, and appropriately, embryos that fail to express this gene fail to develop the inner cell mass.Citation35 Nonetheless, if Nanog is removed from cells that have effectively accomplished pluripotency, they still hold many stem cell features.Citation36 Subsequently, it appears that the major role of Nanog is in sustaining pluripotency. Interestingly, the cells most adjustable to reprogramming have been neural stem cells, which only require Oct4.Citation37 Why certain cell types require fewer factors for reprogramming remains debatable. One possibility is that less differentiated cells are closer to pluripotency and subsequently need fewer factors. Another possibility is that certain cell types already express some of these factors. However, the strongest evidence points to the fundamental role of Oct4 in reprogramming. A recent study showed that Tet-1, a 5-methylcytosine hydroxylase, could replace Oct4 in reprogramming somatic cells and generating fully pluripotent iPSCs,Citation38 which indicates that DNA methylation and hydroxymethylation play important roles in epigenetic remodeling in pluripotency.
All of the three processes discussed so far can prompt terminally differentiated cells to become highly plastic and suitable for therapeutic purposes However, one of the major differences between reprogramming and both transdifferentiation and dedifferentiation is their current potential for in vivo therapy. Although both transdifferentiation and dedifferentiation can be successfully accomplished in vivo, directing pluripotent cells into a new lineage is a complex process, that has so far been successful only in vitro. Although the three processes can induce considerable changes in differentiated cells, each has definite advantages in terms of regenerative medicine. If the aim is to replace cells lost as a consequence of injury or disease, this could be accomplished by reprogramming cells taken from the patient in vitro and afterward differentiating them into the desired cell type, followed by engraftment back into the patient. A simpler in vivo approach would be either to induce cells to dedifferentiate and then proliferate or to induce a more abundant or less specialized cell type to transdifferentiate into the desired cell types. If, however, the objective is to correct a genetic mutation, attempting to transdifferentiate or dedifferentiate any of the patient’s cells would not cure the disease, as the new cells would still contain the mutation. In this situation, the more feasible option would be to reprogram the patient’s own cells in vitro, followed by correcting the damaged gene before differentiating the cells into the desired lineage and injecting them back to the patient.Citation39
MicroRNAs involved in pluripotency
The role of microRNAs (miRNAs) in pluripotency has been investigated by several laboratories. MiRNAs are small noncoding RNAs, typically comprising 19–25 nucleotides, that regulate gene expression post transcriptionally by translational silencing of their targeted messenger RNAs (mRNAs).Citation40 In most cases, the effect of miRNAs in ESC physiology remains unknown. However, the presence of a subset of miRNAs that are only expressed in ESCs () and, most importantly, the failure to create viable ESCs from mice deficient in Dicer,Citation41 the enzyme needed for miRNA processing, suggest an essential role for miRNAs in ESC self-renewal.
Table 1 MicroRNAs involved in pluripotency, self-renewal, differentiation, and reprogramming
DGCR8 is a nuclear protein, essential for miRNA processing. Like the Dicer-null ESCs, DGCR8-deficient ESCs show either delayed or decreased expression of differentiation markers, in addition to delayed kinetics of cell cycle progression.Citation42 Most DGCR8-deficient ESCs are arrested in the G1 phase. This indicates that the fundamental role of the miRNA pathway is to regulate the ESC cycle at the G1-S phase transition. In addition, DGCR8-null ESCs display differentiation defects as they fail to steadily silence the expression of self-renewal markers, including OcT4, Sox2, Nanog, and Rex1.Citation42
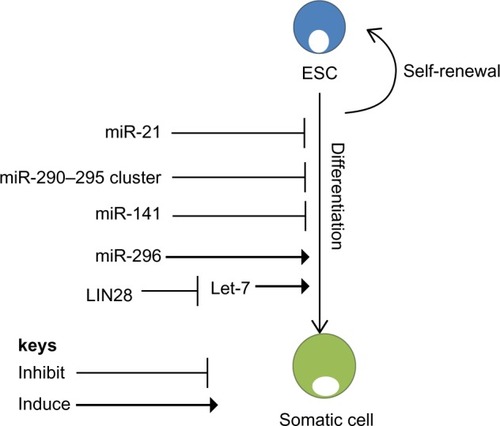
The pattern of expression of miRNA in human ESCs seems to vary extraordinarily from that of differentiated cells.Citation43 Studies showed that the miR-290–295 cluster and miR-296 are peculiar to ESCs and play specific roles in maintaining pluripotencyCitation44 and that their levels are reduced as the stem cells differentiate ().Citation44 In contrast, levels of miR-21 and miR-22 increase significantly after induction of differentiation, indicating that these miRNAs may play a critical role in stem cell differentiation ().Citation43 Other miRNAs, including miR-134, miR-296, and miR-470, were recently found to target coding regions of Oct4, Sox2 and Nanog to promote differentiation.Citation45 Suh et al reported a group of miRNAs that are abundantly expressed in human ESCs. These miRNAs, termed ESC-specific miRNAs, incorporate miR-302a, miR-302b, miR-302c, miR-302d, miR-367, and miR-371–373 ().Citation43 Xu et al showed a critical role for miR-145 in repression of the 3′-untranslated regions of Oct4, Sox2, and Klf4. Loss of miR-145 ameliorated differentiation and increased the expression of the most essential reprogramming factors, indicating its potential role in the creation of iPSCs.Citation46
Figure 2 microRNA regulation of self-renewal and differentiation. ESCs have the potential to self-renew or differentiate into somatic cells. MicroRNAs regulate both self-renewal and differentiation pathways of ESCs. miR-21 and the miR-290–295 cluster are essential for self-renewal of ESCs.Citation43 Another regulator of ESCs is Lin28, which binds to Let-7, promoting self-renewal.Citation51 However, miR-296 promotes differentiation of ESCs.Citation45
Another seemingly important regulator of pluripotency is survivin. Kapinas et al showed that survivin is selectively expressed under the control of miRNAs.Citation47 They showed that miR-203 expression and activity is harmonious with the expression of survivin isoforms and subsequently controls expression of the Oct4 and Nanog transcription factors to maintain pluripotency.Citation47
Efforts to identify the mRNA targets that underlie the capacity of ESC-specific miRNAs have been undertaken and reported by several groups.Citation48 Subramanyam et al demonstrated that miRNAs stimulate reprogramming by simultaneously targeting several discriminating cellular pathways involving regulators of the G1-S cell cycle checkpoint, the mesenchymal-to-epithelial transition, and DNA methyl binding proteins.Citation49 These cellular processes interact with a subset of the mRNAs targeted by these miRNAs. The ESC-specific miRNAs likewise target mRNAs that control apoptosis and bone morphogenetic protein signaling, although the role of these targets in reprogramming has not yet been elucidated.Citation50
