Abstract
We recently reported a novel neuro-immuno co-operation between vasoactive intestinal peptide (VIP) and fraktalkine (FKN) in recruiting human mast cells to the asthmatic airway that provided a classical example of priming effect on mast cells migratory function, but the role of the F-actin in human mast cell chemotaxis’ priming is poorly defined. Therefore the aim of this study was to further investigate the biophysical role of the cytoskeletal element; the F-actin, intracellular reorganization and its polymerization in mast cell priming of chemotaxis function. In the present communication it is shown by immunofluoresence confocal microscopy analysis that physical F-actin intracellular reorganization in a membrane bound manner on human mast cell is involved in VIP-induced priming of human mast cell chemotaxis against FKN. The F-actin reorganization was calcium independent and without modification of its contents as assessed by fluorescence-activated cell scanning analysis. These results identify a novel role for the biophysical association of F-actin in the crosstalk between neuro-inflammatory mediators and mast cells and may be an important target for therapeutic modalities in allergic inflammation.
Introduction
Human airway smooth muscle cells (HASMC) play a significant role in recruiting mast cells to the asthmatic bronchi (CitationBrightling et al 2005; CitationSutcliffe et al 2006). The contribution of fraktalkine (FKN), the CX3C chemokine produced by HASMC to mast cell chemotaxis, needed the cooperation of the neuropeptide vasoactive intestinal peptide (VIP), without altering FcɛR1 or CD117 expression on mast cells (CitationEl-Shazly et al 2006). Similar synergistic cooperation between stem cell factor and tumor necrosis factor-α on intercellular adhesion molecule-1 expression on human mast cells was demonstrated, which further provide insight for cross-talk between different signaling pathways that can help in recruiting mast cells to the allergic inflammatory site (CitationTsang et al 2005).
Chemotaxis primer is an agent that will not induce chemotaxis when it reacts with its specific receptor on the cell surface, but will put the cell in the ready-to-go position to re-stimulation and will do so even with a suboptimal dose of the proper chemokine that attracts the cell type. This scenario of crosstalk between mediators and inflammatory cells is applicable to the pathophysiology of allergic rhinitis and asthma where there is a pool of allergic mediators at different concentrations involved in the allergic site.
The literature review is rich in the different signal transduction pathways and the different protein kinases involved in mast cell chemotaxis. However, apart from its well acknowledged cytoskeletal shape changes, little is known about the role played by the F-actin in the signal transduction of mast cell chemotaxis (CitationJackson et al 2005; CitationSamayawardhena et al 2006; CitationRivera et al 2007; CitationKim et al 2007; CitationSamayawardhena et al 2007).
Accordingly, the current study was designed to look into the role of F-actin in the signal transduction of priming mast cell chemotaxis function. Our recently reported novel clinical crosstalk model of the neuro-immuno-inflamatory axis in recruiting mast cells to the airway; FKN-VIP-mast cells (CitationEl-Shazly et al 2006) were adopted for the following biophysical assay.
Materials and methods
Cell cultures
Human mast cell line (HMC-1) (CitationButterfield et al 1988) was cultured in 10% fetal calf serum/Dubecco’s modified Eagle’s Medium and was passaged every 3–5 days with trypsinethylenediaminetetraacetic acid (EDTA) (Invitrogen Life Technologies, Carlsbad, CA).
Chemotaxis assay
Chemotactic experiments were performed in 48-well microchemotaxis chambers as described elsewhere (CitationBerger et al 2001). As for chemotaxis assay of primed HMC-1, cells were first incubated with 1 μM VIP for 30 minutes (Sigma-Aldrich, St. Louis, MO), washed twice and resuspended in the culture media before inducing their chemotaxis response against 10 ng/ml FKN (R and D Systems, Minneapolis, MN). Mast cells were then counted by microscopy in 5 selected high power fields (magnification × 400). Results of the chemo-taxis experiments were expressed as mast cells/5 high power fields (5hpf).
Microspectrofluorimetry
Changes in HMC-1 intracellular calcium concentration were assessed using the Ca2+-sensitive probe indo-1 (calbiochem) precisely as previously described (CitationBonnet et al 2003). 10 μM ATP was used as positive control that demonstrated the ability of HMC-1 to mobilize intracellular calcium. 1 μM VIP, 10 ng/ml FKN, and 1 μM VIP + 10 ng/ml FKN ability to mobilize intracellular calcium were tested in similar fashion.
