Abstract
In humans, structural and functional changes attributable to aging are more visibly evident in the skin than in any other organ. Estrogens have significant effects on skin physiology and modulate epidermal keratinocytes, dermal fibroblasts and melanocytes, in addition to skin appendages including the hair follicle and the sebaceous gland. Importantly, skin aging can be significantly delayed by the administration of estrogen. This paper reviews the effects of estrogens on skin and the mechanisms by which estrogens can alleviate the changes due to aging that occur in human skin. The relevance of estrogen replacement therapy (HRT) in post-menopausal women and the potential value of selective estrogen receptor modulators (SERMs) as a therapy for diminishing skin aging are also highlighted.
Keywords:
Estrogens and skin biology
A number of studies have shown that estrogens have many important beneficial and protective roles in skin physiology (reviewed in CitationThornton 2002, Citation2005). They have been shown to accelerate cutaneous wound healing (CitationAshcroft and Ashworth 2003), while a significant number of women notice an improvement in inflammatory skin disorders such as psoriasis during pregnancy (CitationDunna and Finlay 1989; CitationBoyd et al 1996; CitationRaychaudhuri et al 2003). Estrogens also offer some degree of protection against skin photoaging (CitationWeinstock 1994; CitationTsukahara et al 2001, Citation2004) and epidemiological studies indicate that the mortality rates from both non-melanoma skin cancers (CitationWeinstock 1994) and melanoma (CitationMiller and MacNeil 1997) are significantly lower in women.
Many of the effects of estrogen on human skin are based on the changes that are seen in post-menopausal women, with a number of studies documenting the differences seen following the menopause, although there is also a variation in skin thickness during the menstrual cycle, with skin thickness lowest at the start of the menstrual cycle, when estrogen and progesterone levels are low, which then increases with the rising levels of estrogen (CitationEisenbeiss 1998). Many women report a sudden onset of skin aging several months after menopausal symptoms begin. The menopause causes hypoestrogenism, accelerating age-related deterioration, which results in thinner skin, an increase in number and depth of wrinkles, increased skin dryness, and decreased skin firmness and elasticity (CitationBrincat 2000). Hormone replacement therapy (HRT) has been shown to increase epidermal hydration, skin elasticity, skin thickness (CitationSator et al 2001), and also reduces skin wrinkles (CitationPhillips et al 2001). Furthermore, the content and quality of collagen and the level of vascularization is enhanced (CitationBrincat et al 1987).
The epidermis
Epidermal thinning is associated with aging, and topical estradiol has been shown to reduce epidermal thinning in aging skin and maintain skin thickness (CitationShah and Maibach 2001). Recent studies have confirmed that administration of estradiol to gonadectomised mice increases epidermal thickness in both sexes (CitationAzzi et al 2005).
In women an increase in the mitotic activity of epidermal keratinocytes occurs in response to estrogens (CitationPunnonen 1972). Furthermore, the stimulation of proliferation and DNA synthesis of human epidermal keratinocytes by estrogens has been demonstrated in vitro (CitationUrano et al 1995; CitationKanda and Watanabe 2004; CitationVerdier-Sevrain et al 2004) and cultured human epidermal keratinocytes have high affinity estrogen binding sites (CitationVerdier-Sevrain et al 2004). Specific antibodies for the two distinct intracellular estrogen receptors (ERα and ERβ) have shown that epidermal keratinocytes from human scalp skin of both sexes predominately express ERβ in situ (CitationThornton et al 2003).
However, primary cultures of epidermal keratinocytes from female breast skin express both ERα and ERβ in vitro (see ).
Figure 1 The expression of ERα and ERβ in human epidermal keratinocytes cultured from human female breast skin. Both nuclear and cytoplasmic expression of ERα and ERβ, with intense staining in the perinuclear region was seen in epidermal keratinocytes derived from female breast skin.
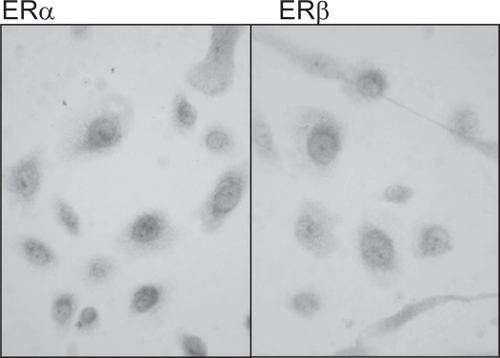
Another study has demonstrated mRNA and protein for ERβ and not ERα is expressed in cultured human epidermal keratinocytes (CitationKanda and Watanabe 2003a, Citation2003b), while a different group have shown that both ERα and ERβ are expressed in cultured human neonatal foreskin keratinocytes (CitationVerdier-Sevrain et al 2004). These differences may be due to differences in anatomical site, since the number of estrogen receptors has been shown to differ with anatomical site, with face skin homogenates having a greater number of receptors than breast or thigh skin (CitationHasselquist et al 1980). A more recent study has shown that there are marked differences in the expression of the two estrogen receptors in human skin derived from different anatomical sites including scalp, breast and abdomen (CitationNelson 2006); in contrast to scalp, which predominantly expresses ERβ (CitationThornton et al 2003), breast skin predominantly expresses ERα in situ (CitationNelson 2006).
Recently, it has been reported that 17β-estradiol conjugated with BSA can stimulate proliferation and DNA synthesis of cultured human keratinocytes (CitationKanda and Watanabe 2004), suggesting that estradiol can stimulate cell proliferation via a cell membrane receptor. This is further supported by another group who demonstrated that incubation of cultured human epidermal keratinocytes with estradiol increased phosphorylation levels of ERK1 and ERK2 kinases within 15 minutes (CitationVerdier-Sevrain et al 2004).
A protective role for estrogens on the human epidermis may also exist since estradiol prevents apoptosis induced by hydrogen peroxide in cultured epidermal keratinocytes by promoting the expression of bcl-2 (CitationKanda and Watanabe 2003a). Furthermore, estradiol inhibits the production of chemokines involved in the attraction of macrophages, which are important in inflammation (CitationKanda and Watanabe 2003b).
The dermis
Autoradiography following administration of systemic or topical estrogen has demonstrated that radio-labeled estradiol binds to dermal fibroblasts in rat and mouse skin (CitationStumpf et al 1974; CitationBidmon et al 1990). More recently, specific antibodies have demonstrated ERβ, but not ERα is expressed by dermal fibroblasts in the papillary dermis of human scalp skin in both sexes (CitationThornton et al 2003), whereas primary cultures of human dermal fibroblasts from female skin have been shown to express both mRNA and protein for ERα and ERβ (CitationHaczynski et al 2002). Although dermal fibroblasts co-express both estrogen receptors, immunocytochemistry showed some variation in their expression; ERβ was predominately nuclear, while ERα was expressed in both the cytoplasm and the nucleus. Furthermore, mRNA levels for ERβ were higher than levels of ERα. A later study by the same group has demonstrated that estradiol up-regulates the expression of ERβ in dermal fibroblasts cultured from postmenopausal women (CitationSurazynski et al 2003), thereby altering the ratio of ERα to ERβ.
Estrogens have been shown to influence skin thickness by stimulating collagen synthesis, maturation and turnover in rats (CitationSmith and Allison 1966) and female guinea pigs (CitationHenneman 1968). In mice, estrogen administration increased hyaluronic acid synthesis by 70% in two weeks (CitationSobel 1965), leading to increased dermal water content. In wound healing models, estrogen reduces wound size and stimulates matrix deposition in both human and murine skin (CitationAshcroft et al 1999; CitationAshcroft and Ashworth 2003), highlighting the effects of estrogens on the dermal fibroblasts.
Estrogen is also believed to provide some protection against photoaging. Ultraviolet (UV)-B exposure is associated with up-regulation of matrix metalloproteinase (MMP) production, leading to an increase in collagen degradation and is also thought to decrease type I and type III collagen synthesis (reviewed in CitationBaumann 2005). CitationTsukahara and colleagues (2001) demonstrated that ovariectomized rats exposed to UVB radiation display an increased formation of deep wrinkles, decreased skin elasticity and marked and advanced curling of dermal elastic fibers compared to rats with normal estrogen levels. Furthermore, the same authors demonstrated that in hairless mice ovariectomy alone decreases skin elasticity, with a further significant increase in skin sagging in the ovariectomized animals exposed to 12 weeks of UV-A irradiation and a significant increase in skin wrinkling following UV-B irradiation (CitationTsukahara et al 2004). In each case this was associated with an increase in elastase activity. This led the authors to suggest that ovariectomy causes an increase in elastase mediated degradation of dermal elastic fibers with a subsequent reduction in skin elasticity, thus explaining the skin changes seen in post-menopausal women.
