Abstract
As the population ages, there is a growing need for effective therapies for the treatment of neurological diseases. A limited number of therapeutics are currently available to improve cognitive function and research is limited by the need for in vivo models. Zebrafish have recently become a focus of neurobehavioral studies since larvae display neuropathological and behavioral phenotypes that are quantifiable and relate to those seen in man. Due to the small size of Zebrafish larvae, assays can be undertaken in 96 well plates and as the larvae can live in as little as 200 μl of fluid, only a few milligrams of compound are needed for screening. Thus in vivo analysis of the effects of compounds can be undertaken at much earlier stages in the drug discovery process. This review will look at the utility of the zebrafish in the study of neurological diseases and its role in improving the throughput of candidate compounds in in vivo screens.
Introduction
Cognitive impairment manifests itself in a number of neurological diseases such as schizophrenia, Huntington’s, Parkinson’s, and Alzheimer’s diseases. As the general population ages, there will be an increased incidence of these disease and disorders and thus a growing need for effective therapies for the treatment of the associated cognitive deficits. A limited number of therapeutics are currently available to improve cognitive function and research is limited by the need for predictive in vivo models.
Introduction to zebrafish
Zebrafish have recently become a focus of neurobehavioral studies since larvae display learning, sleep, drug addiction, and other neurobehavioral phenotypes that are quantifiable and relate to those seen in man (CitationZhdanova et al 2001; CitationCahill 2002; CitationGuo 2004; CitationOrger et al 2004; CitationNinkovic et al 2006). Furthermore, the organization of the zebrafish genome and the genetic pathways controlling signal transduction and development are highly conserved between zebrafish and man (CitationPostlethwait et al 2000). At seven days post fertilization (d.p.f.) the larvae are approximately 4 mm long. Due to this small size of the larvae, assays can be undertaken in 96 well plates and as the larvae can live in as little as 200 μl of fluid; only a few milligrams of compound are needed for screening. Thus in vivo analysis of the effects of compounds can be undertaken at much earlier stages in the drug discovery process than has previously been possible, which is facilitated by the fact that zebrafish are dimethyl sulfoxide (DMSO) tolerant and readily absorb compounds from the water. The relative ease of maintaining large stocks of fish and its high fecundity can provide the investigator with large numbers of larvae to analyze. The above properties have established the zebrafish as an excellent model system that is relevant to studies of human diseases (CitationGrunwald and Eisen 2002). Conventional drug discovery has recently employed systematic, target-based high throughput screening in purified proteins or cells as primary screens with in vivo models as tertiary screens in the cascade after more mechanistic cell assays. While the in vitro screens have been successful at identifying small molecules affecting known mechanisms, there is still the need to identify modulators of complex in vivo phenotypes in the whole organism for less well understood pathways or those that only occur in a physiological/pathophysiological context. The advantages of using larval zebrafish described above allow higher throughput in vivo screening for phenotypic endpoints, and the utility of zebrafish in small molecule screening has been the subject of several reviews (CitationMacRae and Peterson 2003; CitationZon and Peterson 2005; CitationMurphey and Zon 2006; CitationBerger and Currie 2007). However, there are disadvantages to this approach. Namely, that uptake of compound into the zebrafish can be variable and should be measured for accurate interpretation of results (CitationBerghmans et al 2008) and particularly to avoid false negatives, and the larval stage of the zebrafish may not be appropriate in all disease areas.
In a comparison of the zebrafish brain structure with man, there are some differences between teleosts and mammals. Notably, fish have smaller cerebral hemispheres and there are differences in the layout of the forebrain (extensively reviewed by CitationWullimann and Mueller 2004) and the structure and function of the optic tectum (CitationLuque et al 2005). However, the overall organisation of the zebrafish brain is similar to other vertebrates, having similarly defined areas such the hypothalamus and olfactory bulb, encompassing structures of the lateral pallium, which appear to be homologous to the mammalian hippocampus (CitationTropepe and Sive 2003). In addition, the main neurotransmitter systems such as the cholinergic, dopaminergic, and noradrenergic pathways are present and have been mapped throughout the brain (CitationRink and Wullimann 2004; CitationWullimann and Mueller 2004). Zebrafish have a developmentally regulated blood-brain barrier. Functional analysis using fluorescent dyes and anatomical analysis by transmission electron microscopy provides evidence that the zebrafish blood brain barrier is functional at 10 d.p.f. (CitationGoldsmith and Fleming 2007). Additionally, it was also shown that zebrafish paralogues of P-glycoprotein (Pgp) are first detected in the vasculature endothelium of the central nervous system (CNS) at 8 d.p.f., which coincides with the efflux of the Pgp substrate, rhodamine 123 from the zebrafish brain. These data suggest that the zebrafish is a relevant model for the study of neurological diseases.
Zebrafish neurodegenerative disease models
Huntington’s disease
Huntington’s disease (HD) is a monogenic polyglutamine (polyQ) neurodegenerative disorder which results in cognitive deficits in attentional and executive functions along with defects in visual working memory (CitationMontoya et al 2006). Neurodegeneration occurs primarily in the striatal medium-sized spiny neurons which project to the substantia nigra and globus pallidus. HD has a prevalence of approximately 5 in 100,000 worldwide, the median age of onset being 39 (CitationCowan and Raymond 2006). In zebrafish, the huntingtin (Htt) gene has been cloned and sequenced with a 3121 predicted amino acid protein, which has 70% identity with the human peptide sequence (CitationKarlovich et al 1998). Knockdown of Htt using morpholino technology disrupted a number of features of zebrafish development resulting in small head and eyes, delayed or paler pigmentation and colorless hypochromic blood (CitationLumsden et al 2007). In a separate study, a ‘Huntington’s like’ zebrafish was created by inserting mRNA of the N-terminal fragment of Htt with different length polyQ repeats linked to a GFP-fusion protein (CitationSchiffer et al 2007). The increasing polyQ length was associated with an increase in abnormalities and apoptosis in the embryos as early as 24 hours p.f. The embryos containing the Q102-GFP developed inclusions in the cytoplasm, which increased in size by incorporation of the soluble Q102 peptide leading to insoluble deposits. These findings confirmed a previous study where expression of poly Q56 or greater exhibited toxicity and abnormalities in the zebrafish embryos with inclusion bodies formed in more than 70% of embryos (CitationMiller et al 2005). These studies also investigated the effect of aggregation inhibitors which suggested that the prevention of aggregation did not reduce the toxic effect on the fish, implying that the formation of smaller intermediate aggregates were the main cause of toxicity. Thus, these models could be used to screen for novel compounds for the treatment of HD by evaluating either the prevention of aggregate formation, enhanced clearance of aggregates or the reduction in embryo death.