Actin reorganization assessment with phalloidin-FITC
As for actin reorganization and content assessment, after each challenge, HMC-1 were fixed in cold methanol for 20 minutes and permeabilized with 0.1% saponin (Sigma-Aldrich) for another 30 minutes. The cells were then stained with phalloidin-fluorescein isothiocyanate (FITC) (Sigma-Aldrich) labeled for 1 hour in the dark, on ice, and analyzed by fluorescence-activated cell scanning (FACS) or confocal microscopy.
Results and discussion
In the past decade or so, intensive research in the fields of neuropeptides and immune cells has resulted in accumulating evidence that supports the existence of neuroimmune axis (CitationNuma et al 1992; CitationEl-Shazly et al 1996, Citation1997). The 28 amino acid VIP polypeptide, which exists in the parasympathetic nerves, and to a lesser extent in the sensory fibers, is one of the most abundant of the neuropeptides found in the upper and lower airways (CitationGhatei et al 1982; CitationBaranuik et al 1990). It has been detected in the fluids of hypersensitivity reactions (CitationMosimann et al 1993). The biological effects of VIP on airway functions include regulation of vascular tone, airway caliber, and mucus secretion (CitationSaid 1982; CitationPeatfield et al 1983; CitationPalmer et al 1986; CitationStjarne et al 1991).
On inflammatory cells, VIP has the potential to modulate T-lymphocyte and human eosinophil chemotaxis (CitationDelgado et al 1995; CitationEl-Shazly et al 2000) at wide range of doses. However, VIP at the same wide range doses did not induce human mast cell chemotaxis, but acted as a primer to suboptimal dose of 10 ng/ml FKN-induced mast cell chemotaxis (CitationEl-Shazly et al 2006). Nonetheless, little is known about the role of F-actin in mast cell priming of chemotaxis function.
FKN the CX3C chemokine attracts human mast cells in boyden chambers in a classical bell-like shape dose-dependent manner with optimal doses of 25–50 ng/ml through a CX3CR1 receptor stimulation that is linked to PKC epsilon and delta, increase in F-actin contents and rapid shape changes (CitationEl-Shazly et al 2006). Herein, it is shown that the low nonstimulatory dose of 10 ng/ml caused mast cell chemotaxis after the cells have been primed by 1 μM VIP that, per se, did not cause chemotaxis ().
Figure 1 (a) Priming effect of 1 μM VIP on 10 ng/ml FKN-induced mast cells chemotaxis. Upper panel (i, A1–D1) demonstrate the data of the number of mast cells per 5 selected high power field (5 hpf; × 400) ± SEM of 6 independent experiments each performed in triplicate. Asterisks indicate p < 0.05 by paired t-test. Lower panel (ii, A2–D2) demonstrates mast cells in 1 selected HPF as one experiment representative of 6 all showing similar results. (b) Effect of 1μM ATP, 1μM VIP, 10 ng/ml FKN, and 1 μM VIP + 10 ng/ml FKN on mast cells intracellular calcium mobilization. Results are of one representative of 4 independent experiments all showing similar results. Traces are from 27–125 cells. (c) Fluocytometry of F-actin in mast cells. Y-axis represent cell count while X-axis represent the mean fluorescent, where 1 = Control, 2 = Buffer, 3 =VIP (1 μM), 4 = FKN (10 ng/ml), 5 = VIP (1 μM) + FKN (10 ng/ml). Histograms are one representative of 6 different experiments all showing similar results. (d) F-actin dynamics and cell shape changes induced by different stimulants. (A) Control-15 minutes stimulation (Buffer), (B) VIP 1 μM-15 minutes stimulation, (c) VIP 1 μM-30 minutes stimulation, (D) FKN 10 ng/ml-15 minutes stimulation (E) VIP 1μM-15 minutes stimulation, cells were washed twice, and re-stimulated with FKN 10ng/ml for 5 minutes. Images are from one representative of 6 independent experiments which all show similar images.
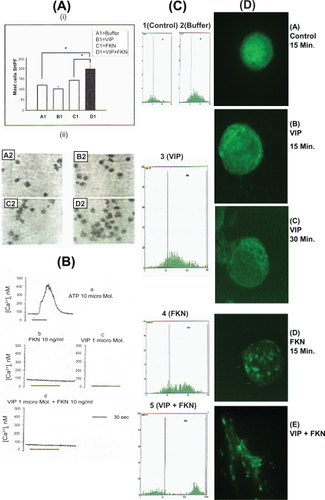
Chemokine-mediated signal transduction is believed to involve (i) Ca2+ mobilization, protein kinase C and heterotrimeric GTP-binding proteins in a classical view (CitationDowney 1994), and (ii) kinases and phosphatases, adaptor proteins, and small GTP-binding proteins in an alternate view (CitationBokoch 1995; CitationBacon et al 1996; CitationKanal et al 1997). Thus, the role of Ca2+ as a second messenger was investigated. Neither FKN nor VIP mobilized intracellular Ca2+ in mast cells. Similar results were obtained after the cells have been primed with VIP () indicating that the VIP-priming effect was Ca2+-independent.