In vitro, murine dermal fibroblasts have been shown to increase collagen synthesis by 76% in response to estrogen (CitationHosokawa et al 1981). Similarly, incubation of cultured human dermal fibroblasts with 17β-estradiol increased type I procollagen secretion (CitationSon et al 2005). This effect was reversed when estrogen was administered with TGF-β1 antibodies, suggesting that TGF-β1 may play a role in increasing collagen secretion in response to estrogen. Other authors have also demonstrated an increase in collagen biosynthesis in vitro in response to 17β-estradiol by measuring radio-labeled proline incorporation (CitationSurazynski et al 2003).
The hair follicle
Hair follicles have a built in rhythm of activity that results in the periodic regeneration of new hairs and the molting of old ones. This rhythm appears to be intrinsic, although it can be influenced by hormonal or other systemic factors. Estrogens significantly influence the hair cycle in a number of mammalian species by inhibiting hair growth (reviewed in CitationThornton 2002, Citation2005; Ohnemus et al 2006). However, the effects of estrogens on the human hair cycle are less clear.
In contrast to the inhibition observed in other mammals, limited studies suggest that estrogens may have a stimulatory effect on human hair growth. During pregnancy there is an increase in the ratio of hair follicles in the growing phase of the cycle; after parturition, these follicles enter the resting phase, resulting in increased hair shedding and a transient thinning of the hair (CitationLynfield 1960). These events have been ascribed to the changes in the level of circulating estrogen during, and immediately following pregnancy. However, it is difficult to accredit this phenomenon entirely to the rise in plasma estrogen as several other hormones and growth factors that modulate hair growth are also altered at this time.
There is some limited trichogram data to suggest that estrogens decrease the resting phase and prolong the growing phase of the hair cycle, hence estrogens are used in the treatment of female pattern hair loss in some countries (CitationSinclair 1999). Further evidence for a stimulatory effect of estrogens on human hair growth comes from the treatment of women with tamoxifen which may result in scalp hair thinning or recession in some women (CitationGateley and Bundred 1997; CitationAyoub et al 1997). Likewise, a common treatment-related side effect of aromatase inhibitors, which inhibit the synthesis of estrogen, is scalp hair thinning in women (CitationBuzdar et al 2001; CitationSimpson et al 2004).
Recent in vitro studies have shown that 17β-estradiol inhibits female scalp hair shaft elongation (CitationNelson 2006), although stimulation occurs in hair follicles derived from frontotemporal male scalp (CitationConrad et al 2004; CitationConrad and Paus 2004). In addition, in female hair follicles the phytoestrogen, genistein inhibits hair shaft elongation to a similar extent as 17β-estradiol. Since genistein preferentially binds to ERβ, this opens the possibility that the inhibition of hair growth in response to 17β-estradiol may be mediated via ERβ rather than ERα (CitationNelson 2006). Therefore the development of selective estrogen receptor ligands may provide important clinical applications for the prevention and treatment of disorders of hair growth.
Melanocytes and melanoma
Chloasma is a common hyperpigmentation of the face seen in pregnant women, often accompanied by increased pigmentation in other areas including the areolae, linea alba and perineal skin, all of which usually fade following parturition (CitationKroumpouzos and Cohen 2001). Oral contraceptives containing estrogen can also result in hyperpigmentation of the face (CitationWade et al 1978) and ointments containing estrogen can produce intense pigmentation of the genitals, mammary areola and linea alba of the abdomen in male and female infants (CitationBeas et al 1969).
The mean age of presentation of malignant melanoma in women is the early fifties, which correlates with the onset of the menopause (CitationDurvasula et al 2002). Melanoma has traditionally been considered to be an estrogen receptor-positive tumor, whose prognosis is adversely affected by estrogen, whether during pregnancy or in association with the oral contraceptive pill or HRT. Recent evidence now disputes this and the relationship between estrogens and malignant melanoma remains controversial.
There is a significant lack of information in relation to HRT and melanoma and the use of steroid hormones in the management of melanoma is limited. Steroid hormone binding activity has been demonstrated in some human melanomas, but only a small percentage of melanomas respond to hormonal manipulation (CitationNeifeld 1996). The relative expressions of the different estrogen receptors in malignant melanoma have yet to be explored, which may be of significance since alterations in the ratio of ERα and ERβ have been suggested in the development and progression of other cancers.
The menopause
Post-menopausal skin has been shown to have increased dryness (CitationSator et al 2004), decreased elasticity (CitationHenry et al 1997; CitationSumino et al 2004), and increased wrinkling (CitationDunn et al 1997). Indeed, many of the effects of estrogen on the human skin have been described based on the changes that are seen following the menopause. Estrogen receptor expression has been shown to be reduced following the menopause (CitationPunnonen et al 1980; CitationNelson and Bulun 2001).
In postmenopausal women skin thickness decreases by 1.13% per postmenopausal year, with an associated decrease in collagen content (2% per post-menopausal year) (CitationBrincat et al 1987). The collagen content (types I and III) of skin is thought to decrease by as much as 30% in the first five years following the menopause (CitationBrincat et al 1985; CitationAffinito et al 1999). Interestingly, this decrease in skin collagen content parallels the reduction in bone mass seen in post-menopausal women (CitationBrincat et al 1987). The decrease in skin thickness and collagen content seen in elderly females appears to correlate more closely with the period of estrogen deficiency than with chronological age (CitationBrincat et al 1985, Citation1987; CitationAffinito et al 1999).
In contrast, another study has demonstrated a closer relationship between chronological age and reduction in skin collagen, than time since menopause (CitationCastelo-Branco et al 1992). However, for the patients in this study the time spent post-menopause was much shorter, therefore the long-term effects of estrogen deficiency may not have become apparent.
A difference in collagen subtypes has also been documented in post-menopausal women. When evaluated by immunohistochemistry, compared to pre-menopausal women, post-menopausal women demonstrate a decrease in collagen types I and III and a reduction in the typeIII/type I ratio within the dermis. Again this correlates more closely with period of estrogen deficiency than with chronological age (CitationAffinito et al 1999). Similar findings have been reported in the arcus tendineous fasciae pelvis, where the collagen type I and type I/type III ratio was decreased in postmenopausal women compared to premenopausal women (CitationMoalli et al 2004). Interestingly, despite these changes the total amount of collagen was not altered.
Hormone replacement therapy
Further understanding of estrogen effects on human skin is derived from comparisons between post-menopausal women taking estrogen replacement therapy and those who have not. An early study reported an increase in epidermal thickness in human female skin in response to six months of oral estrogens (CitationPunnonen 1972). Although a different study did not demonstrate an increase in epidermal thickness, CitationMaheux et al (1994) demonstrated an increased keratinocyte volume and more defined rete ridges in response to oral estrogen.
More recent studies have confirmed that topical estrogen increases epidermal thickness (CitationSon et al 2005; CitationPatriarca et al 2007). Furthermore, CitationSon et al (2005) demonstrated an increase in keratinocyte proliferation and epidermal thickness in response to only two weeks of topical estrogen in the skin of elderly males and females. HRT has also been shown to improve skin hydration when estrogen is administered systemically (CitationPierard-Franchimont et al 1995) or topically (CitationSator et al 2001). Topical application is an efficient method, since due to its small molecular size; estrogen easily penetrates the stratum corneum (CitationDraelos 2005). The increased hydration is believed to be due in part to an increase in the water-holding capacity of the stratum corneum (CitationPierard-Franchimont et al 1995). Additionally, dermal hyaluronic acid and acid mucopolysaccharide levels are increased, which will also improve hydration (reviewed in CitationShah and Maibach 2001).
In a randomized, double-blind, placebo-controlled trial, CitationMaheux et al (1994), demonstrated that after 12 months of oral estrogen therapy, dermal thickness was increased by 30%, resulting in increased skin thickness in post-menopausal nuns. Skin biopsies were taken from the thigh thus eliminating confounding factors such as sun exposure and smoking (CitationMaheux et al 1994). Other authors have also demonstrated increased skin thickness following HRT (CitationBrincat et al 1985; CitationCallens et al 1996; CitationSator et al 2001).
