Abstract
Endoglin (CD105) is a type III auxiliary receptor for the transforming growth factor beta (TGFβ) superfamily. Several lines of evidence suggest that endoglin plays a critical role in maintaining cardiovascular homeostasis. Seemingly disparate disease conditions, including hereditary hemorrhagic telangiectasia, pre-eclampsia, and cardiac fibrosis, have now been associated with endoglin. Given the central role of the TGFβ superfamily in multiple disease conditions, this review provides a detailed update on endoglin as an evolving therapeutic target in the management of cardiovascular disease.
Introduction
Endoglin (CD105) is a type III auxiliary receptor for the transforming growth factor beta (TGFβ) superfamily. Over the past three decades, several lines of evidence suggest that endoglin plays a critical role in maintaining cardiovascular homeostasis. First, mutations in human endoglin result in the autosomal dominant vascular dysplastic syndrome, hereditary hemorrhagic telangiectasia (HTT).Citation1 Second, Endoglin null mice (Eng−/−) die at embryologic day 10–11.5 because of impaired cardiovascular development.Citation2 Third, endoglin expression is increased in human atherosclerotic plaquesCitation3 and in models of balloon angioplasty-mediated vascular injury.Citation4 Fourth, endoglin is abundantly expressed by neovascularized tumors in prostate, breast, and colon cancer.Citation5 Finally, endoglin has recently been shown to regulate cardiac remodeling in heart failureCitation6 and myocardial infarction.Citation7 Given the central role of the TGFβ superfamily in multiple disease conditions, endoglin represents a potentially unique target of therapy for several debilitating conditions involving the cardiovascular system.
Endoglin structure, function, and distribution
Endoglin is a 180 kDa homodimeric integral membrane glycoprotein composed of disulfide-linked subunits. The receptor includes a large 561-amino acid extracellular domain with a single transmembrane domain and a serine/threonine-rich cytoplasmic region, with 47 amino acids in the long isoform (L-endoglin) and 14 residues in the short isoform (S-endoglin).Citation8 The cytoplasmic domain of the predominantly expressed L-endoglin constitutes the region of the protein with the highest degree of conservation in mammalian species. A low-resolution structural analysis of the extracellular domain revealed a dome comprised of antiparallel-oriented monomers with a cavity at one end. Each endoglin monomer is comprised of three well defined domains. A large N-terminus domain extending from Glu26 to Ile359 forms a large flat area and does not show any degree of identity with known protein domains and is therefore referred to as the orphan domain. This domain mediates interactions with ligands of the TGFβ superfamily. Initial studies demonstrated that endoglin can be found in the receptor complex for the TGFβ1 and TGFβ3 isoforms in association with the type II receptor (TBR2).Citation9 Subsequently, it was shown that endoglin also interacts with activin as well as bone morphogenetic proteins (BMP)-2 and BMP7 in association with the ligand binding receptor, which can be either a type I or type II receptor depending on the ligand involved.Citation10 More recent studies have shown that endoglin may bind directly to BMP9 and BMP10.Citation11,Citation12
The orphan domain is followed by two zona pellucida subdomains, organized into an open U-shaped monomer and containing eight conserved cysteine residues localized to Lys362–Asp561.Citation12–Citation14 This zona pellucida domain is highly conserved among endoglin proteins from human, mouse, rat, pig, and dog models, and constitutes more than 30% of the extracellular region, suggesting an important functional role.Citation11,Citation15 The extracellular domain of endoglin also contains consensus motifs for several O-linked and N-linked oligosac-charide chains, with glycosylation accounting for about 30% of the molecular weight. The sequence of human endoglin originally revealed an arginine-glycine-aspartic acid (RGD) tripeptide at Arg399–Asp401, suggesting that endoglin may play a role in cell adhesion.Citation14 However, that sequence was not conserved in other species. A recent study demonstrated that endoglin can bind to the α5β1 integrin via its RGD motif or a functionally equivalent murine TDD peptide that may facilitate leukocyte transmigration.Citation16
The extracellular domain of endoglin can be proteolytically cleaved by matrix metalloproteinase 14 and circulate as soluble endoglin, which may serve as a naturally occurring antagonist for TGFβ signaling and therefore play an important role in cardiovascular disease.Citation6,Citation8,Citation17 The cytoplasmic domain of endoglin is phosphorylated by serine-threonine kinase TBRI and TBR2 TGFβ receptors. Endoglin phosphorylation is thought to influence its subcellular localization by modulating its interaction with adhesive proteins such as zyxin and zyxin-related protein-1.Citation18,Citation19
Endoglin is present at high levels on the vascular endothelium in adults and is expressed early during development on vascular endothelium and on mesenchymal tissue derived from the endocardium.Citation20 Stromal cells of mesenchymal origin,Citation21 smooth muscle cells,Citation22 the placental syncytiotrophoblast, mesenchymal and hematopoietic stem cells, pre-erythroblasts, leukemic cells of lymphoid and myeloid lineage, and activated blood monocytes also express endoglin. In cardiac tissue, endoglin is expressed by the endocardium and fibroblasts, with minimal expression by cardiomyocytes.Citation8,Citation21
Endoglin maintains vascular homeostasis
In quiescent nonproliferative endothelium, TGFβ signal transduction is predominantly mediated by the type I TGFβ receptor, activin receptor-like kinase-5,Citation23 and phosphorylation of signaling mediators known as Smads-2/3. Alternatively, TGFβ-mediated activation of the activin receptor-like kinase-1 pathway phosphorylates Smads-1/5/8, which promotes endothelial proliferation and migrationCitation24 (). Endoglin negatively regulates TGFβ-mediated activin receptor-like kinase-5 signaling in quiescent endothelium, while promoting activin receptor-like kinase-1 signaling in activated endothelium.Citation24,Citation25 Conversely, in the absence of endoglin, activin receptor-like kinase-1 signaling is attenuated, while activin receptor-like kinase-5-mediated growth arrest of endothelial cells is stimulatedCitation24 (). Li et al determined that suppressed endoglin expression enhances the inhibitor effects and further that endoglin overexpression promotes endothelial proliferation.Citation26,Citation27 These pathways have important implications for the development of arteriovenous malformations associated with HHT, as described below.
Figure 1 Endoglin signaling in endothelium.
Abbreviation: ALK, activin receptor-like kinase; P, phospho.
