Abstract
There is growing evidence that accelerated telomeric attrition and/or aberrant telomerase activity contributes to pathogenesis in a number of diseases. Likewise, there is increasing interest to develop new therapies to restore or replace dysfunctional cells characterized by short telomeric length using telomerase-positive counterparts or stem cells. While telomerase adds telomeric repeats de novo contributing to enhanced proliferative capacity and lifespan, it may also increase cellular survival by conferring resistance to apoptosis. Consequently, we sought to determine the involvement of telomerase for reduced apoptosis using ovarian surface epithelial cells. We found that expression of hTERT, the catalytic component of telomerase, was sufficient and specific to reduce caspase-mediated cellular apoptosis. Further, hTERT expression reduced activation of caspases 3, 8, and 9, reduced expression of pro-apoptotic mitochondrial proteins t-BID, BAD, and BAX and increased expression of the anti-apoptotic mitochondrial protein, Bcl-2. The ability of telomerase to suppress caspase-mediated apoptosis was p-jnk dependent since abrogation of jnk expression with jip abolished resistance to apoptosis. Consequently, these findings indicate that telomerase may promote cellular survival in epithelial cells by suppressing jnk-dependent caspase-mediated apoptosis.
Introduction
Telomeric attrition is the prevailing model of a molecular clock (CitationHarley et al 1992) proposed to explain limited cellular proliferation known as the Hayflick limit (CitationHayflick 1965). Telomeres, the linear ends of eukaryotic chromosomes, consist of specialized tandem (TTAGGG)n DNA repeats and associated telomeric binding proteins that serve to protect the ends of chromosomes from recombination, end-to-end fusion and degradation, thereby maintaining chromosomal integrity (CitationBlackburn 1991). While telomeres may also be involved in nuclear organization because they attach to the nuclear matrix (CitationdeLange 1992), they are crucial in solving the ‘end replication’ problem (CitationOlovnikov 1975; CitationHarley et al 1992). That is, in normal somatic cells there is progressive telomeric loss with successive rounds of cellular replication due to the inability of conventional DNA polymerase to fully elongate linear DNA. Telomeric shortening continues until replicative senescence or cellular crisis when critically shortened telomeres no longer effectively stabilize chromosomes and most cells die. Greatest telomeric shortening has been reported in cells from patients with progeroid syndromes (CitationKruk et al 1995; CitationMetcalfe et al 1996) and in epithelial cells (CitationCounter et al 1992). However, the rates and extents of telomeric loss in vivo (7–150 base pairs/year) and in vitro (50–200 base pairs/population doubling) are highly variable among different cell types (CitationHastie et al 1990; CitationAllsopp et al 1995; CitationKruk et al 1995; CitationMetcalfe et al 1996), so that not all cells reach senescent crisis equally.
Occasional immortal cells emerging from crisis usually contain shortened, but stable telomeres due to the re-expression of telomerase, a ribonucleoprotein that elongates telomeric repeats de novo (CitationKim et al 1994). Consequently, telomerase suppresses cellular senescence and extends cellular lifespan (CitationMeyerson et al 1997). Telomerase activity is generally limited to stem cell populations and immortalized tumor cells and is suppressed in normal cells (CitationKim et al 1994). Telomerase consists of an RNA component (hTR) that serves as an internal telomeric template and hTERT, a reverse transcriptase (CitationKim et al 1994). While both protein catalytic hTERT and hTR subunits have been identified and cloned, the protein catalytic subunit is the limiting determinant of telomerase activity (CitationFeng et al 1995; CitationMeyerson et al 1997; CitationWeinrich et al 1997; CitationCounter et al 1998). In addition to these two core components, several telomerase-associated proteins have been identified. They include the hTR RNA binding proteins TEP1, hnRNP proteins (A1, C1/C2, D), survival of motor neuron (SMN), L22, and hStau, while p23 and hsp90 bind hTERT, suggesting a role for these proteins in hTR RNA processing, telomerase assembly, and cellular localization of the telomerase complex (CitationNakayama et al 1997; CitationHolt, Aisner, et al 1999; CitationEversole and Maizels 2000; CitationFord et al 2000, Citation2001; CitationLe et al 2000; CitationFiset and Chabot 2001; CitationKamma et al 2001; CitationBachand et al 2002). While telomerase regulation is complex (CitationUlaner et al 1998; CitationCong et al 1999; CitationGreenberg et al 1999; CitationTakakura et al 1999), we have previously developed a culture model system in which telomerase activity can be perturbed and controlled in ovarian surface epithelial (OSE) cell lines by endogenous and exogenous stresses (CitationAlfonso-De Matte et al 2001; CitationAlfonso-De Matte, Moses-Soto, et al 2002; CitationAlfonso-De Matte, Yang, et al 2002; CitationAlfonso-De Matte and Kruk 2004).
Although the primary function of telomerase is the maintenance of structural integrity at the linear ends of chromosomes contributing to cellular immortalization, recent studies have shown an association between telomerase activity and increased chemotherapeutic resistance consistent with poor prognosis in many cancers (CitationAsai et al 1998; CitationKiyozuka et al 2000; CitationVilla et al 2000). Telomerase appears to mediate its protective effect by conferring resistance to apoptosis (CitationMandal and Kumar 1997; CitationKondo et al 1998; CitationFu et al 1999, Citation2000; CitationTian et al 1999; CitationZhang et al 1999; CitationHerbert et al 1999; CitationHolt, Glinsky, et al 1999; CitationIida et al 2000; CitationZhu et al 2000), suggesting that the function of telomerase is more complex than initially thought. However, the exact mechanism(s) by which telomerase confers resistance to apoptosis leading to enhanced cellular survival remains unclear.