In addition to increased skin thickness, estrogen has also been shown to increase the collagen content of the skin. A randomized, double-blind, placebo-controlled trial carried out by CitationSauerbonn et al (2000) demonstrated an increase of 6.49% in skin collagen fiber content after six months of treatment with an oral combination of estrogen and cytoproterone acetate. Other authors have demonstrated an increase in overall collagen content (CitationBrincat et al 1987) and specifically collagen type III (CitationSavvas et al 1993; CitationSchmidt et al 1996). Another study found no difference in skin thickness or collagen content in upper arm or abdominal skin following oral estrogen therapy (CitationHaapasaari et al 1997); however the patients in this study had only a short period of estrogen deficiency (median time post-menopause 12 months), which may explain this finding. Importantly, the increase in skin collagen content in response to estrogen therapy appears to be related to the initial collagen content. CitationBrincat et al (1987) demonstrated that in women with low skin collagen content, estrogen is initially therapeutic, but later becomes purely prophylactic, whereas for women in the early years post-menopause with only mild collagen loss, estrogen is of prophylactic value only. Indeed, optimum collagen content was seen following two years of treatment in a study which examined the effect of estradiol implants over a period ranging from two to ten years (CitationBrincat et al 1987).
The use of topical estrogen has also been shown to increase skin collagen. CitationVarila et al (1995) demonstrated an increase in collagen content following topical estrogen administration. They also demonstrated an increase in collagen synthesis as shown by increased type I and type III procollagen levels. However, with topical administration, the effect was limited to the area to which the estrogen was directly applied (CitationVarila et al 1995; CitationSchmidt et al 1996). A more recent study examining the effect of estrogen on human skin has demonstrated an increase in collagen content in buttock skin from elderly males and females treated with topical estradiol (CitationSon et al. 2005). Type I procollagen mRNA and protein were significantly increased in males and females, with a significantly greater increase in females compared to males. Similarly, immunohistochemical expression for type I procollagen protein was increased following estradiol treatment. The same group also demonstrated an increase in tropoelastin and fibrillin mRNA and protein in response to estradiol and suggested this can be equated to an increase in elastic fibers. Furthermore, they demonstrated an increase in TGF-β and TGF-β type II receptor mRNA and protein expression in response to estrogen therapy, although TGF-β type-I receptor expression was not increased. Since TGF-β is a growth factor known to stimulate fibroblast proliferation and extracellular matrix (ECM) secretion, they suggested that this is the mechanism by which estrogen increases ECM secretion (CitationSon et al 2005). Furthermore, a decrease in MMP-1 expression was demonstrated in response to topical estrogen, which may also explain increased collagen content in estrogen treated skin. Interestingly, when topical estradiol was applied to the skin of young males, similar effects were also demonstrated (CitationSon et al 2005).
Skin wrinkling is synonymous with aging, but is also affected by environmental and hormonal factors. It occurs due to decreased skin elasticity as a result of elastic degeneration and loss of connective tissue (reviewed in CitationShah and Maibach 2001). In early postmenopausal women monitored for five years, skin elasticity has been shown to decrease by 1.5% per year, a change not seen in women on HRT (CitationHenry et al 1997). Furthermore, conjugated equine estrogen has been shown to increase forearm skin elasticity in postmenopausal women (CitationSumino et al 2004). CitationPunnonen et al (1987) demonstrated that topical estrogen thickens elastic fibers in the papillary dermis, increases their number and improves their orientation, although this effect has not been shown by other authors (CitationBrincat et al 2005). This led to the suggestion that estrogen deficiency plays a role in wrinkle formation and in a large cohort study it was shown that wrinkling is reduced in postmenopausal women administered estrogen (CitationDunn et al 1997).
In post-menopausal women skin wrinkling has also been shown to be improved by topical estrogen therapy after a 24-week treatment period (CitationCreidi et al 1994). However, other authors have demonstrated an improvement in skin elasticity, although there was no improvement in wrinkling in smokers (CitationCastelo-Branco et al 1998). Furthermore, estrogen was not shown to consistently improve the appearance of facial skin over a five-year period, suggesting the effects may be reduced in sun-exposed skin (CitationPierard-Franchimont et al 1999).
More recently, CitationWolff et al (2005) demonstrated decreased facial wrinkling and increased skin rigidity following long-term oral estrogen administration. Women who were at least five years post-menopause and had either taken oral estrogen continuously, or had never used HRT, were examined by a single blind observer who assessed wrinkle formation using an objective photographic wrinkle score and measured skin rigidity using a durometer. They demonstrated that the average wrinkle score was significantly lower in the hormone treated group. Skin rigidity was also significantly lower in the group using HRT (CitationWolff et al 2005). This suggests that estrogen therapy has long-term benefits on skin and supports the use of early and continuous HRT in preventing detrimental skin changes (CitationNaftolin 2005).
Although estrogen administration has been shown to have positive effects on skin by delaying or preventing skin aging manifestations (CitationBrincat 2000; CitationSator et al 2004), understanding the effects of estrogen on skin is complicated by a number of factors. Firstly, when interpreting the results of studies looking at the effects of estrogen on skin it is important to remember that few randomized trials have been carried out and that the majority of trials have been observational studies. The use of different estrogen preparations and doses with, or without, the concomitant use of progesterone or testosterone also complicates this issue. Progestagens are combined with estrogen compounds and given cyclically or continuously since estrogen therapy alone may result in endometrial hyperplasia. Estrogen monotherapy can be used following hysterectomy (CitationSator et al 2004). Thus, making true comparisons and judgments of the isolated effects of estrogen are difficult. Indeed, topical progesterone cream (2%) alone has been shown to increase skin elasticity and decrease wrinkling in peri- and post-menopausal women (CitationHolzer et al 2005).
Controversies surrounding the risks associated with the use of hormone therapy also complicate the issue. HRT was initially introduced to improve menopausal symptoms, reduce the risk of osteoporosis and was thought to reduce the risk of cardiovascular events (CitationStampfer et al 1991). However, a recent study has suggested that HRT may increase the risk of cardiovascular events (CitationGrady et al 2002). It has also been reported that HRT may increase the risk of breast cancer (CitationRossouw et al 2002) and this has led to a significant reconsideration of the risks and benefits of systemic HRT. In response to this, recommendations for the use of HRT have become more stringent and it is now generally accepted that HRT should only be used to relieve menopausal symptoms, in the short-term (CitationHall and Phillips 2004). This means that justifying the use of randomized clinical trials to assess the effects of systemic estrogen on skin is difficult from a risk-benefit point of view.
Despite the controversy that surrounds the use of HRT in post-menopausal women; many authors suggest that the benefits of HRT in skin necessitate the need for further studies of its effects, not only in skin, but in other organ systems (CitationDraelos 2005; CitationNaftolin 2005; CitationWolff et al 2005). Furthermore, as the visual effects that are seen in the skin following the menopause may physiologically mirror the internal effects in other tissues (bone, cardiovascular system, cancer), skin may represent an easily accessible model. The use of topical estrogen could also be considered, but again this requires further investigation to determine the optimal dose for local benefits, while ensuring that systemic effects are avoided (CitationVerdier-Sevrain et al 2006).
Skin and aging
Although estrogen deficiency is associated with skin changes, intrinsic aging also affects skin physiology. Skin undergoes profound changes associated with aging. This is most apparent in the face and other sun-exposed areas. Aging is associated with a decrease in skin thickness due to atrophy of the epidermis, dermis and subcutaneous fat. This is associated with dryness, wrinkling and an increased incidence of proliferative lesions. In the epidermis, aging is associated with a decrease in epidermal thickness, flattening of the dermal papilla and a decrease in melanocyte and Langerhans cell density. Within the dermis, increasing age leads to reduced fibroblast activity, reduced collagen and hyaluronic acid content, more fragmented elastin fibers and decreased vascularity (CitationAshcroft et al 1995). Aging skin has also been shown to have increased pro-enzyme MMP-2 expression (CitationAshcroft et al 1997), suggesting that aged skin is primed for tissue breakdown. A number of environmental factors such as sun exposure and smoking also affect the rate at which skin changes.
Mechanism of estrogen action
Estrogens are the terminal ligand in the biosynthetic pathway of gonadal steroid hormones and are synthesized from androgens by the loss of the C-19 angular methyl group and the formation of an aromatic A ring by the aromatase complex (CitationPayne and Hales 2004). Estrone is derived from androstenedione, whereas estradiol is formed from testosterone. Estrone and estradiol are interconvertable due to different isoenzymes of 17β-hydroxysteroid dehydrogenase (CitationLabrie 2004).
The principle source of estrogen biosynthesis is the ovary in females of reproductive age. In men, estradiol can be produced in peripheral tissues by the actions of aromatase on testosterone (CitationSimpson 1998). Humans, along with some other primates are unusual in that the adrenal cortex secretes large quantities of adrenal androgens, including dehydroepiandrosterone (DHEA), which can then be converted into active steroids in peripheral tissues providing they have the appropriate enzymes (CitationLabrie 2004). DHEA synthesized in the adrenal zona reticularis serves as the main precursor of active estrogens in post-menopausal women. Notwithstanding, the production of DHEA also decreases with age. Serum concentrations are low prior to puberty, reaching a peak in adulthood. However, throughout adult life, levels decline and by the 7th decade are reduced to only 10%–20% of the peak concentrations in both sexes (CitationParker et al 1997). Therefore, with aging the precursor steroids for peripheral estrogen biosynthesis are reduced.