Alzheimer’s disease
Alzheimer’s disease (AD) is the most common cause of dementia, with nearly 50% of dementia cases worldwide being attributed to AD. The prevalence increases from 1.53% of the population between 65–69 years to as high as 30% between 80–85 years (CitationVandenberghe and Tournoy 2005; CitationMount and Downton 2006). AD is characterized histopathologically by amyloid-beta (Aβ) containing plaques and intracellular neurofibrillary tangles consisting of abnormally phosphorylated tau protein in the brain (CitationSelkoe 2000; CitationMudher and Lovestone 2002). Cognitive impairments manifest themselves as progressive episodic memory loss and effects on executive functions, which are usually accompanied by mini-mental state examination scores below 24 (CitationVandenberghe and Tournoy 2005). Zebrafish possess two homologues of amyloid precursor protein (APP) with good (about 70%) homology to human APP (CitationMusa et al 2001). Studies have demonstrated the presence of the functional γ-secretase machinery to produce Aβ in zebrafish (for a full review see CitationNewman et al 2007). Other studies have investigated the tau protein in zebrafish (CitationTomasiewicz et al 2002). Microinjection of four repeat human tau GFP constructs into 1–2 cell stage embryos showed disruption to the cytoskeletal structure and tau trafficking by 48 hours post injection. This eventually led to hyperphosphorylated fibrillar tau staining similar to that seen with neurofibrillary tangles in pathology of AD patients. These models offer the ability to screen for novel therapeutics that that decrease Aβ load and decrease the hyperphosphorylation seen in tauopathies.
Parkinson’s disease
Although predominantly a movement disorder, there are a number of cognitive impairments associated with Parkinson’s disease (PD) (CitationGalvin 2006). These include executive dysfunction and impaired memory retrieval and the prevalence increases from 2.7% per year at 55–64 years to 13.7% at 70–79 years. Parkinson’s disease is characterized neuropathologically by degeneration of dopaminergic neurons and the appearance of intracytoplasmic inclusions called Lewy bodies. Genetically, six genes linked to Parkinsonism have been identified: Parkin, DJ-1, PINK1, α-Synuclein, UCHL-1, and LRRK2 (CitationAbeliovich and Beal 2006). Of these genes two have been studied in zebrafish with preliminary work beginning on the others (CitationPaquet et al 2006; CitationShankaran et al 2006).
Ubiquitin carboxy-terminal hydrolase L1 (UCH-L1) was originally identified in siblings with a strong family history of PD (CitationLeroy et al 1998). In zebrafish, UCH-L1 mRNA was expressed in neuronal cells at 1 d.p.f. (CitationSon et al 2003). UCH-L1 was detected in the diencephalon and ventral region of the mid and hindbrain, which are associated with motoneuron development (although which mammalian motoneurons these were correlated with was not investigated). Interestingly, in the ventral diencephalon, an area functionally homologous to the substantia nigra in humans, the UCH-L1 was co-expressed with tyrosine hydroxylase substantiating the association of UCH-L1 with dopaminergic neurons.
Patients with autosomal recessively inherited DJ-1 mutations typically present with early onset PD. In embryonic and adult zebrafish DJ-1 is expressed throughout the body with higher abundance in the brain, eyes, heart and muscle of the adult (CitationBretaud et al 2007). The zebrafish DJ-1 protein has high homology with human (83%) and mouse DJ-1 (80%). Knockdown of DJ-1 in the zebrafish did not affect the number of dopaminergic neurons in a similar manner to the mouse DJ-1 null mutant (CitationChen et al 2005). In keeping with the suggested role of DJ-1, the knockdown zebrafish embryos were more susceptible to oxidative stress and had significantly elevated SOD1 levels. Furthermore, simultaneous knockdown of DJ-1 and p53 caused dopaminergic neuronal loss demonstrating a strong interaction between these genes.
The main pharmacological approach to studying PD in animal models has been using 1-methythl-4-phenyl-1, 2, 3, 6-tetrahydropyridine (MPTP) which reproduces some of the effects of idiopathic Parkinson’s disease (CitationSmeyne and Jackson-Lewis 2005). Zebrafish embryos treated with MPTP demonstrated a loss of TH and DAT- expressing neurons which could be rescued using the monoamine oxidase-B inhibitor deprenyl (CitationBretaud et al 2004; CitationLam et al 2005; CitationMcKinley et al 2005). A reduction was also seen in the locomotor activity mimicking motor effects seen in PD patients; however this did not always occur simultaneously with the reduction in dopaminergic cells (CitationSalzmann et al 2006) and in the adult a reduction is seen in the locomotor activity without an effect being seen in dopaminergic cells (CitationAnichtchik et al 2004). The utility of these various models will aid screening for novel compounds for both the hereditary and the idiopathic forms of PD.
Schizophrenia
Schizophrenia affects approximately 1% of the world’s population and is characterized by neuronal dysfunction resulting in deficits in a number of different cognitive areas such as visual and verbal memory and learning and attention (CitationNuechterlein et al 2004; CitationTamminga 2006). Patients with schizophrenia and other conditions including Huntington’s disease have been shown to exhibit impaired prepulse inhibition (PPI) (CitationSwerdlow et al 1995; CitationBraff et al 2001). PPI is a neurological phenomenon in which a weak prestimulus or prepulse suppresses the response to a subsequent startling stimulus and is highly conserved among vertebrates. A recent study of sensorimotor gating in zebrafish described an assay for PPI in 6 d.p.f. larvae (CitationBurgess and Granato 2007). In this study, the effective interstimulus interval for inhibition along with the dopaminergic and glutamergic modulation of PPI was found to be comparable to that in mammals. Again, using this paradigm will allow the screening of novel therapeutics for schizophrenia at an earlier stage of the drug discovery process.