Accelerated telomeric erosion and/or aberrant telomerase activity have been implicated in the pathogenesis of many diseases including: progeroid disorders (CitationKruk et al 1995; CitationMetcalfe et al 1996; CitationDu et al 2004); cardiovascular disease (CitationOh et al 2001; CitationSerrano and Andres 2004); neurological disorders (CitationPanossian et al 2003; CitationZhang et al 2003); and acquired anemia (CitationMitchell et al 1999; CitationVulliamy et al 2001; CitationGreenwood and Lansdorp 2003; CitationOhshima et al 2003; CitationPolychronopoulou and Koutroumba 2004). Given the growing interest in the potential for progenitor cell therapy to restore or repair dysfunctional cells by using telomerase-positive/stem cells to enhance cell survival, we sought to determine whether telomerase-mediated suppression of apoptosis contributes to increased cellular survival in epithelial cells.
Materials and methods
Cell culture and transfections
Normal ovarian surface epithelial cell lines (IOSE-80, FHIOSE 1816-686, IMCC3, IMCC5) were used as previously described (CitationKruk et al 1990, Citation1999). All cell lines were previously determined to be telomerase-negative (CitationKruk et al 1999). Cells were maintained in Medium 199/MCDB 105 (1:1; Sigma Scientific, St Louis, MO, USA) supplemented with 10 μg gentamicin (GIBCO BRL, Grand Island, NY, USA) and 5%–10% fetal bovine serum (FBS; Hyclone, Logan UT, USA) in a humidified 5% CO2/95% air atmosphere. Cisplatin (CP), staurosporine (STS), and tumor necrosis factor-α (TNF-α) were employed to induce apoptosis and were obtained from Sigma Scientific.
Cells were transfected with plasmid constructs of wild-type (wt) hTERT, dominant-negative (DN) hTERT, jip, or jnk using the previously described lipofectamine transfection protocol (CitationAlfonso-De Matte, Yang, et al 2002). Transfections were confirmed via Western blot analysis. Transfection efficiency was measured in parallel cultures transfected with green fluorescent protein (GFP) cDNA under identical conditions.
Telomerase assay
To quantitatively detect changes in telomerase, cells were assayed for telomerase activity using the telomerase polymerase chain reaction-enzyme-linked immunosorbent assay (PCR-ELISA; Roche Molecular Biochemicals, Indianapolis, IN, USA) as described previously (CitationAlfonso-De Matte et al 2001). Briefly, cells were washed with Dulbecco’s phosphate buffered saline (DPBS), trypsinized and centrifuged at 500 g for 5 min. Pellets were washed twice in DPBS, then resuspended in 200 μl of CHAPS lysis buffer and kept on ice for 30 min, after which time the lysates were centrifuged at 100 000 g for 60 min at 4°C. Lysates were assayed using the Bio-Rad detergent corrected (DC) Protein Assay (Bio-Rad, Hercules, CA, USA) for determination of protein concentration following detergent solubilization. In order to perform the telomerase PCR-ELISA assay within the linear range, cell extracts equivalent to 2 μg of protein were used. Following PCR-ELISA, telomerase activity was detected using a Dynex-MRX plate reader (Dynex Technologies, Chantilly, VA, USA) and recorded as absorbance units. The values were expressed as relative absorbance at 450 nm ± SE.
Telomeric length determination
Telomeric lengths were determined by Southern blot analysis as described previously (CitationKruk et al 1995). Briefly, cells were washed with phosphate buffered saline (PBS), lysed in 10 mM TrisHCl/1mM EDTA/0.5% SDS/0.1 mg of proteinase K per ml (Sigma Scientific) at 37°C. DNA was isolated by standard high salt extraction, treated with 100 μg RNase A per ml, and resuspended in 10 mM TrisHCl/1 mM EDTA, pH 7.2. The DNA concentration was determined by the absorbance at 260 nm. The DNA was digested to completion with HinfI (1 unit per μg DNA; New England Biolabs, Beverly, MA, USA). DNA (1–2 μg) was loaded onto 0.5% alkaline agarose gels, resolved by electrophoresis, transferred to nylon membranes in 1 N NaOH, and hybridized in Church’s hybridization buffer at 42°C with (γ-32P) adenosine triphosphate (ATP) (Dupont/NEN, Wilmington, DE, USA) end-labeled (TTAGGG)4 (Midland Certified Reagent Company, Midland, TX, USA). Membranes were washed in 0.5X saline–sodium citrate (SSC)/0.1% Sodium dodecyl sulfate (SDS) at 42°C, and the telomeres visualized by autoradiography. Mean telomeric lengths were calculated using ImageQuant software (Molecular Dynamics, Sunnyvale, CA, USA) based on center of mass calculations and expressed in kb ± SE. Mean telomeric lengths were determined from at least two autoradiograms from each of three biological experiments.
Cell growth
Cell growth was determined by the MTS colorimetric assay (Promega, Madison, WI, USA). The assay was performed in 96 well microtiter plates as described previously (CitationJohnson and Kruk 2002) and is based on soluble formazan production by dehydrogenase enzymes found in metabolically active cells. Cells were plated at 2.5 × 103 cells per well. Absorbance was determined at 490 nm using a Dynex MRX plate reader and the results expressed as the mean absorbance ± SE.
DNA ladders
Cellular apoptosis, as performed previously (CitationJohnson et al 2004), was examined via DNA laddering by electrophoresis of 1 μg of high salt-extracted DNA on a 2% agarose gels followed by SYBR green I staining (FMC Bioproducts, Rockland, ME, USA).