Despite these reports, regulators of miRNA biogenesis have been demonstrated to be among the handful of factors that can change differentiated cells into iPSCs. This is the case with the RNA-binding protein Lin28, which can proficiently change fibroblasts into iPSCs when transduced together with Oct4, Sox2, and Nanog.Citation51 Evidently, Lin28 is an ESC-specific RNA-binding protein that interferes with and suppresses the action of let-7 miRNAs.Citation52 let-7 miRNA is expressed at low levels in ESCs, and is rapidly induced upon differentiation.Citation53 Additionally, Lin28 expression is activated by c-Myc in numerous human and mouse tumor models ().Citation54 These data provide evidence that Lin28 has a central role in blocking miRNA-mediated differentiation in stem cells and in substitution of c-Myc throughout iPSC formation. Evidently, using miRNAs to control the differentiation of stem cells can prompt diverse tissues to be generated from stem cells for the purpose of cell therapy. For instance, expression of miR-181 results in an increase in the number of cells entering the B-lymphoid lineage, but not the number of cells entering the T-lymphoid lineage, while ectopic expression of miR-142 or miR-223 has the opposite effect.Citation55
Epigenetic modifications in pluripotency
Pluripotency of ESCs is maintained by epigenetic factors closely associated with the pluripotency transcription factor network ().Citation56 Epigenetic modifications of gene expression include DNA methylation and histone modification, each of which adjusts how genes are expressed without modifying the underlying DNA sequence. Epigenetic factors involved in maintaining the pluripotency of ESCs must be activated throughout the reprogramming process,Citation57 as during reprogramming, pluripotency genes that are hypermethylated in somatic cells must be demethylated and activated. The genomes of ESCs express an open chromatin state that is highly accessible to transcription factors.Citation58 The open chromatin status is locally stimulated by chromatin-remodeling factors, such as the SWI-SNF complex and Chd1. Knockdown of Chd1 blocks the iPSC, while overexpression of the SWI-SNF complex enhances its formation.Citation59,Citation60
Table 2 List of epigenetic markers and their role in reprogramming
Histone modifications
During iPSC generation, somatic cell chromatin requires reorganization to an ESC-like state.Citation61 It seems that the chromatin reorganization event occurs in a coordinated and sequential manner. Rearrangement of heterochromatin, distinguished by the presence of histone H3 lysine 9 trimethylation (H3K9me3) and heterochromatin protein 1, occurs before activation of Nanog, while enrichment of euchromatin markers happens simultaneously with activation of Nanog.Citation62 Pluripotent ESCs are known for active histone markers, including H3K4me3, but as ESCs differentiate, the regions marked by repressive H3K9me3 and H3K27me3 extend and spread ().Citation63 H3K9 methylation is a tangible barrier to reprogramming, so decreasing the levels of H3K9 methyltransferases or overexpressing H3K9 demethylases enhances the efficiency of creating iPSCs ().Citation64,Citation65 ESCs thus express bivalent histone modifications, ie, H3K4me3 and H3K27me3, which hinder many differentiation genes.Citation66
DNA methylation
Like histone modifications, DNA methylations are well established in ESCs and iPSCs.Citation67 Methylation is mediated by DNA methyltransferases, which stimulate the addition of Met groups to Cys residues in CpG islands. In general, regions rich in CpG that overlap gene promoters are unmethylated and their expression is decreased.Citation68 Notwithstanding this, many promoters in CpG islands undergo methylation changes during differentiation, like promoters of pluripotency genes, which are free from methylation in undifferentiated ESCs and iPSCs, but become methylated at differentiation.Citation69 Indeed, for ESCs to differentiate, DNA demethylation is needed as DNA methyltransferase-depleted cells conserve their stem cell characteristics, indicating that DNA methylation is essential for the maintenance of stemness.Citation70
Chromatin remodeling
DNA is packaged into chromatin by associations with histone and non-histone proteins. The main unit of chromatin is the nucleosome, structured by the wrapping of 146 base pairs of DNA around a core of two copies, each composed of four histone proteins (H2A, H2B, H3, and H4).Citation71 Pluripotent ESCs are characterized by a global chromatin structure that is generally dynamic and specific to the transcriptional machinery. Upon differentiation, chromatin is changed into a more repressive state.Citation58 Components of the chromatin structure also play a key role in controlling gene expression profiles in pluripotent ESCs. Direct interaction between reprogramming factors and chromatin regulators may be essential for pluripotency. For example, Oct4 can interact with subunits of the BAF chromatin-remodeling complex,Citation25,Citation72 which improves reprogramming and could stimulate the binding of transcription factors to nucleosomal sites.Citation59 Also, it has been shown that overexpression of Baf155/Brg1 (an ATP-dependent chromatin-remodeling complex) enhances reprogramming by enhancing the binding of Oct4 to its pluripotency targets.Citation59 Additionally, Chd1, the chromatin-remodeling factor, is required to maintain the open chromatin state of pluripotent stem cells and is also required for efficient reprogramming of fibroblasts to the pluripotent stem cell state.Citation60 Thus, somatic cell reprogramming using chromatin-remodeling molecules represents an efficient method of generating reprogrammed cells.
Recent work in iPSC programming showed that if cells are deficient in Mbd3, a nuclear protein of the Mbd3/NuRD (nucleosome remodeling and acetylation) complex, the efficiency of reprogramming induced by Oct4, Sox2, Klf4, and c-Myc increases to nearly 100%.Citation73 It is also worth pointing out that the peremptory reprogramming of Mbd3-depleted somatic cells reported by Brumbaugh et al has been challenged by a more recent study showing a need for functioning of Mbd3/NuRD for efficient reprogramming.Citation73 Santos et al have showed that depletion of Mbd3 led to reduction in the efficiency of reprogramming and that overexpression of Mbd3 facilities reprogramming of neural stem cells when combined with expression of Nanog, but not with other tested reprogramming factors.Citation74 It seems that a number of differences, including choice of reprogramming cassettes and reprogramming culture conditions, have influenced the differences between these results. Also, 5-azacytidine, a DNA methyltransferase inhibitor, encourages transformation of partially reprogrammed cells to iPSCs, and upgrades overall reprogramming efficiency when used at the late stage of reprogramming.Citation75
The ability to establish patient-matched ESC lines is currently challenged by the fact that there are abnormal epigenetic modifications during the reprogramming process. The above data thus provide strong evidence that epigenetic modifications have significant control over pluripotency, and should be considered when using iPSCs for cell therapy.