Estrogen receptor: non-genomic signaling
While the best described mode of estrogen signaling is mediated via two related proteins (ERα and ERβ) that belong to the nuclear receptor superfamily (see below), a further level of complexity in estrogen signaling is now apparent due to studies demonstrating rapid effects in response to estrogens that cannot be attributed to signaling via the classical intracellular nuclear receptors and their genomic pathways (CitationLevin 2002). These rapid effects, at rates much faster than can be attributed to genomic signaling, have led to the view that cell membrane forms of estrogen receptors exist that are coupled to cytosolic signal transduction proteins able to initiate different signaling cascades via conventional second messengers (CitationNadal et al 2001). Estrogens have been shown to activate second messengers such as adenylate cyclase and cAMP (CitationAronica et al 1994), phospholipase C (CitationLieberherr et al 1993), protein kinase C (CitationMarino et al 2002) and the mitogen-activated protein kinase (MAPK) (CitationShaul 1999; CitationRussel et al 2000). Interestingly, estrogen can also activate ligand or voltage-gated ion channels (CitationPappas et al 1995; CitationRazandi et al 1999; CitationNadal et al 2001) resulting in the increase in levels of intracellular calcium (CitationBenten et al 2001).
The term “activators of nongenotropic estrogen-like signaling (ANGELS)” has been introduced to define small nonphenolic molecules that mimic the nongenotropic actions of estrogens, but lack their classical genotropic effects (CitationKousteni et al 2003). ANGELS appear to represent a novel class of compounds that differ mechanistically from classical estrogens. One such compound is estren, which is completely devoid of classical genotropic actions (CitationKousteni et al 2003), but can reverse bone loss in sex steroid-deficient mice (CitationKousteni et al 2002).
Classical mechanism of action: genomic signaling
Two distinct intracellular estrogen receptors (ERα and ERβ) have been identified that belong to the superfamily family of nuclear hormone receptors (CitationMoverare et al 2002). It is now clear that estrogens have important effects on many non-reproductive tissues and the expression of the two estrogen receptors is tissue dependent. In addition to the male and female reproductive tissues, ERα and ERβ are expressed in tissues as diverse as bone, brain, lung, bladder, thymus, pituitary, hypothalamus, heart, kidney, adrenal, the cardiovascular system and the skin including the hair follicle (reviewed in CitationThornton 2002, Citation2005).
The two estrogen receptors are distinct proteins encoded by separate genes that are located on different chromosomes (CitationEnmark et al 1997). The ERα and ERβ proteins share approximately 97% homology in the DNA binding domain, with only a few amino acids differing in this region. However, in the ligand binding domain they only share 59% homology (CitationGustafsson 1999), while they share little homology in the amino terminal domain, hinge domain and COOH domain (CitationMoverare et al 2002). With such a difference in the ligand binding domain, it could be anticipated that the receptors would bind estradiol with different affinities; this however is not the case, since 17β-estradiol has a similar affinity for both receptors. Ligand binding analysis has shown an average dissociation constant of 0.5 nM for 17β-estradiol on mouse ERβ and 0.2 nM on mouse ERα (CitationGiguere 1998), suggesting ERβ has a slightly lower affinity for 17β-estradiol. Many synthetic estrogens also bind both receptors with a similar affinity (CitationKuiper et al 1997).
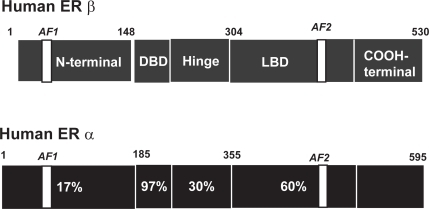
Similar to all intracellular steroid receptors, ERα and ERβ are ligand activated nuclear transcription factors that enhance target-gene transcription upon binding to chromatin. Activation of the target gene by 17β-estradiol activates both ERα and ERβ to increase transcriptional activities when dimers of the liganded receptors bind to estrogen response elements (ERE) which are specific DNA palindrome sequences located in the promoter region of estrogen-regulated target genes (CitationKlinge 2001). Activation of transcription also requires the recruitment of a large coactivator complex composed of p160 coactivators including GRIP1 and SRC-1 and the histone acetyltransferases p300/CREB-binding protein and pCAF (CitationWebb et al 2003). While the DNA binding domain mediates ERE recognition, the mediation of coactivator recruitment occurs via distinct activation functions (AF) located in the N-terminal domain (AF-1) and the ligand binding domain (AF-2) (). Coactivators are tissue-specific and there is some evidence that ERα and ERβ differ in their requirements for coactivators in a cell and tissue dependent manner (CitationSmith and O’Malley 2004).
Figure 2 Schematic representation of the modular structures of ERα and ERβ The modular structure of the two estrogen receptors (ERα and ERβ) illustrating several distinct functional domains. The numbers within the ERα domains represent the percentage homology between the two receptors. There is little homology between activation function-1 (AF-1) in the N-terminal domain, whereas the core sequences of AF-2 in the ligand binding domain (LBD) are identical. The numbers above each receptor represent the amino acid position of each boundary. The DNA binding domain (DBD) shows the greatest degree of homology.
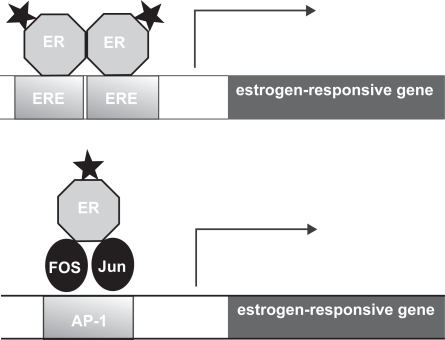
In addition to the well characterized mode of action via EREs, estrogen receptors may also interact with other transcription factors bound to their cognate DNA binding sites through protein-protein interactions. For example, both ERα and ERβ enhance the transcription of genes that contain AP-1 sites, the cognate binding site for the Jun/Fos complex (CitationPaech et al 1997) (see ). ERα and ERβ can also enhance the transcription of genes that contain binding sites for other transcription factors (CitationElgort et al 1996). While the precise mechanism of ER action at AP-1 sites is still unclear, studies suggest that there are striking differences from ER action at EREs (CitationWebb et al 1999).
Figure 3 Models of estrogen action at a classical estrogen response element (ERE) and an ER-dependent AP-1 response element. The stars represent the ligand bound to the estrogen receptor (ER). In the classical mode of action activated ERs bind as dimers to the ERE which is comprised of 2 inverted hexanucleotide repeats, and activates gene transcription. In a different mode of action, the ligand activated ER can mediate gene transcription via an AP-1 enhancer element that requires the AP-1 transcription factors Fos and Jun for transcriptional activation. The difference between the actions of the SERMs tamoxifen and raloxifene are due to their ability/inability to activate these pathways. Tamoxifen inhibits the transcription of genes regulated by the classical ERE, but activates the transcription of genes regulated by an AP-1 element. In contrast raloxifene does not have the ability to activate AP-1. Therefore there are numerous combinations by which SERMS can modulate the estrogen receptor in a tissue-specific manner.
Since ERα and ERβ are often co-localized within many tissues, it was assumed that they would exert their effects on different target genes. However, there is evidence that ERα and ERβ can form both homodimers and heterodimers and that ERα homodimers and ERα/ERβ heterodimers are formed in preference to ERβ homodimers (CitationCowley et al 1997). It has been proposed that in a cell where both estrogen receptors are present, the overall estrogen responsiveness may be determined by the ERα:ERβ ratio (CitationStenn and Paus 2001). Furthermore, where ERs are binding to AP-1 sites, binding of ERα or ERβ homodimers can have opposing effects on gene transcription (CitationPaech et al 1997; CitationKushner et al 2000). Thus, this creates the possibility that ERα and ERβ can work antagonistically or synergistically (CitationMatthews and Gustafsson 2003).
Selective estrogen receptor modulators (SERMs)
Clinically, estrogens are widely used in the form of both oral contraceptives and HRT. Although they are highly effective both as a contraceptive and managing menopausal symptoms in women, nevertheless the use of estrogens has also been implicated as a risk factor in breast and uterine cancer (CitationRossouw et al 2002).