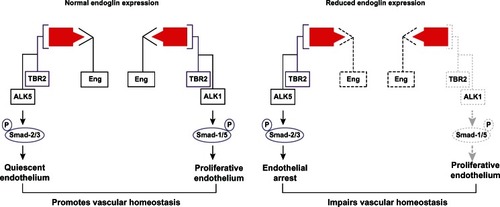
Further substantiating the role of endoglin in maintaining vascular homeostasis, cells from Endoglin heterozygous (Eng+/−) mice exhibit lower levels of endothelial nitric oxide synthase (eNOS), which results in less nitric oxide generation and more eNOS-derived superoxide production due to an uncoupling of eNOS activity.Citation28 Endoglin may modulate vascular homeostasis via its colocalization to endothelial membrane-associated caveolae.Citation26 Consistent with this observation, resistance arteries from Eng+/− mice demonstrate impaired myogenic responses and enhanced eNOS-dependent vasodilatation despite reduced eNOS levels. These findings may reflect increased stabilization of eNOS protein expression and enhanced calcium-induced activation of eNOS by endoglin.Citation28,Citation29 Subsequent data have also shown that TGFβ1 directly induces eNOS-dependent vasorelaxation, which is blocked by high circulating levels of soluble endoglinCitation18 and may contribute to elevated vascular resistance.Citation17 In addition to regulating eNOS activity, endoglin expression may correlate with myeloperoxidase activity and expression of vascular cell adhesion molecule 1, inducible nitric oxide synthase, and CD68 during ischemia-reperfusion injury.Citation30
Endoglin modulates cardiac development
The importance of endoglin in hematopoiesis, angiogenesis, cardiovascular development, and vascular remodeling has been well establishedCitation31 and is supported by the observation that Endoglin null mice (Eng−/−) die at embryonic day 10–11.5 as a result of cardiovascular abnormalities.Citation2,Citation20,Citation32,Citation33 The critical role of endoglin in cardiovascular development is further supported by high levels of endoglin expression in the endocardial cushion during valve formation and heart septation.Citation34 Originating from the lateral plate mesoderm, both cardiac myocytes and cardiac endothelial progenitor cells ultimately constitute the primitive spongy heart tube. A subpopulation of endothelial cells in the region of the future cardiac valve and septal formation transforms into mesenchymal cells, which migrate into the cardiac jelly and begin to express α-actin.Citation35 A critical aspect of this process is the ability of the developing myocardium to induce endothelial transformation via endocardial-myocardial signaling pathways, which is mediated in part by signaling through TGFβ1, TGFβ3, and endoglin.Citation35–Citation37 The role of endoglin in valve formation was recently highlighted by a systems biology approach which identified a haplotype within the endoglin (ENG) gene that is strongly associated with the presence of a bicuspid aortic valve in patients.Citation38
Regulators of endoglin expression
Endoglin is localized to human chromosome 9 and contains 15 exons of which 13 code for the extracellular domain. The 5′ flanking region of the gene lacks consensus TATA and CAAT boxes, but contains two regions rich in G-C and consensus motifs for SP1, ETS, GATA, AP-2, NFkB, MAD, and response elements for TGFβ, glucocorticoids, vitamin D, and estrogen.Citation39 Importantly, both endoglin and type I collagen promoters contain Smad-binding elements in close proximity to Sp1 binding sites, suggesting a possible mechanism by which TGFβ1 may coactivate endoglin and type I collagen expression.Citation40 Factors known to induce endoglin expression in various cell types include angiotensin II,Citation41 TGFβ,Citation6,Citation42 hypoxia-inducible factor-1,Citation43 hypercholesterolemia,Citation44 and balloon-mediated vascular injury via the Kruppel-like zinc finger transcription factor, KLF6.Citation4 The inflammatory cytokine, tumor necrosis factor-alpha (TNFα), attenuates endoglin expression by endothelial cells in vitro.Citation26
Several pharmacologic agents also influence endoglin expression. Tranexamic acid is a competitive inhibitor of plasminogen activation to plasmin, and at much higher concentrations, a noncompetitive inhibitor of plasmin. In endothelial cells, tranexamic acid promotes endoglin gene and protein expression and TGFβ1 signaling via activin receptor-like kinase-1.Citation45 Conversely, inhibitors of angiotensin-converting enzyme, such as trandolapril,Citation46 and angiotensin type I receptor inhibitors, such as valsartan, losartan, and candesartan, attenuate angiotensin II-induced endoglin expression.Citation42 The 3-hydroxy-3-methyl-glutaryl-CoA reductase inhibitor, atorvastatin, also decreases endoglin expression in mouse models of hyperlipidemia, which appears to correlate with the hypolipidemic effects of statin therapy.Citation44,Citation47
Acting as a biosensor for humoral and mechanical stimuli, the vascular endothelium plays a primary role in maintaining local and systemic homeostasis under conditions of mechanical stress. As a central marker of endothelial cell integrity and function, the effect of hemodynamic forces and mechanical deformation on endoglin activity remains largely uncharacterized. Several lines of evidence suggest that endoglin expression is regulated by mechanical forces such as stretch and strain. First, the evolutionarily conserved zona pellucida domain in the extracellular region of endoglin is mechanically responsive in fruit flies.Citation15 Second, in vivo models of placental hypertension and balloon-induced arterial injury are characterized by increased endoglin expression in vascular tissue.Citation4,Citation17 Third, endoglin expression is increased in hemorrhoidal tissue and correlates with an increased propensity for thrombosis, suggesting that increases in venular wall tension may modulate endoglin expression and vascular thromboresistance.Citation48 Recent data showed that mechanically stretching myoblasts increases endoglin expression; a process which appears to be regulated by microRNA-208A.Citation49
As a central regulator of cardiovascular homeostasis, a functional role for endoglin has been identified in several important clinical disease conditions, including HHT, pre-eclampsia, cardiac remodeling, cancer, pulmonary hypertension, and other fibroproliferative disorders. Several of these conditions are discussed in detail below.
Hereditary hemorrhagic telangiectasia
HHT, known historically as Osler-Weber-Rendu syndrome, is a disorder of vascular homeostasis affecting small and large vessels. The prevalence of HHT varies depending on the population studied, from as low as about 1:39,000 in northern England to as high as about 1:1330 in the Afro-Caribbean population of Curaçao and Bonaire, with an average prevalence estimated at 1:5000–1:8000.Citation50–Citation53 The hallmark of HHT is the formation of arteriovenous malformations, which may occur in small vessels, leading to cutaneous telangiectases, nosebleeds, and gastrointestinal bleeds, or in large vessels of the lungs, brain, and liver. Important clinical sequelae of large-vessel arteriovenous malformations include hemorrhage, stroke, and brain abscess for pulmonary arteriovenous malformations, and pulmonary hypertension and high output heart failure arteriovenous malformations in the liver. Rupture of arteriovenous malformations, either spontaneous or associated with pregnancy or other physiological stressors, represents a significant source of morbidity in patients with HHT.Citation51,Citation54–Citation57 The diagnosis of HHT is based on clinical observations that are summarized in consensus criteria known as the “Curaçao criteria”.Citation56,Citation58
Mutations in endoglin (ENG, OMIM#187300) and activin receptor-like kinase-1 (also known as ACVRL1, OMIM#600376) genes lead to HHT1 and HHT2, respectively, and account for >80% of cases.Citation59 Mutations in Smad-4 (MADH4, OMIM #175050) are also associated with 2%–3% of cases with a combined juvenile polyposis-HHT syndrome.Citation59 Two other loci have been shown to be in linkage disequilibrium with HHT; one on chromosome 5, defining HHT3, and the other on chromosome 7, defining HHT4.Citation60,Citation61 The precise genes involved remain to be identified and would give clues about the pathways deficient in HHT.
