Abstract
Recent studies have suggested that some variants of bipolar disorder (BD) may be due to hyperconnectivity between orbitofrontal (OFC) and temporal pole (TP) structures in the dominant hemisphere. Some initial MRI studies noticed that there were corpus callosum abnormalities within specific regional areas and it was hypothesized that developmentally this could result in functional or effective connectivity changes within the orbitofrontal-basal ganglia-thalamocortical circuits. Recent diffusion tensor imaging (DTI) white matter fiber tractography studies may well be superior to region of interest (ROI) DTI in understanding BD. A “ventral semantic stream” has been discovered connecting the TP and OFC through the uncinate and inferior longitudinal fasciculi and the elusive TP is known to be involved in theory of mind and complex narrative understanding tasks. The OFC is involved in abstract valuation in goal and sub-goal structures and the TP may be critical in binding semantic memory with person–emotion linkages associated with narrative. BD patients have relative attenuation of performance on visuoconstructional praxis consistent with an atypical localization of cognitive functions. Multiple lines of evidence suggest that some BD alleles are being selected for which could explain the enhanced creativity in higher-ability probands. Associations between ROI’s that are not normally connected could explain the higher incidence of artistic aptitude, writing ability, and scientific achievements among some mood disorder subjects.
Introduction
There are reliable differences in regional callosal area across subjects with specific relationships to the organization of linguistic processes. Normal limit differences in width or regional fiber composition of the corpus callosum do have predictable effects on the patterns of inter- and intra-hemispheric of communication and hence in the transfer of information throughout the cerebral cortex (CitationClarke and Zaidel 1994). Developmentally speaking, thickness or thinning of specific regions of the callosum could accentuate the canalization of specific white matter pathways both interhemispherically and intrahemispherically. Between subject differences in the variance of regional fiber composition could even result in unique co-configurations of white matter tracts in men and women. Bipolar disorder has long been known to be concordant with either regionally reduced white matter density or more recently hyperconnectivity between specific regions of the human brain. However only recently has the disorder been reconceptualized theoretically as a complex disorder of connectivity.
In the cognitive neuropsychiatry and cognitive neuropsychology literature it has been shown that electrophysiological synchronous patterns of activation can coordinate up to four sensory regional modules in a single cyclic 100 millisecond reverbatory whole-brain circuit linking frontal and posterior cortical networks (CitationSinger 2004). In normal individuals, language and praxis are co-localized to the left hemisphere with the left and right hemispheres sensitive to abstract and concrete concepts, respectively. In contrast in atypical language localized subjects language and praxis may be localized to the right and left hemispheres, respectively (CitationKnecht et al 2003). In some of these individuals, abstract concepts may show a preponderance of association with the right hemisphere and concomitantly, concrete concepts will be largely processed with left hemisphere networks (CitationBinder et al 2005).
In certain variants of bipolar disorder in susceptible individuals, the atypical co-localization of praxis and concrete concepts could function as a highly sensitive mapping mechanism in concert with the right hemisphere’s abstract linguistic mapping function. However, this is only one of many possible subtypes. For our current purposes, we will focus on recent neuroanatomical studies that have clear theoretical implications for explaining the disorder with a particular emphasis on the “ventral semantic stream”. Convergent findings across disciplines implicate this recently articulated white matter pathway in the etiology of bipolar disorder.
However not all bipolar disorder subjects show atypical patterns of cortical localization. Bipolar disorder is reliably associated with higher-order impairments fundamentally presenting as dysregulated mood homeostatic mechanisms. These dysregulated homeostatic mood mechanisms originate in the prefrontal to the limbic to the reticular formation top-down afferents. As a result, white or gray matter deficiencies anywhere in the distributed prefrontal neural networks will result in impairments in functional or effective connectivity such that the disorder can be conceived of as the end result of any combination of structural brain abnormalities. Such sophisticated system views of top-down prefrontal mediated networks could well benefit from some of the third generation computational modeling techniques that use both diffusion tensor imaging fiber tracking (eg, tractography) and vector autoregressive modeling techniques (CitationGoebel et al 2004).
Indeed, recently just such approaches have left the theoretical cognitive neuroscience laboratory and entered neuropsychiatric research and practice. That bipolar disorder etiology is extremely heterogeneous yet convergent in resultant symptomologic profile suggests dysfunction in specific neuropsychiatric functional systems. Already, genetic linkage studies have identified 10 regions of the human genome that carry with it susceptibility to development of bipolar disorder (CitationBerrittini 2006). However for such a serious mental illness it is difficult to explain the maintenance of these harmful alleles. One hypothesis would be that atypical co-localization of functions confers on unique individuals enhanced signal-noise ratios in the sequestration of praxis and object-based systems to the left hemisphere. The idea that atypical co-localization of unique combinations of modules in a hemisphere could confer on anomalous individuals emergent properties or an ability is a serious possibility and the idea is not new (CitationBasso et al 1985).
If sequestration of praxis and object-based systems to the left hemisphere enhances the accuracy, as opposed to speed, of motor response then evolutionary mechanisms must have led to the selection for such a trait. Recent studies supporting the existence of both a direct and indirect ventral semantic stream sensitive to semantic paraphasias but not to phonemic paraphasias is congruent with this hypothesis (CitationDuffau et al 2005). Classical lesion studies also have shown that left-sided lesions of the frontal cortex and basal ganglia are apt to cause organic depression whereas euphoria and mania can be triggered by right-sided basal frontotemporal and subcortical damage (CitationMayberg 2006). These studies to be reviewed suggest any number of combinations of intercommissural inhibition or disinhibition or excitatory mechanisms could be operative.
Moreover, post-injury studies are congruent with the commissural fiber hypothesis that developmentally speaking large scale compensatory modulation of functional neural networks occurs. Similarly, organic depression or mania can result after specific cortical or subcortical lesions. Finally the question must be asked why would such a serious mental illness that is so detrimental persist in the population with such high basal allelic frequencies? Evolutionary hypotheses suggest that low mood could possibly confer adaptive traits under ancestral conditions by increasing the probability of highly realistic assessments pertaining to disengagement or flexibility from salient goal objectives of an individual in rapidly changing environments. This valuation and re-valuation of goals and sub-goal structures will be reviewed here in terms of functional neuroanatomy.