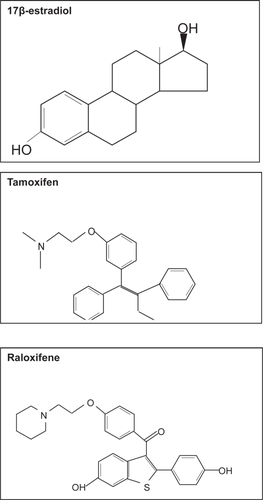
Tamoxifen is a non-steroidal triphenylethylene (), which is widely used in the management of women with hormonally responsive early and metastatic breast cancer, since it acts as an antagonist of estrogen action in breast tissue (CitationCataliotti et al 2006). Although tamoxifen blocks estrogen action in breast cancer cells, conversely it stimulates proliferation of uterine cells by acting as an estrogen agonist, thereby resulting in an increase in the risk of endometrial cancer by three fold (CitationStygar et al 2003).
Figure 4 Molecular structure of 17β-estradiol, tamoxifene and raloxifene.
The study of the selective biological effects of tamoxifen in different tissues has led to the embodiment of the concept of selective estrogen receptor modulators or SERMs. SERMs bind to estrogen receptors and can act as either estrogen agonists or antagonists depending on the target tissue. Current SERMs generally act as agonists in liver, in bone by inhibiting bone resorption, and on the cardiovascular system by reducing the markers of cardiovascular risk such as LDL. They are often antagonists in tissues such as breast and brain, but exhibit a mixed response in the uterus (CitationJordan 1998; CitationDiel 2002). The undesirable uterine side-effects of tamoxifen have provided the drive for the development of the “ideal” SERM.
More recently a new SERM, raloxifene has been introduced (). Raloxifene is derived from a benzothiophene series of anti-estrogens and is now approved for the prevention and treatment of osteoporosis in post-menopausal women (reviewed in CitationNeven and Vergotta 2001). While raloxifene exhibits antiestrogen activity in the breast, in contrast to tamoxifen, it lacks uterotrophic activity and thus signifies an improved agonist/antagonist profile (CitationStygar et al 2003).
Since the three-dimensional structure of tamoxifen and raloxifene differ to 17β-estradiol () binding of these ligands to the estrogen receptor in breast tissue results in a conformational change that blocks AF-2 activity. This results in differences in their ability to recruit essential co-activators, thus eliminating the estrogen effect (CitationSporn 2004). Although tamoxifen inhibits the transcription of genes that are regulated by a classical ERE, tamoxifen activates the AP-1 target gene in uterine cells (CitationKushner 2000) in an AF-independent manner. Raloxifene, however, does not have the ability to activate AP-1 which would account for the differences seen in their effects on the uterus. This gives numerous combinations by which SERMS can modulate estrogen receptors in a tissue-specific manner. Understanding why SERMs exhibit different estrogen activities will help to identity new SERMs with more favourable profiles.
Since hypoestrogenism has detrimental effects on many other tissues including the skeleton, the CNS and the cardiovascular system, many consider the ideal SERM to have estrogenic activity in bone, the cardiovascular system, vagina and CNS, yet have antiestrogenic activity in the breast and uterus. However, since neither tamoxifen nor raloxifene possess this spectrum of properties, there is clearly a market for the development of specific SERMs to address these issues.
SERMs and skin biology
Despite the well documented effects of estrogen on skin physiology and aging, there is very limited data on the effect of SERMs on the skin. One of the adverse effects of tamoxifen include hot flashes and vaginal dryness (reviewed in CitationNeven and Vergotta 2001), but there have been no studies to determine effects on skin thickness, collagen content, elastic fibers or the formation of wrinkles. A histopathological assessment of rat skin following subcutaneous administration of tamoxifen observed that tamoxifen treatment resulted in the appearance of abnormal hair follicles, epidermal atrophy and increased dermal fibrosis, particularly around the hair follicles (CitationInaloz et al 2002). There have been reports of tamoxifen treatment causing diffuse thinning of the hair with moderate receding of the frontal hair line (CitationAyoub et al 1997) and the development of alopecia on the crown, which was reversed when treatment was stopped (CitationGateley and Bundred 1997). Although alopecia is reported on the data and patient information sheet for proprietary tamoxifen, alopecia is not reported in the datasheets for the generic form of tamoxifen (CitationGateley and Bundred 1997).
We have recently reported that tamoxifen alone has no effect on human hair shaft elongation in organ culture, suggesting it is not an estrogen agonist (CitationNelson 2006). However, a 10-fold excess of tamoxifen incubated in combination with 17β-estradiol eliminated the inhibitory effect of 17β-estradiol, suggesting that tamoxifen acts as an antagonist of estrogen in the female scalp hair follicle (CitationNelson 2006).
Notwithstanding, there have been some studies that suggest tamoxifen may be useful in the treatment of keloids. Tamoxifen has been shown to inhibit the proliferation of fibroblasts cultured from keloid biopsies, and to inhibit their contraction (CitationHu et al 1998). Keloid dermal fibroblasts have also been shown to secrete higher levels of TGF-β1 than fetal dermal fibroblasts, which can be counteracted by incubation with tamoxifen (CitationMikulec et al 2001).
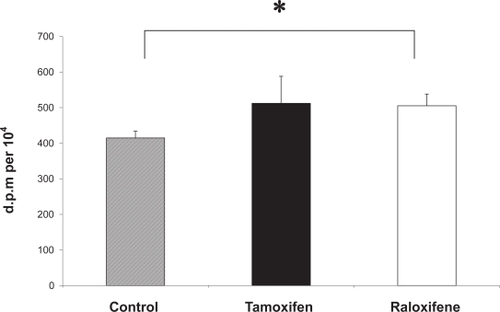
Together these provide evidence that tamoxifen may improve wound healing and improve dermal scarring. Cell migration is a key element of the wound healing process and a recent study has shown that estrogen and tamoxifen induce cytoskeleton remodeling and migration in endometrial cancer cells (CitationAcconcia et al 2006). Another fundamental aspect of cutaneous wound healing is an increase in dermal fibroblast proliferation, and recent studies in our laboratory have shown that monolayers of human dermal fibroblasts that have been mechanically wounded in vitro demonstrate a significantly increased rate of both migration and proliferation in response to 17β-estradiol (CitationStevenson et al 2005). Furthermore, we have recently demonstrated that the proliferative activity of mechanically wounded cultured human dermal fibroblasts is significantly increased in the presence of tamoxifen and raloxifene (), supporting the application of SERMs as potential therapeutic agents to improve wound healing.
Figure 5 Tamoxifen and raloxifene increase the DNA synthesis of mechanically wounded human dermal fibroblasts in vitro. A 3H-thymidine assay was used to measure DNA synthesis in wounded dermal fibroblasts (n = 3) in the presence of 100 nM tamoxifen (black bar) and 100 nM raloxifene (white bar) and vehicle control (0.0001% absolute ethanol (hatched bar). All assays were performed in triplicate dishes. Data are presented as patient mean +/– SEM. * p < 0.05.
Other groups using cultured human dermal fibroblasts have shown that the levels of expression of ERα and ERβ can be modulated by 17β-estradiol, tamoxifen and raloxifene (CitationHaczynski et al 2004). Another study has shown that raloxfine has a stronger positive stimulatory effect on collagen biosynthesis than 17β-estradiol (CitationSurazynski et al 2003) and that in contrast to estradiol, raloxifene inhibits the expression of MMP-9. Tamoxifen has been used in clinical trials for patients with melanoma since the late 1970s. Initially it showed promise as a single agent and then was combined with chemotherapy in 1984 (CitationRusthoven 1998). Since then, a number of phase II clinical trials have combined tamoxifen with different chemotherapeutic agents with overall response rates ranging from 8% to 60%. However, treatment of melanoma with tamoxifen still remains controversial, since it is still unclear whether the strength of evidence from the randomized trials outweighs the combined evidence from numerous nonrandomized trials (CitationRusthoven 1998).
In vitro, tamoxifen has also been shown to inhibit the proliferation of a melanoma cell line (SK-Mel–28) (Lama et al 1999). Cell attachment to plastic and invasion through fibronection in vitro of the highly metastatic cutaneous melanoma cell line (A375-SM) is also reduced in the presence of tamoxifen (CitationDewhurst et al 1997). Others have shown that tamoxifen can induce cell death in malignant melanoma cell lines and this cytoxicity may be due to inactivation of the insulin-like growth factor receptor (IGF-1R) (Kanter-Lewensohn et al 1999). Clearly, malignant melanomas and the derived cell lines may be heterogeneous and the actions of estrogens in these tissues is worthy of further study.
Potential SERMs in development
Recently much emphasis has been placed on developing SERMS, which have tissue specific estrogen actions that can separate the positive biological effects from the undesirable ones. Several SERM trials are underway but are being evaluated in terms of traditional outcomes such as bone mineral density, breast and endometrial cancer, and cardiovascular risk. A number of compounds are currently undergoing Phase III clinical trials (http://www.clinicaltrials.gov). These include lasofoxifene (Pfizer), arzoxifene (Lilly) and bazedoxifene (Wyeth) (see ).