Transgenic mouse models of deficient expression of endoglin or ALK-1 recapitulate clinical features of HHT.Citation62,Citation63 In Eng−/− mice, complete loss of endoglin is embryonically lethal due to impaired angiogenesis, while heterozygosity leads to the clinical syndrome of HHT1 with variable penetrance.Citation2,Citation62 In humans with HHT1, endoglin expression in peripheral blood monocytes and newborn umbilical vein endothelial cells is half of control levels and mutant proteins are rarely detected.Citation59 These and other data support the role of haploinsufficiency in the pathogenesis of HHT1 as opposed to reduced local expression of endoglin through a “second hit” process or interference by the mutated allele with the function of the normal allele.Citation59,Citation62
Mutations in ALK-1 and to a lesser degree ENG are associated with pulmonary arterial hypertension, which can occur in the absence of HHT.Citation64–Citation66 Both endoglin and ALK-1 can associate with the BMP type II receptor, which is primarily responsible for most familial forms of pulmonary arterial hypertension.Citation23 The mechanistic link between pulmonary arterial hypertension and HHT was recently studied in adult ALK-1+/− and Eng+/− mice, which develop signs of pulmonary arterial hypertension that can be attenuated by treatment with the antioxidant, tempol.Citation67,Citation68 These findings highlight the importance of eNOS-derived superoxide in these mice due to uncoupling of eNOS, which contributes to impaired vascular tone in either HHT or pulmonary arterial hypertension. Furthermore, endoglin and ALK-1 are coexpressed in the terminal-most segments of the pulmonary vasculature, which is most affected in mouse models and where most HHT-associated arteriovenous malformations tend to form.Citation69 However, physiologically, pulmonary arterial hypertension is associated with increased pulmonary vascular resistance, while arteriovenous malformations are associated with abnormal vasodilatation.Citation70
An important treatment option for patients with HHT is embolization of visceral arteriovenous malformations, which can prevent major adverse clinical events, including stroke, high-output heart failure, pulmonary hypertension, and hemorrhage. Hormonal and antiangiogenic agents have also been explored as potential therapy for HHT. Clinical trials of estrogen preparations including the estrogen receptor antagonist, tamoxifen, and the selective estrogen receptor modulator, raloxifene, can reduce episodes of epistaxis and transfusion requirements.Citation71,Citation73,Citation74 Inhibition of angiogenesis through the use of agents such as thalidomide, lenalidomide, and bevacizumab reduces the incidence of nasal and gastrointestinal bleeding in some patients.Citation75–Citation80 However, the lack of randomized controlled trial data for bevacizumab and the absence of long-term safety or efficacy data for antiangiogenesis therapy limits enthusiasm for these HHT treatment approaches, which remain experimental and limited to highly selected patients. The therapeutic action of thalidomide in HHT derives in part from increased platelet-derived growth factor expression at low doses and a direct antiangiogenic effect at high doses.Citation77 Raloxifene and tranexamic acid increase in vitro mRNA and protein expression of endoglin and ALK-1 in endothelial cells.Citation45,Citation71 Whether future therapies such as gene transfer methods can target the underlying genetic defect associated with HHT remains unknown.Citation81,Citation82 Further insight into the role of endoglin and the TGFβ signaling pathway in the pathogenesis of HHT, together with evolving molecular approaches, may lead to novel therapeutic approaches for this debilitating condition.
Pre-eclampsia
Pre-eclampsia is a systemic disorder occurring in 3% of pregnant women after 20 weeks of gestation with symptoms of new-onset hypertension and proteinuria defined as a systolic blood pressure > 140 mmHg or diastolic blood pressure > 90 mmHg in a previously normotensive patient and at least 0.3 g of protein in a 24-hour urine specimen. Eclampsia is defined as pre-eclampsia with grand mal seizures without an identifiable cause.Citation83,Citation84 The incidence of maternal death associated with pre-eclampsia has been estimated at one in 100,000 live births.Citation85 Maternal complications include central nervous system involvement, acute renal or liver failure, and hematological dysfunction.Citation86 Incident pre-eclampsia is associated with increased risk of subsequent cardiovascular morbidity, including hypertension and ischemic heart disease, although it is not clear if pre-eclampsia is causal or reflects an underlying predisposition to cardiovascular disease.Citation87 Fetal effects of pre-eclampsia include prematurity, fetal growth restriction, oligohydramnios, and placental abruption.Citation88,Citation89 Notable risk factors for the development of pre-eclampsia include a prior personal or family history of pre-eclampsia, advanced maternal age, obesity, and pregestational diabetes.Citation90
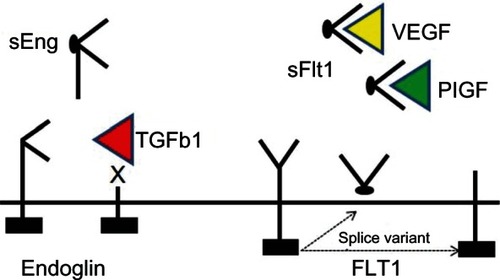
The pathophysiology of pre-eclampsia remains poorly understood. Early changes in the placenta prior to the appearance of clinical manifestations of pre-eclampsia include failure of migrating placental cells to express endothelial surface adhesion markers, resulting in incomplete invasion of maternal arteries by the developing trophoblast.Citation91–Citation94 Pathological evidence of placental ischemia has been observed in some cases of pre-eclampsia,Citation95 which is thought to trigger release of antiangiogenic factors, including soluble endoglin and soluble fms-like tyrosine kinase (sFlt1) into the maternal circulation.Citation96,Citation97 sFlt1 antagonizes the angiogenic factors, vascular endothelial growth (VEGF) factor and placental growth factor, while soluble endoglin antagonizes TGFβ1 and TGFβ3 activity.Citation9,Citation17,Citation19,Citation97 Furthermore, systemic disruption of vascular hemostasis caused by elaboration of these antiangiogenic factors promotes pre-eclampsia.Citation86,Citation98
The VEGF and TGFβ axes are the major signaling pathways implicated in the pathogenesis of pre-eclampsia (). sFlt1, or the type 1 VEGF receptor, can be generated by either alternative splicing of the Flt1 gene or by cleavage of the Flt1 extracellular domain in response to hypoxia or thrombin.Citation99,Citation100 Elevated levels of sFlt1 in the maternal circulation correspond to a fall in VEGF and placental growth factor levels.Citation97,Citation101–Citation103 A putative role for sFlt1 in pre-eclampsia is supported by studies showing that administration of exogenous sFlt1 to pregnant rats induces severe pre-eclampsia, while removal of sFlt1 by immunoprecipitation normalizes the angiogenic response of cells derived from placental villous explants.Citation17,Citation96 Similarly, pre-eclamptic placentae express increased levels of membrane-bound and soluble endoglin.Citation17 A recent study demonstrated that oxysterol activation promotes matrix metalloproteinase 14-mediated cleavage of soluble endoglin in cells of trophoblast origin,Citation104 which suggests that hypoxia and oxidative stress are important triggers for release of soluble endoglin. Once in the circulation, soluble endoglin antagonizes TGFβ1-induced vasodilation, thereby promoting hypertension.Citation28,Citation105,Citation106 A rise in circulating maternal soluble endoglin heralds the onset of symptoms in patients with pre-eclampsia.Citation102,Citation107,Citation108 Individually and in concert with sFlt1, soluble endoglin induces a pre-eclamptic state in pregnant rats.Citation17 Investigation into the regulation of sFlt and soluble endoglin in pre-eclampsia is ongoing.Citation109
Figure 2 Circulating angiogenic peptides in pre-eclampsia.
Abbreviations: TGFβ1, transforming growth factor beta-1; PIGF, placental growth factor; VEGF, vascular endothelial growth factor; sEng, soluble endoglin; mRNA, messenger RNA.