These neuropsychologic and evolutionary studies suggest that for some subtypes of bipolar disorder the functional neuroanatomic organizational principles of the cerebral cortex could function quite differently. Moreover, these distinct patterns of cortical organization in susceptible individuals might be particularly apt to benefit the accuracy of semantic-motor associations, which confer adaptive motor performance, yet also might increase the prevalence of the frequency of deleterious mood-instability traits in kinship groups. In other words, some individuals with certain variants of bipolar disorder would be expected to possess atypical, beneficial, highly developed, selected for cognitive abilities (eg, writing, artistic aptitude or creativity) that could explain the maintenance of highly deleterious endophenotypes which reduce life-expectancy at the single subject level.
Bipolar disorder, corpus callosum, and diffusion tensor imaging
It is hypothesized that bipolar disorder subjects have an associated cognitive endophenotype that presumably has evolved through beneficial traits conferred under ancestral conditions (CitationBerrittini 2006). Moreover functional imaging studies have suggested that there are laterality effects in terms of the localization of the functional neuroanatomical substrates of mood homeostatic mechanisms in bipolar disorder patients (CitationMayberg 2006). Also early comprehensive studies of atypically cognitively localized subjects were viewed as essential in understanding the deep principles by which language is represented in the brain (CitationBasso et al 1985). The same hypothesis would also hold true for atypically localized bipolar disorder subjects. Finally if the processing of language attributes such as concreteness and abstractness of words is lateralized (CitationBinder et al 2005), and atypically localized subjects thus have different patterns of localization of modules (CitationSinger 2004; CitationKnecht et al 2003) then differences in anatomical structures such as the callosum could canalize the development of atypical white matter tracts in anomalous individuals (CitationClarke and Zaidel 1994; CitationGoebel et al 2004; CitationDuffau et al 2005). Hence we will review recent diffusion tensor imaging studies in bipolar disorder patients to show evidence of how the specific canalization of white matter tracts occurs with development.
Thirty-three bipolar disorder subjects were compared with 40 normal healthy age, gender, SES and ethnicity matched cohorts. Age and gender were found to be nonsignificant covariates for fractional anisotropy (FA) and were dropped from further analysis (CitationWang et al 2008). Fractional anisotropy is a measure of the degree to which white matter fibers in the human brain are aligned in a specific direction within a three-dimensional space in the cerebral cortex. There were no significant correlations between clinical factors with the bipolar disorder groups and FA (eg, rapidity of cycling, mood state, medication status, history of substance abuse or co-morbidities). Only 9% or 3 subjects had co-morbidities and thus for all intents and purposes this was an undifferentiated bipolar disorder sample. FA was lowest in the mid-body of the corpus callosum in bipolar subjects, compared to normal subjects (p < 0.001) and also significantly lower in the genu in bipolar subjects; although of an order of magnitude less (p < 0.01). When average FA values were examined in the mid-body of the corpus callosum bipolar subjects’s FA values were approximately (FA = 0.4). In healthy subjects, in the anterior or posterior regions of the corpus callosum FA values were approximately 0.5. This suggests that most of the variance in FA was occurring within the mid-body of the corpus callosum.
Recent studies have suggested that subcortical hyper-intensities are not found in all bipolar disorder subjects, nor are prevalences or severity of these focal white matter lesions significantly different from that of healthy subjects (CitationStrakowski et al 1993). This implies that callosal abnormalities may figure more prominently in these mood disorders (CitationYurgelun-Todd et al 2007). Incidentally, there was significantly higher fractional anisotropy in bipolar disorder subjects in the medial aspect of the genu compared to normal controls in spite of the small sample size (effect size = 0.48). Of note was that normal healthy subjects demonstrated higher axonal integrity in the splenium compared with the genu whereas the bipolar patients showed similar FA values across both areas. Hence, region of interest and voxel-based comparisons of diffusion tensor studies of bipolar disorder subjects have been confusing with some studies showing higher or lower FA values in the anterior cingulate dependent on specific methodology or statistic used.
Moreover, the splenium has been shown to possess larger diameter axonal fibers and less unmyelinated small diameter fibers than the genu (CitationAboitiz et al 1992) which could account for this anterior-posterior dichotomy. However, there is controversy over whether these differences are artifacts of relatively greater connectivity within frontal lobes (eg, oblique fibers) in the FA matrix (CitationPurves and Seltzer 1986) compared with the splenium, or whether these differences are a result of composition of regional fibers. Fibers oblique to the x,y,z directions would appear as decreased FA values denoting less white matter when in actual fact these oblique fibers could be more dense than fibers in the x,y,z direction. The white matter underlying the frontal lobes is notoriously heterogeneous in terms of the directions in which fibers are orientated due to the highly top-down and distributed nature of the connections within virtually all areas of the cortex throughout the brain (CitationDenes and Pizzamiglio 1999). Finally, in the Yurgelun-Todd study there was no effect for white matter hyperintensities; that is, only 1 of the 11 bipolar subjects displayed even 1 hyperintensity. When this subject was removed from the analysis, observed differences between patients and controls persisted in the genu (CitationYurgelun-Todd et al 2007).
Another study of bipolar disorder subjects demonstrated significantly less fractional anisotropy in the anterior cingulate (p = 0.003) in the largest sample to date (CitationWang et al 2008). However the clinical value of this finding is questionable. The actual magnitude of the difference indicated that bipolar disorder subjects had only 7% lesser FA values than their normal counterparts in the anterior cingulum. In the posterior cingulum no significant FA difference levels were found between groups. However there were small absolute levels of differences between groups in the posterior cingulate measurements.
Therefore, for white matter tracts the important factor may well be functional connectivity (CitationGoebel et al 2004) and it does not seem likely that small FA differences would on the whole lead to reduced functionality. This is because anterior and posterior cingulum FA value differences were of the same magnitude and basal levels of FA were within the same range. Moreover, absolute FA values in the anterior cingulum are fraught with measurement idosyncrasies since many oblique fibers are present and automated tractography vectoring was not used. Indeed, asymmetrical white matter connectivity has been recently demonstrated in a DTI tractography study by CitationHouenou and colleagues (2007), implying that careful consideration of micro- and macrostructural white matter asymmetries may be important for group and single-case analysis.