SDS-PAGE and Western blot analysis
As described previously, Western blot analysis was performed to observe changes in members of the apoptotic cascade associated with telomerase expression (CitationAlfonso-De Matte, Moses-Soto, et al 2002; CitationJohnson and Kruk 2002; CitationJohnson et al 2004). Cells were trypsinized, pelleted at 500 g for 5 min, lysed in ice cold CHAPS lysis buffer and 15 μg protein were added to 4X loading buffer (250 mM Tris pH 6.8, 8% SDS, 20% glycerol, 0.012% bromophenol blue, 4% β-mercaptoethanol), electrophoresed in 12.5 % SDS-polyacrylamide gels, transferred to nitrocellulose membranes (Amersham Pharmacia Biotech, Piscataway, NJ, USA) via semi-dry or wet transfer. All membranes were blocked for 1 hour with 5% non-fat milk Tris Buffered Saline plus 0.1% Tween-20 (T-TBS) and incubated overnight at 4°C in primary antibody. Membranes were incubated and developed according to the Enhanced Chemiluminescent Protocol (Amersham Pharmacia Biotech). After initial blotting, membranes were reprobed for actin to ensure even loading. All antibodies were purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA).
Statistical analysis
Samples for telomerase PCR-ELISA, MTL, and growth assays were run in triplicate and the data subjected to Student t test analysis for determination of statistical significance between treated and untreated samples.
Results
Telomerase increases mean telomeric length and cell growth
In agreement with others (CitationKim et al 1994; CitationMeyerson et al 1997; CitationWeinrich et al 1997; CitationCounter et al 1998; CitationLuiten et al 2003), we found that transfection with hTERT cDNA was sufficient to activate telomerase expression in ovarian surface epithelial cells (). Likewise, long-term expression of hTERT resulted in increased mean telomeric lengths to levels more than double their telomerase-negative counterparts () and indicative of functional telomerase activity with hTERT transfection. In addition, cells transfected with hTERT demonstrated increased growth capacity (). That is, cell yield in hTERT-positive cells was found to be three times greater than their telomerase-negative counterparts in short term culture assays.
Telomerase enhances cell survival
To determine whether telomerase conferred increased cell survival, IOSE cells ± hTERT cDNA were treated with 25μM cisplatin (CP) for 2 hours and assayed for cell growth. Cisplatin suppressed cell growth in parental cell lines 50%–70% of untreated controls (). In contrast, CP was only able to suppress cell growth in hTERT-positive cells by approximately 30% (). While increased growth rates in telomerase-positive cells may contribute to reduced cytotoxicity, parallel cultures were examined for DNA laddering to determine whether reduced apoptosis contributed to reduced cytotoxicity in telomerase-positive cells. In agreement with the cytotoxicity reported above, IOSE cells demonstrated significant DNA laddering and fragmentation by 48 hours following treatment with CP (). In contrast, telomerase-positive cells showed only marginal DNA laddering by 72 hours following CP, indicative of reduced apoptosis in telomerase-positive cells.
Telomerase reduces caspase 3-mediated apoptosis
To determine whether reduced apoptosis in telomerase-positive cells was caspase 3-dependent, we treated telomerase-positive and negative IOSE cells with 25 μM CP for 2 hours, 20J/m2 ultraviolet (UV) or 1 μM STS for 4 hours and examined cell lysates for levels of activated (cleaved) caspase 3 (). All three agents were able to induce significant activation of caspase 3 cleavage in telomerase-negative cells. However, we noted reduced caspase 3 cleavage by all three apoptosis-inducing agents in telomerase-positive cells. In addition, some samples were examined for DFF45, a known cleavage target of caspase 3 to ensure caspase 3 activity (). We found increased cleavage of DFF45 in telomerase-negative cells compared with telomerase-positive cells. To ensure that telomerase was sufficient and specific to reduce caspase 3-mediated apoptosis, we co-transfected telomerase-positive cells with a dominant negative (DN) hTERT cDNA to determine whether inhibition of telomerase restored caspase 3 activation. As expected, DN hTERT suppressed telomerase activity (), but levels of telomerase were unaffected by STS. When these cultures were examined for caspase 3 activation, DN hTERT was able to restore STS-induced caspase 3 activation to levels found in control cells while hTERT alone suppressed caspase 3 activation ().
Telomerase reduces caspase and mitochondrial apoptotic activation
To identify additional targets of telomerase-mediated anti-apoptosis, IOSE cells ± hTERT were treated with 1 ng/ml TNF-α, which induces apoptosis by both the intrinsic and extrinsic apoptotic pathways. TNF-α did not significantly alter telomerase activity in telomerase-positive cells (). However, activation of caspases 3, 8, and 9 in telomerase-positive cells was reduced by up to 50% compared with their telomerase-negative counterparts following treatment with TNF-α (–).
Likewise, telomerase altered pro- and anti-apoptotic mitochondrial protein expression. Densitometric analysis revealed that treatment with TNF-α resulted in up to an 84% reduction in Bcl-2 levels in telomerase-negative cells, but up to a 172% increase in Bcl-2 expression in telomerase-positive cells (), suggesting an anti-apoptotic tendency in the latter cells. When we examined the samples for levels of pro-apoptotic proteins, we found that activation of tBID (by cleavage of BID) was more pronounced in telomerase-negative cells (). Further, when the levels of t-BID to Bcl-2 were compared, the ratio of t-BID/Bcl-2 was more than 7 times greater in telomerase-negative cells (). Likewise, while BAX levels did not change significantly during TNF-α treatment course (), densitometric analysis revealed that BAX levels were consistently 20%–25% lower in telomerase-positive cells. When the levels of BAX or BAD to Bcl-2 were compared, the ratios of BAX/Bcl-2 and BAD/Bcl-2 were up to 30 times and 8 times greater, respectively, in telomerase-negative cells indicating an increased propensity for apoptosis in these cells compared with telomerase-positive cells (). In contrast, telomerase did not appear to alter the levels of the inhibitor of apoptosis protein (IAP), XIAP ().