Signaling pathways involved in pluripotency
Wnt/β-catenin signaling pathway
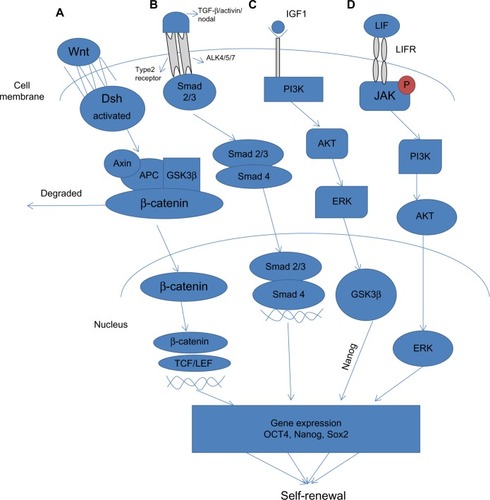
The Wnt signaling pathways function through groups of surface proteins, such as Wnt, which has been linked to embryonic development. β-catenin, a part of the cadherin cell adhesion complex, assumes an essential role in E-cadherin-mediated cell-cell adhesion and is an essential intermediate in the Wnt signaling pathway. It is well established that human and murine ESCs can be maintained in an undifferentiated state by the activation of Wnt signaling, which upregulates the expression of Oct4, Nanog, and Rex1 through the action of β-catenin ().Citation76 The role of Wnt signaling in the reprogramming processes is confounding since high levels of Wnt activity in glycogen synthase kinase (GSK)-3 double-knockout mice were shown to prompt differentiation of ESCs.Citation77 (GSK-3 is a serine/threonine protein kinase that has been demonstrated to phosphorylate β-catenin, thus targeting it for recession).
Figure 3 Signaling pathways involved in pluripotency. (A) Wnt signaling pathway is stimulated via binding of Wnt proteins with frizzled receptors. This subsequently activates Dsh, which replaces GSK-3β from the Axin-APC complex, preventing degradation of β-catenin. Subsequently, β-catenin translocates into the nucleus where it associates with TCF/LEF proteins to activate transcription of Wnt target genes that cause self-renewal. (B) TGF-β signals through two main receptors, ie, the type II receptor and the Activin receptor. Upon activation, ligand-specific Smad proteins (Smad2/3) are phosphorylated and form a complex with Smad4, which translocates to the nucleus to activate gene expression. (C) The PI3K/Akt pathway is activated by IGF-1. Activation of PI3K activates Akt, which modulates the threshold of Smad2/3 activity and inhibits Erk and maintains GSK-3β activity, which in turn induces expression of Nanog. (D) The Jak/Stat signal pathway is principally initiated by LIF, which likewise initiates PI3K through which Akt and Erk are activated, ultimately resulting in upregulation of expression of Klf4 and Myc.
Wnt has been definitively shown to regulate dedifferentiation. Without Wnt signaling, β-catenin is phosphorylated by functional interaction with GSK-3β and hence targeted to degradation by the ubiquitin-proteasome system.Citation78 Another putative “reprogrammer” enacted by Wnt signaling might be T-cell factor/lymphocyte enhancer factor, which indicates a high level of colocalization with Oct4, Sox2, and Nanog ().Citation79 Additionally, Wnt could activate endogenous c-Myc directly, substituting the need for exogenous c-Myc. This supports the evidence that Wnt signaling can promote reprogramming of somatic cells to pluripotency.Citation80
TGF-β signaling pathway
Transforming growth factor-beta (TGF-β) is a protein that controls proliferation and cellular differentiation in different capacities in most cells. The signaling role of TGF-β in stem cell reprogramming has been investigated by Ichida et al, who showed that inhibiting TGF-β signaling replaces Sox2 in reprogramming by inducing the transcription of Nanog ().Citation81 It is well established that bone morphogenic protein 4, a member of the TGF-β superfamily, participates with leukemia inhibitory factor in maintaining pluripotency in murine ESCs. Under serum-free culture conditions, leukemia inhibitory factor alone prompted neural differentiation of murine ESCs. This effect was annulled by treatment with bone morphogenic protein 4, which maintained the undifferentiated state of ESCs, and without serum.Citation82
In 2009, Maherali et al demonstrated that hindrance of TGF-β receptor I kinase with an Activin-like kinase 5 inhibitor upgraded both the proficiency and kinetics of murine embryonic fibroblasts reprogrammed by Oct4, Sox2, Klf4, and c-Myc. Activation of the TGF-β signaling pathway on the other hand blocked reprogramming.Citation83 This discrepancy suggests that hindrance of TGF-β acts only on direct reprogramming pathways that are already working in ESCs. Inhibition of TGF-β could replace Sox2 and c-Myc in induction of iPSCs. This highlights the dual role of TGF-β as an inducer of pluripotency by replacing Sox2 and c-Myc.Citation83 Inhibition of TGF-β signaling also promotes the mesenchymal-to-epithelial transition process, which is a critical step in somatic reprogramming.Citation32 TGF-β prevents mesenchymal-to-epithelial transition by inhibiting both upregulation of epithelial markers and downregulation of the mesenchymal transcriptional repressor Snail.Citation32
PI3K/Akt signaling pathway
Phosphoinositide-3-kinases (PI3Ks) are lipid kinases that induce generation of phosphatidylinositol trisphosphate, a signaling lipid, upon stimulation by numerous growth factor receptor tyrosine kinases, such as epidermal growth factor, fibroblast growth factor, and platelet-derived growth factor. Akt1, a serine/threonine kinase, is one of the fundamental players in this pathway. It modulates the functions of different cellular responses, including cell proliferation, growth, adhesion, and death, in addition to inducing tumorigenesis.Citation84 Activation of Akt1 signaling is sufficient to maintain the pluripotency of murine ESCs without leukemia inhibitory factor ().Citation85 However, despite what might be expected, treatment of murine ESCs with the PI3K inhibitor LY294002 promoted a loss of ESC characteristics, even in the presence of leukemia inhibitory factor.Citation86 Further, initiation of PI3K signaling is critical to the advancement of dedifferentiation in embryonic germ cells from primordial germ cells.Citation87 The role of the PI3K pathway in ESC biology and in reprogramming remains largely unknown. Nakamura et al showed that activation of Akt signaling stimulated reprogramming after fusion of ESCs with thymocytes or murine embryonic fibroblasts, which prompted the formation of ESC-like hybrid cells.Citation88 Supporting these results, it has been shown that ERas, an ESC-specific Ras gene, is closely associated with Akt in enhancing reprogrammingCitation89 and modulation of Akt signaling by genetic or chemical means.Citation89
Jak-Stat signaling pathway