At present, the primary mode of therapy for pre-eclampsia is accelerated delivery of the placenta. The available evidence strongly implicates an imbalance of the antiangiogenic factors, sFlt1 and soluble endoglin, and the proangiogenic factors, placental growth factor, TGFβ, and VEGF, in the pathogenesis of pre-eclampsia. Strategies targeting maternal depletion of soluble endoglin or sFlt1 would plausibly de-escalate the disease process. Given the growing appreciation for the role of sFlt1 and soluble endoglin in the pathophysiology of pre-eclampsia, novel therapeutic approaches include the use of antibodies and small molecules to sequester or limit synthesis of antiangiogenic molecules. Alternatively, modulating the balance of angiogenic factor levels has been recently explored in a study that showed improvement in blood pressure and renal function after administration of exogenous VEGF in a preclinical model of pre-eclampsia.Citation110–Citation112 More recently, directly removing sFlt1 from the maternal circulation by extracorporeal apheresis using a dextran sulfate column has been introduced as a potential therapeutic approach for pre-eclampsia.Citation113 Other approaches include induction of hemoxygenase-1 with cobalt protoporphyrin in a rat model of pre-eclampsiaCitation114 and direct inhibition of matrix metalloproteinase 14 to prevent the release of soluble endoglin.Citation115,Citation116 Limiting fetal consequences of any treatment approach is mandatory and remains a challenging limitation to the development of new treatments.
Cardiac remodeling
Heart failure is a major cause of global morbidity and mortality that affects nearly 23 million individuals worldwide.Citation117,Citation118 Heart failure commonly occurs secondary to hypertensive heart disease, myocardial infarction, infection, and inherited cardiomyocyte dysfunction. A decline in cardiac function activates several signaling cascades that promote cardiomyocyte hypertrophy and cardiac fibrosis, a process known as cardiac remodeling.Citation119 At each phase of cardiac remodeling from acute load to compensatory hypertrophy, various signaling cascades are implicated. Among these, TGFβ1 promotes the cardiac hypertrophy and fibrosis associated with heart failure. In response to angiotensin II, TGFβ1 is increased, converts fibroblasts into myofibroblasts, and generates extracellular matrix proteins, such as type I collagen.Citation120 Excess collagen deposition exaggerates mechanical stiffness of the left ventricle, impairs myocyte contractility, disrupts electrical coupling, and worsens tissue hypoxia,Citation119 thereby promoting heart failure.
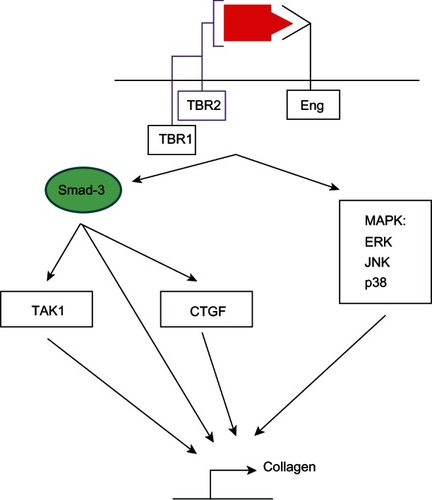
TGFβ1-induced fibrosis involves both canonical and noncanonical signaling (). Among the canonical pathways, Smad-3 plays a central role.Citation120 First, Smad-3 mediates TGFβ1-induced activation of a TGFβ-response elementCitation57 located in the promoter region of type I collagen.Citation121 Second, Smads-2/3 and Smad-4 form a complex with activated transcription factor-2 to stimulate collagen synthesis. Activated transcription factor-2 phosphorylation requires activation of TGFβ-activated kinase.Citation122 Third, Smad-3 mediates TGFβ1-induced expression of connective tissue growth factor via a TGFβ1 response element located in the connective tissue growth factor promoter.Citation123 Connective tissue growth factor is a critical downstream mediator of tissue fibrosis. Noncanonical pathways contributing to TGFβ1-induced collagen synthesis involve activation of mitogen-activated protein kinases including: extracellular regulated kinases, jun-n-terminal kinases, and p38 kinase.Citation120 While all three cascades are activated in advanced heart failure, the extracellular regulated kinase pathway is required for TGFβ1-induced connective tissue growth factor expression and collagen synthesis.Citation124 In cardiac fibroblasts, connective tissue growth factor is an important downstream mediator of TGFβ1-induced fibrosis because it promotes binding of TGFβ1 to TBR2 and blocks inhibition of the pathway by Smad-7.Citation124,Citation125 Given its central role in stimulating fibrosis, TGFβ1 has been nonselectively targeted in heart failure models using multiple approaches, none of which have had clearly beneficial therapeutic effects.Citation126,Citation127
Figure 3 Canonical and noncanonical transforming growth factor beta-1/endoglin signaling in fibrosis.
Abbreviations: CTGF, connective tissue growth factor; ERK, extracellular regulated kinase; JNK, jun-n-terminal kinase; Eng, endoglin; MAPK, mitogen-activated protein kinase.
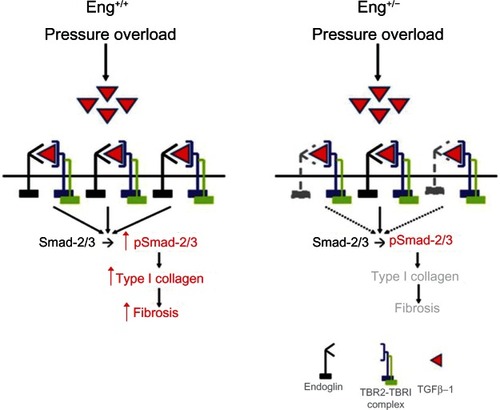
Recently, endoglin expression was found to be increased in left ventricular tissue from patients with end-stage heart failure and in a murine model of thoracic aortic constriction-induced heart failure.Citation6 Further analysis showed that endoglin is highly expressed by cardiac fibroblasts and endothelium, but poorly on cardiomyocytes. Compared with Eng+/+ wild-type mice, Eng+/− mice demonstrated improved survival, limited cardiac fibrosis, and enhanced myocardial capillarity after thoracic aortic constriction (). Loss-of-function studies in vitro confirmed the dependence of TGFβ1 activity on endoglin expression in human cardiac fibroblasts. Paradoxically, adenoviral-mediated overexpression of full-length endoglin also blocked TGFβ1-induced collagen synthesis. Further study showed that levels of soluble endoglin were elevated in the conditioned medium after treatment with the adenovirus, thereby implicating soluble endoglin as a negative regulator of TGFβ1 activity. This observation was confirmed by adenoviral-mediated overexpression of human soluble endoglin or treatment with recombinant human soluble endoglin in vitro. Treatment with adenoviral-mediated overexpression of human soluble endoglin attenuated cardiac fibrosis in wild-type mice after thoracic aortic constriction. These findings suggest that endoglin is required for TGFβ1 signaling in cardiac fibroblasts and that selectively inhibiting TGFβ1 signaling by reducing endoglin activity attenuates cardiac fibrosis and improves survival in a mouse model of heart failure. In contrast with the functional role of endoglin in promoting TGFβ1 signaling, soluble endoglin limits TGFβ1 signaling, type I collagen synthesis, and ultimately cardiac fibrosis.
Figure 4 Functional role of endoglin in cardiac fibrosis.