Assays for assessing learning and memory in zebrafish
Mild cognitive impairment, a risk factor for AD, consists of a number of etiologies and is characterized by a slight impairment (usually memory) in cognitive function of everyday activities (CitationLevey et al 2006). Its prevalence in epidemiological studies ranges from 3%–19% in adults over 65 years old, with half of adults diagnosed with MCI progressing to dementia (CitationGauthier et al 2006). There have been a number of models developed to facilitate the study of learning and memory in zebrafish. Nonassociative learning has been studied in larval zebrafish by monitoring the reduction in a startle response to a series of acoustic stimuli (CitationBest et al 2007). This habituation to the stimuli was modulated by current therapies for improving cognitive ability in Alzheimer’s disease patients, namely donepezil and memantine, as well as by the phosphodiesterase (PDE) 4 inhibitor rolipram, a compound shown to increase learning and memory in rodent studies. Associative learning has been demonstrated by visual discrimination in a T maze with food reward linked to certain colours (CitationColwill et al 2005). The adult zebrafish learned to associate one of a pair of colours with food and reversal and extinction were also shown in this system. A T maze has also been used to determine the roles of glutamergic transmission (CitationSwain et al 2004) and histamine in learning and memory in adult zebrafish (CitationPeitsaro et al 2003). Furthermore, adult zebrafish have been shown to display quick and reliable learning by swimming to certain locations within a tank to avoid adverse stimulus, such as a moving net (CitationArthur and Levin 2001) or electrical shocks (CitationXu et al 2007) and show cognitive flexibility in that they show reversal when the contingencies are changed over. Active avoidance to avoid mild electrical shocks was used in a similar experimental paradigm to demonstrate the possible involvement of cell adhesion molecule L1.1 in memory consolidation (CitationPradel et al 2000). A three compartment maze with a central chamber and a choice of moving to the left or right chambers has been used to demonstrate spatial and non spatial visual discrimination learning (CitationArthur and Levin 2001) and to demonstrate the improvement in learning in response to nicotine treatment (CitationLevin et al 2006a) in adult zebrafish. A spatial alternation paradigm has been used to show that adult zebrafish learn to alternate between ends of the test tank for food reward, rapidly achieving high correct response levels (CitationWilliams et al 2002). This behavior is recalled if the zebrafish are tested again after 10 days. This type of learning can be impaired in zebrafish treated with ethanol (10 mM) or lead (10 μM) for the first 24 hours post fertilization of development as they demonstrated significant learning and memory deficits when treated in the three compartment maze assay (CitationCarvan et al 2004). The progress in the area of learning and memory experimental paradigms in zebrafish suggest the possible utility of this model organism in behavioral pharmacology studies.
Pharmacology mediating cognition
One of the key challenges with CNS drug discovery is the need to isolate specific areas of the brain mediating a particular disorder or disease. Therapies have become available for cognitive impairment, although these have been dominated by two classes of drug, acetylcholinesterase inhibitors and N-methyl-D-aspartate (NMDA) antagonists exemplified by donepezil and memantine, respectively. Since a number of publications and reviews have covered these therapies, the pathways they mediate and their effects in both preclinical models (CitationLevin and Simon 1998; CitationLevin et al 2006b; CitationCosman et al 2007) and clinical trials (CitationBurns et al 1999; CitationBirks 2006; CitationGeerts and Grossberg 2006; CitationRingman and Cummings 2006; CitationRobinson and Keating 2006), as well as the effects of these pathways on zebrafish behavior (CitationLevin and Chen 2004; CitationNam et al 2004; CitationLevin et al 2006a; CitationMcDearmid and Drapeau 2006), the latter part of this review will concentrate on four other interesting targets; these being phosphodiesterases, histamine H3 receptors, 5HT6 receptors, and AMPAkines, which are currently being investigated preclinically and the utility of zebrafish to aid in the characterization of these targets.
Phosphodiesterase inhibitors
The role of phosphodiesterases in cognition have been well documented (for reviews see CitationRose et al 2005; CitationHebb and Robertson 2007). Currently, in mammals there are 11 distinct families encoding 21 genes. Of these, PDE 1, 2, 4, and 10 are primarily located in the mammalian brain. PDE1B and PDE10 are located in the striatum and expression levels of both were found to decline in the HD mouse models R6/1 and R6/2, suggesting a role for these PDEs in the deficits associated with HD (CitationHebb et al 2004). The PDE 4 A, B, and C isoforms are distributed throughout regions of the brain, with PDE4A and 4D observed in the hippocampus along with 4D, and all three isoforms were seen to different extents in the cortical regions (CitationCherry and Davis 1999). Furthermore, although the levels of PDE4A were low in the substantia nigra there was no change in levels between the control and HD mice (CitationHebb et al 2004). Since the levels of PDE4s may not change in Huntington’s disease it may provide a viable target for the associated cognitive deficits.
Treatment of rodents with rolipram, a nonselective PDE4 inhibitor, demonstrated either enhancement of memory and long term potentiation or a reversal of pharmacologically induced memory deficit (CitationImanishi et al 1997; CitationBarad et al 1998; CitationZhang et al 2000; CitationZhang and O’Donnell 2000). Other studies with the PDE5 inhibitor sildenafil, also demonstrated cognitive improvement in the object recognition and attenuated the scopolamine induced learning deficit in mice suggesting this family as a possible therapeutic target (CitationPrickaerts et al 2002; CitationDevan et al 2004). Futhermore, more recent studies have investigated PDE10A as a therapeutic target for treating schizophrenia and HD (CitationHebb et al 2004; CitationSiuciak et al 2006). However, despite the substantial data to support the role of PDEs in learning and memory, so far only the only reported studies for the clinical use of phosphodiesterase inhibitors, has been in the treatment of depression using rolipram (reviewed by CitationZhu et al 2001; CitationRenau 2004).