Figure 6 Molecular structure of SERMs in phase III clinical trials.
The safety and effectiveness of lasofoxifene in reducing new spinal fractures in women with osteoporosis is currently being investigated; (PEARL: Postmenopausal Evaluation And Risk-Reduction with Lasofoxifene). This study is expected to enroll 7500 women and the primary outcomes will investigate new radiographic vertebral fractures at 3 years, while the secondary outcomes will compare clinical fractures, non-vertebral fractures, bone mineral density, breast cancer, cardiovascular events, and gynecological safety events at 3 years.
Eli Lilly and Company is sponsoring a study of another SERM, arzoxifene, in a phase III clinical trial to determine its effects on bone mass and the uterus of postmenopausal women. This study will determine the effects of arzoxifene on bone mineral density at the spine and hip as well as the effects on the uterus. Secondary outcomes will include the effects of arzoxifene on blood tests that measure changes in bone rebuilding in postmenopausal women with low bone density; also the effects on blood lipids and other blood markers of heart disease and effects on breast density will be determined.
A third SERM in phase III clinical trial sponsored by Wyeth is bazedoxifene. The purpose of this study is to determine whether bazedoxifene/conjugated estrogens combinations are effective in the treatment of moderate to severe vasomotor symptoms associated with menopause and whether they are effective in the treatment of vaginal atrophy. The effectiveness of bazedoxifene/conjugated estrogens combinations for the prevention of endometrial hyperplasia and osteoporosis in postmenopausal women is also under investigation. However, none of these studies have been designed to assess the effects of these SERMs in the skin and it’s appendages in the women studied.
Conclusion
Evident changes associated with skin aging are thinning of the skin, increased dryness and reduced vascularity. The protective function of the skin can become impaired and aging is also associated with impaired wound healing, hair loss and skin cancer. Clearly the skin is an important estrogen target tissue, yet we still do not fully understand the molecular processes involved and the mechanisms by which estrogens and related compounds regulate skin function and delay skin aging. The widespread and constant expression of estrogen receptors in human skin may provide protection for the skin and its appendages, which are continuously exposed to oxidative damage due to UVB radiation.
However, the mechanisms of estrogen signaling are complex and intricate, involving multiple signaling pathways. There are at least three different receptors that may act independently, synergistically, or have opposing actions. Gene transcription also requires specialized co-activators, which are tissue and cell specific. A class of compounds that demonstrate estrogenic activity in some tissues conversely act as estrogen antagonists in other tissues. This has led to the embodiment of the term selective estrogen receptor modulators (SERMs). With the development of new generation SERMs to prevent osteoporosis and reduce the incidence of cardiovascular disease in postmenopausal women, it is important to understand their effects on the skin and the hair follicle.
Undesirable effects on the skin may lead to chronic wound healing or an increased incidence of skin cancer; in addition women find the adverse occurrence of hair loss very distressing. Conversely, there may be SERMs currently in use or in development that may act as estrogen agonists in human skin and the hair follicle and may potentially provide a useful therapy for clinical application to delay the aging manifestations of the skin. However, research into the potential use of SERMs as a therapy for symptoms of skin aging appears to be very much in its infancy.
In order to understand more comprehensively the mechanism of estrogen action on skin aging, we need to improve our understanding of the complex interactions of estrogen signaling pathways. In addition to SERMs, new advances in the development of selective ligands for ERα and ERβ, and activators of nongenotrophic estrogen-like signaling (ANGELS) may also provide a basis for further interventions in pathological processes that involve the impairment of estrogen action in human skin. This may have important implications for therapies for skin aging, hair growth, skin cancer and wound healing.
Abbreviations
| ERβ: | = | estrogen receptor beta; |
| ERα: | = | estrogen receptor alpha; |
| HSP: | = | heat shock protein; |
| ERE: | = | estrogen response elements; |
| DHEA: | = | dehydroepiandrosterone; |
| 17 β -HSD: | = | 17β-hydroxysteroid dehydrogenase; |
| SERM: | = | selective estrogen receptor modulators; |
| ANGELS: | = | activators of nongenotrophic estrogen-like signaling; |
| HRT: | = | hormone replacement therapy; |
| MMP: | = | matrix metalloproteinase; |
| ECM: | = | extracellular matrix |
Acknowledgements
We are indebted to the following for their help and encouragement relating to various aspects of this article, Professor David Sharpe, Dr Louisa Nelson and Dr Ian Laing.
References
- AcconciaFBarnesCJKumarR2006Estrogen and tamoxifen induce cytoskeletal remodeling and migration in endometrial cancer cellsEndocrinology14712031216339197
- AffinitoPPalombaSSorrentinoC1999Effects of postmenopausal hypoestrogenism on skin collagenMaturitas332394710656502
- AronicaSMKrausWLKatzenellenbogenBS1994Estrogen action via the cAMP signaling pathway: stimulation of adenylate cyclase and cAMP-regulated gene transcriptionProc Natl Acad Sci USA918517218078914
- AroraSVevesACaballaroAE1998Estrogen improves endothelial functionJ Vasc Surg2711416 discussion 1147.9652476
- AshcroftGSAshworthJJ2003Potential role of estrogens in wound healingAm. J. Clin. Dermatol47374314572296
- AshcroftGSHoranMAFergusonMWJ1995The effects of aging on cutaneous wound-healing in mammalsJ Anat1871267591970
- AshcroftGSGreenwell-WildTHoranMA1999Topical estrogen accelerates cutaneous wound healing in aged humans associated with an altered inflammatory responseAm J Pathol15511374610514397
- AshcroftGSHoranMAHerrickSE1997Age-related differences in the temporal and spatial regulation of matrix metalloproteinases (MMPs) in normal skin and acute cutaneous wounds of healthy humansCell Tissue Res290581919369533
- AyoubJ-P MValeroVHortobagyiGN1997Tamoxifen–induced female androgenetic alopecia in a patient with breast cancerAnnals of Internal Medicine12674569139575
- AzziLEl-AlfyMMartelC2005Gender differences in mouse skin morphology and specific effects of sex steroids and dehydroepiandrosteroneJ Invest Dermatol12422715654949
- BaumannL2005A dermatologist’s opinion on hormone therapy and skin agingFertil Steril8428990 discussion 295.16084863
- BeasFVargasLSpadaRP1969Pseudoprecocious puberty in infants caused by a dermal ointment containing estrogensJ Pediatr75127305790395
- BentenWPMStephanCLieberherrM2001Estradiol signalling via sequestrable surface receptorsEndocrinol142166977
- BidmonHJPittsJDSolomonHF1990Estradiol distribution and penetration in rat skin after topical application, studied by high resolution autoradiographyHistochemistry9543542286532
- BoydASMorrisLFPhillipsCM1996Psoriasis and pregnancy: hormone and immune system interactionInt J Dermatol35169728655230
- BrincatMVersiEMonizCF1987Skin collagen changes in post-menopausal women receiving different regimens of estrogen therapyObstet Gynecol7012373601260
- BrincatMMonizCJStuddJW1985Long-term effects of the menopause and sex hormones on skin thicknessBr J Obstet Gynaecol9225693978054
- BrincatMP2000Hormone replacement therapy and the skinMaturitas351071710924836
- BrincatMPBaronYMGaleaR2005Estrogens and the skinClimacteric81102316096167
- BuzdarJDoumaNDavidsonR2001Brady Phase III, multicenter, double-blind, randomized study of letrozole, an aromatase inhibitor, for advanced breast cancer versus megestrol acetateJournal of Clinical Oncology1933576611454883
- CallensAVaillantLLecomteP1996Does hormonal skin aging exist? A study of the influence of different hormone therapy regimens on the skin of postmenopausal women using non-invasive measurement techniquesDermatology193289948993951
- Castelo–BrancoCDuranMGonzalez-MerloJ1992Skin collagen changes related to age and hormone replacement therapyMaturitas15113191345134
- Castelo-BrancoCFiguerasFMartinez de OsabaMJ1998Facial wrinkling in postmenopausal women. Effects of smoking status and hormone replacement therapyMaturitas2975869643520