The suboptimal dose of 10 ng/ml FKN did not cause significant increase of F-actin contents or shape changes, but reorganized the intracellular F-actin homogeneously in a clump-like manner within the spherical cell (compare FACS results 1 and 2 with 4 in and confocal images A with D in ). However, the same suboptimal dose caused a rapid increase in the F-actin contents and shape changes after the cells were primed by a physiological dose of VIP (compare FACS results 1 and 2 with 5 in and confocal images A with E in ). VIP alone did not cause any increase in the F-actin contents or shape changes but caused a characteristic peripheral, membrane bound intracellular reorganization of F-actin in the spherical cells in a time-dependent manner (compare FACS results of 1 and 2 with 3 in and confocal images A and B with C in ).
The crosstalk between different agonist stimulants of cell receptors may indicate a novel role for F-actin physical reorganization independent from shape changes and increase in its contents in mast cells migration. In this context it is possible that high and low affinity receptors of chemokines (FKN) on the surface of mast cells are differently linked to F-actin changes, ie, the optimal dose will cause a rapid increase in F-actin contents that is associated with the rapid intracellular reorganization and shape changes through PKC epsilon and delta (CitationEl-Shazly et al 2006), while the suboptimal dose will only reorganize the F-actin homogeneously in a cluster-like manner as shown in this study. On the other hand, chemotaxis primers of mast cells such as VIP may induce a calcium-independent change in the signal transduction towards tyrosine kinases and p38 MAPK as we reported elsewhere (CitationEl-Shazly et al 2006) that is linked to peripheral reorganization of the F-actin in the spherical cell in a membrane-bound manner without increasing the F-actin contents or causing shape changes as demonstrated in this study. This is further supported by previous reports linking the involvement of p38-MAPK with the physical reorganization of F-actin (CitationKutsuna et al 2004; CitationKobayashi et al 2006; CitationJu et al 2007; CitationKim et al 2008).
Taken collectively, the results presented herein may identify a significant role for the unique pattern of the Ca2+-independent F-actin biophysical dynamics in human mast cells migratory response to crosstalk between mediators and cell type at the inflammatory site. If so, then antagonists at the level of cell receptors may provide a therapeutic target for F-actin in treating mast cells in allergic inflammation.
Acknowledgements
The author wishes to acknowledge Prof. Tunon De Lara J. M. and Dr Patrick Berger for their critical discussion as coauthors in related previously published work and for the materials used. Also a special thank is due to Prof. Marthan R. for the laboratory facilities at the Laboratoire de Physiologie Cellulaire Respiratoire, Universite Victor Segalen Bordeaux 2-France. The author reports no conflicts of interest in this work.
References
- BaconKBSzaboMCYsselH1996RANTES induces tyrosine kinase activity of stably complexed p125 FAK ands ZAP-70 in human T cellsJ Exp Med184873829064347
- BaraniukJNOkayamaMLundgrenJD1990Vasoactive intestinal peptide in human nasal mucosaJ Clin Invest86825312203825
- BergerPTunon-de-LaraJMSavineauJP2001Tryptase-induced PAR-2-mediated Ca2+ signaling in human airway smooth muscle cellsJ Appl Physiol919951003
- BokochGM1995Chemoattractant signaling and leukocyte activationBlood861649607654998
- BonnetSDumas-de-La-roqueEBégueretH2003Dehydroepiandrosterone (DHEA) prevents and reverses chronic hypoxic pulmonary hypertensionProc Natl Acad Sci U S A10094889312878719
- BrightlingCEAmmitAJKaurD2005The CXCL10/CXCR3 axis mediates human lung mast cell migration to asthmatic airway smooth muscleAm J Respir Crit Care Med1711103815879427
- ButterfieldJHWeilerDDewaldG1988Establishment of an immature mast cell line from a patient with mast cell leukemiaLeuk Res12345553131594
