Abstract
Antiretroviral drugs have saved and extended the lives of millions of individuals infected with HIV. The major classes of anti-HIV drugs include reverse transcriptase inhibitors, protease inhibitors, integrase inhibitors, and entry/fusion inhibitors. While antiretroviral drug regimens are not commonly used to treat other types of retroviral infections, there are instances where there is a perceived need for re-evaluation of the benefits of antiretroviral therapy. One case in point is that of feline leukemia virus (FeLV), an infection of companion felines. While vaccines exist to prevent FeLV infection and spread, they have not eliminated FeLV infection. For FeLV-infected felines and their human companions, antiretroviral therapy would be desirable and of practical importance if good options were available. Here, we discuss FeLV biology and current treatment options, and propose that there is a need for antiretroviral treatment options for FeLV infection. The comparative use and analysis of antiretroviral therapy can provide new insights into the mechanism of antiretroviral drug action.
Introduction
Retroviruses are a large group of RNA viruses that are found in all vertebrates. They share many common features, such as similarities in genetic organization and mechanism of replication, and in particular for their encoding for a reverse transcriptase. These viruses are a significant source of morbidity and mortality in both humans and animals. In humans, HIV is responsible for a world-wide pandemic that continues to be a significant cause of morbidity and mortality worldwide. In animals, feline leukemia virus (FeLV) represents a significant source of mortality and is the leading cause of infectious deaths in cats. Besides its role in feline mortality, FeLV has played an important role in advancing the understanding of retroviruses in general. In fact, the characterization of FeLV as well as other animal retroviruses such as bovine leukemia virus and Rous sarcoma virus, led to the concepts and techniques that later enabled the discovery and characterization of human retroviruses including HIV.Citation1 Additionally, FeLV was 1) the first retrovirus in which a vaccine was developed, 2) the first retrovirus for which a practical diagnostic test was developed, and 3) the first retrovirus that elicited the development of a program whose goal was to control its spread.Citation1 Despite the significant achievements and understanding of FeLV biology, FeLV is still a significant source of morbidity and mortality in felines and the treatment options for infected cats are ineffective, toxic, or cost-prohibited. Here, we discuss FeLV epidemiology, pathogenicity, and current treatments as well as future drug targets that may advance the field of FeLV treatment.
Epidemiology
FeLV is highly transmissible through saliva and nasal secretions as well as through coitus and vertical transmission from queen to kitten. Because the primary mode of transmission is through the oronasal route, its prevalence is often dictated by the extent of animal-to-animal contact. For example, the high prevalence of FeLV in the stray cat population (40%) increases the risk of infection for indoor–outdoor cats compared to indoor-only cats. Similarly, animal-to-animal contact contributes to the different rates of FeLV infection in single cat households (4%–11%) compared to multicat households whose prevalence has been reported to be as high as 70%.Citation2,Citation4,Citation5
Containment of FeLV is difficult due to transmission routes, the time between infection and the onset of symptoms, and the ability of latently infected cats to become viremic. In fact, exposure of cats to FeLV usually leads to one of three outcomes, two of which can contribute to the spread of disease.Citation2–Citation8 The first outcome is accounted for by the 10% of exposed cats that become latently infected without a detectable viremia. While these cats would not seem to be a source of infection, they can become viremic, and subsequently shed virus into the environment. The second outcome is represented by 40% of cats exposed to FeLV, and is characterized by persistent viremia and antigenemia. These cats are chronically infected, and therefore represent a significant source of viral shedding in the environment. The third outcome includes cats that become infected with FeLV, but are then able to clear the virus to the point where it is undetectable by standard testing methods. Approximately 50% of cats exposed to FeLV fall into this last group and these cats are not considered to be a reservoir for viral spread.Citation2,Citation4
FeLV replication