In zebrafish, 2’,3’-cyclic-nucleotide 3’- phosphodiesterase was first reported as being induced in an optic nerve regeneration study (CitationBallestero et al 1999). Further searches of the National Center for Biotechnology Information (NCBI) protein sequence finder revealed zebrafish possess proteins with similarity to the phosphodiesterases 1, 3–7, 9–11. Two enzymes of primary interest are the PDE4 (http://www.ncbi.nlm.nih.gov/BLAST, accession number: CAK10806), with an identity to the human PDE4C1 of 63%, rat 4D 67% and mouse 4C 61%, and PDE10A (http://www.ncbi.nlm.nih.gov/BLAST, accession number: NP 957396), which has an identity with the human, rat, and mouse PDE10A of 83% (for summary see ). Specific orthologues of the other splice forms may also exist but have yet to be reported. As yet, little has been reported in zebrafish as to the effects of PDE inhibitors on learning and memory; though this lab has reported that rolipram enhances the acoustic startle response in 7 d.p.f. larval zebrafish and delays habituation to consecutives tones (CitationBest et al 2007). These effects in zebrafish are similar to those seen in previous rat studies (CitationKehne et al 1991), suggesting the presence of functional phosphodiesterase 4 enzymes or at the very least high-affinity rolipram binding sites. Clearly more investigations are needed to elucidate the distribution of PDEs in fish and their role in fish behavior and cognition.
Table 1 Homology of the rat and zebrafish with the human receptors which have been associated with learning and memory
Histamine 3 receptor antagonists
The histamine 3 (H3) receptor is a G protein coupled receptor identified in the 1980s and pursued as a drug target for a number of indications (for reviews see CitationWitkin and Nelson 2004; CitationEsbenshade et al 2006; CitationBonaventure et al 2007). H3 receptors are centrally located with mRNA distributed in regions connected to memory and learning, such as the hippocampus and cortex (CitationLovenberg et al 1999). In recombinant receptor systems H3 receptors have been shown to have constitutive activity (reviewed by CitationArrang et al 2007) and research has centred on investigating inverse agonists/ antagonists for cognitive disorders. Thioperamide has been used to examine the effect of H3 receptor antagonists on learning and memory and clobenpropit has been shown to reverse scopolamine-induced learning deficits as well as increasing social memory, attention and inhibitory avoidance. However, thioperamide and clobenpropit impair fear conditioning suggesting H3 antagonists may be specific in their role in mediating cognition (CitationPassani et al 2004; CitationWitkin and Nelson 2004; CitationEsbenshade et al 2006; CitationBonaventure et al 2007). A number of pharmaceutical companies have programs based around H3 antagonists with clinical candidates reported in Phase I (see CitationEsbenshade et al 2006).
Three histamine receptors H1, H2, and H3 have been cloned and expressed in zebrafish (CitationPeitsaro et al 2007). When compared with the distribution in mammalian brain, the histaminergic neurons are more tightly located around the posterior recess and are more ventral to the mammillary body than in rodents. However, the rostrocaudal distribution is very similar to that seen in the rat and overall the projections patterns are highly conserved (CitationKaslin and Panula 2001). Comparison of the H3 receptor peptide sequence showed 50% identity with the human (see ) (CitationPeitsaro et al 2007), and binding studies demonstrated the H3 receptor was expressed throughout the zebrafish brain with the greatest intensity in the optic tectum and hypothalamus (CitationPeitsaro et al 2000). Zebrafish at 5 d.p.f. treated with thioperamide (100 μM) demonstrated a decrease in locomotor activity although no toxicity assessment was given to determine whether this decrease was an effect was specific antagonism or a reduction due to generalised toxicity. Other studies in goldfish have demonstrated involvement of H1 and H2 in inhibitory avoidance (CitationCofiel and Mattioli 2006).
5HT6 receptor antagonists
5-Hydoxytryptamine (5-HT) receptors so far are composed of seven different subtypes which have been implicated in mediating cognition (CitationMeneses 1999, Citation2007). They are all found in areas of the brain connected with learning and memory, but attention has focussed on 5-HT6, which appears to have a greater role in long-term memory (for review see CitationMitchell and Neumaier 2005). A number of antagonists (eg, RO4368554, SB-271046) have been shown to reverse scopolamine-induced deficits in passive avoidance assays in rats as well as enhancing object recognition (CitationFoley et al 2004; CitationMitchell and Neumaier 2005; CitationHirst et al 2006; CitationSchreiber et al 2007), although the consistency of the effect at this target has been disputed, which raises questions as to brain penetrance and selectivity of the various compounds used (CitationRussell and Dias 2002). Again, despite the literature supporting their effect in learning and memory, there are currently no reports of clinical trials for 5HT6 antagonists.
In zebrafish, 5-HT neurons are expressed in the spinal cord as early as 1 d.pf. with populations in the telencephalon, hindbrain and throughout the brain by 5 d.p.f. (CitationDrapeau et al 2002; CitationMcLean and Fetcho 2004). In the adult these neurons cluster in the area of the hypothalamus and habenula (CitationKaslin and Panula 2001). A search of the NCBI protein sequence finder database and blast search shows that zebrafish have at least three recognised 5-HT receptors namely 1, 2 and 7 (http://www.ncbi.nlm.nih.gov/BLAST). This has been confirmed with studies using methysergide a 5-HT1,2,7 antagonist which modulated locomotor activity in larval zebrafish (CitationDrapeau et al 2002; CitationBrustein and Drapeau 2005). Studies in goldfish have demonstrated the presence of 5-HT1A in the retina (CitationSchmeer et al 2001). Interestingly, a blast search using the human 5-HT6 peptide sequence revealed 54% identity with a hypothetical protein (accession number: XP 696681) which also has 52% and 50% identity with the rat and mouse 5-HT6 sequences respectively (see ).