- DelgadoMDe la FuenteMMartinezC1995Pituitary abenylate cyclase-activating polypeptides inhibit the mobility of murine thymocytes and splenic lymphocytes: Comparison with and implication of cAMPJ Neuro Immunol6213746
- DowneyGP1994Mechanisms of leukocyte motility and chemotaxisCurr Opin Immunol6113248172673
- El-ShazlyABergerPGirodetPO2006Fraktalkine produced by airway smooth muscle cells contributes to mast cell recruitment in asthmaJ Immunol1761860816424217
- El-ShazlyAMasuyamaKEuraM1996Immunoregulatory role of substance P in human eosinophil migratory functionImmunol Invest251912019157054
- El-ShazlyAMasuyamaKIshikawaT1997Mechanisms involved in activation of human eosinophil exocytosis by substance P: An in vitro model of sensory neuroimmunomodulationImmunol Invest26615299399104
- El-ShazlyAMasuyanmaKTsunodaN2000Non-specific activation of human eosinophil functional responses by vasoactive intestinal peptideAllergology Int491926
- GhateiMASheppardMO’ShaughnessyDJ1982Regulatory peptides in the mammalian respiratory tractEndocrinology1111248546180890
- JacksonNEWangHWTedlaN2005IL-15 induces mast cell migration via a pertussis toxin-sensitive receptorEur J Immunol3523768516047340
- JuYWangTLiY2007Coxsackievirus B3 affects endothelial tight junctions: possible relationship to ZO-1 and F-actin, as well as p38 MAPK activityCell Biol Int3112071317544707
- KimSJJeongHJParkRK2007The regulatory effect of SC-236 (4-[5-(4chlorophenyl)-3-(trifluoromethyl)-1-pyrazol-1-l]benzenesulfonamide) on stem cell factor induced migration of mast cellsToxicol Appl Pharmacol2201384517320132
- KimMSKewalramaniGPuthanveetilP2008Acute diabetes moderates trafficking of cardiac lipoprotein lipase through p38 mitogen-activatid protein kinase-dependent actin cytoskeleton organizationDiabetes57647617942824
- KnallCWorthenGSJohnsonGL1997Interleukin 8-stimulated phosphatidylinositol-3-kinase activity regulates the migration of human neutrophils independent of ectracellular signal-regulated kinase and p38 mitogen-activated protein kinasesProc Natl Acad Sci U S A94305279096344
- KobayashiMNishitaMMishimaT2006MAPKAPK-2-mediated LIM-kinase activation is critical for VEGF-induced actin remodeling and cell migrationEMBO J257132616456544
- KutsunaHSuzukiKKamataN2004Actin reorganization and morphological changes in human neutrophils stimulated by TNF, GM-CSF, and G-CSF: the role of MAP kinasesAm J Physiol Cell Physiol2865564
- MosimannBLWhiteMVHohmanRJ1993Substance P, calcitonin gene-related peptide, and vasoactive intestinal peptide increase in nasal secretions after allergen challenge in atopic patientsJ Allegy Clin Immunol9295104
- NumaoTAgrawalDK1992Neuropeptides modulate human eosinophil chemotaxisJ Immunol1493309151385521
- PalmerJBDCussFMCBarnesPJ1986VIP and PHM and their role in nonadrenergic inhibitory responses in isolated human airwayJ Appl Physiol61132283781947
- PeatfieldACBarnesPJBratcherC1983Vasoactive intestinal peptide stimulates tracheal submucosal gland secretion in ferretAm Rev Respir Dis12889936870072
- RiveraJOliveraA2007Src family kinases and lipid mediators in control of allergic inflammationImmunol Rev2172556817498064
- SaidSI1982Vasoactive peptides in the lung, with special reference to vasoactive intestinal peptideExp Lung Res334386762962
- SamayawardhenaLAHuJSteinPL2006Fyn kinase acts upstream of Shp2 and p38 mitogen-activated protein kinase to promote chemotaxis of mast cells towards stem cell factorCell Signal1814475416442778
- SamayawardhenaLAKapurRCraigAW2007Involvement of Fyn kinase in Kit and integrin-mediated Rac activation, cytoskeletal reorganization, and chemotaxis of mast cellsBlood10936798617213284
- StjarnePLacroixJSAnggardA1991Compartment analysis of vascular effects of neuropeptides and capsaicin in pig nasal mucosaActa Physiol Scand141335421713399
- SutcliffeAKaurDPageS2006Mast cell migration to Th2 stimulated airway smooth muscle from asthmaticsThorax616576216601090
- TsangCMWongCKIpWK2005Synergistic effect of SCF and TNF-alpha on the up-regulation of cell-surface expression of ICAM-1 on human leukemic mast cell line (HMC)-1 cellsJ Leukoc Biol782394715800027