- CataliottiLBuzdarAUNoguchiS2006Comparison of anastrozole versus tamoxifen as preoperative therapy in postmenopausal women with hormone receptor-positive breast cancer: the Pre-Operative “Arimidex” Compared to Tamoxifen (PROACT) trialCancer106209510316598749
- CliftonVLCromptonRReadMA2005Microvascular effects of corticotropin-releasing hormone in human skin vary in relation to estrogen concentration during the menstrual cycleJ Endocrinol186697616002537
- ConradFOhnemusUBodoE2004Estrogens and human scalp hair growth-still more questions than answersJ Invest Dermatol122840215086574
- ConradFPausR2004Estrogens and the hair follicleJ German Soc Dermatol241223
- CowleySMHoareSMosselmanS1997Estrogen receptors alpha and beta form heterodimers on DNAJ Biol Chem27219858629242648
- CreidiPFaivreBAgacheP1994Effect of a conjugated oestrogen (Premarin) cream on ageing facial skin. A comparative study with a placebo creamMaturitas19211237799828
- DewhurstLOGeeJWRennieIG1997Tamoxifen, 17beta-oestradiol and the calmodulin antagonist J8 inhibit human melanoma cell invasion through fibronectinBr J Cancer7586089062408
- DielP2002Tissue-specific estrogenic response and molecular mechanismsToxicol Lett1272172412052661
- DraelosZD2005Topical and oral estrogens revisited for antiaging purposesFertil Steril842912 discussion 295.16084864
- DunnLBDamesynMMooreAA1997Does estrogen prevent skin aging? Results from the First National Health and Nutrition Examination Survey (NHANES I)Arch Dermatol133339429080894
- DunnaSFFinlayAY1989Psoriasis: improvement during and worsening after pregnancyBr J Dermatol1205842730848
- DurvasulaRAhmedSMVashishtA2002Hormone replacement therapy and malignant melanoma: to prescribe or not to prescribe?Climacteric519720012051116
- EisenbeissCWelzelJSchmellerW1998The influence of female sex hormones on skin thickness: evaluation using 20 MHz sonographyBr J Dermatol13946279767291
- ElgortMGZouAMarschkeKB1996Estrogen and estrogen receptor antagonists stimulate transcription from the human retinoic acid receptor-alpha 1 promoter via a novel sequenceMol Endocrinol10477878732679
- EnmarkEPelto-HuikkoMGrandienK1997Human estrogen receptor beta-gene structure, chromosomal localization, and expression patternJ Clin Endocrinol Metab824258659398750
- GateleyCABundredNJ1997Alopecia and breast diseaseBritish Medical Journal3144819056798
- GiguereVTremblayATremblayGB1998Estrogen receptor beta: re-evaluation of estrogen and antiestrogen signallingSteroids6333599618797
- GradyDHerringtonDBittnerV2002Cardiovascular disease outcomes during 6.8 years of hormone therapy: Heart and Estrogen/progestin Replacement Study follow-up (HERS II)JAMA288495712090862
- GustafssonJA1999Estrogen receptor beta – a new dimension in estrogen mechanism of actionJ Endocrinol1633798310588810
- HaapasaariKMRaudaskoskiTKallioinenM1997Systemic therapy with estrogen or estrogen with progestin has no effect on skin collagen in postmenopausal womenMaturitas27153629255750
- HaczynskiJTarkowskiRJarzabekK2002Human cultured skin fibroblasts express estrogen receptor alpha and betaInt J Mol Med101495312119550
- HaczynskiJTarkowskiRJarzabekK2004Differential effects of estradiol, raloxifene and tamoxifen on estrogen receptor expression in cultured human skin fibroblastsInt J Mol Med13903815138633
- HallGKPhillipsTJ2004Skin and hormone therapyClin Obstet Gynecol474374915166870
- HasselquistMBGoldbergNSchroeterA1980Isolation and characterization of the estrogen receptor in human skinJ Clin Endocrinol Metab5076827350189
- HennemanDH1968Effect of estrogen on in vivo and in vitro collagen biosynthesis and maturation in old and young female guinea pigsEndocrinology83678905685958
- HenryFPierard–FranchimontCCauwenberghG1997Age-related changes in facial skin contours and rheologyJ Am Geriatr Soc4522029033524
- HolzerGRieglerEHonigsmannH2005Effects and side-effects of 2% progesterone cream on the skin of peri- and postmenopausal women: results from a double-blind, vehicle-controlled, randomized studyBr J Dermatol1536263416120154
- HosokawaMIshiiMInoueK1981Estrogen induces different responses in dermal and lung fibroblasts: special reference to collagenConnect Tissue Res9115206458448
- HuDHughesMACherryGW1998Topical tamoxifen – a potential therapeutic regime in treating excessive dermal scarring?Br J Plast Surg5146299849367
- InalozHSDeveciEInalozSS2002The effects of tamoxifen on rat skinEur J Gynaecol Oncol2350211876393
- JordanVC1998Designer estrogensSci Am2796079768522
- KandaNWatanabeS2003a17beta-estradiol inhibits oxidative stress-induced apoptosis in keratinocytes by promoting Bcl-2 expressionJ Invest Dermatol1211500914675202
- KandaNWatanabeS2003b17beta-estradiol inhibits the production of interferon-induced protein of 10 kDa by human keratinocytesJ Invest Dermatol1204111912603854
- KandaNWatanabeS200417beta-estradiol stimulates the growth of human keratinocytes by inducing cyclin D2 expressionJ Invest Dermatol1233192815245432
- Kanter-LewensohnLGirnitaLGirnitaA2000Tamoxifen-induced cell death in malignant melanoma cells: possible involvement of the insulin-like growth factor-1 (IGF-1) pathwayMol Cell Endocrinol165131710940491
- KlingeCM2001Estrogen receptor interaction with estrogen response elementsNucleic Acids Res2929051911452016
- KousteniS2002Reversal of bone loss in mice by nongenotropic signaling of sex steroidsScience298843612399595
- KousteniSHanLChenJR2003Kinase-mediated regulation of common transcription factors accounts for the bone-protective effects of sex steroidsJ Clin Invest11116516412782668
- KroumpouzosGCohenLM2001Dermatoses of pregnancyJ Am Acad Dermatol4511911423829
- KuiperGGCarlssonBGrandienK1997Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and betaEndocrinology138863709048584
- KushnerPJAgardDAGreeneGL2000Estrogen receptor pathways to AP-1Steroid Biochem Mol Biol7431117
- LabrieF2004Adrenal androgens and intracrinologySemin Reprod Med2229930915635498
- LamaGAngelucciCBruzzeseN1998Sensitivity of human melanoma cells to oestrogens, tamoxifen and quercetin: is there any relationship with type I and II oestrogen binding site expression?Melanoma Res8313229764806
- LevinER2002Cellular functions of plasma membrane estrogen receptorsSteroids67471511960623
- LieberherrMGrosseBKachkacheM1993Cell signaling and estrogens in female rat osteoblasts: a possible involvement of unconventional nonnuclear receptorsJ Bone Mineral Res8136576
- LynfieldYL1960Effect of pregnancy on the human hair cycleJ Invest Dermatol35323713764567
- MaheuxRNaudFRiouxM1994A randomized, double-blind, placebo-controlled study on the effect of conjugated estrogens on skin thicknessAm J Obstet Gynecol17064298116726
- MarinoMAcconciaFBrescianiF2002Distinct nongenomic signal transduction pathways controlled by 17beta-estradiol regulate DNA synthesis and cyclin D(1) gene transcription in HepG2 cellsMol Biol Cell133720912388769
- MatthewsJGustafssonJA2003Estrogen signaling: a subtle balance between ER alpha and ER betaMol Interv32819214993442
- MikulecAAHanasonoMMLumJ2001Effect of tamoxifen on transforming growth factor beta1 production by keloid and fetal fibroblastsArch Facial Plast Surg31111411368663
- MillerJGMac NeilS1997Gender and cutaneous melanomaBr J Dermatol136657659205495
- MoalliPATalaricoLCSungVW2004Impact of menopause on collagen subtypes in the arcus tendineous fasciae pelvisAm J Obstet Gynecol190620715041990
- MoverareSLindbergMKFaergemannJ2002Estrogen receptor alpha, but not estrogen receptor beta, is involved in the regulation of the hair follicle cycling as well as the thickness of epidermis in male miceJ Invest Dermatol1191053812445192
- NadalARoperoABFuentesE2001The plasma membrane estrogen receptor: nuclear or unclear?Trends in Pharm Sci22597911730951
- NaftolinF2005Prevention during the menopause is critical for good health: skin studies support protracted hormone therapyFertil Steril842934 discussion 295.16084865
- NeifeldJP1996Endocrinology of melanomaSemin Surg Oncol1240268914204
- NelsonLMessengerDKarooAG200317beta-eatradiol but not 17alpha-estradiol inhibits human hair growth in whole follicle organ cultureJ. Invest. Dermatol121821a14632201
- NelsonLD2006The role of oestrogen in skinPhD ThesisBradford, UKSchool of Life Sciences, University of Bradford
- NelsonLRBulunSE2001Estrogen production and actionJ Am Acad Dermatol45S1162411511861
- NevenPVergoteI2001Tamoxifen, screening and new oestrogen receptor modulatorsBest Pract Res Clin Obstet Gynaecol153658011476559
- t et al. 2006The hair follicle as an estrogen target and sourceEndocr Rev728 [Epub ahead of print].