The Jak-Stat signaling pathway transmits information from chemical signals outside the cell through the cell membrane and into gene promoters in the DNA in the nucleus, causing DNA transcription and activity. Activation of the Janus kinase/signal transducer and activator of transcription 3 (Jak/Stat3) signaling pathway maintains murine ESC pluripotency.Citation90 The Jak/Stat3 signaling pathway is principally initiated by leukemia inhibitory factor, which likewise initiates the PI3K/protein kinase B (also known as Akt) and extracellular signal-regulated kinase 1/2 (Erk1/2) pathways in ESCs ().Citation91 Artificially activated Stat3 sustains self-renewal of murine ESCs in the absence of leukemia inhibitory factor.Citation92 Little is known about the downstream mechanism of Stat3 in maintenance of pluripotency, despite reports that leukemia inhibitory factor/Stat3 upregulates expression of Klf4 and Myc in murine ESCs.Citation93
Activin/Nodal pathway
Nodal, a member of the TGF-β superfamily, acts by binding to heteromeric complexes between type I and type II Activin receptors, which in turn act through the Smad2/Smad3 signaling pathway.Citation94 Recent data indicates that overexpression of the Nodal growth factor in human ESCs can obstruct their default neuroectoderm differentiation by formation of embryoid bodies.Citation95 Nodal itself is expressed in human ESCs and disappears rapidly upon differentiation, indicating that Nodal signaling could be included in the maintenance of pluripotency. Restraint of Nodal does not induce differentiation of human ESCs. However, hindrance of the Activin/Nodal/TGF-β signaling pathway induces differentiation of human ESCs, showing that the Activin/Nodal pathway is fundamental for support of pluripotency.Citation96 Evidently, understanding the molecular mechanisms and signaling pathways that regulate stemness is a mandatory step toward the design of rational clinical treatments.
Toll-like receptors involved in pluripotency
Toll-like receptors (TLRs) are a class of proteins with a key role in the innate immune response. TLR signaling comprises two different pathways, ie, a myeloid differentiation primary response gene (MyD 88)-dependent pathway and a MyD88-independent pathway.Citation97 The MyD88-dependent pathway is common to all TLRs, except for TLR3.Citation98 The TLR3 pathway is initiated by viral double-stranded RNA and is independent of MyD88. To investigate the role of this pathway in reprogramming using retroviral plasmid-containing stem cell factors (Oct4, Sox2, Klf4, and c-Myc), knockdown of TLR3 signaling diminished the pluripotency gene expression induced by Oct4. Similarly, short hairpin RNA knockdown of TLR3 diminished the ability of Oct4 to induce expression of the target pluripotent genes.Citation99 This indicates that TLR3 knockdown inhibits activation of downstream pluripotency genes when utilizing retroviral vectors or mRNA to overexpress the reprogramming factors and reduces the efficiency and yield of human iPSCs. Taken together, stimulation of TLR3 produces substantial changes in the expression of epigenetic modifiers to enhance chromatin rebuilding and nuclear reprogramming and stimulates pluripotency genes and generation of human iPSC colonies.Citation99
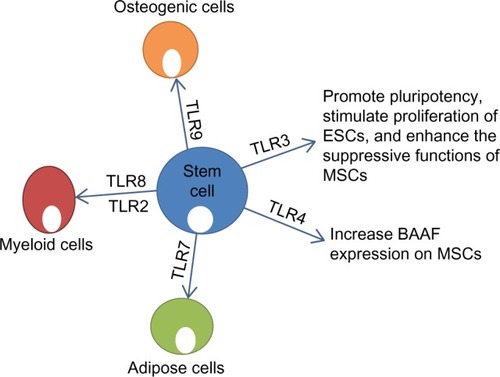
Emerging data suggest that TLRs may play a role in regulation of stem cell fate. TLR ligands have been shown to contribute to differentiation of mesenchymal stem cells (MSCs). Supplementation of adipogenic differentiation medium with TLR7 and TLR2 agonists induced MSCs to produce a significantly high percentage of adipose cells, with more promising results achieved with TLR7 ().Citation100 Also, TLR9 increased the osteogenic differentiation of MSCs ().Citation100 Human bone marrow hematopoietic stem cells could be differentiated into myeloid cells just by stimulation with TLR7/8 agonists.Citation101 TLR2 agonists have been shown to have similar effects in hematopoietic stem cell differentiation into the myeloid lineage ().Citation101 Interestingly, TLR7/8 agonists were more effective at inducing CD11+CD14− cells, whereas TLR2 was more effective at inducing CD11+CD14+ cells.Citation101 On the basis of these findings, it can be suggested that TLR signaling can influence the outcome of stem cell differentiation, and that differentiation is not produced through the same signaling pathways.
Figure 4 Effect of stimulation of TLRs on function and differentiation of stem cells. TLR3 plays a role in maintaining pluripotency and enhancing the function of MSCs.Citation99 TLR4 increases expression of BAAF on MSCs. TLR7 induces adipogenic differentiation of MSCs.Citation98 TLR8/2 induces production of myeloid cells from HSCs.Citation100 TLR9 induces osteogenic differentiation of MSCs.Citation98
Human MSCs have been shown to produce proinflammatory cytokines, such as interleukin (IL)-1β, IL-6, IL-8, CCL5, and tumor necrosis factor-alpha, in response to TLR agonists.Citation102 TLR2 agonists stimulated CD34+ progenitors to secrete granulocyte-macrophage colony-stimulating factor.Citation101 It has been shown that production of cytokines in response to TLR stimulation occurs through nuclear factor kappa B signaling.Citation103 MSCs from diverse sources express TLRs at the mRNA level, although expression at a protein level appears to be low (ie, compared with monocytes). Detection of TLRs in MSCs by flow cytometry has also been troublesome.Citation104 Pevsner-Fischer et al demonstrated that murine MSCs express many TLRs, and specifically TLR2, which was essential for their differentiation capacity.Citation105 Like murine MSCs, human MSCs also express many TLRs.Citation106 In murine MSCs, TLR signaling may be connected to the multipotency of MSCs, as MyD88-deficient bone marrow MSCs failed to efficiently differentiate into chondrogenic and osteogenic lineages.Citation105 In human MSCs, only activation of TLR9 has been shown to affect adipogenic differentiation.Citation100
Giuliani et al have shown that TLRs can modulate the function of MSCs.Citation107 TLR-primed adult and ESCs were more resistant than unprimed stem cells to natural killer-induced killing.Citation107 In addition, TLR3-primed but not TLR4-primed MSCs showed enhanced suppressive functions against natural killer cells.Citation107 On the other hand, Yan et al showed that TLR4 priming of MSCs increased the expression of B-cell activating factor,Citation108 which indicates that TLR4 has an important role in B-lymphocyte immune regulation of MSCs ().