White matter diffusion tensor imaging tractography
Of unique interest in terms of the neuroanatomical basis of the disorder was the finding by CitationHouenou and colleagues (2007) that only in bipolar disorder patients was the left subgenual cingulate (SC) and amygdalo-hippocampal (AH) complex hyperconnected. That is, a leftward asymmetry was observed with respect to the number of reconstructed fibers between the SC and AH [t(15) = −3.27, p = 0.05]. The authors speculated that this connectivity most probably represented the uncinate fasciculus since that is the most important connection between the temporal pole and the ventral prefrontal or orbitofrontal cortex (CitationEbeling and von Cramon 1992), innervating central nuclei of the amygdala with the orbitofrontal regions. In addition, this leftward asymmetry was manifested in a twofold increase in the volume of the left unicinate fasiculus in bipolar disorder patients compared with normal healthy controls [t(30) = −2.73, p = 0.01]. Since both numbers of fibers and volume of fibers was significant implies that Houenou’s tractography study is suggestive of a trend. Recall that the number of fibers and the volume of a white matter tract and its function are dependent on the: (i) diameter of axons within a tract; (ii) the proportion of small, medium, and large axons, (iii) the specific intrahemispheric or commissural tract in question which vary in fiber composition and thus density (CitationClarke and Zaidel 1994; CitationAboitiz et al 1992; CitationPurves and Seltzer 1986; CitationDenes and Pizzamiglio 1999).
The findings of reduced white matter density in the subgenual regions of the cingulate and frontal white matter give rise to another line of inquiry (CitationWang et al 2008). Perhaps the reduced diffuse anisotropy in the frontal lobes is a consequence of localized left uncinate hyper-connectivity; assuming that total white matter in an individual brain stays somewhat constant. Moreover, distributions relating to the number of reconstructed fibers normalized for seed mask volume between the left subgenual cingulate and hippocampus/amygdala was twice to fourfold greater in bipolar subjects compared to healthy controls (). This potential difference in number of fibers and thickness of this tract suggests that it is functional and not an anatomical artifact. These findings of uncinate hyperconnectivity are in contrast to specific studies showing reduced fractional anisotropy of the fasciculus in schizophrenia patients suggesting a fundamental distinction in etiology between the two disorders (CitationKubicki et al 2002).
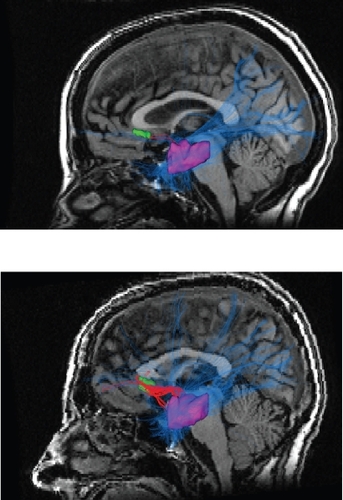
Figure 1 In the top picture is the diffusion tensor imaging tractography output for a healthy control. In the bottom picture is a representative tractography output from a bipolar disorder patient. All patients were on lithium monotherapy or combined therapy consisting of lithium, atypical neuroleptics and selective serotonin reuptake inhibitors. The bipolar disorder sample consisted of 8 males and 8 females with a mean age of 41 years. Note the much greater density and thickness of the red colored fibers in the bipolar disorder subjects in the bottom picture. The red tortuous pathway consists of reconstructed fibers connecting the left subgenual cingulate which is depicted in green and the left amygdalo-hippocampal complex depicted in purple. The blue lines represent reconstructed fibers connecting the left amygdalo-hippocampal complex to the rest of the brain. Copyright © 2007, Nature Publishing Group. Reproduced with permission from CitationHouenou J, Wessa M, Douaud G, et al. 2007. Increased white matter connectivity in euthymic bipolar patients: Diffusion tensor tractography between the subgenual cingulate and the amygdalo-hippocampal complex. Mol Psychiatry, 12:1001–10.
Validity of diffusion tensor imaging ROI approaches?
Jones and colleagues conducted a technical and methodological study of diffusion tensor imaging (DTI) white matter tract characteristics and of tractography using schizophrenia patients with known anatomical abnormalities (CitationJones et al 2006). The results highlight a number of limitations with voxel-based mapping and region of interest approaches using DTI. Among the methodological findings was that age was a significant covariate which if not incorporated into the analysis could skew the results in favor of higher or lower FA and mean diffusivity values in the patient groups. The age-specific effects on fractional anisotropy and mean diffusivity have been previously found in a well-defined cognitive skill (reading) with characteristic developmental peaks and plateaus in performance indices (CitationBeaulieu et al 2004; CitationSnook et al 2004). DTI studies must use age as a covariate in developmental or neuropathologic studies; many previous DTI studies have not used such an approach. However, as the authors of this study note there are some problems with DTI studies of fractional anisotropy and mean diffusivity using only region of interest (ROI) or voxel-based mapping; although it must be kept in mind that the best course of action is for DTI experimenters to use the method which is most congruent with the goals of the study (CitationWang et al 2008).
In the face of criticism regarding the weaknesses in region of interest or voxel-based mapping of diffusion tensor imaging within a single plane, some of the originators of the technique of DTI advocated for the superiority of diffusion tensor tractography. These experts noted that “…compared to the conventional approach of taking measurements from manually drawn ROIs, our method [DTI-MRI-tractography] presents several advantages: (1) it allows us to investigate specific tracts that are hypothesized to be involved in the pathogenesis of [the disorder in question]; (2) it provides data from the entire tract rather than from a limited portion of it; and (3) it is less prone to operator-induced bias…” (CitationJones et al 2006, p. 235). In sum, experts in the use of diffusion tensor imaging have shown that in a specific clinical sample of neuropsychiatric patients diffusion tensor imaging magnetic resonance tractography possesses clinical research potential. It is only a matter of time before such rich sources of qualitative information are readily and widely available to the neuroradiologist, behavioral neurologist, and cognitive neuropsychologist.
When tractography and region of interest or volume of interest methods were compared, the hypothesis that tractography could provide unique information was borne out in the CitationHouenou and colleagues (2007) study. It was found that fibers between the pons and the cerebellum belonging to the cerebellar peduncle were successfully reconstructed, whereas nonexistent fibers between the subgenual cingulate and the pons were not reconstructed. A recent theoretical review of the topic by specialists made several compelling arguments for the use of tractography over DTI regions of interest analysis (CitationKanaan et al 2006).