- PaechKWebbPKuiperGG1997Differential ligand activation of estrogen receptors ERalpha and ERbeta at AP1 sitesScience2771508109278514
- PappasTCGametchuBWatsonCS1995Membrane estrogen receptors identified by multiple antibody labelling and impeded-ligand bindingFASEB J9404107896011
- ParkerCRMixonRLBrissieRM1997Ageing alters zonation in the adrenal cortex of menJ Clin Endocrinol Metab8238989019360559
- PatriarcaMTGoldmanKZDos SantosJM2007Effects of topical estradiol on the facial skin collagen of postmenopausal women under oral hormone therapy: a pilot studyEur J Obstet Gynecol Reprod Biol130202516797821
- PayneAHHalesDB2004Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormonesEndor Rev2594770
- PhillipsTJDemircayZSahuM2001Hormonal effects on skin agingClin. Geriatr Med176617211535422
- Pierard-FranchimontCLetaweCGoffinV1995Skin water-holding capacity and transdermal estrogen therapy for menopause: a pilot studyMaturitas2215148538484
- Pierard-FranchimontCCornilFDehavayJ1999Climacteric skin ageing of the face – a prospective longitudinal comparative trial on the effect of oral hormone replacement therapyMaturitas32879310465376
- PunnonenR1972Effect of castration and peroral estrogen therapy on the skinActa Obstet Gynecol Scand Suppl213444511789
- PunnonenRLovgrenTKouvonenI1980Demonstration of estrogen receptors in the skinJ Endocrinol Invest321721
- PunnonenRVaajalahtiPTeisalaK1987Local oestriol treatment improves the structure of elastic fibers in the skin of postmenopausal womenAnn Chir Gynaecol Suppl20239413477986
- RaychaudhuriSPNavareTGrossJ2003Clinical course of psoriasis during pregnancyInt J Dermatol425182012839599
- RazandiMPedramAGreeneGl1999Cell membrane and nuclear estrogen receptors (ERs) originate from a single transcript: studies of ERalpha and ERbeta expressed in Chinese hamster ovary cellsMol Endocrinol13307199973260
- RossouwJEAndersonGLPrenticeRL2002Risks and benefits of estrogen plus progestin in healthy postmenopausal women: principal results From the Women’s Health Initiative randomized controlled trialJAMA2883213312117397
- RusselKSHaynesMPSinhaD2000Human vascular endothelial cells contain membrane binding sites for estradiol, which mediate rapid intracellular signallingProc Natl Acad Sci USA975930510823945
- RusthovenJJ1998The evidence for tamoxifen and chemotherapy as treatment for metastatic melanomaEur J Cancer34Suppl 3S3169849407
- SatorPGSchmidtJBRabeT2004Skin aging and sex hormones in women – clinical perspectives for intervention by hormone replacement therapyExp Dermatol13Suppl 4364015507111
- SatorPGSchmidtJBSatorMO2001The influence of hormone replacement therapy on skin ageing: a pilot studyMaturitas39435511451620
- SauerbronnAVFonsecaAMBagnoliVR2000The effects of systemic hormonal replacement therapy on the skin of postmenopausal womenInt J Gynaecol Obstet68354110687834
- SavvasMBishopJLaurentG1993Type III collagen content in the skin of postmenopausal women receiving oestradiol and testosterone implantsBr J Obstet Gynaecol10015468476808
- SchmidtJBBinderMDemschikG1996Treatment of skin aging with topical estrogensInt J Dermatol35669748876303
- ShahMGMaibachHI2001Estrogen and skin: an overviewAm J Clin Dermatol21435011705091
- ShaulPW1999Rapid activation of endothelial nitric oxide synthase by estrogenSteroids64283410323670
- SimpsonER1998Genetic mutations resulting in estrogen insufficiency in the maleMol Cell Endocrinol1455599922099
- SimpsonDCurranMPPerryCM2004Letrozole: a review of its use in postmenopausal women with breast cancerDrugs6412133015161328
- SinclairR1999Hair structure and function In Sinclair R, Banfiel C, Dawber R Handbook of diseases of the hair and scalpNew YorkBlackwell, Oxford Press
- SmithQTAllisonDJ1966Studies on the uterus, skin and femur of rats treated with 17-beta-oestradiol benzoate for one to twenty-one daysActa Endocrinol (Copenh)535986106012539
- SmithCLO’MalleyBW2004Coregulator functions: a key to understanding tissue specificity of selective receptor modulatorsEndocr Rev25457114769827
- SobelHLeeKDHewlettMJ1965Effect of estrogen on acid glycosaminoglycans in skin of miceBiochem Biophys Acta10122595852522
- SonEDLeeJYLeeS2005Topical application of 17beta-estradiol increases extracellular matrix protein synthesis by stimulating tgf-Beta signaling in aged human skin in vivoJ Invest Dermatol12411496115955089
- SpornMBDowsettSAMershonJ2004Role of raloxifene in breast cancer prevebtion: clinical evidence and potential mechanisms of actionClin Ther268304015262454
- StampferMJColditzGAWillettWC1991Postmenopausal estrogen therapy and cardiovascular disease. Ten-year follow-up from the nurses‘ health studyN Engl J Med325756621870648
- StennKSPausR2001Controls of hair follicle cyclingPhysiol Rev814499411152763
- StevensonSNelsonLDHuqS2005Oestrogens and wound healing: migration, proliferation and secretion of paracrine factors by human dermal fibroblasts in vitroSoc for Endocrinol Annual Meeting. URL: http://www.endocrine-abstracts.org10P80
- StumpfWESarMJoshiSG1974Estrogen target cells in the skinExperientia3019684360727
- StygarDMuravitskayaNErikssonB2003Effects of SERM (selective estrogen receptor modulator) treatment on growth and proliferation in the rat uterusReprod Biol Endocrinol14012777179
- SuminoHIchikawaSAbeM2004Effects of aging and postmenopausal hypoestrogenism on skin elasticity and bone mineral density in Japanese womenEndocr J511596415118265
- SurazynskiAJarzabekKHaczynskJ2003Differential effects of estradiol and raloxifene on collagen biosynthesis in cultured human skin fibroblastsInt J Mol Med12803914533013
- ThorntonMJ2002The biological actions of estrogens on skinExp Dermatol1148750212473056
- ThorntonMJ2005Oestrogen functions in skin and skin appendagesExpert Opin Ther Targets96172915948678
- ThorntonMJTaylorAHMulliganK2003Estrogen receptor beta (ERβ) is the predominant estrogen receptor in human scalpExp Dermatol121819012702147
- TsukaharaKMoriwakiSOhuchiA2001Ovariectomy accelerates photoaging of rat skinPhotochem Photobiol735253111367575
- TsukaharaKNakagawaHMoriwakiS2004Ovariectomy is sufficient to accelerate spontaneous skin ageing and to stimulate ultraviolet irradiation-induced photoageing of murine skinBr J Dermatol1519849415541076
- UranoRSakabeKSeikiK1995Female sex hormone stimulates cultured human keratinocyte proliferation and its RNA- and protein-synthetic activitiesJ Dermatol Sci9176848664215
- VarilaERantalaIOikarinenA1995The effect of topical oestradiol on skin collagen of postmenopausal womenBr J Obstet Gynaecol10298598652490
- Verdier-SevrainSBonteFGilchrestB2006Biology of estrogens in skin: implications for skin agingExp Dermatol15839416433679
- Verdier-SevrainSYaarMCantatoreJ2004Estradiol induces proliferation of keratinocytes via a receptor mediated mechanismFaseb J181252415208259
- WadeTRWadeSLJonesHE1978Skin changes and diseases associated with pregnancyObstet Gynecol5223342683665
- WebbPNguyenPValentineC1999The estrogen receptor enhances AP-1 activity by two distinct mechanisms with different requirements for receptor transactivation functionsMol Endocrinol1316728510517669
- WebbPNguyenPKushnerPJ2003Differential SERM effects on corepressor binding dictate ERα activity in vivoJ Biol Chem27869122012482846
- WeinstockMA1994Epidemiologic investigation of nonmelanoma skin cancer mortality: the Rhode Island Follow-Back StudyJ Invest Dermatol1026S9S8006441
- WolffEFNarayanDTaylorHS2005Long-term effects of hormone therapy on skin rigidity and wrinklesFertil Steril84285816084862