FeLV was first described by Jarrett et al in 1964 who isolated viral particles from lymphomas obtained from infected cats.Citation9 Using electron microscopy, Jarrett et al described the infectious agent as being similar in appearance to murine leukemia virus (MuLV). Later studies confirmed FeLV to be a retrovirus. FeLV is a “simple” retrovirus, in that it encodes for 3 genes common to all retroviruses (gag, pol, and env), but lacks many of the additional genes found in complex retroviruses such as HIV.Citation6,Citation7,Citation10 As with other retroviruses, the gag gene encodes for structural proteins while the pol gene encodes for the enzymatic proteins necessary for reverse transcription of the FeLV genome, integration of its DNA into the host genome, and processing of viral proteins. Finally, env encodes for 2 envelope proteins that determine cellular tropism, including p15E, a transmembrane protein, and the associated external envelope protein, gp70.Citation1 Amino acid variation in the virus envelope protein has led to the division of FeLV into 4 different subtypes that defines their cell tropism: A, B, C, and T (see ).Citation11–Citation13 Subtype A is considered to be the founder, transmitted form of FeLV, with all other subtypes arising through mutations in FeLV-A Env or by recombination events with one of the endogenous FeLVs (enFeLV) contained within the cat genome.Citation1,Citation14 The enFeLV has an incomplete genome, is not replication competent, and is theorized to have originated hundreds of thousands of years ago when a cat ate a mouse that was viremic with murine leukemia virus (MuLV). Such an event enabled the incorporation of the MuLV genome into the genome of the cat’s germ line cells.Citation15,Citation16
Table 1 Description of feline leukemia virus (FeLV) subtypes, their tropism, and prevalence in infected cats
Cell-free FeLV gains entry into target cells when the envelope protein binds to the appropriate host receptor that is dependent on the FeLV subtype (). Once inside of the target cell, the RNA genome is reverse transcribed into viral DNA by the viral protein reverse transcriptase.Citation17–Citation20 The viral DNA is then transported into the nucleus where it integrates into the host genome through the enzymatic action of viral integrase. The integrated viral DNA is then transcribed to produce RNA that serves as both viral progeny as well as mRNA for the translation of viral proteins. Translated Gag and Pol proteins are then trafficked to the cell membrane where the new virions bud from the cell membrane. As the virion buds, proteolytic cleavage of viral proteins causes structural changes in the virion that are necessary for viral maturation and the formation of infectious virus particles.Citation17–Citation20
Pathobiology
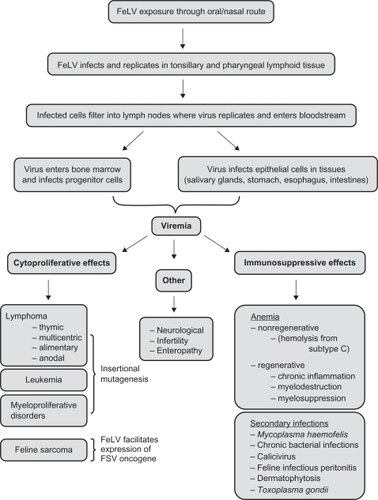
FeLV usually enters the feline host through the oronasal route either through mutual grooming, biting, or a shared food source.Citation21–Citation23 In the pharynx, FeLV infects the tonsillary B-lymphocytes and monocytes which can enter the draining lymph nodes.Citation24 The draining lymph nodes serve as a site of replication and as an entry point for the virus to enter the bloodstream. Once in the bloodstream, the virus can gain access to and infect cells in the bone marrow.Citation1 This represents a critical point in the infection process as it is thought that a persistent infection can be avoided if the immune system can mount an appropriate response before cells in the bone marrow are infected.Citation25 Once the virus becomes systemic, it infects epithelial cells in the intestines, stomach, trachea, and salivary glands and is shed into the environment. Persistently infected cats can demonstrate symptoms of disease anywhere from weeks to years after infection. FeLV-mediated disease typically falls into one of two major categories – cytoproliferative or immunosuppressive.Citation1
Cytoproliferative diseases associated with FeLV include leukemias, lymphomas, fibrosarcoma, and associated myeloproliferative disorders.Citation4,Citation26 Most cytoproliferative diseases are attributed to insertional mutagenesis, a process by which FeLV DNA integrates at a site in the cat’s genome that disrupts or deregulates expression of proteins involved in the regulation of cell cycle, cell survival, or apoptosis.Citation27–Citation32 In contrast, FeLV plays an indirect role in the formation of feline fibrosarcoma. Specifically, fibrosarcoma is caused by dual infection by both FeLV and feline sarcoma virus (FSV). FSV is a replication-defective virus that encodes an oncogene that drives cellular transformation.Citation33–Citation36 Therefore, FeLV serves as a helper virus, providing FSV with the proteins necessary for its replication, thereby allowing for expression of the FSV oncogene.Citation33–Citation36