AMPA receptor potentiators
Alpha-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid (AMPA) receptors are glutamate receptors comprising four different subunits GluRA1-A4, are found throughout the CNS mediating fast excitatory transmission (for a comprehensive review see CitationO’Neill et al 2004). Early studies have shown the presence of multiple AMPA receptors in the CA1/CA2 region of the hippocampus suggesting this as a target for cognition (CitationWenthold et al 1996). Ampakines are a diverse class of compounds which positively modulate glutamateric AMPA receptors with pyrrolidones (eg, piracetam) and piperidines (CX516) being examples (see CitationO’Neill et al 2004; CitationO’Neill and Dix 2007). These compounds have been studied in a number of different rodent memory tasks such as spatial memory, delayed-nonmatch-to sample and extinction learning and have been shown to improve performance (CitationStaubli et al 1994; CitationHampson et al 1998; CitationZushida et al 2007). However, despite some early encouraging data (CitationLynch and Gall 2006), recent clinical results have been mixed with the ampakine farampator increasing short-term memory trial of the symbol digit recall test in elderly patients (CitationWezenberg et al 2007) whereas CX516 used as an add-on therapy did not alter the composite cognitive score compared to the placebo group in schizophrenic patients (CitationGoff et al 2007) or improve cognitive outcomes in patients with fragile X syndrome (CitationBerry-Kravis et al 2006) suggesting these compounds may not have been dosed adequately or that they need to be used more selectively for cognitive impairments.
Zebrafish possess the AMPA receptors subunits GluR1-4 with each one consisting of a subtype A and B (NCBI protein database; http://www.ncbi.nlm.nih.gov/sites/entrez). A blast search between the subtypes shows a similarity between the A and B subtypes ranges from 82% for GluR1, 88% GluR3 to 92% for GluR2 and 4. Compared with human, rat, and mouse, the zebrafish receptors have very good identity, the 1A subtype has 71% identity with the human, rat, and mouse GluR1 sequences. The GluR2-4A receptors have higher identities with human, rat and mouse of between 85%–90% with the equivalent protein sequence (see ). AMPA receptors have been found and studied in the retina, hindbrain, spinal cord, and Mauthner neurons of zebrafish (CitationAli et al 2000; CitationYazulla and Studholme 2001; CitationPatten and Ali 2007). They are also associated with the neuromuscular junction in zebrafish (CitationTodd et al 2004). Not much has been reported in the zebrafish literature as to the affect of AMPAkines on their learning and memory; however studies on nonassociative learning in this lab investigating piracetam have found that it increased the ASR and prolonged the habituation (unpublished data) similar to the other cognition enhancers tested.
Conclusion
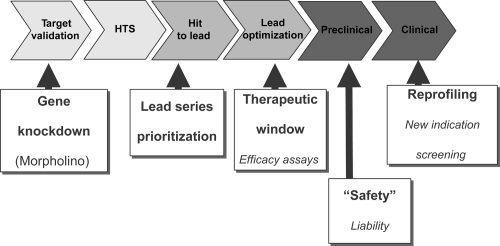
It is now recognised that zebrafish possess a great deal of similarity to mammals and are an extremely useful model for screening compounds at several stages of the drug discovery process (see ). As more is reported on the efficacy models in zebrafish its utility as an early stage screening tool for the discovery of CNS therapeutics should enable increased throughput of in vivo analysis of novel compounds for neurological disorders. In this review we have described the application of the zebrafish models to neurodegenerative disorders, schizophrenia and learning and memory. Several of these diseases affect an aging population and are adult onset raising questions as to the appropriateness of using a rapidly developing larval system. However, early phenotypes are observable and in comparison with rodents the zebrafish larvae are not foetal but are closer to juveniles in that the nervous system is mature, organs are functioning and tissue architecture is fully developed by the time at which many of the assays are performed. In this review we have also described four validated pharmacological targets which are being investigated preclinically for impaired learning and memory; phosphodiesterases, histamine 3, 5HT-6 and AMPA, and have illustrated how zebrafish may be used in the assessment of these targets. As more information is reported on neurological assays in zebrafish, the utility of this model organism as an early stage screening tool for CNS disorders should help increase in vivo throughput and ameliorate the cost associated with drug screening in mammals.
Figure 1 Utility of zebrafish in a screening and drug discovery strategy. Flow diagram showing where zebrafish can aid the drug development and discovery process but either increasing throughput or assessing candidate compounds more economically.