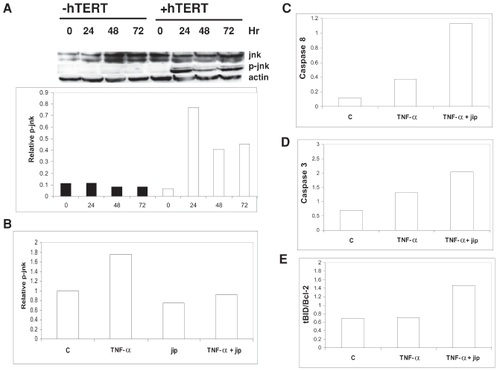
Telomerase-mediated anti-apoptosis is jnk-dependent
To identify a molecular signaling pathway involved in telomerase-mediated suppression of apoptosis, we assessed the contribution of jnk to suppress apoptosis since we found that induction of apoptosis in telomerase-positive cells was associated with a 2-fold increase in the levels of phosphorylated (p)-jnk compared with telomerase-negative cells (). In contrast, transfection with GFP as a procedural control was not associated with significant increases in p-jnk expression since p-jnk levels in GFP-transfected cells did not differ from untreated cells by more than 10% (data not shown). Co-transfection with jip, an inhibitor of jnk, not only reduced levels of p-jnk in telomerase-positive cells by 25%, but also abolished TNF-α-mediated induction of p-jnk in these cells (). Likewise, inhibition of jnk with jip in telomerase-positive cells restored apoptotic sensitivity as indicated by increased caspase 3 and 8 cleavage as well as increased levels of tBID/Bcl-2 following treatment with TNF-α (–).
Discussion
Apoptosis, programmed cell death, plays an important role in many normal and pathophysiological processes and is triggered when cellular death factors outweigh cellular survival factors. Aspartic acid-specific cysteine proteases (caspases), synthesized as zymogens, are the critical mediators of apoptosis (CitationCohen 1997). Whether activated by membrane-bound death receptors (CitationWalczak and Krammer 2000) or by stress-induced mitochondrial perturbation with subsequent cytochrome c release (CitationBratton et al 2000; CitationLoeffler and Kroemer 2000), the respective initiator caspases 8 and 9 function to activate downstream caspases 3 and 7 (CitationNicholson 1999). Once triggered, caspases lead to stepwise cellular destruction by disrupting the cytoskeleton, shutting down DNA replication and repair, degrading chromosomal DNA, and, finally, disintegrating the cell into apoptotic bodies (CitationNagata 2000).
In the present study, several lines of evidence suggest that telomerase confers resistance to epithelial cell apoptosis. First, activation of telomerase by transfection with hTERT resulted in reduced cisplatin-induced cytotoxicity as well as reduced DNA ladder formation. This is consistent with previous reports where telomerase activity in primary cultures of cancer cells was inversely related to chemosensitivity (CitationFaraoni et al 1999). Similarly, in a pilot study, CitationTakahashi et al (2000) showed that ovarian cancer patients responding to platinum therapy had low levels of telomerase whereas 50% of non-responders demonstrated elevated telomerase activity. Likewise, reduced apoptosis in pancreatic cancer cells after exposure to etoposide was associated with elevated telomerase activity (CitationSato et al 2000).
Second, ectopic expression of telomerase reduced apoptosis by suppressing caspase 3 cleavage in OSE cells. This appears to be the major pathway for telomerase-mediated resistance to apoptosis and has likewise been reported by CitationYamada et al (2003) who showed that overexpression of hTERT suppressed caspase 3 activity in K562 hematopoietic cells and CitationLuiten et al (2003) who showed reduced caspase 3 activation following ectopic expression of hTERT in T cells. However, there have been reports of telomerase-mediated resistance to apoptosis through caspase 3-independent means (CitationRen et al 2001), which serves to highlight the potential complexity of telomerase-mediated resistance to apoptosis. Interestingly, we also found that telomerase-mediated suppression of caspase 3 activation occurred in response to a number of apoptotic insults including CP, UV, and STS. Consequently, it appears that telomerase may confer global resistance to apoptosis and may be related to the propensity of many DNA-damaging agents to target the telomeric TTAGGG DNA sequence (CitationRamirez et al 2003).
Third, hTERT expression was sufficient and specific to confer resistance to apoptosis since co-transfection with dominant negative (DN) hTERT inhibited telomerase activity and abolished resistance to apoptosis. Likewise, we and others have shown that inhibition of telomerase in a variety of cancer cell types resulted in enhanced drug-induced apoptosis associated with increased caspase 3 activity (CitationKondo et al 1998; CitationKraemer et al 2004; CitationShoup et al 2004). Consequently, since telomerase re-activation is a crucial step for cellular immortalization and malignant transformation, inhibition of telomerase may act as a tumor-suppressive mechanism and have clinical utility as adjuvant therapy for enhanced chemosensitization.
Fourth, our data also indicate that telomerase suppresses an early step in the apoptotic cascade since, in addition to caspase 3 cleavage, telomerase re-activation suppressed cleavage of the upstream caspases 8 and 9, but did not affect levels of the caspase 3 inhibitor, XIAP. Further, we found that telomerase-mediated resistance to apoptosis was jnk-dependent because apoptosis associated with ectopic expression of hTERT resulted in elevated levels of p-jnk while suppression of jnk expression by jip abolished resistance to apoptosis. Interestingly, jnk promotes both pro-and anti-apoptotic functions, dependent upon differential phosphorylation of pro- and anti-apoptotic Bcl-2 family members. That is, jnk-mediated cleavage of BID to truncated (t)-BID results in mitochondrial release of cytochrome C whereas its phosphorylation of BAD at theronine 201 inhibits inactivation of the anti-apoptotic protein, BCL-Xl, by BAD (CitationDeng et al 2003; CitationYu et al 2004). In agreement, we found that re-expression of telomerase resulted in increased Bcl-2 expression consistent with studies showing that telomerase activity is higher in colorectal and cervical cancers expressing high levels of Bcl-2 (CitationMandal and Kumar 1997; CitationIida et al 2000). Further, the ratio of pro-apoptotic proteins tBID, BAD, or BAX/Bcl-2 remained low in telomerase-positive cells favoring resistance to apoptosis.