Besides having cytoproliferative effects, FeLV also mediates a significant loss of immune function. This immune suppression is due to a progressive loss of T and B lymphocytes as well as neutrophils. Immune suppression leads to secondary infections such as bacterial or fungal infections that would not be a significant source of morbidity in an otherwise healthy cat.Citation37–Citation40 Although the exact mechanism of FeLV-mediated immunosuppression is not clear, evidence supports 3 distinct mechanisms of action. First, immune suppression may be a result of FeLV-mediated myeloproliferative disorder.Citation2,Citation14 This disorder leads to an over-proliferation of incompetent mature or immature white blood cells within the bone marrow that eventually overcrowd hematopoietic cells, thereby decreasing red blood cell production and leading to a hindrance of the immune system. Second, the virus may be cytopathic or induce cellular apoptosis, although most evidence indicates that this may be specific for FeLV subtype T. Third, it has been suggested that the transmembrane envelope protein, p15E, may have immunosuppressive properties.Citation41–Citation44 For example, p15E has been reported to inhibit production of mitogenic lymphokines in T cells and has been shown to inhibit lymphocyte function without affecting receptor function.Citation42,Citation43 Additionally, while p15E is not known to be cytotoxic, lymphocyte populations that are exposed to FeLV decline in size.Citation41–Citation44 FeLV-mediated immunosuppression allows for secondary infections such bacterial, parasitic, and other viral infections. One example includes the blood borne parasitic infection, hemobartonellosis, which is seen with the subgroup C infections and results in anemia with a hemolytic aspect.Citation1
Other diseases are associated with FeLV in which the mechanisms remain unclear. For example, neurological diseases and infertility are seen in FeLV-infected cats, though it is not clear how FeLV replication causes these disorders. Also, FeLV-C is known to cause a nonregenerative aplastic anemia. While the mechanism is not clear, it has been suggested that the anemia may be due to FeLV’s use of the heme export receptor, FLVCR1, leading to a toxic accumulation of heme in erythroid progenitor cells and decreasing their numbers.Citation45,Citation46 provides a summary of FeLV pathogenesis.
Figure 1 Pathogenesis of feline leukemia virus (FeLV) showing common entry route of virus, sites of viral replication, and pathological consequences of infection.
Therapeutics: vaccines and limitations
Besides containment of infected cats, commercially available vaccines for FeLV such as Pfizer’s Leukocell® or Merial’s Purevax® are marketed and may significantly reduce FeLV spread and viral reservoir development. These particular vaccines are derived from chemically inactivated antigens or are of recombinant viral origin, respectively, and do not offer sterilizing immunity (ie, these vaccines essentially prime the cat’s immune system enabling it to clear the virus upon exposure).Citation8,Citation47,Citation48 The American Association of Feline Practitioners reserves the vaccination regimen for high-risk populations such as indoor–outdoor cats and catteries rather than including it as part of the core vaccines.
There are several reasons why these FeLV vaccines are likely reserved strictly for the high-risk populations. The link between FeLV vaccine and feline sarcoma is likely to play an important role in why these vaccines are not part of the core vaccines. Vaccine-associated sarcoma (VAS) is an aggressive malignant tumor which requires aggressive surgery and often chemotherapy to treat.Citation2,Citation14,Citation49 The association of VAS is a significant reason why many owners and breeders choose not to vaccinate their cats against FeLV. A second reason why the FeLV vaccine is not part of the core vaccines is that a closed population of indoor-only cats is not susceptible to FeLV, making the risk of VAS greater than the risk of infection. A third factor that might limit the use of the FeLV vaccine is that its efficacy is still not known and is difficult to determine. Further complicating the issue is that the biological response to FeLV exposure differs significantly among cats. For example, 50% of cats clear the virus, 40% become persistently infected, and another 10% become latently infected. Finally, efficacy may need to be determined by using age-matched groups, given that younger cats are more susceptible to a pathogenic infection than older individuals.Citation2,Citation14,Citation50 Analyzing the result from studies that have examined efficacy is difficult because of the differences in the study design, viral strain used, and age at which cats were challenged with virus.