Lee et al observed that TLR ligands such as lipopolysaccharide, a ligand of TLR4, and Poly (I:C), a ligand of TLR3, stimulated the proliferation of murine ESCs (ESD3) developed on gelatin-coated dishes in standard ESC culture medium containing leukemia inhibitory factor ().Citation99 The TLR ligands, lipopolysaccharide and Poly (I:C), were likewise found to induce ESCs to form primary embryoid bodies.Citation109 Evidently, the cells from the lipopolysaccharide-stimulated primary embryoid bodies were still capable of forming the secondary embryoid bodies more efficiently in the presence of lipopolysaccharide, suggesting that lipopolysaccharide-stimulated ESCs retain competence for self-renewal.Citation110
TLR signaling plays a similarly significant role in promoting differentiation of murine ESCs into hematopoietic progenitors.Citation109 Adding lipopolysaccharide to the ESC hematopoietic differentiation culture favorably increased the differentiation of myeloid lineage progenitors.Citation109 Also, treatment with the TLR4 ligand stimulated hematopoietic progenitors to differentiate from Twist-2-deficient ESCs,Citation111 indicating the important role of the transcriptional repressor Twist-2 in controlling hematopoietic differentiation from ESCs.Citation109
It appears that TLRs and their ligands can serve as regulators of stem cell proliferation and differentiation and might affect the maintenance of MSC multipotency. Our understanding of the biological significance of functional TLRs in stem cells and their role in dedifferentiation, transdifferentiation, and reprogramming seem to be evolving, and further investigation is warranted.
Conclusion and perspectives
One of the most significant challenges in regenerative medicine is being able to provide transplantable cells.Citation112 Stem cell transplantation has shown impressive results in the treatment of a number of intractable diseases, such as leukemia, lymphoma, and immune deficiency disorders. However, use of stem cells for solid tissue transplantation has not been met with similar success.Citation113 Issues with safety and production efficiency have hampered the progress and clinical applications of stem cell therapy, especially utilizing iPSCs.Citation114 Evidently, the stress generated during reprogramming prompts the selection of iPSCs in which stress regulatory genes are mutated, rendering iPSCs more prone to tumor development.Citation115 In this review, we have attempted to explain the role of miRNAs, epigenetic modifications, signaling pathways, and TLRs in pluripotency and the potential application of this knowledge in facilitating stem cell therapy.
The role of ESC-specific miRNAs in effective reprogramming of ESCs looks very promising. ESC-specific miRNAs alone can accomplish successful reprogramming, leading to the conclusion that these miRNAs have an important role in the process. The primary advantage of miRNAs is that, unlike transcription factors, they are more effective and take less time to induce cell redirection. However, the mechanism by which ESC-specific miRNAs redirect somatic cells to assume pluripotency remains unknown. miRNA-based reprogramming may herald a new era in regenerative medicine, and the introduction of miRNAs directly into patients to change the fate of certain cells does seem highly promising.
Epigenetic modifications have been shown to be closely connected to the pluripotent state of ESCs, as in the case of H3K4me3, which was shown to support the self-renewal capacity of ESCs.Citation116 Further understanding of the regulatory connections between epigenetic modifications and pluripotency will aid in the generation of “high-quality” iPSCs suitable for therapeutic application.
Although the signaling pathways involved in dedifferentiation, transdifferentiation, and reprogramming have not been completely elucidated, recent evidence suggests that these pathways may have a critical role in application of stem cell therapy.Citation117 The Wnt/β-catenin, TGF-β, PI3K/Akt, Activin/Nodal, and Jak-Stat signaling pathways all have an essential role in pluripotency.Citation118 Recognizing which signaling pathways play a key role during dedifferentiation, transdifferentiation, and reprogramming could conceivably allow development of new approaches to control these processes through better control of the process of pluripotency. By activating or inhibiting certain signaling pathways, pluripotency can be turned on and off.
Understanding of the molecular mechanisms that control pluripotency or stimulate differentiation will enhance application of stem cell therapy in the clinical setting. Of special importance is the role of TLRs in embryonic and adult stem cells, with new reports suggesting that TLRs may play a dual activation and regulatory role in the immune system. This role may be more pronounced in regulation of mammalian stem cells, and highlights a previously unrecognized role for activation of innate immunity in pluripotency. These emerging data should have significant implications for understanding stem cell biology and applying stem cell therapy in the clinical setting.
Despite the large volume of research in the field, regenerative medicine has shown slow progress at the bedside, largely because of issues related to the safety of ESCs and iPSCs and the limited efficiency of reprogramming technology. Most efforts to reprogram somatic cells to behave like stem cells have focused on genetic modifications. Data presented in this review highlight important, novel, and nongenetic factors that can significantly alter somatic cells, enhance pluripotency, and provide an efficient means for reprogramming and stem cell therapy.
Acknowledgments
The Center of Excellence for Stem Cells and Regenerative Medicine is funded by a grant (number 5300) from The Science and Technology Development Fund in Egypt.
Disclosure
The authors report no conflicts of interest in this work.