References
- AbeliovichABealMFParkinsonism genes: culprits and cluesJ Neurochem20069910627216836655
- AliDWBussRRDrapeauPProperties of miniature glutamatergic EPSCs in neurons of the locomotor regions of the developing zebrafishJ Neurophysiol2000831819110634865
- AnichtchikOVKaslinJPeitsaroNNeurochemical and behavioural changes in zebrafish Danio rerio after systemic administration of 6-hydroxydopamine and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridineJ Neurochem2004884435314690532
- ArrangJMMorissetSGbahouFConstitutive activity of the histamine H(3) receptorTrends Pharmacol Sci2007124913951
- ArthurDLevinEDSpatial and non-spatial visual discrimination learning in zebrafish (Danio rerio)Animal Cognition2001412531
- BallesteroRPDybowskiJALevyGCloning and characterization of zRICH, a 2’,3’-cyclic-nucleotide 3’-phosphodiesterase induced during zebrafish optic nerve regenerationJ Neurochem19997213627110098837
- BaradMBourtchouladzeRWinderDGRolipram, a type IV-specific phosphodiesterase inhibitor, facilitates the establishment of long-lasting long-term potentiation and improves memoryProc Natl Acad Sci USA1998951502059844008
- BergerJCurriePThe role of zebrafish in chemical geneticsCurr Med Chem20071424132017896989
- BerghmansSButlerBGoldsmithPEarly drug safety assessment for the cardiac, visual and gut functions using the zebrafishJ Pharmacol Toxicol Methods2008in press
- Berry-KravisEKrauseSEBlockSSEffect of CX516, an AMPA-modulating compound, on cognition and behavior in fragile X syndrome: a controlled trialJ Child Adolesc Psychopharmacol2006165254017069542
- BestJDBerghmansSHuntJJNon-associative learning in larval zebrafishNeuropsychopharmacology2007620 Epub ahead of print
- BirksJCholinesterase inhibitors for Alzheimer’s diseaseCochrane Database Syst Rev20061CD00559316437532
- BonaventurePLetavicMDugovicCHistamine H3 receptor antagonists: from target identification to drug leadsBiochem Pharmacol20077310849617129577
- BraffDLGeyerMASwerdlowNRHuman studies of prepulse inhibition of startle: normal subjects, patient groups, and pharmacological studiesPsychopharmacology (Berl)20011562345811549226
- BretaudSAllenCInghamPWp53-dependent neuronal cell death in a DJ-1-deficient zebrafish model of Parkinson’s diseaseJ Neurochem200710016263517166173
- BretaudSLeeSGuoSSensitivity of zebrafish to environmental toxins implicated in Parkinson’s diseaseNeurotoxicol Teratol2004268576415451049
- BrusteinEDrapeauPSerotoninergic modulation of chloride homeostasis during maturation of the locomotor network in zebrafishJ Neurosci200525106071616291933
- BurgessHAGranatoMSensorimotor gating in larval zebrafishJ Neurosci20072749849417475807
- BurnsARossorMHeckerJThe effects of donepezil in Alzheimer’s disease – results from a multinational trialDement Geriatr Cogn Disord1999102374410325453
- CahillGMClock mechanisms in zebrafishCell Tissue Res2002309273412111534
- CarvanMJ3rdLoucksEWeberDNEthanol effects on the developing zebrafish: neurobehavior and skeletal morphogenesisNeurotoxicol Teratol2004267576815451040
- ChenLCagniardBMathewsTAge-dependent motor deficits and dopaminergic dysfunction in DJ-1 null miceJ Biol Chem2005280214182615799973
- CherryJADavisRLCyclic AMP phosphodiesterases are localized in regions of the mouse brain associated with reinforcement, movement, and affectJ Comp Neurol199940728730110213096
- CofielLPMattioliRInvolvement of histamine receptors in the acquisition of inhibitory avoidance in Carassius auratusProg Neuropsychopharmacol Biol Psychiatry20063012465016631292
- ColwillRMRaymondMPFerreiraLVisual discrimination learning in zebrafish (Danio rerio)Behav Processes200570193115967284
- CosmanKMBoyleLLPorsteinssonAPMemantine in the treatment of mild-to-moderate Alzheimer’s diseaseExpert Opin Pharmacother200782031417257090
- CowanCMRaymondLASelective neuronal degeneration in Huntington’s diseaseCurr Top Dev Biol200675257116984809
- DevanBDSierra-MercadoDJrJimenezMPhosphodiesterase inhibition by sildenafil citrate attenuates the learning impairment induced by blockade of cholinergic muscarinic receptors in ratsPharmacol Biochem Behav200479691915582676
- DrapeauPSaint-AmantLBussRRDevelopment of the locomotor network in zebrafishProg Neurobiol2002688511112450489
- EsbenshadeTAFoxGBCowartMDHistamine H3 receptor antagonists: preclinical promise for treating obesity and cognitive disordersMol Interv2006677885916565470
- FoleyAGMurphyKJHirstWDThe 5-HT(6) receptor antagonist SB-271046 reverses scopolamine-disrupted consolidation of a passive avoidance task and ameliorates spatial task deficits in aged ratsNeuropsychopharmacology2004299310014571256
- GalvinJECognitive change in Parkinson diseaseAlzheimer Dis Assoc Disord2006203021017132978
- GauthierSReisbergBZaudigMMild cognitive impairmentLancet200636712627016631882
- GeertsHGrossbergGTPharmacology of acetylcholinesterase inhibitors and N-methyl-D-aspartate receptors for combination therapy in the treatment of Alzheimer’s diseaseJ Clin Pharmacol2006468S16S16809810
- GoffDCLambertiJSLeonACA Placebo-Controlled Add-On Trial of the Ampakine, CX516, for Cognitive Deficits in SchizophreniaNeuropsychopharmacology2007
- Goldsmith P, Fleming A (2007). Screening methods employing zebrafish and the blood brain barrier. IN OFFICE, E. P (Ed.).
- GrunwaldDJEisenJSHeadwaters of the zebrafish – emergence of a new model vertebrateNat Rev Genet200237172412209146
- GuoSLinking genes to brain, behavior and neurological diseases: what can we learn from zebrafish?Genes Brain Behav20043637415005714
- HampsonRERogersGLynchGFacilitative effects of the ampakine CX516 on short-term memory in rats: enhancement of delayed-nonmatch-to-sample performanceJ Neurosci199818274079502831
- HebbALRobertsonHARole of phosphodiesterases in neurological and psychiatric diseaseCurr Opin Pharmacol20077869217113826