CitationKanaan and colleagues (2006) used a novel quantitative diffusion tensor tracking algorithm which differed from Houenou’s diffusion tensor deflection method in a number of ways. Kanaan’s method was verified in schizophrenia patients and compared with both region of interest and voxel-based analysis. It has a number of advantages over DTI-deflection in that (i) principal orientations of the diffusion tensor uses an automated B-spline-interpolated continuous tensor field, (ii) the fibers were tracked in both directions, and finally (iii) these reconstructed fibers were compared for neuroanatomic accuracy by comparison with in-house MAT-LAB visualization scripts and neuroanatomic atlases. The highly experienced London Institute of Psychiatry DTI group have thus developed an important method for improving the tractography method and therefore the Houenou tractography findings are provocative but require replication using different methodologies and across laboratories. That said it is important to mention that the Houneou findings show some signs of reliability and validity since single subject maps showed significant cross-subject agreement.
These authors noted conceptual and practical difficulty with ROI and voxel-based mapping including difficulties in replication across methods, high variability in white matter tracts and lack of standard anatomical references. The authors speculated that contradictory findings with respects to ADC, (apparent diffusion coefficient or rate of speed of diffusion) and FA in frontal white matter could be entirely artifactual and that the true gold-standard for connectivity analysis may be individualized and group DTI tractography analysis. It seems tentatively then that the subgenual cingulate – amygdalo – hippocampal connections are a reliable finding and can explain many associated symptoms of bipolar disorder. However it is unclear what the functional correlates of such hyperconnected white matter tracts are.
The discovery of a ventral semantic stream
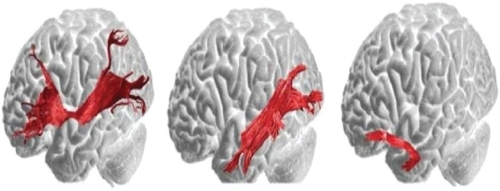
Are there dual routes to language as in the visual system? Electrocortical and subcortical stimulation studies suggest a dorsal phonological route and ventral semantic stream coursing from the anterior temporal lobe to the frontobasal region (CitationMandonnet et al 2007). Intraoperative direct stimulation was used by the investigators to confirm anatomofunctional correlations at cortical and subcortical levels in patients undergoing left temporal pole gliomas. It is important to mention that aside from the glioma, there was no reason to suspect that subjects had any atypical patterns of white matter connectivity. Indeed, CitationCatani and ffytche (2005) have provided maps, depicted in below, of the ventral stream semantic pathways in normal healthy human subjects. If the inferior occipitofrontal and the inferior longitudinal fasciculus are essential pathways in the ventral semantic stream, then an understanding of temporal pole functions which are connected to the ventral prefrontal cortex directly and indirectly by the uncinate fasciculus will be required.
Figure 2 Left illustration depicts the direct pathway from the posterior cortices to the prefrontal cortex. The inferior occipitofrontal fasciculus connects the posterior temporal areas and the orbital and lateral prefrontal regions. Middle illustration consists of the first segment of the indirect pathway. The inferior longitudinal fasciculus links the posterior occipitotemporal regions with the temporal pole and this fiber tract courses underneath the inferior occipitofrontal fasciculus. Right illustration depicts the second component of the indirect pathway. The uncinate fasciculus connects the anterior and medial temporal lobe with the orbitofrontal regions. Copyright © 2005, Oxford University Press. Adapted with permission from CitationCatani M, ffytche DH. 2005. The rises and falls of disconnection syndromes. Brain, 128:2224–39.
Cortical – subcortical stimulation was carried out in 17 glioma patients. These presurgical mapping studies provided evidence in favor of the existence of a main pathway underlying the semantic system located in the left hemisphere (CitationDuffau et al 2005). This pathway joins the posterior and superior temporal areas with the inferior and dorsolateral prefrontal cortex and is identified as the inferior fronto-occipital fasciculus. The fasciculus narrows at the junction of the frontal and temporal lobes and passes through the anterior floor of the external capsule with terminal radiations reaching the middle and inferior temporal gyri as well as all the way to lingual and fusiform gyri. Electrostimulations anywhere along the route in the superficial white matter underlying cortical areas or in deep subcortical nuclei resulted in semantic paraphasias. Due to the density of mappings, the accuracy of magnetic resonance imaging (MRI) guided point locations with the aid of human brain atlases, and the elicitation of nonsemantic paraphasias (eg, phonemic paraphasias) in deviations off this route, the authors named this fasciculus a de facto “semantic loop”.
CitationVigneau and colleagues (2006) in their review of 129 phonology, sentential or semantic processing neuroimaging studies published between 1992 and 2004, postulated that the temporal pole and the anterior portion of the fusiform gyrus were part of a semantic network that was in part instantiated by the inferior longitudinal fasciculus. This fasciculus is also known as the occipitotemporal fasciculus, which links the posterior inferior temporal region with the temporal pole. Refined semantic information was then hypothesized to be relayed by the temporal pole through the uncinate fasciculus on through to the orbitofrontal cortex. The inferior longitudinal fasciculus (ILF) runs laterally and inferiorly to the lateral wall of the temporal horn whereas the inferior occiptofrontal fasciculus runs just medial of the ILF and above the optic pathways. Speculation and controversy arose about whether the indirect fiber pathway between the inferior-posterior occiptotemporal region and the temporal pole consisted of straight line fibers or consecutive U-shaped fiber junctions (CitationTusa and Ungerleider 1985). Diffusion tensor imaging tractography studies of the human occipitotemporal lobe have since confirmed that both straight line fibers and looping U-shaped fibers are present, thereby conferring in the ILF a potential fast and slow computational route.
Although stimulation of the inferior occipitofrontal fasciculus did result in reliable semantic paraphasias, electrical stimulation of the inferior longitudinal fasciculus did not induce any naming disturbances; nor did resection of a portion of it cause language problems (CitationMandonnet et al 2007). However, the authors note that lack of an effect for resection or stimulation does not mean that the ILF is not involved in semantics; due to plasticity from slow-growing lesions the functions could conceivably be redistributed within the ipsilateral hemisphere or contralateral homotypic regions (CitationHeilman and Valenstein 2003). On the basis of their electrostimulation findings, the investigators hypothesized that there were two pathways constituting a ventral semantic stream (CitationMandonnet et al 2007). A direct dominant pathway consisting of the occipitofrontal fasciculus connecting posterior temporal regions and the prefrontal cortex; as well as an indirect pathway consisting of the inferior longitudinal fasciculus (posterior occipitotemporal → temporal pole → uncinate fasciculus → orbitofrontal cortex). Finally, CitationDuffau (2008) makes the interesting observation that at the level of sentence comprehension the inferior direct occipitofrontal fasciculus has been found to make links with phonology and linguistic comprehension units, implying a role for this ventral semantic stream in language acquisition and development (CitationSakai 2005).