Age-dependent telomeric attrition in normal somatic cells, including endothelial cells, smooth muscle cells, cardiomyocytes, lymphocytes, and neurons, impairs cellular function so that telomeric shortening is thought to contribute to the pathogenesis of chronic diseases of aging including heart disease, diabetes, and Alzheimer’s disease (CitationJeanclos et al 1998; CitationHalvorsen et al 2000; CitationOh et al 2001, Citation2003; CitationObana et al 2003; CitationPanossian et al 2003; CitationZhang et al 2003; CitationNakashima et al 2004; CitationSerrano and Andres 2004). Accelerated telomeric shortening associated with progeroid and DNA damage repair syndromes, including Werner’s syndrome, Xeroderma pigmentosum, Ataxia telangiectasia, and Bloom’s syndrome (CitationDu et al 2004; CitationKruk et al 1995; CitationMetcalfe et al 1996; CitationMondello et al 1997), similarly contributes to accelerated cellular senescence and reduced viability. Likewise, aberrant telomerase regulation has been reported in developmental abnormalities, cancer, and disorders of the hematopoietic system. Specifically, in aplastic anemia and Fanconi’s anemia, the mean telomeric length of peripheral blood cells is considerably shorter than normal counterparts despite increased levels of telomerase, which is considered a result of high turnover in hematopoietic stem cells and high rates of telomere breaks (CitationGreenwood and Lansdorp 2003; CitationOhshima et al 2003; CitationPolychronopoulou and Koutroumba 2004). In addition, autosomal dominant dyskeratosis congenita is associated with mutations in the hTR component of telomerase while X-linked dyskeratosis congenita is due to mutations of dyskerin, a protein involved in RNA processing and that also resides within the telomerase complex (CitationMitchell et al 1999; CitationVulliamy et al 2001). Telomerase dysfunction in either form of dyskeratosis congenita results in progressive bone marrow failure. The potential clinical use of telomerase-positive cells or stem cells to expand proliferative capacity and replace damaged somatic cells could reverse or prevent symptoms associated with progeroid disorders, age-related diseases, or diseases associated with dysfunctional telomerase regulation and telomere maintenance. Indeed, CitationRoy et al (2004) showed that when fetal neuroepithelial cells were transfected with hTERT and transplanted at sites of spinal cord injury, they differentiated into functional neurons and became incorporated into the tissue at the damaged site. Using TERT transgenic mice, CitationOh et al (2003) showed that telomerase expression could delay cardiac myocyte cell cycle exit and stimulate cardiac hypertrophy. Though further studies are warranted, our data indicate that restoration of hTERT expression and telomerase activity in epithelial cells increases cell survival possibly by conferring resistance to apoptotic insults important in the pathogenesis of disease.
Figures
Figure 1 hTERT expression extends mean telomeric length and increases cell growth.
IOSE686 and IOSE80 cells were transfected ± hTERT cDNA and assayed for telomerase activity by PCR-ELISA (A), mean telomeric length by Southern blot analysis (B), and short term growth in cell culture by MTS assay (C). Telomerase activity and cell growth are expressed as the absorbance at 450 nm and 490 nm, respectively, ± SE while mean telomeric length is expressed as average kb ± SE.
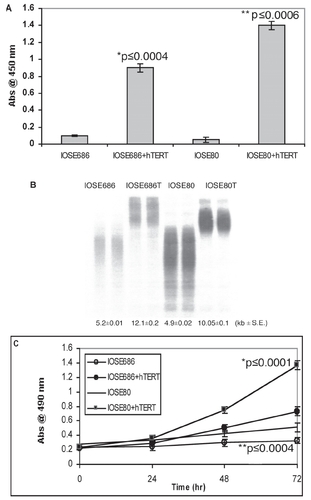
Figure 2 Telomerase enhances cell survival.
IMCC3 and IOSE80 cells were transfected ± hTERT cDNA, treated with 25 μM CP for 2 hr and then assayed for cell growth by MTS (A) or apoptosis by DNA laddering (B). Cell growth is expressed as the absorbance at 490 nm ± SE.
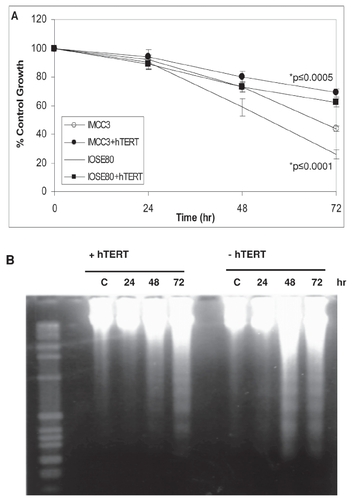
Figure 3 Telomerase confers resistance to caspase 3-dependent apoptosis.
IOSE cells transfected ± hTERT cDNA were treated with 25 μM CP for 2 hours, 20J/m2 (A), or 1 μM STS for 4 hours (B), and assayed for activated caspase 3 (17 kDa and 19 kDa bands) or DFF45 cleavage (45 kDa band) by Western blot analysis. (C) Control IOSE cells, IOSE cells transfected with hTERT, and IOSE cells transfected with both hTERT and DN hTERT cDNA were treated ± 1 μM STS and examined for telomerase activity by PCR-ELISA. Telomerase activity was expressed as the absorbance at 450 nm ± SE. Samples from (C) were analyzed by Western immunoblot for activated caspase 3 (D). Actin served as a loading control for all Western immunoblots.