Abbreviation: TGFβ1, transforming growth factor beta-1.
In a separate study, circulating levels of soluble endoglin were significantly increased in patients with suspected left ventricular dysfunction and were strongly correlated with predictors of mortality in heart failure, such as elevated left ventricular end-diastolic pressure, reduced left ventricular ejection fraction, and worsening New York Heart Association class symptoms.Citation128 Soluble endoglin levels demonstrated superior sensitivity, specificity, accuracy, and predictive value compared with atrial and brain natriuretic peptides for identifying subjects with increased left ventricular end-diastolic pressure. Furthermore, levels were reduced in patients receiving medical therapy for congestive heart failure and correlated with reduced cardiac filling pressures. These findings identify elevated soluble endoglin levels as a biomarker of increased cardiac filling pressure and perhaps a potential biomarker of endoglin activity in heart failure.Citation129 These findings also represent an important paradox in endoglin biology. Expression of membrane-associated endoglin is increased in heart failure and correlates with a direct increase in circulating soluble endoglin. However, membrane-associated endoglin promotes adverse cardiac remodeling by driving cardiac fibrosis, while soluble endoglin may serve as a negative feedback loop to limit TGFβ1/endoglin signaling. The exact functional role of soluble endoglin remains to be determined, but appears to serve as a potentially important diagnostic marker of heart failure as well as a participant in the cardiac remodeling associated with heart failure.
The role of soluble endoglin in heart failure remains poorly understood. First, the mechanism underlying increased expression of soluble endoglin in heart failure is unknown. Proteolytic cleavage of soluble endoglin from membrane endoglin may occur both locally in cardiac tissue or systemically because levels of matrix metalloproteinase 14 are known to be elevated in heart failure.Citation130 Second, the mechanism by which higher soluble endoglin levels interrupt TGFβ1 signaling remains poorly characterized. First, endoglin may modulate signaling via several TGFβ-family ligands and soluble endoglin could serve as a ligand trap for ligands of the TGFβ superfamily.Citation10 However, recent studies indicate that BMP9 and BMP10 may be the only ligands that bind to soluble endoglin with high affinity.Citation12 Second, soluble endoglin may promote alternate signaling pathways that indirectly inhibit TGFβ1 signaling, such as BMP7.Citation131 Finally, release of soluble endoglin by ectodomain shedding may render the receptor nonfunctional by removing the primary active component of endoglin, the extracellular domain, and leaving behind a nonfunctional cytoplasmic tail that cannot interact with TBR2.
Further supporting the role of endoglin in cardiac remodeling, studies of acute myocardial infarction have shown that Eng+/− mice exhibit showed neoangiogenesis in the infarct zone, progressive left ventricular dilatation, worsening cardiac function, and elevated cardiac filling pressures compared with wild-type controls. In this model, adverse cardiac remodeling in Eng+/− mice was partially rescued by intravenous injection of mononuclear cells from healthy human donors with normal endoglin expression but not by those from patients with HHT1 harboring mutant endoglin.Citation7
In contrast with heart failure, circulating levels of soluble endoglin in patients presenting with acute myocardial infarction were significantly lower than in healthy controls, and decreased further in the first 48 hours of admission.Citation132 Reduced levels of soluble endoglin were an independent predictor of short-term and long-term cardiovascular mortality. Unlike other known circulating angiogenic proteins, including sFlt1 and VEGF, levels of soluble endoglin are not altered by treatment with the anticoagulants commonly used to manage patients with myocardial infarction.Citation133 Whether alterations in soluble endoglin levels correlate with atherosclerotic burden remains under investigation.Citation134 Collectively, these data suggest that both membrane-associated and soluble endoglin serve important roles in cardiac remodeling due to heart failure or acute myocardial infarction, and may serve as potential targets of therapy for these debilitating conditions.
In summary, the role of endoglin as a critical mediator of cardiovascular homeostasis has been well established over the past two decades. Studies have now begun to explore the potential utility of targeting endoglin and the signaling program it regulates as a novel therapeutic approach for patients suffering from cardiovascular diseases, including HHT1, pre-eclampsia, pulmonary hypertension, heart failure, myocardial infarction, atrial fibrillation, pulmonary hypertension, and atherosclerosis.
Disclosure
The authors report no conflicts of interest in this work.
References
- McAllisterKAGroggKMJohnsonDWEndoglin, a TGF-beta binding protein of endothelial cells, is the gene for hereditary haemor-rhagic telangiectasia type 1Nat Genet1994843453517894484
- LiDYSorensenLKBrookeBSDefective angiogenesis in mice lacking endoglinScience199928454191534153710348742
- ConleyBASmithJDGuerrero-EsteoMBernabeuCVaryCPEndoglin, a TGF-beta receptor-associated protein, is expressed by smooth muscle cells in human atherosclerotic plaquesAtherosclerosis2000153232333511164421
- BotellaLMSanchez-ElsnerTSanz-RodriguezFTranscriptional activation of endoglin and transforming growth factor-beta signaling components by cooperative interaction between Sp1 and KLF6: their potential role in the response to vascular injuryBlood2002100124001401012433697
- LiCGuoBBernabeuCKumarSAngiogenesis in breast cancer: the role of transforming growth factor beta and CD105Microsc Res Tech200152443744911170303
- KapurNKWilsonSYunisAAReduced endoglin activity limits cardiac fibrosis and improves survival in heart failureCirculation2012125222728273822592898
- van LaakeLWvan den DriescheSPostSEndoglin has a crucial role in blood cell-mediated vascular repairCirculation2006114212288229717088457
- BernabeuCConleyBAVaryCPNovel biochemical pathways of endoglin in vascular cell physiologyJ Cell Biochem200710261375138817975795
- CheifetzSBellonTCalesCEndoglin is a component of the transforming growth factor-beta receptor system in human endothelial cellsJ Biol Chem19922672719027190301326540
- BarbaraNPWranaJLLetarteMEndoglin is an accessory protein that interacts with the signaling receptor complex of multiple members of the transforming growth factor-beta superfamilyJ Biol Chem199927425845949872992
- AltAMiguel-RomeroLDonderisJStructural and functional insights into endoglin ligand recognition and bindingPLoS One201272e2994822347366