Functional neuroanatomy of the temporal pole
CitationArnold and colleagues (1994) showed that neuropathological changes to the temporal pole occur in Alzheimer’s and Pick’s disease, and CitationBraak and colleagues (1993) demonstrated that during the initial stages of Alzheimer’s, the temporal pole is among the first regions to show neurofibrillary tangles. Episodic memory impairments are considered among the earliest symptoms of the onset of Alzheimer’s disease (CitationBassett et al 2006) which is congruent with the functions of the temporopolar cortex (CitationNakamura and Kubota 1996). Cholinergic modulation of the anterior temporal pole could play an essential role in its functions, since CitationFlynn and Mash (1986) reported that the affinity and concentration of nicotinic acetylcholine receptors was decreased in this region in Alzheimer’s disease patients. Like electrical stimulation of the medial temporal lobe, recent electrophysiological studies have shown that electrical stimulation of the anterior ventromedial temporal lobe can also result in the generation of simple or complex seizures (CitationMunari et al 1993).
In (on the next page), the ventral temporopolar cortex is represented by TPv and the perirhinal cortex is comprised of Brodmann’s area 35 and 36. Damage to the ventral temporal pole produces loss of memory for familiar objects or associative agnosia but not apperceptive agnosia, suggesting that the resemblance of objects including faces associated with personal recollective memory, is bound here. TEO is involved in the construction of singular objects and TE is critical for visual recognition and memory of objects, whereas V4 is involved in color perception. TH and TF appear to be related to spatial and tactual function and together are sometimes referred to as the parahippocampal cortex involved in place learning.
Figure 3 Copyright © 1996, Elsevier. Adapted with permission from CitationNakamura K, Kubota K. 1996. The primate temporal pole: Its putative role in object recognition and memory. Behav Brain Res, 77:53–77.
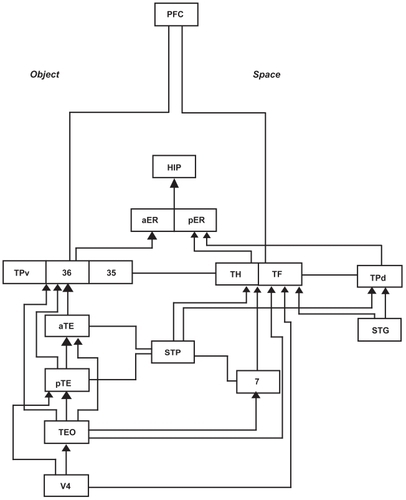
Brodmann’s area 7 is the superior parietal lobe involved in spatial location memory encoding and STP refers to the superior temporal polysensory area which runs posterior to anterior and is involved in complex auditory and spatial processing. TPd is the dorsal temporal pole and is involved in auditory spatial object construction with the superior temporal gyrus usually associated with Wernicke’s area language comprehension functions. The anterior and posterior entorhinal cortex provides the major route for information into the hippocampus. Interestingly, the temporal pole does not receive direct innervation from the prefrontal cortex, but rather dorsal routes are through the parahippocampal cortex and ventral routes are through the perirhinal cortex involved in recognition memory.
Temporal pole and theory of mind
The temporal pole (TP) receives projections from three sensory systems within the temporal lobe proper. The dorsolateral TP receives projections from the third-order auditory association cortex whereas the ventral TP receives projections from the extrastriate cortex at the terminal end of the occipitotemporal cortex (CitationOlson et al 2007). An important role for the medial aspect of the temporal pole has been found in the awareness of internal somatic states through afferents received from the insula (CitationCritchley 2004). Some of the most interesting studies to date on the temporal pole function have had to do with the organic induction of rapidly vacillating mood states with TP damage. CitationMurai and Fujimoto (2003) found that brain injury to the TP resulted in rapidly cycling bipolar disorder and CitationGlosser and colleagues (2000) established that anterior temporal resections, especially within the right lobe were apt to produce depression, anxiety, and organic mood disorder.
Three separate reports, as noted by Olson, have found that anterior temporal pole damage can cause acquired bipolar disorder (CitationJorge et al 1993; CitationBrooks and Hobyln 2005; CitationCarran et al 2003). Finally, CitationOlson and colleagues (2007, p. 1720) argue that based on reviews of studies of Kluver-Bucy syndrome in monkeys and man, that TP regions are instrumental in supporting group social behaviors such as the tendency to band together to form societies, to participate in family life and to interact with others in their respective social groups (CitationOlson et al 2007). Another ubiquitous finding pertaining to the anterior temporal poles are its involvement with high level face processing, such that perceptual recognition of faces is intact, yet there is impaired multimodal recognition of faces.
CitationBarton and colleagues (2001, Citation2004) have shown that associative prosopagnosia may occur independent of the integrity of memory functions, and that face priming across modalities may be ineffectual with patients with anterior temporal lobe damage. The anterior temporal lobes appear critical then to linking person-specific memories to the structural description of a face and such multi-modal deficits have extended to deficits in recognition by voice (CitationGainotti et al 2003) name of person (CitationSnowden et al 2004), or handwriting (CitationGentileschi et al 2001). This is analogous to loss of access to person-specific knowledge which is a type of high-level semantic deficit. CitationOlson and colleagues (2007) noted that over 19 studies have found reliable temporal polar activation with theory of mind tasks in which another’s thoughts or beliefs must be inferred (CitationFletcher et al 1995a; CitationGoel et al 1995; CitationBaron-Cohen et al 1999; CitationBrunet et al 2000; CitationCastelli et al 2000; CitationGallagher et al 2000; CitationVogeley et al 2001; CitationBerthoz et al 2002; CitationFerstl and von Cramon 2002; CitationGallagher et al 2002; CitationCalarge et al 2003;Sehultz et al 2003; CitationGerman et al 2004; CitationIacoboni et al 2004; CitationOhnishi et al 2004; CitationWalter et al 2004; CitationMitchell et al 2006; CitationSaxe and Powell 2006). Similar types of tasks also have resulted in reliable temporal polar activation. Detection of deception, moral decision making, inference of the emotional state of others, and ratings of how much one feels the negative emotions of others also resulted in polar activation (CitationOlson et al 2007). Some groups have concluded that the temporal polar regions are inextricably linked with mentalizing and that this region is a key element in a general neural network involved in making mental state attributions about the actions of others (CitationFrith 2001; CitationFrith and Frith 2003; CitationVollm et al 2006).