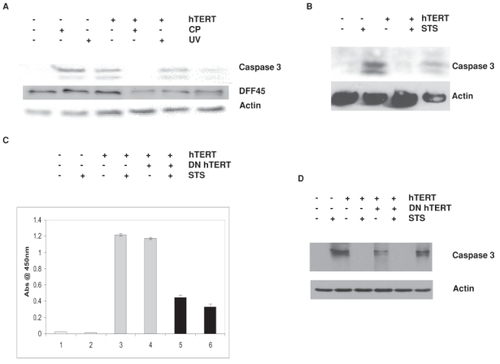
Figure 4 Telomerase-mediated anti-apoptosis is caspase-dependent.
IOSE cells transfected ± hTERT cDNA were treated with 1ng/ml TNF-α for 24 hours and assayed for telomerase activity by PCR-ELISA for up to 72 hours following treatment (A). Telomerase activity was expressed as the absorbance at 450 nm ± SE. Samples from (A) were also analyzed by Western immunoblot for procaspase 3, activated caspase 3 (B), cleaved caspase 9 (C), and cleaved caspase 8 (D). Actin served as a loading control for all Western immunoblots. Densitometric analysis of Western blots is provided in graphical presentation below respective blots.
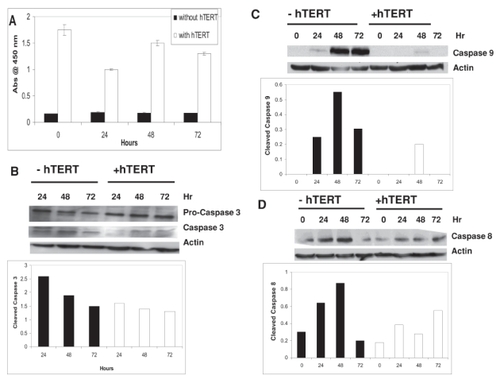
Figure 5 Telomerase alters pro- and anti-apoptotic mitochondrial protein expression.
IOSE cells transfected ± hTERT cDNA were treated with 1 mg/ml TNF-α for 24 hours and analyzed by Western immunoblot for Bcl-2 (A), tBID and BID (B), BAX (C), and XIAP (D). Actin served as a loading control for all Western immunoblots. Densitometric analysis of Western 2blots for the ratios of t-BID/Bcl-2, BAX/Bcl-2 and BAD/Bcl-2 are provided in graphical representation in (E–G), respectively.
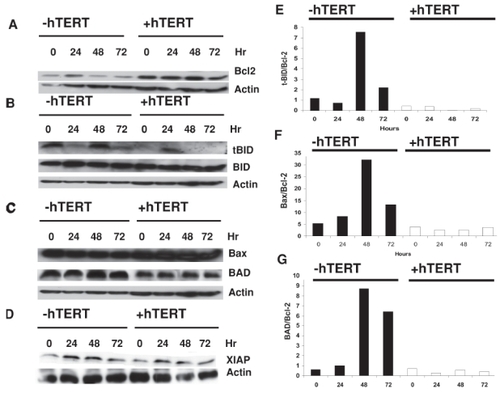
Figure 6 Telomerase-mediated anti-apoptosis is jnk-dependent.
(A) IOSE cells transfected ± hTERT cDNA were treated with 1 ng/ml TNF-α and analyzed by Western immunoblot for p-jnk and jnk. Densitometric analysis of Western blots for the levels of p-jnk is provided in graphical representation below the Western immunoblot. IOSE cells, transfected with hTERT cDNA ± jip cDNA, were treated with 1 ng/ml TNF-α for 24 hours and analyzed by Western immunoblot for p-jnk (B), cleaved caspase 8 (C), cleaved caspase 3 (D), and levels of tBID/Bcl-2 (E). The results are presented as densitometric analyses of Western blots in graphical presentation. Actin served as a loading control for all Western immunoblots.
Acknowledgements
We thank Ms Farheen S Qureshi for technical assistance and Drs. Vernon Mar and Nelly Auersperg for generous gifts of hTERT constructs and cells. This work was supported, in part, by US Army Department of Defense grant # DAMD17-02-1-0670 (Project #2) to PAK.