- CastonguayRWernerEDMatthewsRGSoluble endoglin specifically binds bone morphogenetic proteins 9 and 10 via its orphan domain, inhibits blood vessel formation, and suppresses tumor growthJ Biol Chem201128634300343004621737454
- GougosALetarteMIdentification of a human endothelial cell antigen with monoclonal antibody 44G4 produced against a pre-B leukemic cell lineJ Immunol19881416192519333262644
- GougosALetarteMPrimary structure of endoglin, an RGD-containing glycoprotein of human endothelial cellsJ Biol Chem199026515836183641692830
- JovineLQiHWilliamsZLitscherEWassarmanPMThe ZP domain is a conserved module for polymerization of extracellular proteinsNat Cell Biol20024645746112021773
- RossiESanz-RodriguezFElenoNEndothelial endoglin is involved in inflammation: role in leukocyte adhesion and transmigrationBlood2013121240341523074273
- VenkateshaSToporsianMLamCSoluble endoglin contributes to the pathogenesis of preeclampsiaNat Med200612664264916751767
- ConleyBAKolevaRSmithJDEndoglin controls cell migration and composition of focal adhesions: function of the cytosolic domainJ Biol Chem200427926274402744915084601
- Sanz-RodriguezFGuerrero-EsteoMBotellaLMBanvilleDVaryCPBernabeuCEndoglin regulates cytoskeletal organization through binding to ZRP-1, a member of the Lim family of proteinsJ Biol Chem200427931328583286815148318
- BourdeauADumontDJLetarteMA murine model of hereditary hemorrhagic telangiectasiaJ Clin Invest1999104101343135110562296
- GougosASt JacquesSGreavesAIdentification of distinct epitopes of endoglin, an RGD-containing glycoprotein of endothelial cells, leukemic cells, and syncytiotrophoblastsInt Immunol19924183921371694
- AdamPJCleshamGJWeissbergPLExpression of endoglin mRNA and protein in human vascular smooth muscle cellsBiochem Biophys Res Commun1998247133379636649
- MachadoRDAldredMAJamesVMutations of the TGF-beta type II receptor BMPR2 in pulmonary arterial hypertensionHum Mutat200627212113216429395
- LebrinFGoumansMJJonkerLEndoglin promotes endothelial cell proliferation and TGF-beta/ALK1 signal transductionEMBO J200423204018402815385967
- LebrinFDeckersMBertolinoPTen DijkePTGF-beta receptor function in the endotheliumCardiovasc Res200565359960815664386
- LiCHampsonINHampsonLKumarPBernabeuCKumarSCD105 antagonizes the inhibitory signaling of transforming growth factor beta1 on human vascular endothelial cellsFASEB J2000141556410627280
- Pece-BarbaraNVeraSKathirkamathambyKEndoglin null endothelial cells proliferate faster and are more responsive to transforming growth factor beta1 with higher affinity receptors and an activated Alk1 pathwayJ Biol Chem200528030278002780815923183
- ToporsianMGrosRKabirMGA role for endoglin in coupling eNOS activity and regulating vascular tone revealed in hereditary hemorrhagic telangiectasiaCirc Res200596668469215718503
- JerkicMRivas-ElenaJVSantibanezJFEndoglin regulates cyclooxygenase-2 expression and activityCirc Res200699324825616840721
- DochertyNGLopez-NovoaJMArevaloMEndoglin regulates renal ischaemia-reperfusion injuryNephrol Dial Transplant20062182106211916751653
- ChenCZLiMde GraafDIdentification of endoglin as a functional marker that defines long-term repopulating hematopoietic stem cellsProc Natl Acad Sci USA20029924154681547312438646
- ChoSKBourdeauALetarteMZuniga-PfluckerJCExpression and function of CD105 during the onset of hematopoiesis from Flk1(+) precursorsBlood200198133635364211739167
- BorgesLIacovinoMMayerhoferTA critical role for endoglin in the emergence of blood during embryonic developmentBlood2012119235417542822535663
- QuRSilverMMLetarteMDistribution of endoglin in early human development reveals high levels on endocardial cushion tissue mesen-chyme during valve formationCell Tissue Res199829223333439560476
- MolinDGBartramUVan der HeidenKExpression patterns of Tgfbeta1-3 associate with myocardialisation of the outflow tract and the development of the epicardium and the fibrous heart skeletonDev Dyn2003227343144412815630
- Nomura-KitabayashiAAndersonGASleepGEndoglin is dispensable for angiogenesis, but required for endocardial cushion formation in the midgestation mouse embryoDev Biol20093351667719703439
- SridurongritSLarssonJSchwartzRRuiz-LozanoPKaartinenVSignaling via the Tgf-beta type I receptor Alk5 in heart developmentDev Biol2008322120821818718461
- WootenECIyerLKMontefuscoMCApplication of gene network analysis techniques identifies AXIN1/PDIA2 and endoglin haplotypes associated with bicuspid aortic valvePLoS One201051e883020098615
- RiusCSmithJDAlmendroNCloning of the promoter region of human endoglin, the target gene for hereditary hemorrhagic telangiectasia type 1Blood19989212467746909845534
- BotellaLMSanchez-ElsnerTRiusCCorbiABernabeuCIdentification of a critical Sp1 site within the endoglin promoter and its involvement in the transforming growth factor-beta stimulationJ Biol Chem200127637344863449411432852
- LiDChenHMehtaJLAngiotensin II via activation of type 1 receptor upregulates expression of endoglin in human coronary artery endothelial cellsHypertension20013851062106711711498
- ChenKMehtaJLLiDJosephLJosephJTransforming growth factor beta receptor endoglin is expressed in cardiac fibroblasts and modulates profibrogenic actions of angiotensin IICirc Res200495121167117315539634
- Sanchez-ElsnerTBotellaLMVelascoBLangaCBernabeuCEndoglin expression is regulated by transcriptional cooperation between the hypoxia and transforming growth factor-beta pathwaysJ Biol Chem200227746437994380812228247
- PospisilovaNSemeckyVJamborovaGEndoglin expression in hyper-cholesterolemia and after atorvastatin treatment in apoE-deficient miceJ Pharm Pharm Sci20069338839717207421
- FernandezLAGarrido-MartinEMSanz-RodriguezFTherapeutic action of tranexamic acid in hereditary haemorrhagic telangiectasia (HHT): regulation of ALK-1/endoglin pathway in endothelial cellsThromb Haemost200797225426217264955
- PrietoMRodriguez-PenaAArevaloMEffect of the long-term treatment with trandolapril on endoglin expression in rats with experimental renal fibrosis induced by renal mass reductionKidney Blood Press Res2005281324015475654
- ShyuKGWangBWChenWJKuanPHungCRMechanism of the inhibitory effect of atorvastatin on endoglin expression induced by transforming growth factor-beta1 in cultured cardiac fibroblastsEur J Heart Fail201012321922620156938
- ChungYCHouYCPanACEndoglin (CD105) expression in the development of haemorrhoidsEur J Clin Invest200434210711214764073
- ShyuKGWangBWWuGJLinCMChangHMechanical stretch via transforming growth factor-beta1 activates microRNA208a to regulate endoglin expression in cultured rat cardiac myoblastsEur J Heart Fail2013151364522941949
- DakeishiMShioyaTWadaYGenetic epidemiology of hereditary hemorrhagic telangiectasia in a local community in the northern part of JapanHum Mutat200219214014811793473
- KjeldsenADVasePGreenAHereditary haemorrhagic telangiectasia: a population-based study of prevalence and mortality in Danish patientsJ Intern Med19992451313910095814