References
- BrockesJPKumarAPlasticity and reprogramming of differentiated cells in amphibian regenerationNat Rev Mol Cell Biol20023856657412154368
- MasonCDunnillPA brief definition of regenerative medicineRegen Med2008311518154457
- RiaziAMKwonSYStanfordWLStem cell sources for regenerative medicineMethods Mol Biol2009482559019089350
- MuneokaKAllanCHYangXLeeJHanMMammalian regeneration and regenerative medicineBirth Defects Res C Embryo Today200884426528019067422
- PossKDWilsonLGKeatingMTHeart regeneration in zebrafishScience200229856012188219012481136
- JoplingCSleepERayaMMartiMRayaAIzpisua BelmonteJCZebrafish heart regeneration occurs by cardiomyocyte dedifferentiation and proliferationNature2010464728860660920336145
- LienCLSchebestaMMakinoSWeberGJKeatingMTGene expression analysis of zebrafish heart regenerationPLoS Biol200648e26016869712
- OdelbergSJInducing cellular dedifferentiation: a potential method for enhancing endogenous regeneration in mammalsSemin Cell Dev Biol200213533534312324215
- NicolayBNBayarmagnaiBMoonNSBenevolenskayaEVFrolovMVCombined inactivation of pRB and hippo pathways induces dedifferentiation in the Drosophila retinaPLoS Genet201064e100091820421993
- GrafTEnverTForcing cells to change lineagesNature2009462727358759419956253
- TsonisPAMadhavanMTancousEEDel Rio-TsonisKA newt’s eye view of lens regenerationInt J Dev Biol2004488–997598015558488
- ThitoffARCallMKDel Rio-TsonisKTsonisPAUnique expression patterns of the retinoblastoma (Rb) gene in intact and lens regeneration-undergoing newt eyesAnat Rec A Discov Mol Cell Evol Biol2003271118518812552633
- DayRCBeckCWTransdifferentiation from cornea to lens in Xenopus laevis depends on BMP signalling and involves upregulation of Wnt signallingBMC Dev Biol2011115421896182
- XieHYeMFengRGrafTStepwise reprogramming of B cells into macrophagesCell2004117566367615163413
- Rodriguez-UbrevaJCiudadLGomez-CabreroDPre-B cell to macrophage transdifferentiation without significant promoter DNA methylation changesNucleic Acids Res20114051954196822086955
- KimJEfeJAZhuSDirect reprogramming of mouse fibroblasts to neural progenitorsProc Natl Acad Sci USA2011108197838784321521790
- IedaMFuJDDelgado-OlguinPDirect reprogramming of fibroblasts into functional cardiomyocytes by defined factorsCell2010142337538620691899
- CirilloLALinFRCuestaIFriedmanDJarnikMZaretKSOpening of compacted chromatin by early developmental transcription factors HNF3 (FoxA) and GATA-4Mol Cell20029227928911864602
- SzaboERampalliSRisuenoRMDirect conversion of human fibroblasts to multilineage blood progenitorsNature2010468732352152621057492
- SapirTShternhallKMeivar-LevyICell-replacement therapy for diabetes: generating functional insulin-producing tissue from adult human liver cellsProc Natl Acad Sci USA2005102227964796915899968
- BeyretEIzpisua BelmonteJCWorming toward transdifferentiation, one (epigenetic) step at a timeDev Cell201430664164225268170
- ZurynSAhierAPortosoMTransdifferentiation. Sequential histone-modifying activities determine the robustness of transdifferentiationScience2014345619882682925124442
- de VriesWNEvsikovAVBroganLJReprogramming and differentiation in mammals: motifs and mechanismsCold Spring Harb Symp Quant Biol200873333819022741
- TakahashiKYamanakaSInduction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factorsCell2006126466367616904174
- PardoMLangBYuLAn expanded Oct4 interaction network: implications for stem cell biology, development, and diseaseCell Stem Cell20106438239520362542
- RoddaDJChewJLLimLHTranscriptional regulation of Nanog by OCT4 and SOX2J Biol Chem200528026247312473715860457
- AmbrosettiDCBasilicoCDaileyLSynergistic activation of the fibroblast growth factor 4 enhancer by Sox2 and Oct-3 depends on protein-protein interactions facilitated by a specific spatial arrangement of factor binding sitesMol Cell Biol19971711632163299343393
- NishimotoMFukushimaAOkudaAMuramatsuMThe gene for the embryonic stem cell coactivator UTF1 carries a regulatory element which selectively interacts with a complex composed of Oct-3/4 and Sox-2Mol Cell Biol19991985453546510409735
- WangZXTehCHKuehJLLufkinTRobsonPStantonLWOct4 and Sox2 directly regulate expression of another pluripotency transcription factor, Zfp206, in embryonic stem cellsJ Biol Chem200728217128221283017344211
- NicholsJZevnikBAnastassiadisKFormation of pluripotent stem cells in the mammalian embryo depends on the POU transcription factor Oct4Cell19989533793919814708
- NiwaHToyookaYShimosatoDInteraction between Oct3/4 and Cdx2 determines trophectoderm differentiationCell2005123591792916325584
- LiRLiangJNiSA mesenchymal-to-epithelial transition initiates and is required for the nuclear reprogramming of mouse fibroblastsCell Stem Cell201071516320621050
- Abu-RemailehMGersonAFaragoMOct-3/4 regulates stem cell identity and cell fate decisions by modulating Wnt/beta-catenin signallingEMBO J200229193236324820736927
- MasuiSNakatakeYToyookaYPluripotency governed by Sox2 via regulation of Oct3/4 expression in mouse embryonic stem cellsNat Cell Biol20079662563517515932
- SilvaJNicholsJTheunissenTWNanog is the gateway to the pluripotent ground stateCell2009138472273719703398
- ChambersISilvaJColbyDNanog safeguards pluripotency and mediates germline developmentNature200745071731230123418097409
- KimJBGreberBArauzo-BravoMJDirect reprogramming of human neural stem cells by OCT4Nature2009461726464964319718018
- GaoYChenJLiKReplacement of Oct4 by Tet1 during iPSC induction reveals an important role of DNA methylation and hydroxymethylation in reprogrammingCell Stem Cell201312445346923499384
- JoplingCBoueSIzpisua BelmonteJCDedifferentiation, transdifferentiation and reprogramming: three routes to regenerationNat Rev Mol Cell Biol2010122798921252997
- YingSYChangDCMillerJDLinSLThe microRNA: overview of the RNA gene that modulates gene functionsMethods Mol Biol200634211816957363
- BernsteinBEMikkelsenTSXieXA bivalent chromatin structure marks key developmental genes in embryonic stem cellsCell2006125231532616630819
- WangYMedvidRMeltonCJaenischRBlellochRDGCR8 is essential for microRNA biogenesis and silencing of embryonic stem cell self-renewalNat Genet200739338038517259983