References
- Alfonso-De MatteMYChengJQKrukPA2001Ultraviolet irradiation-and dimethyl sulfoxide-induced telomerase activity in ovarian epithelial cell linesExp Cell Res267132711412034
- Alfonso-De MatteMYMoses-SotoHKrukPA2002Calcium induces telomerase activity in ovarian epithelial cell linesArch Biochem Biophys3992394411888211
- Alfonso-De MatteMYYangHChengJQ2002C-Jun kinase-mediated regulation of telomerase in ovarian surface epithelial cellsCancer Res624575812183409
- Alfonso-De MatteMYKrukPA2004Phosphotidylinositol-triphosphate kinase- and c-jun N-terminal kinase-dependent induction of telomerase by calcium requires pyk2Cancer Res6423614729602
- AllsoppRCChangEKashefi-AazamM1995Telomere shortening is associated with cell division in vitro and in vivoExp Cell Res2201942007664836
- AsaiAKiyozukaYYoshidaR1998Telomere length, telomerase activity and telomerase RNA expression in human esophageal cancer cells: correlation with cell proliferation, differentiation and chemosensitivity to anticancer drugsAnticancer Res181465729673357
- BachandFBoisvertFMCoteJ2002The product of the survival motor neuron (smn) gene is a human telomerase-associated proteinMol Cell Biol1331923202
- BlackburnEH1991Structure and function of telomeresNature350569721708110
- BrattonSBMacFarlaneMCainK2000Protein complexes activate distinct caspase cascades in death receptor and stress-induced apoptosisExp Cell Res256273310739648
- CohenGM1997Caspases: The executioners of apoptosisBiochem J3261169337844
- CongYSWenJBacchettiS1999The human telomerase catalytic subunit hTERT: organization of the gene and characterization of the promoterHum Mol Genet8137429887342
- CounterCMAvilionAALeFeuvreCE1992Telomere shortening associated with chromosome instability is arrested in immortal cells which express telomerase activityEMBO J11192191582420
- CounterCMMeyersonMEatonEN1998Telomerase activity is restored in human cells by ectopic expression of hTERT (hEST2), the catalytic subunit of telomeraseOncogene161217229528864
- deLangeT1992Human telomeres are attached to the nuclear matrixEMBO J11717241537344
- DengYRenXYangL2003A JNK-dependent pathway is required for TNF-α induced apoptosisCell115617014532003
- DuXShenJKuganN2004Telomere shortening exposes functions of the mouse Werner and Bloom syndrome genesMol Cell Biol2484374615367665
- EversoleAMaizelsN2000In vitro properties of the conserved mammalian protein hnrnp d suggest a role in telomere maintenanceMol Cell Biol2054253210891483
- FaraoniIGrazianiGTurrizianiM1999Suppression of telomerase activity as an indicator of drug-induced cytotoxicity against cancer cells: in vitro studies with fresh human tumor samplesLab Invest79993100510462037
- FengJFunkWWangS1995The human telomerase RNA componentSci269123641
- FisetSChabotB2001Hnrnp a1 may interact simultaneously with telomeric DNA and the human telomerase RNA in vitroNuc Acids Res29226875
- FordLPShayJWWrightWE2001The Ia antigen associated with the human telomerase ribonucleoprotein and influences telomere length in vivoRNA710687511497426
- FordLPSuhJMWrightWE2000Heterogeneous nuclear ribonucleoproteins c1 and c2 associates with the RNA component of human telomeraseMol Cell Biol2090849111074006
- FuWBegleyJGKillenMW1999Anti-apoptotic role of telomerase in pheochromocytoma cellsJ Biol Chem27472647110066788
- FuWKillenMCulmseeC2000The catalytic subunit of telomerase is expressed in developing brain neurons and serves a cell survival-promoting functionJ Mol Neurosci1431510854032
- GreenbergRAO’HaganRCDengH1999Telomerase reverse transcriptase gene is a direct target of c-Myc but is not functionally equivalent in cellular transformationOncogene1812192610022128
- GreenwoodMJLansdorpPM2003Telomeres, telomerase, and hematopoietic stem cell biologyArch Med Res344899514734088
- HalvorsenTLBeattieGMLopezAD2000Accelerated telomere shortening and senescence in human pancreatic islet cells stimulated to divide in vitroJ Endocrinol166103910856888
- HarleyCBVaziriHCounterCM1992The telomere hypothesis of cellular agingExp Gerontol27375821459213
- HastieNDDempsterMDunlopMG1990Telomere reduction in human colorectal carcinoma and with ageingNature34686682392154
- HayflickL1965The limited in vitro lifetime of human diploid cell strainsExp Cell Res376143614315085
- HerbertBSPittsAEBakerSI1999Inhibition of human telomerase in immortal human cells leads to progressive telomere shortening and cell deathProc Natl Acad Sci U S A961427781
- HoltSEAisnerDLBaurJ1999aFunctional requirement of p23 and hsp90 in telomerase complexesGenes Dev138172610197982
- HoltSEGlinskyVVIvanovaAB1999bResistance to apoptosis in human cells conferred by telomerase function and telomere stabilityMol Carcinogenesis252418
- IidaAYamaguchiAHiroseK2000Telomerase activity in colorectal cancer and its relationship to bcl-2 expressionJ Surg Oncol732192310797335
- JeanclosEKrolewskiASkurnickJ1998Shortened telomere length in white blood cells of patients with IDDMDiabetes4748269519758
- JohnsonNCKrukPA2002BRCA1 Zinc RING finger domain disruption alters caspase response in ovarian surface epithelial cellsCancer Cell Intl2116
- JohnsonNCDanHCChengJQ2004Loss of XIAP in BRCA1 185delAG human ovarian surface epithelial cells results in aberrant active caspase 3 ubiquitinationExp Cell Res29891615242757
- KammaHFujimotoMFujiwaraM2001Interaction of hnrnp a2/b1 isoforms with telomeric ssdna and the in vitro functionBiochem Biophys Res Commun2806253011162566
- KimNWPiatyszekMAProwseKR1994Specific association of human telomerase activity with immortal cells and cancerSci266201115
- KiyozukaYYamamotoDYangJ2000Correlation of chemosensitivity to anticancer drugs and telomere length, telomerase activity and telomerase RNA expression in human ovarian cancer cellsAnticancer Res202031210769656
- KondoYKondoSTanakaY1998Inhibition of telomerase increases the susceptibility of human malignant glioblastoma cells to cisplatin-induced apoptosisOncogene16224389619833
- KraemerKFuesselSKotzschM2004Chemosensitization of bladder cancer cells by human telomerase reverse transcriptase antisense treatmentJ Urol1722023815540782