- PorteousMEBurnJProctorSJHereditary haemorrhagic telangiectasia: a clinical analysisJ Med Genet19922985275301518020
- WestermannCJRosinaAFDe VriesVde CoteauPAThe prevalence and manifestations of hereditary hemorrhagic telangiectasia in the Afro-Caribbean population of the Netherlands Antilles: a family screeningAm J Med Genet A2003116A432432812522784
- FerenceBAShannonTMWhiteRIJrZawinMBurdgeCMLife-threatening pulmonary hemorrhage with pulmonary arteriovenous malformations and hereditary hemorrhagic telangiectasiaChest19941065138713907956388
- SabbaCPasculliGCirulliAHereditary hemorrhagic teleangiectasia (Rendu-Osler-Weber disease)Minerva Cardioangiol200250322123812107402
- ShovlinCLGuttmacherAEBuscariniEDiagnostic criteria for hereditary hemorrhagic telangiectasia (Rendu-Osler-Weber syndrome)Am J Med Genet2000911666710751092
- TrembathRCThomsonJRMachadoRDClinical and molecular genetic features of pulmonary hypertension in patients with hereditary hemorrhagic telangiectasiaN Engl J Med2001345532533411484689
- FaughnanMEPaldaVAGarcia-TsaoGInternational guidelines for the diagnosis and management of hereditary haemorrhagic telangiectasiaJ Med Genet2011482738719553198
- AbdallaSALetarteMHereditary haemorrhagic telangiectasia: current views on genetics and mechanisms of diseaseJ Med Genet20064329711015879500
- Bayrak-ToydemirPMcDonaldJAkarsuNA fourth locus for hereditary hemorrhagic telangiectasia maps to chromosome 7Am J Med Genet A2006140202155216216969873
- ColeSGBegbieMEWallaceGMShovlinCLA new locus for hereditary haemorrhagic telangiectasia (HHT3) maps to chromosome 5J Med Genet200542757758215994879
- BourdeauACymermanUPaquetMEEndoglin expression is reduced in normal vessels but still detectable in arteriovenous malformations of patients with hereditary hemorrhagic telangiectasia type 1Am J Pathol2000156391192310702408
- SrinivasanSHanesMADickensTA mouse model for hereditary hemorrhagic telangiectasia (HHT) type 2Hum Mol Genet200312547348212588795
- CottinVDupuis-GirodSLescaGCordierJFPulmonary vascular manifestations of hereditary hemorrhagic telangiectasia (Rendu-Osler disease)Respiration200774436137817641482
- GirerdBMontaniDCouletFClinical outcomes of pulmonary arterial hypertension in patients carrying an ACVRL1 (ALK1) mutationAm J Respir Crit Care Med2010181885186120056902
- SimonneauGRobbinsIMBeghettiMUpdated clinical classification of pulmonary hypertensionJ Am Coll Cardiol200954Suppl 1S43S5419555858
- JerkicMKabirMGDaviesAPulmonary hypertension in adult Alk1 heterozygous mice due to oxidative stressCardiovasc Res201192337538421859819
- ToporsianMJerkicMZhouYQSpontaneous adult-onset pulmonary arterial hypertension attributable to increased endothelial oxidative stress in a murine model of hereditary hemorrhagic telangiectasiaArterioscler Thromb Vasc Biol201030350951720042709
- MahmoudMBorthwickGMHislopAAArthurHMEndoglin and activin receptor-like-kinase 1 are co-expressed in the distal vessels of the lung: implications for two familial vascular dysplasias, HHT and PAHLab Invest2009891152519015642
- MacheCJGamillschegAPopperHHHaworthSGEarly-life pulmonary arterial hypertension with subsequent development of diffuse pulmonary arteriovenous malformations in hereditary haemorrhagic telangiectasia type 1Thorax2008631858618156574
- AlbinanaVBernabeu-HerreroMEZarrabeitiaRBernabeuCBotellaLMEstrogen therapy for hereditary haemorrhagic telangiectasia (HHT): effects of raloxifene on Endoglin and ALK1 expression in endothelial cellsThromb Haemost2010103352553420135064
- van CutsemERutgeertsPVantrappenGTreatment of bleeding gastrointestinal vascular malformations with oestrogen-progesteroneLancet199033586959539551970032
- YanivEPreisMShevroJNagerisBHadarTAnti-estrogen therapy for hereditary hemorrhagic telangiectasia – a long-term clinical trialRhinology201149221421621743879
- ShovlinCLSulaimanNLGovaniFSJacksonJEBegbieMEElevated factor VIII in hereditary haemorrhagic telangiectasia (HHT): association with venous thromboembolismThromb Haemost20079851031103918000608
- BowcockSJPatrickHELenalidomide to control gastrointestinal bleeding in hereditary haemorrhagic telangiectasia: potential implications for angiodysplasias?Br J Haematol2009146222022219438503
- KarnezisTTDavidsonTMEfficacy of intranasal bevacizumab (Avastin) treatment in patients with hereditary hemorrhagic telangiectasia-associated epistaxisLaryngoscope2011121363663821344445
- LebrinFSrunSRaymondKThalidomide stimulates vessel maturation and reduces epistaxis in individuals with hereditary hemorrhagic telangiectasiaNat Med201016442042820364125
- SimondsJMillerFMandelJDavidsonTMThe effect of bevacizumab (Avastin) treatment on epistaxis in hereditary hemorrhagic telangiectasiaLaryngoscope2009119598899219194865
- Dupuis-GirodSGinonISaurinJCBevacizumab in patients with hereditary hemorrhagic telangiectasia and severe hepatic vascular malformations and high cardiac outputJAMA2012307994895522396517
- MitchellAAdamsLAMacQuillanGTibballsJvan den DriesenRDelriviereLBevacizumab reverses need for liver transplantation in hereditary hemorrhagic telangiectasiaLiver Transpl200814221021318236396
- HighKAThe gene therapy journey for hemophilia: are we there yet?Blood2012120234482444822829631
- MaguireAMSimonelliFPierceEASafety and efficacy of gene transfer for Leber’s congenital amaurosisN Engl J Med2008358212240224818441370
- ACOG Committee on Practice Bulletins–ObstetricsACOG Practice Bulletin. Diagnosis and management of preeclampsia and eclampsia. Number 33, Jan 2002Obstet Gynecol200299115916716175681
- HutcheonJALisonkovaSJosephKSEpidemiology of pre-eclampsia and the other hypertensive disorders of pregnancyBest Pract Res Clin Obstet Gynaecol201125439140321333604
- MacKayAPBergCJAtrashHKPregnancy-related mortality from preeclampsia and eclampsiaObstet Gynecol200197453353811275024
- MaynardSEKarumanchiSAAngiogenic factors and preeclampsiaSemin Nephrol2011311334621266263
- BellamyLCasasJPHingoraniADWilliamsDJPre-eclampsia and risk of cardiovascular disease and cancer in later life: systematic review and meta-analysisBMJ2007335762797417975258
- OdegardRAVattenLJNilsenSTSalvesenKAAustgulenRRisk factors and clinical manifestations of pre-eclampsiaBJOG2000107111410141611117771
- SibaiBMRamadanMKUstaISalamaMMercerBMFriedmanSAMaternal morbidity and mortality in 442 pregnancies with hemolysis, elevated liver enzymes, and low platelets (HELLP syndrome)Am J Obstet Gynecol19931694100010068238109
- DuckittKHarringtonDRisk factors for pre-eclampsia at antenatal booking: systematic review of controlled studiesBMJ2005330749156515743856
- RedmanCWPlatelets and the beginnings of preeclampsiaN Engl J Med199032374784802374568
- MeekinsJWPijnenborgRHanssensMMcFadyenIRvan AssheAA study of placental bed spiral arteries and trophoblast invasion in normal and severe pre-eclamptic pregnanciesBr J Obstet Gynaecol199410186696747947500