- SuhMRLeeYKimJYHuman embryonic stem cells express a unique set of microRNAsDev Biol2004270248849815183728
- HoubaviyHBMurrayMFSharpPAEmbryonic stem cell-specific microRNAsDev Cell20035235135812919684
- TayYZhangJThomsonAMLimBRigoutsosIMicroRNAs to Nanog, Oct4 and Sox2 coding regions modulate embryonic stem cell differentiationNature200845572161124112818806776
- XuNPapagiannakopoulosTPanGThomsonJAKosikKSMicroRNA-145 regulates OCT4, SOX2, and KLF4 and represses pluripotency in human embryonic stem cellsCell2009137464765819409607
- KapinasKKimHMandevilleMmicroRNA-mediated survivin control of pluripotencyJ Cell Physiol20152301637024891298
- JudsonRLBabiarzJEVenereMBlellochREmbryonic stem cell-specific microRNAs promote induced pluripotencyNat Biotechnol200927545946119363475
- SubramanyamDLamouilleSJudsonRLMultiple targets of miR-302 and miR-372 promote reprogramming of human fibroblasts to induced pluripotent stem cellsNat Biotechnol201129544344821490602
- LipchinaIElkabetzYHafnerMGenome-wide identification of microRNA targets in human ES cells reveals a role for miR-302 in modulating BMP responseGenes Dev201125202173218622012620
- YuJVodyanikMASmuga-OttoKInduced pluripotent stem cell lines derived from human somatic cellsScience200731858581917192018029452
- NewmanMAThomsonJMHammondSMLin-28 interaction with the Let-7 precursor loop mediates regulated microRNA processingRNA20081481539154918566191
- KumarMSErkelandSJPesterRESuppression of non-small cell lung tumor development by the let-7 microRNA familyProc Natl Acad Sci USA2008105103903390818308936
- ChangTCZeitelsLRHwangHWLin-28B transactivation is necessary for Myc-mediated let-7 repression and proliferationProc Natl Acad Sci USA200910693384338919211792
- ChenCZLiLLodishHFBartelDPMicroRNAs modulate hematopoietic lineage differentiationScience20043035654838614657504
- OrkinSHHochedlingerKChromatin connections to pluripotency and cellular reprogrammingCell2011145683585021663790
- YoungRAControl of the embryonic stem cell stateCell2011144694095421414485
- MeshorerEMisteliTChromatin in pluripotent embryonic stem cells and differentiationNat Rev Mol Cell Biol20067754054616723974
- SinghalNGraumannJWuGChromatin-remodeling components of the BAF complex facilitate reprogrammingCell2010141694395520550931
- Gaspar-MaiaAAlajemAPolessoFChd1 regulates open chromatin and pluripotency of embryonic stem cellsNature2009460725786386819587682
- FussnerEDjuricUStraussMConstitutive heterochromatin reorganization during somatic cell reprogrammingEMBO J20113091778178921468033
- MattoutABiranAMeshorerEGlobal epigenetic changes during somatic cell reprogramming to iPS cellsJ Mol Cell Biol20113634135022044880
- WenBWuHShinkaiYIrizarryRAFeinbergAPLarge histone H3 lysine 9 dimethylated chromatin blocks distinguish differentiated from embryonic stem cellsNat Genet200941224625019151716
- ChenJLiuHLiuJH3K9 methylation is a barrier during somatic cell reprogramming into iPSCsNat Genet2013451344223202127
- SoufiADonahueGZaretKSFacilitators and impediments of the pluripotency reprogramming factors’ initial engagement with the genomeCell20121515994100423159369
- ZhaoXDHanXChewJLWhole-genome mapping of histone H3 Lys4 and 27 trimethylations reveals distinct genomic compartments in human embryonic stem cellsCell Stem Cell20071328629818371363
- SmithZDMeissnerADNA methylation: roles in mammalian developmentNat Rev Genet201314320422023400093
- BartkeTVermeulenMXhemalceBRobsonSCMannMKouzaridesTNucleosome-interacting proteins regulated by DNA and histone methylationCell2010143347048421029866
- FarthingCRFiczGNgRKGlobal mapping of DNA methylation in mouse promoters reveals epigenetic reprogramming of pluripotency genesPLoS Genet200846e100011618584034
- JacksonMKrassowskaAGilbertNSevere global DNA hypomethylation blocks differentiation and induces histone hyperacetylation in embryonic stem cellsMol Cell Biol200424208862887115456861
- HenikoffSAhmadKAssembly of variant histones into chromatinAnnu Rev Cell Dev Biol20052113315316212490
- van den BergDLSnoekTMullinNPAn Oct4-centered protein interaction network in embryonic stem cellsCell Stem Cell20106436938120362541
- BrumbaughJHochedlingerKRemoving reprogramming roadblocks: Mbd3 depletion allows deterministic iPSC generationCell Stem Cell201313437938124094318
- dos SantosRLTostiLRadzisheuskayaAMBD3/NuRD facilitates induction of pluripotency in a context-dependent mannerCell Stem Cell201415110211024835571
- MikkelsenTSHannaJZhangXDissecting direct reprogramming through integrative genomic analysisNature20084547200495518509334
- SatoNMeijerLSkaltsounisLGreengardPBrivanlouAHMaintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitorNat Med2004101556314702635
- YingJLiHYuJWNT5A exhibits tumor-suppressive activity through antagonizing the Wnt/beta-catenin signaling, and is frequently methylated in colorectal cancerClin Cancer Res2008141556118172252
- PolakisPThe adenomatous polyposis coli (APC) tumor suppressorBiochim Biophys Acta199713323F127F1479196022
- TamWLLimCYHanJT-cell factor 3 regulates embryonic stem cell pluripotency and self-renewal by the transcriptional control of multiple lineage pathwaysStem Cells20082682019203118467660
- MarsonAForemanRChevalierBWnt signaling promotes reprogramming of somatic cells to pluripotencyCell Stem Cell20083213213518682236
- IchidaJKBlanchardJLamKA small-molecule inhibitor of TGF-beta signaling replaces Sox2 in reprogramming by inducing NanogCell Stem Cell20095549150319818703
- YingQLNicholsJChambersISmithABMP induction of Id proteins suppresses differentiation and sustains embryonic stem cell self-renewal in collaboration with STAT3Cell2003115328129214636556
- MaheraliNHochedlingerKTGF-beta signal inhibition cooperates in the induction of iPSCs and replaces Sox2 and cMycCurr Biol200919201718172319765992
- CantleyLCThe phosphoinositide 3-kinase pathwayScience200229655731655165712040186
- WatanabeSUmeharaHMurayamaKOkabeMKimuraTNakanoTActivation of Akt signaling is sufficient to maintain pluripotency in mouse and primate embryonic stem cellsOncogene200625192697270716407845
- LiuNLuMFengXMExogenous Nanog alleviates but is insufficient to reverse embryonic stem cells differentiation induced by PI3K signaling inhibitionJ Cell Biochem200910661041104719229869