Many owners choose not to vaccinate their cats for fear of VAS and because the efficacy of the vaccine is not clear. The unvaccinated cat population as well as the stray cat population remains at risk for FeLV and is a significant source for FeLV transmission. The prevalence of FeLV in the cat population is evident in veterinary care where there is a significant demand for the treatment of FeLV.
Therapeutics: antiviral drug targets and opportunities
The lack of effective treatment options leads most owners to choose palliative care for FeLV-infected cats. Palliative care may include medications to treat infections, pain management, nutritional support, or any other care with the goal of keeping the cats comfortable and improving their quality of life.
Although some antivirals and immune modulators have been reported to improve the quality or quantity of life for FeLV infected cats, no studies have convincingly shown that any antivirals or immune modulators actually improve the quality or quantity of life for FeLV-infected felines in a clinically useful form. Among antivirals used to treat FeLV, azidothymidine (AZT) was the first antiretroviral used to treat HIV. AZT is a nucleoside analog that becomes incorporated into the viral DNA during reverse transcription of the RNA genome to double-stranded DNA. AZT lacks the 3′ hydroxyl group necessary for DNA polymerization, which results in termination of viral DNA synthesis. Although AZT is the primary antiviral used clinically to treat FeLV, there is little, if any, literature to support its ability to improve the course of the disease once infection is established. Some studies have suggested that treatment of cats 24 hours prior to infection up until 24 hours after infection might delay or minimize infection,Citation51,Citation52 but there have been no well-designed studies to address the efficacy of AZT to prolong the lives of cats with established infection. The studies that indicate that AZT might be efficacious demonstrated that even minor improvements in disease indicators were associated with drug-related toxicities. For example, animals treated with daily doses of 30 and 60 mg/kg had elevated antibody titers, suggesting that their immune system was better able to respond to infection, although all animals receiving these doses demonstrated drug-related toxicities.Citation53 Thus, the main limitation for the use of AZT is its lack of apparent efficacy at tolerable doses.
Similar to antivirals, studies reporting the efficacy of immune modulators, lack appropriate controls and/or have not been independently verified. One of the immune modulators with little to no support for efficacy is lymphocyte T-cell immune modulator (LTCI). LTCI is a protein produced by a thymic stromal epithelial cell line whose manufacturers claim induces cytokines that activate CD8 cytotoxic T cells to attack virally infected cells. It is also claimed that LTCI leads to clinical improvement in FeLV-infected cats. These claims are not documented in the peer-reviewed literature, and the data supplied by the manufacturers do not argue strongly for improvements that would translate into an improvement in the quality of life or length of life for FeLV-infected animals.Citation54
Another immune modulator, inactivated parapox ovis virus, strain D1701 (Baypamun®, Bayer, Leverkusen, Germany), is reported to nonspecifically activate the immune system and improve or cure FeLV-infected cats. Baypamun is reported to increase neutrophil counts and increase the production of interferon, interleukins, and tumor necrosis factor. While initial reports by Hörber and Mayr and Hörber et al reported that Baypamun cured 80% to 100% of FeLV-infected cats,Citation55,Citation56 numerous independent studies have failed to find a difference in clinical response between cats treated with Baypamun and those treated with placebo.Citation57
Staphylococcus protein A (SPA) is perhaps the only immune modulator with evidence to support its ability to improve FeLV-infected cats. SPA is a bacterial polypeptide purified from the cell walls of Staphylococcus aureus Cowan I. Although its mechanism of action is not clear, it has been shown to bind preferentially to IgG in the form of an immune complex rather than its monomeric form. It has been speculated that SPA may bind to IgG that is bound to a “blocking factor” associated with antigen–antibody complexes and that this blocking factor may facilitate tumor or virus escape from immunological control. Therefore, SPA removes antigen–antibody-blocking factor complexes, allowing the immune system to react to viral invasion. Others have suggested that SPA may stimulate the immune system by inducing antibody synthesis, expression of interferon, and by potentiating the natural killer activity of lymphocytes. Although some studies have shown that SPA can improve the life expectancy of FeLV-infected cats, the treatment regimens described in these studies are too cost-prohibitive and demanding to be clinically useful.Citation58 For example, the treatment regimens required whole-body irradiation followed by treatments given twice weekly. Additionally, it took 14 to 45 treatments to clear the virus and even this many treatments did not elicit a response from all of the cats.Citation59 Given the conflicting results on treatment efficacy in addition to the cost and time associated with the treatments, SPA is not a clinically feasible treatment option.