- KrukPAMaines-BandieraSLAuerspergN1990A simplified method to culture human ovarian surface epitheliumLab Invest6313262374399
- KrukPARampinoNJBohrVA1995DNA damage and repair in telomeres: relation to agingProc Natl Acad Sci U S A92258627816828
- KrukPAGodwinAKHamiltonTC1999Telomeric instability and reduced proliferative potential in ovarian surface epithelial cells from women with a family history of cancerGynecol Oncol732293610329039
- LeSSternglanzRGreiderCW2000Identification of two RNA-binding proteins associated with human telomerase RNAMol Cell Biol119991010
- LoefflerMKroemerG2000The mitochondrion in cell death control: certainties and incognitaExp Cell Res256192610739647
- LuitenRMPeneJYsselH2003Ectopic hTERT expression extends the life span of human CD4+ helper and regulatory T-cell clones and confers resistance to oxidative stress-induced apoptosisBlood10145121912586632
- MandalMKumarR1997Bcl-2 modulates telomerase activityJ Biol Chem221418379162048
- MetcalfeJAParkhillJCampbellL1996Accelerated telomere shortening in ataxia telangiectasiaNature Genet1335038673136
- MeyersonMCounterCMEatonEN1997hEST2, the putative human telomerase catalytic subunit gene, is up-regulated in tumor cells and during immortalizationCell90785959288757
- MitchellJRWoodECollinsK1999A telomerase component is defective in the human disease dyskeratosis congenitalNature402551510591218
- MondelloCRiboniRCasatuA1997Chromosomal instability and telomere length variations during the life span of human fibroblast clonesExp Cell Res236385969367622
- NagataS2000Apoptotic DNA fragmentationExp Cell Res256121810739646
- NakashimaHOzonoRSuyamaC2004Telomere attrition in white blood cell correlating with cardiovascular damageHypertension Res Clin Exp2731925
- NakayamaJSaitoHNakamuraH1997Tlp1: a gene encoding a protein component of mammalian telomerase is a novel member of wd repeats familyCell88875849118230
- NicholsonDW1999Caspase structure, proteolytic substrates, and function during apoptotic cell deathCell Death Differ610284210578171
- ObanaNTakagiSKinouchiY2003Telomere shortening of peripheral blood mononuclear cells in coronary disease patients with metabolic diseaseInt Med421503
- OhHTaffetGEYoukerKA2001Telomerase reverse transcriptase promotes cardiac muscle cell proliferation, hypertrophy, and survivalProc Natl Acad Sci U S A98103081311517337
- OhHWangSCPrahashA2003Telomere attrition and Chk2 activation in human heart failureProc Natl Acad Sci U S A10053788312702777
- OhshimaKKarubeKShimazakiK2003Imbalance between apoptosis and telomerase activity in myelodysplastic syndromes: possible role in ineffective hemopoiesisLeuk Lymphoma4413394612952227
- OlovnikovAM1975A theory of marginotomy. The incomplete copying of template margin in enzymic synthesis of polynucleotides and biological significance of the phenomenonJ Theor Biol41181904754905
- PanossianLAPorterVRValenzuelaHF2003Telomere shortening in T cells correlates with Alzheimer’s disease statusNeurobiol Aging24778412493553
- PolychronopoulouSKoutroumbaP2004Telomere length variation and telomerase activity expression in patients with congenital and acquired aplastic anemiaActa Hematol11112531
- RamirezRCarracedoJJimenezR2003Massive telomere loss is an early event of DNA damage-induced apoptosisJ Biol Chem2788364212409303
- RenJGXiaHLTianYM2001Expression of telomerase inhibits hydroxyl radical-induced apoptosis in normal telomerase negative human lung fibroblastsFEBS Lett488133811163759
- RoyNSNakanoTKeyoungHM2004Telomerase immortalization of neuronally restricted progenitor cells derived from the human fetal spinal cordNature Biotech22297305
- SatoNMizumotoKKusumotoM2000Up-regulation of telomerase activity in human pancreatic cancer cells after exposure to etoposideBrit J Cancer8218192610839297
- SerranoALAndresV2004Telomeres and cardiovascular disease. Does size matter?Circ Res945758415031270
- ShoupBLLowellNEKrukPA2004Inhibition of telomerase improves chemosensitivity in cisplatin resistant ovarian cancer cellsCancer Therapy250110
- TakahashiMKigawaJOishiT2000Alteration of telomerase activity in ovarian cancer after chemotherapyGynecol Obstet Invest49204810729763
- TakakuraMKyoTHiranoH1999Cloning of human telomerase catalytic subunit (hTERT) gene promoter and identification of proximal core promoter sequences essential for transcriptional activation in immortalized and cancer cellsCancer Res5955179973199
- TianX-XPangJCSToSST1999Restoration of wild-type PTEN expression leads to apoptosis, induces differentiation, and reduces telomerase activity in human glioma cellsJ Neuropathol Exp Neurol58472910331435
- UlanerGAHuJ-FVuTH1998Telomerase activity in human development is regulated by human telomerase reverse transcriptase (hTERT) transcription and by alternate splicing of hTERT transcriptsCancer Res584168729751630
- VillaRFoliniMPeregoP2000Telomerase activity and telomere length in human ovarian cancer and melanoma cell lines: correlation with sensitivity to DNA damaging agentsIntl J Oncol169951002
- VulliamyTMarroneAGoldmanF2001The RNA component of telomerase is mutated in autosomal dominant dyskeratosis congenitalNature413432511574891
- WalczakHKrammerPH2000The CD95 (APO-1/Fas) and the TRAIL (APO-2L) apoptosis systemsExp Cell Res256586610739652
- WeinrichSLPruzanRMaL1997Reconstitution of human telomerase with the template RNA component hTR and the catalytic protein subunit hTERTNat Genet174985029398860
- YamadaOAkiyamaMKawauchiK2003Overexpression of telomerase confers a survival; advantage through suppression of TFR1 gene expression while maintaining differentiation characteristics in K562 cellsCell Transplant123657712911124
- YuCMinemotoYZhangJ2004JNK suppresses apoptosis via phosphorylation of the proapoptotic Bcl-2 family protein BADMol Cell133294014967141
- ZhangJKongQZhangZ2003Telomere dysfunction of lymphocytes in patients with Alzheimer diseaseCognit Behav Neurol16170614501538
- ZhangXMarVZhouW1999Telomere shortening and apoptosis in telomerase-inhibited human tumor cellsGenes Dev1323889910500096
- ZhuHFuWMattsonMP2000The catalytic subunit of telomerase protects neurons against amyloid beta-peptide-induced apoptosisJ Neurochem751172410854254