A primary function of the anterior temporal poles then is to associate either the visual ventral temporopolar region, or the auditory dorsal temporopolar region, with visceral emotional responses to facilitate the “…storage of perception-emotion linkages forming the basis of a personal semantic memory…(p. 1725) (CitationOlson et al 2007). There also appears to be some evidence that the temporal polar regions are important for attachment processes, since bilateral TP lesions induce loss of normal emotional attachment in mother monkeys to their infants and to peer monkeys (CitationKling et al 1976). It is possible that propositional knowledge pertaining to the self and others is bound in the temporal pole, and is imbued with emotional connections via proximity with the amygdala-hippocampal complex, resulting in emergent properties of attachment, personal semantic memory and attendant formation of social groups.
Temporal pole and narrative
In ten separate studies involved in basic semantic processes associated with narrative the temporal pole has been shown to be reliably activated (CitationDuffau 2008). These include categorical decisions pertaining to the living versus nonliving character of visually presented words and pictures (CitationBright et al 2004) or retrieval of a word associated with an intransitive gesture (CitationDamasio et al 2001). CitationJames and Gauthier (2004) found that retrieval of previously learned semantic features of a novel object reliably activated the temporopolar cortex, and CitationNoppeney and Price (2004) similarly made this discovery when subjects were visually presented with abstract word triads and had to choose which of the two words were most alike.
Visual presentation of four different categories of words (CitationPerani et al 1999), passive listening to words (CitationPrice et al 1996) or visual presentation of single nouns and judgements as to whether the noun could apply to a human being or not (CitationScott et al 2003), all activated TP. An n-back task requiring subjects to determine if the current object is the same as the previously displayed object (CitationSevostianov et al 2002), and a semantic association task requiring subjects to match word and picture triplets of the same category for meaning (CitationVandenbergh et al 1996), both resulted in a TP signal. In sum, high-level object-orientated actions imbued with abstract categorical semantics were particularly apt to activate the TP; particularly in PET studies where signal fade-out due to inhomogeneity artifacts were not likely to skew the results.
More specifically, CitationDuffau (2008) reviewed studies directly implicating the TP in the integration of a coherent narrative structure. When subjects judged the plausibility of metaphors or sentences (CitationBottini et al 1994), or when they read stories (CitationFletcher et al 1995b), read sentences (CitationStowe et al 1998), or were simply listening to intelligible semantic content (CitationScott et al 2000), the TP was activated in every instance. If subjects were required to integrate the content of a sentence such that lexical decisions were made on sentences ending with an expected versus unexpected word, a temporopolar region signal resulted (CitationBaumgaertner et al 2002). Passive listening to stories, or tasks requiring participants to provide the number of narrators in different stories (CitationCrinion et al 2003), or reading a text in a nonnative language (CitationVingerhoets et al 2003), all resulted in foci in the TP. Attending to a speaker’s voice (Citationvon Kriegstein et al 2003), or judging the syntactic or semantic plausibility of Chinese or English sentences resulted in reliable activation in TP (CitationLuke et al 2002).
In a recent study listening to narrative texts activated the posterior-anterior extent of the left superior temporal sulcus as far forward as the temporal pole (CitationCrinion et al 2006), and critically a Wernicke’s area lesion also impaired anterior left temporal lobe function. The implication is that clinical studies of language in stroke patients have underestimated the role of the left anterior temporal pole in comprehension of narrative speech since routinely only lesion studies have been carried out on these patients and only a functional neuroimaging study could demonstrate such a “virtual lesion”. Passages of prose were read twice to subjects during successive scans with the requirement to remember them. Narratives consisted of either standard readily comprehensible stories or unusual stories for which the global theme was difficult to extract. Mental frameworks for understanding the stories were provided in the form of relevant, irrelevant or no visual cues. CitationMaguire and colleagues (1999) found that both the medial ventral orbitofrontal cortex and the left anterior temporal pole were associated with general processes of comprehension of the narratives, with accompanying mental templates for understanding (CitationMaguire et al 1999).
Orbitofrontal cortex connectivity
In their influential review of the functional neuroanatomy of the orbital frontal cortex CitationZald and Kim (1996) note that this “nose-brain” could be derived from two ancestral lines of cortex. They postulated that an archicortex moiety arose from ancient hippocampal formation structures involved in spatial localization, and that a paleocortex moiety evolved from a primitive olfactory core responsible for identifying objects. Early single-cell stimulation monkey studies showed that neurons within the orbital frontal cortex had receptive fields which were bilateral, exhibited azimuthal sensitivity and were sensitive to moving stimuli. Many of these stimuli were either excited or inhibited by auditory sounds such that for both vision and audition a variety of on and off effects was found. Some of these cells showed audiovisual interaction presumably due to convergence at the cortical cell level. In one example, an auditory stimulus of a characteristic frequency had a powerful inhibitory effect on many neurons and this inhibitory effect was found to negate the excitation caused by a simultaneous visual stimulus (Benevento et al 1997). Subsequently it was shown that both complex patterns and objects can elicit such effects in the orbitofrontal cortex (CitationWilson et al 1993).
There appears to be one functional cortical zone within this transitional region and it is comprised of three key regions: the temporal pole, insula, and orbitofrontal cortex (CitationMorecraft et al 1992). The mediodorsal nucleus of the thalamus is the primary source of thalamic innervation of the orbitofrontal cortex (CitationRay and Price 1993). In addition to the direct connection between the mediodorsal (MD) nucleus and the orbitofrontal cortex, there are seven separate indirect pathways from MD to [substrate x] to orbitofrontal cortex. These intermediaries through the indirect pathway include the amygdala, primary olfactory cortex, entorhinal cortex, perirhinal cortex, temporal pole, superior temporal gyrus, inferior temporal gyrus and the insula. It is hypothesized that the direct route carries detailed perceptual information whereas projections through the MD possess more integrated refined and multimodal information. Little known is that the orbitofrontal cortex is the only region that possesses a strong association with the amygdala. The lateral nucleus of the amygdala is a crucial weigh station for sensory input and the central nucleus is a source of major input into the brain stem and hypothalamic structures. Collectively, these amygdalic structures control species-specific survival responses coordinated with endocrine, autonomic and involuntary behavioral action correlates (CitationDavis 1992).