Treatment options, drug targets, and the need for FeLV-based antiretrovirals
The significant morbidity and mortality associated with FeLV combined with the significant knowledge of FeLV biology and availability of antiretrovirals developed for HIV should facilitate the identification and development of new treatment options for FeLV. Perhaps one of the main problems with treating FeLV is that early treatment is almost a prerequisite for success. Since insertional mutagenesis is responsible for FeLV-mediated lymphomas and leukemias, a quick decrease in viral loads would ensure fewer integration events, thereby decreasing the likelihood of oncogenesis. Given the success in anti-HIV drugs at quickly decreasing viral loads, it is reasonable to assume that some of these drugs could be repositioned for the treatment of FeLV. The most likely classes of anti-HIV drugs that could be repositioned include the nucleoside reverse transcriptase inhibitors (NRTIs). Many of the other classes of anti-HIV drugs are structure-based small-molecule inhibitors and would therefore be unlikely to possess anti-FeLV activities. Although there are a number of NRTIs that could be repositioned to treat FeLV, in vitro and in vivo studies indicate that NRTIs may not be readily repositioned to treat FeLV infections. Biochemical studies have shown that reverse transcriptase from oncoretroviruses such as FeLV have a higher fidelity and a significantly lower susceptibility to certain nucleoside analogs compared with lentiviruses.Citation58 The differences in susceptibility to nucleoside analogs between lentiviral reverse transcriptases and oncoretroviral reverse transcriptases suggest that nucleoside analogs used to treat HIV will have different susceptibilities for FeLV.
Conclusion
In human medicine, improvement in treatment options is driven by the demand of those affected by the disease. A prominent example of this demand and supply is seen with conditions such as cancer and HIV, where the impact on society fueled the funding and motivation necessary to rapidly expand treatment options in a short time. Although diseases of companion animals do not have the same affect on society as those that affect humans, there is an increasing awareness of the benefits of companion animals in improving the health and well-being of humans and a trend for demanding better treatment options at whatever cost. Antiretroviral drugs and molecular tools are readily available to determine if drugs used to treat HIV could be repositioned to treat FeLV. An important factor in assessing and instituting these potential treatments is adequate funding of well-designed studies that have sufficient numbers and controls to clearly define the treatment’s efficacy and potential toxicity.
Acknowledgements
Supported by NIH grants R01 GM56615 (LMM and SEP), T32 RR18719 (WMG), and T32 CA09138 (CLC).
Disclosure
The authors report no conflicts of interest.
References
- LevyJARetroviridaeSpringer19932
- CoteEClincial Veterinary Advisor: Dogs and CatsSt LouisMosby Elsevier2007
- EssexMCotterSMCarpenterJLFeline virus-induced tumors and the immune response: recent developmentsAm J Vet Res19733468098124350543
- EttingerSFFeldmanECTextbook of Veterinary Internal Medicine6th edSt. LouisElsevier Saunders2004
- RogersonPJarrettWMackeyLEpidemiological studies on feline leukemia virus infection. I. A serological survey in urban catsInt J Cancer1975155781785166947
- LevyJCrawfordCHartmannKAmerican Association of Feline Practitioners’ feline retrovirus management guidelinesJ Feline Med Surg200810330031618455463
- LutzHAddieDBelákSFeline leukaemia. ABCD guidelines on prevention and managementJ Feline Med Surg200911756557419481036
- SparkesAHFeline leukaemia virus: a review of immunity and vaccinationJ Small Anim Pract19973851871949179815