- KimuraTTomookaMYamanoNAKT signaling promotes derivation of embryonic germ cells from primordial germ cellsDevelopment2008135586987918216173
- NakamuraTInoueKOgawaSEffects of Akt signaling on nuclear reprogrammingGenes Cells200813121269127719032345
- YuYLiangDTianQStimulation of somatic cell reprogramming by ERas-Akt-FoxO1 signaling axisStem Cells201432234936323765875
- NiwaHBurdonTChambersISmithASelf-renewal of pluripotent embryonic stem cells is mediated via activation of STAT3Genes Dev19981213204820609649508
- NiwaHOgawaKShimosatoDAdachiKA parallel circuit of LIF signalling pathways maintains pluripotency of mouse ES cellsNature2009460725111812219571885
- MatsudaTNakamuraTNakaoKSTAT3 activation is sufficient to maintain an undifferentiated state of mouse embryonic stem cellsEMBO J199918154261426910428964
- HallJGuoGWrayJOct4 and LIF/Stat3 additively induce Kruppel factors to sustain embryonic stem cell self-renewalCell Stem Cell20095659760919951688
- SchierAFNodal signaling in vertebrate developmentAnnu Rev Cell Dev Biol20031958962114570583
- VallierLReynoldsDPedersenRANodal inhibits differentiation of human embryonic stem cells along the neuroectodermal default pathwayDev Biol2004275240342115501227
- InmanGJNicolasFJCallahanJFSB-431542 is a potent and specific inhibitor of transforming growth factor-beta superfamily type I activin receptor-like kinase (ALK) receptors ALK4, ALK5, and ALK7Mol Pharmacol2002621657412065756
- KawaiTAkiraSThe role of pattern-recognition receptors in innate immunity: update on Toll-like receptorsNat Immunol201011537338420404851
- KaishoTAkiraSToll-like receptor function and signalingJ Allergy Clin Immunol2006117597998716675322
- LeeJSayedNHunterAActivation of innate immunity is required for efficient nuclear reprogrammingCell2012151354755823101625
- Hwa ChoHBaeYCJungJSRole of toll-like receptors on human adipose-derived stromal cellsStem Cells200624122744275216902195
- Romieu-MourezRFrancoisMBoivinMNBouchentoufMSpanerDEGalipeauJCytokine modulation of TLR expression and activation in mesenchymal stromal cells leads to a proinflammatory phenotypeJ Immunol2009182127963797319494321
- SioudMFloisandYTLR agonists induce the differentiation of human bone marrow CD34+ progenitors into CD11c+ CD80/86+ DC capable of inducing a Th1-type responseEur J Immunol200737102834284617853407
- ZhaoJLMaCO’ConnellRMConversion of danger signals into cytokine signals by hematopoietic stem and progenitor cells for regulation of stress-induced hematopoiesisCell Stem Cell201414444545924561084
- van den BerkLCJansenBJSiebers-VermeulenKGToll-like receptor triggering in cord blood mesenchymal stem cellsJ Cell Mol Med2009139B3415342620196781
- Pevsner-FischerMMoradVCohen-SfadyMToll-like receptors and their ligands control mesenchymal stem cell functionsBlood200710941422143217038530
- TomchuckSLZwezdarykKJCoffeltSBWatermanRSDankaESScandurroABToll-like receptors on human mesenchymal stem cells drive their migration and immunomodulating responsesStem Cells20082619910717916800
- GiulianiMBennaceur-GriscelliANanbakhshATLR ligands stimulation protects MSC from NK killingStem Cells201432129030024123639
- YanHWuMYuanYWangZZJiangHChenTPriming of Toll-like receptor 4 pathway in mesenchymal stem cells increases expression of B cell activating factorBiochem Biophys Res Commun2014448221221724780395
- LeeSHHongBSharabiAHuangXFChenSYEmbryonic stem cells and mammary luminal progenitors directly sense and respond to microbial productsStem Cells20092771604161519544467
- ChanRJJohnsonSALiYYoderMCFengGSA definitive role of Shp-2 tyrosine phosphatase in mediating embryonic stem cell differentiation and hematopoiesisBlood200310262074208012791646
- SharabiABLeeSHGoodellMAHuangXFChenSYEnhanced generation of myeloid lineages in hematopoietic differentiation from embryonic stem cells by silencing transcriptional repressor Twist-2Cloning Stem Cells200911452353320025523
- BhagavatiSStem cell therapy: challenges aheadIndian J Pediatr201582328629124992980
- OwakiTShimizuTYamatoMOkanoTCell sheet engineering for regenerative medicine: current challenges and strategiesBiotechnol J20149790491424964041
- MedvedevSPShevchenkoAIZakianSMInduced pluripotent stem cells: problems and advantages when applying them in regenerative medicineActa Naturae201022182822649638
- DuinsbergenDSalvatoriDErikssonMMikkersHTumors originating from induced pluripotent stem cells and methods for their preventionAnn N Y Acad Sci2009117619720419796248
- OkanoMBellDWHaberDALiEDNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian developmentCell199999324725710555141
- ShoniMLuiKOVavvasDGProtein kinases and associated pathways in pluripotent state and lineage differentiationCurr Stem Cell Res Ther20149536638724998240
- LiuKSongYYuHZhaoTUnderstanding the roadmaps to induced pluripotencyCell Death Dis20145e123224832604
- MeltonCJudsonRLBlellochROpposing microRNA families regulate self-renewal in mouse embryonic stem cellsNature2010463728162162620054295
- MeltonCBlellochRMicroRNA regulation of embryonic stem cell self-renewal and differentiationAdv Exp Med Biol201169510511721222202
- Anokye-DansoFTrivediCMJuhrDHighly efficient miRNA-mediated reprogramming of mouse and human somatic cells to pluripotencyCell Stem Cell20118437638821474102
- AngYSTsaiSYLeeDFWdr5 mediates self-renewal and reprogramming via the embryonic stem cell core transcriptional networkCell2011145218319721477851
- BuganimYFaddahDAChengAWSingle-cell expression analyses during cellular reprogramming reveal an early stochastic and a late hierarchic phaseCell201215061209122222980981
- MansourAAGafniOWeinbergerLThe H3K27 demethylase Utx regulates somatic and germ cell epigenetic reprogrammingNature2012488741140941322801502
- LiangGHeJZhangYKdm2b promotes induced pluripotent stem cell generation by facilitating gene activation early in reprogrammingNat Cell Biol201214545746622522173
- OnderTTKaraNCherryAChromatin-modifying enzymes as modulators of reprogrammingNature2012483739159860222388813
- HuangfuDMaehrRGuoWInduction of pluripotent stem cells by defined factors is greatly improved by small-molecule compoundsNat Biotechnol200826779579718568017
- LiangGTaranovaOXiaKZhangYButyrate promotes induced pluripotent stem cell generationJ Biol Chem201028533255162552120554530