A critical and often overlooked point is that of how the prefrontal cortex exerts control over the hippocampal formation and associated structures. The only major prefrontal afferent to the hippocampal area is from the orbitofrontal cortex to the entorhinal cortex (EC), and although the EC is intimately connected with the hippocampus the orbitofrontal cortex does not possess direct connections with the hippocampus (CitationVan Hoesen et al 1975). This suggests a terminal pathway ultimately meeting at the orbitofrontal cortex site of modulation for all prefrontally-mediated alterations of mnemonic functions associated with the medial temporal lobe. Finally, dense projections from the medial and lateral orbitofrontal cortex and gyrus rectus reach the lateral hypothalamus – one of the major output pathways of the limbic system (CitationJohnson et al 1968). The orbitofrontal cortex has direct projections with the frontal eye fields and dorsolateral prefrontal cortex such that visual search and working memory also can be influenced by the value of stimuli (CitationMorecraft et al 1992).
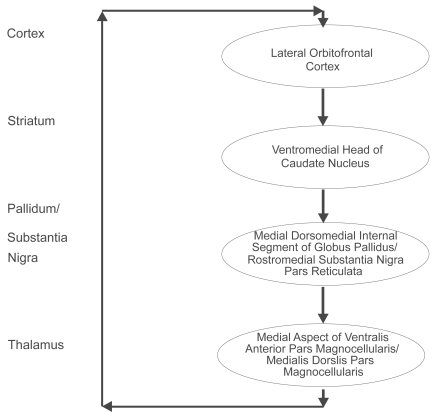
Five distinct types of basal ganglia-thalamocortical circuits have been described and the lateral orbitofrontal circuit is most relevant to an understanding of the basal ganglia-cortical loops () (CitationAlexander et al 1986). By way of example, bilateral lesions of the lateral orbitofrontal area, and/or lesions to the caudate nucleus to which the lateral orbitofrontal afferents fibers project, will result in perseverative interference with a monkey’s capacity to make appropriate switches in behavioral set (CitationDivac and Manning et al 1967; CitationMishkin et al 1978). Hence an essential function of the basal ganglia is to integrate the sensory input arriving from the neocortex with contextually appropriate motor or cognitive functioning programs. The striatum sends projections to the globus pallidus and the substantia nigra. The direct striatal-globus pallidus/substantia nigra pathway inhibits the globus pallidus/substantia nigra output nuclei and thus causes disinihibition of the thalamus. Activation of the direct loop facilitates the triggering of programs at the cortical level through double inhibition. Discrete portions of the mediodorsal nucleus of the thalamus are important in emotional and cognitive functioning through their connections with the orbitofrontal and dorsolateral prefrontal cortex, respectively (CitationGoldman-Rakic and Porrino 1985). Lesions to this feedback system can result in disinhibition of emotional circuits leading to depression, mania and/or obsessive compulsive disorder (CitationChamberlain et al 2005).
Figure 4 The lateral orbitofrontal basal ganglia-thalamocortical circuit. The ventromedial head of the caudate nucleus receives input from the superior temporal gyrus, the inferior temporal gyrus and the anterior cingulate area. The closed loop of the lateral orbitofrontal circuit is completed by the return from these two regions of the thalamus to the lateral orbitofrontal cortex. Copyright © 1986. Redrawn and adapted from CitationAlexander GE, DeLong MR, Strick PL. 1986. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci, 9:357–81.
shows how the medial orbitofrontal loop follows a unidirectional course thereby allowing affectively valued stimuli to influence motor programs via the basal ganglia. Recall that affective valuation is different from affective valence since valence denotes an instinctual approach or avoidance behavior, whereas valuation can be abstract without stimulus features and connotates instrumental behavior. The interaction of the medial temporal hippocampal – amygdalic structures and the orbitofrontal cortex is of a magnitude greater in terms of complexity than the separate study of either of these regions. Effective and functional connectivity studies using functional MRI may be especially illuminating with respect to the functional neuroanatomy of orbital and temporal pole hyperconnected regions in the bipolar patient. For instance the ventromedial head of the caudate nucleus is the source of afferent fibers emanating from the auditory stream of the superior temporal gyrus, the visual stream of the inferior temporal gyrus and the anterior cingulate gyrus. Connections between these regions allows the imbuement of stimulus-specific value and context-specific determination of reward and hence goal states in the brain.
Figure 5 The medial orbitofrontal – basal ganglia loop. Copyright © 1996, American Psychiatric Publishing. Adapted with permission from CitationZald DH, Kim SW. 1996. Anatomy and function of the orbital frontal cortex. I: Anatomy, neurocircuitry, and obsessive-compulsive disorder. J Neuropsychiatry Clin Neurosci, 8:125–38.
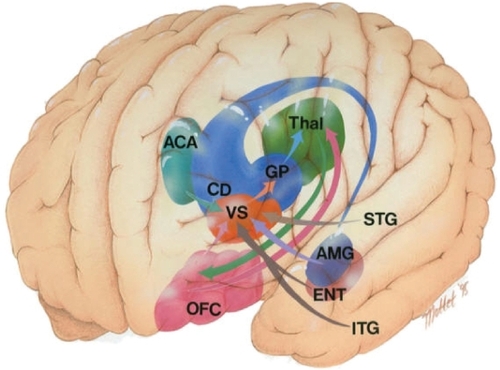
The medial orbitofrontal cortex (pink) has direct reciprocal connections with the accessory basal amygdala (purple) (CitationCarmichael and Price 1995). The ventromedial temporal pole area 38 is also reciprocally connected with the medial aspect of the orbitofrontal cortex (CitationCarmichael and Price 1995; CitationMoran et al 1987; CitationVogt and Pandya 1987). The auditory association areas of the dorsal temporal pole are reciprocally connected with the lateral orbitofrontal cortex (CitationCarmichael and Price 1995; CitationMoran et al 1987; CitationVogt and Pandya 1987). Finally the lateral orbitofrontal cortex has reciprocal connectivity with the area TE and frontal eye fields; this highlights the OFC control over sensorimotor processing (CitationCarmichael and Price 1995).