- HebbALRobertsonHADenovan-WrightEMStriatal phosphodiesterase mRNA and protein levels are reduced in Huntington’s disease transgenic mice prior to the onset of motor symptomsNeuroscience20041239678114751289
- HirstWDSteanTORogersDCSB-399885 is a potent, selective 5-HT6 receptor antagonist with cognitive enhancing properties in aged rat water maze and novel object recognition modelsEur J Pharmacol20065531091917069795
- ImanishiTSawaAIchimaruYAmeliorating effects of rolipram on experimentally induced impairments of learning and memory in rodentsEur J Pharmacol199732127389085037
- KarlovichCAJohnRMRamirezLCharacterization of the Huntington’s disease (HD) gene homologue in the zebrafish Danio rerioGene1998217117259795179
- KaslinJPanulaPComparative anatomy of the histaminergic and other aminergic systems in zebrafish (Danio rerio)J Comp Neurol20014403427711745628
- KehneJHBoulisNMDavisMEffects of the phosphodiesterase inhibitor rolipram on the acoustic startle response in ratsPsychopharmacology (Berl)199110527361660609
- LamCSKorzhVStrahleUZebrafish embryos are susceptible to the dopaminergic neurotoxin MPTPEur J Neurosci20052117586215845104
- LeroyEBoyerRAuburgerGThe ubiquitin pathway in Parkinson’s diseaseNature199839545129774100
- LeveyALahJGoldsteinFMild cognitive impairment: an opportunity to identify patients at high risk for progression to Alzheimer’s diseaseClin Ther200628991100116990077
- LevinEDChenENicotinic involvement in memory function in zebrafishNeurotoxicol Teratol200426731515451037
- LevinEDLimpuangthipJRachakondaTTiming of nicotine effects on learning in zebrafishPsychopharmacology (Berl)2006a1845475216175402
- LevinEDMcClernonFJRezvaniAHNicotinic effects on cognitive function: behavioral characterization, pharmacological specification, and anatomic localizationPsychopharmacology (Berl)2006b1845233916220335
- LevinEDSimonBBNicotinic acetylcholine involvement in cognitive function in animalsPsychopharmacology (Berl)1998138217309725745
- LovenbergTWRolandBLWilsonSJCloning and functional expression of the human histamine H3 receptorMol Pharmacol1999551101710347254
- LumsdenALHenshallTLDayanSHuntingtin-deficient zebrafish exhibit defects in iron utilization and developmentHum Mol Genet2007
- LuqueMAPerez-PerezMPHerreroLInvolvement of the optic tectum and mesencephalic reticular formation in the generation of saccadic eye movements in goldfishBrain Res Rev2005493889716111565
- LynchGGallCMAmpakines and the threefold path to cognitive enhancementTrends Neurosci2006295546216890999
- MacRaeCAPetersonRTZebrafish-based small molecule discoveryChem Biol200310901814583256
- McDearmidJRDrapeauPRhythmic motor activity evoked by NMDA in the spinal zebrafish larvaJ Neurophysiol2006954011716207779
- McKinleyETBaranowskiTCBlavoDONeuroprotection of MPTP-induced toxicity in zebrafish dopaminergic neuronsBrain Res Mol Brain Res20051411283716209898
- McLeanDLFetchoJROntogeny and innervation patterns of dopaminergic, noradrenergic, and serotonergic neurons in larval zebrafishJ Comp Neurol2004480385615515022
- MenesesA5-HT system and cognitionNeurosci Biobehav Rev19992311112510643820
- MenesesADo serotonin(1–7) receptors modulate short and long-term memory?Neurobiol Learn Mem2007875617217224282
- MillerVMNelsonRFGouvionCMCHIP suppresses polyglutamine aggregation and toxicity in vitro and in vivoJ Neurosci20052591526116207874
- MitchellESNeumaierJF5-HT6 receptors: a novel target for cognitive enhancementPharmacol Ther20051083203316005519
- MontoyaAPriceBHMenearMBrain imaging and cognitive dysfunctions in Huntington’s diseaseJ Psychiatry Neurosci20063121916496032
- MountCDowntonCAlzheimer disease: progress or profit?Nat Med200612780416829947
- MudherALovestoneSAlzheimer’s disease-do tauists and baptists finally shake hands?Trends Neurosci20022522611801334
- MurpheyRDZonLISmall molecule screening in the zebrafishMethods2006392556116877005
- MusaALehrachHRussoVADistinct expression patterns of two zebrafish homologues of the human APP gene during embryonic developmentDev Genes Evol2001211563711862463
- NamRHKimWLeeCJNMDA receptor-dependent long-term potentiation in the telencephalon of the zebrafishNeurosci Lett20043702485115488332
- NewmanMMusgraveFILardelliMAlzheimer disease: amyloidogenesis, the presenilins and animal modelsBiochim Biophys Acta200717722859717208417
- NinkovicJFolchertAMakhankovYVGenetic identification of AChE as a positive modulator of addiction to the psychostimulant D-amphetamine in zebrafishJ Neurobiol2006664637516470869
- NuechterleinKHBarchDMGoldJMIdentification of separable cognitive factors in schizophreniaSchizophr Res200472293915531405
- O’NeillMJBleakmanDZimmermanDMAMPA receptor potentiators for the treatment of CNS disordersCurr Drug Targets CNS Neurol Disord200431819415180479
- O’NeillMJDixSAMPA receptor potentiators as cognitive enhancersIDrugs2007101859217351873
- OrgerMBGahtanEMutoABehavioral screening assays in zebrafishMethods Cell Biol200477536815602905
- PaquetDSchmidBHaassCAnalysis of the function of the Parkinson’s disease gene pink1 in zebrafish20067th International Conference on Zebrafish Development and GeneticsJune 14–18University of Wisconsin-Madison, Madison, Wisconsin USA
- PassaniMBLinJSHancockAThe histamine H3 receptor as a novel therapeutic target for cognitive and sleep disordersTrends Pharmacol Sci2004256182515530639
- PattenSAAliDWAMPA receptors associated with zebrafish Mauthner cells switch subunits during developmentJ Physiol200758110435617412769
- PeitsaroNAnichtchikOVPanulaPIdentification of a histamine H(3)-like receptor in the zebrafish (Danio rerio) brainJ Neurochem2000757182410899947
- PeitsaroNKaslinJAnichtchikOVModulation of the histaminergic system and behaviour by alpha-fluoromethylhistidine in zebrafishJ Neurochem2003864324112871584
- PeitsaroNSundvikMAnichtchikOVIdentification of zebrafish histamine H1, H2 and H3 receptors and effects of histaminergic ligands on behaviorBiochem Pharmacol20077312051417266939