- JarrettWFCrawfordEMMartinWBDavieFA virus-like particle associated with leukemia (lymphosarcoma)Nature196420256756914195054
- ChenHBechtelMKShiYPathogenicity induced by feline leukemia virus, Rickard strain, subgroup A plasmid DNA (pFRA)J Virol1998729704870569696797
- JarrettOFeline leukaemia virus diagnosisVet Rec1980106245136254235
- JarrettOHardyWDJrGolderMCHayDThe frequency of occurrence of feline leukaemia virus subgroups in catsInt J Cancer1978213334337204584
- SarmaPSLogTSubgroup classification of feline leukemia and sarcoma viruses by viral interference and neutralization testsVirology19735411601694351607
- KahnCThe Merck Veterinary Manual9th edWhitehouse StationMerck & Co, Inc2005
- BenvenisteRESherrCJTodaroGJEvolution of type C viral genes: origin of feline leukemia virusScience1975190421788688852892
- BenvenisteRETodaroGJHomology between type-C viruses of various species as determined by molecular hybridizationProc Natl Acad Sci U S A19737012331633204357865
- CoffinJMGenetic diversity and evolution of retrovirusesCurr Top Microbiol Immunol19921761431641600751
- CoffinJMStructure, replication, and recombination of retrovirus genomes: some unifying hypothesesJ Gen Virol19794217126215703
- CoffinJMStructure and classification of retrovirusesLevyJAThe Retroviridae1New York, NYPlenum Press19921949
- CoffinJMRetroviridae: The viruses and their replicationFieldsBNKnipeDMHowleyPMFields Virology23rd edPhiladelphiaLippincott-Raven199617671847
- RickardCGPostJENoronhaFBarrLMA transmissible virus-induced lymphocytic leukemia of the catJ Nat Cancer Inst196942698710145793196
- EssexMKleinGSnyderSPHarroldJBCorrelation between humoral antibody and regression of tumours induced by feline sarcoma virusNature (Lond)19712331951964939178
- CattoriVTandonRRiondBPepinACLutzHHofmann-LehmannRThe kinetics of feline leukaemia virus shedding in experimentally infected cats are associated with infection outcomeVet Microbiol2009133329229618774240
- RojkoJLHooverEAMathesLEKrakowkaSOlsenRGInfluence of adrenal corticosteroids on the susceptibility of cats to feline leukemia virus infectionCancer Res197939937893791225025
- RojkoJLHooverEAMathesLEOlsenRGSchallerJPPathogenesis of experimental feline leukemia virus infectionJ Natl Cancer Inst1979633759768224237
- HardyWDJrHessPWMacEwenEGMcClellandAJZuckermanEEEssexMCotterSMJarretOBiology of feline leukemia virus in the natural environmentCancer Res1976362 pt 2582588175919
- FujinoYLiaoCPZhaoYSIdentification of a novel common proviral integration site, flit-1, in feline leukemia virus induced thymic lymphomaVirology20093861162219203775
- FujinoYOhnoKTsujimotoHMolecular pathogenesis of feline leukemia virus-induced malignancies: insertional mutagenesisVet Immunol Immunopathol20081231–213814318313764
- FujinoYSatohHHisasueMMasudaKOhnoKTsujimotoHDetection of the integrated feline leukemia viruses in a cat lymphoid tumor cell line by fluorescence in situ hybridizationJ Hered200394325125512816966
- LevyLSLobelle-RichPAOverbaughJflvi-2, a target of retroviral insertional mutagenesis in feline thymic lymphosarcomas, encodes bmi-1Oncogene199387183318388390036
- LevyLSStarkeyCRPrabhuSLobelle-RichPACooperating events in lymphomagenesis mediated by feline leukemia virusLeukemia199711Suppl 32392419209353
- TsujimotoHFultonRNishigakiKA common proviral integration region, fit-1, in T-cell tumors induced by myc-containing feline leukemia virusesVirology199319628458488396812
- SnyderSPTheilenGHTransmissible feline fibrosarcomaNature1969221107410755774407
- NaharroGRobbinsKCReddyEPGene product of v-fgr onc: hybrid protein containing a portion of actin and a tyrosine-specific protein kinaseScience1984223463163666318314
- NaharroGTronickSRRasheedSGardnerMBAaronsonSARobbinsKCMolecular cloning of integrated Gardner-Rasheed feline sarcoma virus: genetic structure of its cell-derived sequence differs from that of other tyrosine kinase-coding onc genesJ Virol19834736116196312085