- ZhouYDamskyCHFisherSJPreeclampsia is associated with failure of human cytotrophoblasts to mimic a vascular adhesion phenotype. One cause of defective endovascular invasion in this syndrome?J Clin Invest1997999215221649151787
- ZhouYFisherSJJanatpourMHuman cytotrophoblasts adopt a vascular phenotype as they differentiate. A strategy for successful endovascular invasion?J Clin Invest1997999213921519151786
- SalafiaCMPezzulloJCGhidiniALopez-ZenoJAWhittingtonSSClinical correlations of patterns of placental pathology in preterm pre-eclampsiaPlacenta199819167729481787
- AhmadSAhmedAElevated placental soluble vascular endothelial growth factor receptor-1 inhibits angiogenesis in preeclampsiaCirc Res200495988489115472115
- MaynardSEMinJYMerchanJExcess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsiaJ Clin Invest2003111564965812618519
- GilbertJSRyanMJLaMarcaBBSedeekMMurphySRGrangerJPPathophysiology of hypertension during preeclampsia: linking placental ischemia with endothelial dysfunctionAm J Physiol Heart Circ Physiol20082942H541H55018055511
- KendallRLThomasKAInhibition of vascular endothelial cell growth factor activity by an endogenously encoded soluble receptorProc Natl Acad Sci USA1993902210705107098248162
- KapurNKHeffernanKSYunisAAElevated soluble fms-like tyrosine kinase-1 levels in acute coronary occlusionArterioscler Thromb Vasc Biol201131244345021071694
- ClarkDESmithSKHeYA vascular endothelial growth factor antagonist is produced by the human placenta and released into the maternal circulationBiol Reprod1998596154015489828203
- LevineRJLamCQianCSoluble endoglin and other circulating antiangiogenic factors in preeclampsiaN Engl J Med200635510992100516957146
- LevineRJMaynardSEQianCCirculating angiogenic factors and the risk of preeclampsiaN Engl J Med2004350767268314764923
- Valbuena-DiezACBlancoFJOujoBOxysterol-induced soluble endoglin release and its involvement in hypertensionCirculation2012126222612262423110859
- JerkicMRivas-ElenaJVPrietoMEndoglin regulates nitric oxide-dependent vasodilatationFASEB J200418360961114734648
- Lopez-NovoaJMBernabeuCThe physiological role of endoglin in the cardiovascular systemAm J Physiol Heart Circ Physiol20102994H959H97420656886
- GuYLewisDFWangYPlacental productions and expressions of soluble endoglin, soluble fms-like tyrosine kinase receptor-1, and placental growth factor in normal and preeclamptic pregnanciesJ Clin Endocrinol Metab200893126026617956952
- GilbertJSGilbertSAAranyMGrangerJPHypertension produced by placental ischemia in pregnant rats is associated with increased soluble endoglin expressionHypertension200953239940319075097
- CudmoreMAhmadSAl-AniBNegative regulation of soluble Flt-1 and soluble endoglin release by heme oxygenase-1Circulation2007115131789179717389265
- BergmannAAhmadSCudmoreMReduction of circulating soluble Flt-1 alleviates preeclampsia-like symptoms in a mouse modelJ Cell Mol Med2010146B1857186719538465
- LiZZhangYYing MaJRecombinant vascular endothelial growth factor 121 attenuates hypertension and improves kidney damage in a rat model of preeclampsiaHypertension200750468669217724276
- SuzukiHOhkuchiAMatsubaraSEffect of recombinant placental growth factor 2 on hypertension induced by full-length mouse soluble fms-like tyrosine kinase 1 adenoviral vector in pregnant miceHypertension20095451129113519786649
- ThadhaniRKisnerTHagmannHPilot study of extracorpo-real removal of soluble fms-like tyrosine kinase 1 in preeclampsiaCirculation2011124894095021810665
- GeorgeEMCockrellKAranayMCsongradiEStecDEGrangerJPInduction of heme oxygenase 1 attenuates placental ischemia-induced hypertensionHypertension201157594194821383306
- DevyLHuangLNaaLSelective inhibition of matrix metalloproteinase-14 blocks tumor growth, invasion, and angiogenesisCancer Res20096941517152619208838
- OverallCMKleifeldOTumour microenvironment – opinion: validating matrix metalloproteinases as drug targets and anti-targets for cancer therapyNat Rev Cancer20066322723916498445
- CowieMRMosterdAWoodDAThe epidemiology of heart failureEur Heart J19971822082259043837
- Lloyd-JonesDAdamsRCarnethonMHeart disease and stroke statistics – 2009 update: a report from the American Heart Association Statistics Committee and Stroke Statistics SubcommitteeCirculation20091193e21e18119075105
- BerkBCFujiwaraKLehouxSECM remodeling in hypertensive heart diseaseJ Clin Invest2007117356857517332884
- LeaskATGFbeta, cardiac fibroblasts, and the fibrotic responseCardiovasc Res200774220721216919613
- RitzenthalerJDGoldsteinRHFineASmithBDRegulation of the alpha 1(I) collagen promoter via a transforming growth factor-beta activation elementJ Biol Chem19932681813625136318514794
- QiaoBPadillaSRBenyaPDTransforming growth factor (TGF)-beta-activated kinase 1 mimics and mediates TGF-beta-induced stimulation of type II collagen synthesis in chondrocytes independent of Col2a1 transcription and Smad3 signalingJ Biol Chem200528017175621757115743758
- LeaskAHolmesABlackCMAbrahamDJConnective tissue growth factor gene regulation. Requirements for its induction by transforming growth factor-beta 2 in fibroblastsJ Biol Chem200327815130081301512571253
- ChenMMLamAAbrahamJASchreinerGFJolyAHCTGF expression is induced by TGF- beta in cardiac fibroblasts and cardiac myocytes: a potential role in heart fibrosisJ Mol Cell Cardiol200032101805181911013125
- Chuva de Sousa LopesSMFeijenAKorvingJConnective tissue growth factor expression and Smad signaling during mouse heart development and myocardial infarctionDev Dyn2004231354255015376321
- FrantzSHuKAdamekATransforming growth factor beta inhibition increases mortality and left ventricular dilatation after myocardial infarctionBasic Res Cardiol2008103548549218651091
- KuwaharaFKaiHTokudaKTransforming growth factor-beta function blocking prevents myocardial fibrosis and diastolic dysfunction in pressure-overloaded ratsCirculation2002106113013512093782
- KapurNKHeffernanKSYunisAAUsefulness of soluble endoglin as a noninvasive measure of left ventricular filling pressure in heart failureAm J Cardiol2010106121770177621126621
- YanavitskiMGivertzMMNovel biomarkers in acute heart failureCurr Heart Fail Rep20118320621121681444
- PetersonJTLiHDillonLBryantJWEvolution of matrix metalloprotease and tissue inhibitor expression during heart failure progression in the infarcted ratCardiovasc Res200046230731510773235
- ZeisbergMHanaiJSugimotoHBMP-7 counteracts TGF-beta1-induced epithelial-to-mesenchymal transition and reverses chronic renal injuryNat Med20039796496812808448
- Cruz-GonzalezIPabonPRodriguez-BarberoAIdentification of serum endoglin as a novel prognostic marker after acute myocardial infarctionJ Cell Mol Med200812395596118494936
- KapurNKShenoyCYunisAADistinct effects of unfractionated heparin versus bivalirudin on circulating angiogenic peptidesPLoS One201274e3434422509290
- NachtigalPZemankova VecerovaLRathouskaJStraskyZThe role of endoglin in atherosclerosisAtherosclerosis2012224141122460049