- PostlethwaitJHWoodsIGNgo-HazelettPZebrafish comparative genomics and the origins of vertebrate chromosomesGenome Res200010189090211116085
- PradelGSchmidtRSchachnerMInvolvement of L1.1 in memory consolidation after active avoidance conditioning in zebrafishJ Neurobiol20004338940310861564
- PrickaertsJvan StaverenWCSikAEffects of two selective phosphodiesterase type 5 inhibitors, sildenafil and vardenafil, on object recognition memory and hippocampal cyclic GMP levels in the ratNeuroscience20021133516112127092
- RenauTEThe potential of phosphodiesterase 4 inhibitors for the treatment of depression: opportunities and challengesCurr Opin Investig Drugs20045349
- RingmanJMCummingsJLCurrent and emerging pharmacological treatment options for dementiaBehav Neurol20061751616720956
- RinkEWullimannMFConnections of the ventral telencephalon (subpallium) in the zebrafish (Danio rerio)Brain Res200410112062015157807
- RobinsonDMKeatingGMMemantine: a review of its use in Alzheimer’s diseaseDrugs20066615153416906789
- RoseGMHopperADe VivoMPhosphodiesterase inhibitors for cognitive enhancementCurr Pharm Des20051133293416250839
- RussellMGDiasRMemories are made of this (perhaps): a review of serotonin 5-HT(6) receptor ligands and their biological functionsCurr Top Med Chem200226435412052198
- SalzmannJAnichtchikOVBestJDEffect of MPTP treatment on locomotor activity and dopaminergic cells in the zebrafish larvae2006Workshop – Neurodegeneration; basic mechanisms of motor and cognitive dysfunctionsSeptember 2–4Cracow, Poland
- SchifferNWBroadleySAHirschbergerTIdentification of anti – prion compounds as efficient inhibitors of polyglutamine protein aggregation in a zebrafish modelJ Biol Chem2007282919520317170113
- SchmeerCObregonFUrbinaMFurther characterization of 5-HT1A receptors in the goldfish retina: role of cyclic AMP in the regulation of the in vitro outgrowth of retinal explantsNeurochem Res2001262132311495544
- SchreiberRVivianJHedleyLEffects of the novel 5-HT(6) receptor antagonist RO4368554 in rat models for cognition and sensorimotor gatingEur Neuropsychopharmacol2007172778816989988
- SelkoeDJToward a comprehensive theory for Alzheimer’s disease. Hypothesis: Alzheimer’s disease is caused by the cerebral accumulation and cytotoxicity of amyloid beta-proteinAnn N Y Acad Sci2000924172511193794
- ShankaranSSchmidBKahlePFunctional Analysis of Parkinson’s Disease Gene LRRK2 in Zebrafish20067th International Conference on Zebrafish Development and GeneticsJune 14–18University of Wisconsin-Madison, Madison, Wisconsin USA
- SiuciakJAChapinDSHarmsJFInhibition of the striatum-enriched phosphodiesterase PDE10A: a novel approach to the treatment of psychosisNeuropharmacology2006513869616780899
- SmeyneRJJackson-LewisVThe MPTP model of Parkinson’s diseaseBrain Res Mol Brain Res2005134576615790530
- SonOLKimHTJiMHCloning and expression analysis of a Parkinson’s disease gene, uch-L1, and its promoter in zebrafishBiochem Biophys Res Commun2003312601714680807
- StaubliURogersGLynchGFacilitation of glutamate receptors enhances memoryProc Natl Acad Sci USA199491777818290599
- SwainHASigstadCScalzoFMEffects of dizocilpine (MK-801) on circling behavior, swimming activity, and place preference in zebrafish (Danio rerio)Neurotoxicol Teratol200426725915451036
- SwerdlowNRPaulsenJBraffDLImpaired prepulse inhibition of acoustic and tactile startle response in patients with Huntington’s diseaseJ Neurol Neurosurg Psychiatry1995581922007876851
- TammingaCAThe neurobiology of cognition in schizophreniaJ Clin Psychiatry200667e1117081078
- ToddKJSlatterCAAliDWActivation of ionotropic glutamate receptors on peripheral axons of primary motoneurons mediates transmitter release at the zebrafish NMJJ Neurophysiol2004918284014762151
- TomasiewiczHGFlahertyDBSoriaJPTransgenic zebrafish model of neurodegenerationJ Neurosci Res2002707344512444595
- TropepeVSiveHLCan zebrafish be used as a model to study the neurodevelopmental causes of autism?Genes Brain Behav200322688114606692
- VandenbergheRTournoyJCognitive aging and Alzheimer’s diseasePostgrad Med J2005813435215937198
- WentholdRJPetraliaRSBlahosJIIEvidence for multiple AMPA receptor complexes in hippocampal CA1/CA2 neuronsJ Neurosci199616198298604042
- WezenbergEVerkesRJRuigtGSAcute effects of the ampakine farampator on memory and information processing in healthy elderly volunteersNeuropsychopharmacology20073212728317119538
- WilliamsFEWhiteDMesserWSA simple spatial alternation task for assessing memory function in zebrafishBehav Processes2002581253212044689
- WitkinJMNelsonDLSelective histamine H3 receptor antagonists for treatment of cognitive deficiencies and other disorders of the central nervous systemPharmacol Ther200410312015251226
- WullimannMFMuellerTTeleostean and mammalian forebrains contrasted: Evidence from genes to behaviorJ Comp Neurol20044751436215211457
- XuXScott-ScheiernTKempkerLActive avoidance conditioning in zebrafish (Danio rerio)Neurobiol Learn Mem20078772716861014
- YazullaSStudholmeKMNeurochemical anatomy of the zebrafish retina as determined by immunocytochemistryJ Neurocytol2001305519212118162
- ZhangHTCrissmanAMDorairajNRInhibition of cyclic AMP phosphodiesterase (PDE4) reverses memory deficits associated with NMDA receptor antagonismNeuropsychopharmacology20002319820410882846
- ZhangHTO’DonnellJMEffects of rolipram on scopolamine-induced impairment of working and reference memory in the radial-arm maze tests in ratsPsychopharmacology (Berl)20001503111610923759
- ZhdanovaIVWangSYLeclairOUMelatonin promotes sleep-like state in zebrafishBrain Res2001903263811382414
- ZhuJMixEWinbladBThe antidepressant and antiinflammatory effects of rolipram in the central nervous systemCNS Drug Rev200173879811830756
- ZonLIPetersonRTIn vivo drug discovery in the zebrafishNat Rev Drug Discov20054354415688071
- ZushidaKSakuraiMWadaKFacilitation of extinction learning for contextual fear memory by PEPA: a potentiator of AMPA receptorsJ Neurosci2007271586617202483