- NaharroGDunnCYRobbinsKCAnalysis of the primary translational product and integrated DNA of a new feline sarcoma virus, GR-FeSVVirology198312525025076601328
- HardyWDJrMcClellandAJZuckermanEEFeline leukemia virus nonproducer lymphosarcomas of cats as a model for the etiology of human leukemiasHaematol Blood Transfus1981264924946274764
- HooverEAPerrymanLEKocibaGJEarly lesions in cats inoculated with feline leukemia virusCancer Res19733311451524345148
- PerrymanLEHooverEAYohnDSImmunologic reactivity of the cat: immunosuppression in experimental feline leukemiaJ Natl Cancer Inst1972495135713654346537
- TraininZWernickeDUngar-WaronHEssexMSuppression of the humoral antibody response in natural retrovirus infectionsScience198322045998588596302837
- HebebrandLCOlsenRGMathesLENicholsWSInhibition of human lymphocyte mitogen and antigen response by a 15,000-dalton protein from feline leukemia virusCancer Res1979392 Pt 1443447216488
- LafradoLJLewisMGMathesLEOlsenRGSuppression of in vitro neutrophil function by feline leukaemia virus (FeLV) and purified FeLV-p15EJ Gen Virol198768Pt 25075133029288
- MathesLEOlsenRGHebebrandLCImmunosuppressive properties of a virion polypeptide, a 15,000-dalton protein, from feline leukemia virusCancer Res1979393950955218725
- QuackenbushSLMullinsJIHooverEAColony forming T lymphocyte deficit in the development of feline retrovirus induced immunodeficiency syndromeBlood19897325095162537113
- QuigleyJGBurnsCCAndersonMMCloning of the cellular receptor for feline leukemia virus subgroup C (FeLV-C), a retrovirus that induces red cell aplasiaBlood20009531093109910648427
- TailorCSWillettBJKabatDA putative cell surface receptor for anemia-inducing feline leukemia virus subgroup C is a member of a transporter superfamilyJ Virol19997386500650510400745
- Hofmann-LehmannRTandonRBorettiFSReassessment of feline leukaemia virus (FeLV) vaccines with novel sensitive molecular assaysVaccine20062481087109416198454
- SparkesAHFeline leukaemia virus and vaccinationJ Feline Med Surg2003529710012670435
- MadewellBRGiegerTLPesaventoPAKentMSVaccine site-associated sarcoma and malignant lymphoma in cats: a report of six cases (1997–2002)J Am Anim Hosp Assoc2004401475014736905
- FlynnJNHanlonLJarrettOFeline leukaemia virus: protective immunity is mediated by virus-specific cytotoxic T lymphocytesImmunology2000101112012511012762
- NelsonPSellonRNovotneyCTherapeutic effects of diethylcarbamazine and 3′-azido-3′-deoxythymidine on feline leukemia virus lymphoma formationVet Immunol Immunopathol1995461–21811947618257
- TavaresLRonekerCJohnstonKLehrmanSNde NoronhaF3′-Azido-3′-deoxythymidine in feline leukemia virus-infected cats: a model for therapy and prophylaxis of AIDSCancer Res19874712319031943034403
- HaschekWMWeigelRMScherbaGZidovudine toxicity to cats infected with feline leukemia virusFundam Appl Toxicol19901447647752163339
- GingerichDALymphocytic T-cell immunomodulator (LTCI): Review of the immunopharmacology of a new veterinary biologicIntern J Appl Res Vet Med2008626168
- MayrBHörberDParamunisierung FeLV-positiver Katzen – ein Bericht aus der PraxisKleintierprax199237515518
- HörberDSchnablWMayrBPraxiserfahrungen bei der Paramunisierung FeLV-positiver Katzen mit Baypamun HK. TierärztlUmschau199247556560
- HartmannKBlockAFerkGBeerBVollmarALutzHTreatment of feline leukemia virus (FeLV) infectionVet Microbiol1999691–211111310515279
- DayNKEngelmanRWLiuWTTrangLGoodRARemission of lymphoma leukemia in cats following ex vivo immunosorption therapy using Staphylococcus protein AJ Biol Response Mod1984332782856086846
- OperarioDJReynoldsHMKimBComparison of DNA polymerase activities between recombinant feline immunodeficiency and leukemia virus reverse transcriptasesVirology2005335110612115823610