Orbitofrontal cortex review
A neuroeconomical view of the function of the orbitofrontal cortex is that it represents the relative value of potential choices at a reductionistic level (CitationPadoa-Schioppa 2007). Moreover, this view suggests that orbitofrontal cortex neurons encode economic value independently of visuomotor contingencies, consistent with an abstract object-based model rather than one action-based. Lesions of the ventromedial prefrontal cortex often have been associated with reversal learning (CitationRoberts 2006). Reversal learning is exemplified by the Iowa Gambling Task wherein subjects must choose between 4 decks of cards (CitationBechara et al 1997). On each trial a card is drawn and the subject either wins or is provided with a win and a loss. As CitationFellows (2005, Citation2007) noted two of the decks are associated with large wins but even larger losses and the other two decks are associated with smaller wins but even smaller losses. The second set of decks is the conservative choice or advantageous one. Critically, the order of cards in each deck is fixed so that after large wins on several sets of consecutive trials, (on the disadvantageous decks), large losses begin to accrue. After large losses accrue, healthy and nonorbitally lesioned patients choose spontaneously to change to the advantageous decks, whereas orbitally lesioned patients perseverate on the decks which initially provided large wins.
To demonstrate more generally that the Iowa Gambling Task (IGT) is dependent on reversal learning required an ingenious design change that eliminated the requirement for reversal learning (CitationFellows and Farah 2005). Two conditions were incorporated into the new design. There was the original IGT and a shuffled version of the IGT in which the cards were shuffled so that there was no opportunity for delayed reversal learning. In the original design, ventromedial lesion patients performed poorly; in the same task where there was no reversal learning switch, they performed normally. Interestingly, the dorsolateral prefrontal lesioned patients performed poorly in both tasks, perhaps as the consequence of generalized working memory impairment. These results show conclusively that difficulties that ventromedial lesion patients exhibit in the study, result not from impaired decision making, but rather from reversal learning as a deficit in flexible reinforcement using knowledge of contingencies. Finally, CitationFellows (2007) noted that the orbitofrontal cortex also may be involved in a simpler type of task in which the subject must determine the relative value between a number of choices. This preference judgment was found not to correlate with reversal learning, suggesting that the two tasks may rely on different subregions of the orbito-frontal cortex.
Orbitofrontal cortex: Stimuli, rewards, and future actions
The orbitofrontal cortex is involved in at least five different separate functions. Firstly, the orbitofrontal cortex encodes the value of stimuli as these stimuli are perceived, allowing for goal-weighting and goal/sub-goal structuring. Such valuation encoding allows for long-term and immediate future planning, specifically with respect to reinforcement contingencies. Secondly, the orbitofrontal cortex is involved in maintaining expectations of future reward thereby providing information on specific courses of action or particular stimulus choices. The orbital regions also play an essential role in coordinating error signals with midbrain regions, which could conceivably underlie the learning of reward predictions. The orbital gyri are also involved in encoding the underlying implicit rules or structure of an abstract problem in order to guide predictions of future reward. However such future reward encoding does not constitute decision making per se but rather implicit goal weight structuring or simple logical operator functions. Finally, orbital regions appear to function in computing reward relevant decisions as well as in representing the reinforcement contingencies of a particular decision in association with dorsolateral prefrontal structures (CitationO’Doherty 2007).
CitationO’Doherty (2007) notes that among the main functions of the orbital gyri or orbitofrontal cortex is assisting in making adaptive decisions under conditions of uncertainty, as well as in the face of changing contingencies (CitationO’Doherty 2007; CitationBechara et al 1994; CitationRolls et al 1994; CitationHornak et al 2004). Subsequent studies show that neurons within the orbitofrontal cortex encode the state-dependent reward value of not only intrinsically hedonic olfactory (CitationAnderson et al 2003) and gustatory stimuli (CitationSmall et al 2003), but also that of more neutral somatosensory, auditory, and visual stimuli. In this context, the perceptual-emotional associations formed in the anterior temporal pole could be the terminal neural network input upon which the relative valuation orbitofrontal network operates.
The orbitofrontal cortex also has also been shown to respond to secondary reinforcers such as money or social praise (CitationBreiter et al 2001; CitationElliott et al 1997). However, the orbitofrontal cortex responds not only to rewards but also to aversive stimuli like foul tastes or actual monetary loss in economic games (CitationO’Doherty et al 2001a; CitationO’Doherty et al 2001b). Moreover, there seems to be a medial/lateral parcellation of function across the orbitofrontal cortex. Both CitationO’Doherty and colleagues (2001a) and CitationUrsu and Carter (2005) found that the medial aspects of the orbitofrontal cortex blood flow increased in response to rewards and that lateral aspects of the brain region were found to respond to broad classes of aversive stimuli (CitationUrsu and Carter 2005). Other functional neuroimaging studies have made similar discoveries, suggesting that the medial/lateral distinction might be a general feature of functional organization in the orbitofrontal cortex (CitationO’Doherty 2007).
Similarly, avoidance of an aversive outcome activated the medial orbitofrontal cortex and conversely, failure to obtain a reward activated the lateral orbital surface (CitationKim et al 2006) suggesting some type of encoding of logical operators within the region. At least two studies have shown that anticipation of receipt of rewards or highly aversive stimuli, in association with predictive cues, led to anterior and rostral orbitofrontal cortex activation (CitationO’Doherty et al 2002; CitationGottfried et al 2002). As with the increasingly abstract future-focused behavior gradiations found within the posterior to anterior temporal lobe, a similar type of anticipatory behavior continuum appears to operate within the orbitofrontal cortex. Finally, distinctions have been made by CitationO’Doherty (2007) between stimulus substitution and signalling predictive mechanisms that could well involve some implicit nonconscious algorithm functions in the orbitofrontal cortex (CitationO’Doherty 2007). The essence of this finding is that it is extremely adaptive for the animal or human organism to have a predictive mechanism that would signal an impending affectively laden event without eliciting a conscious representation of that event.
To always require conscious, rather than unconscious and automatic representations, is computationally cumbersome, inefficient, and slow and therefore maladaptive in life or death situations. This signalling mechanism would in effect allow for the animal to distinguish cues that predict a stimulus from those of the actual unconditioned stimulus (eg, rewarding food versus aversive stimuli). This is because different behavioral responses are elicited when anticipating an event as opposed to immediately experiencing it. As CitationO’Doherty (2007, p. 260) notes if “…stimulus substitution were to be the only mechanism in place then a CS would be indistinguishable from the UCS from the point of view of the animal…”. To provide a good example of this, if there was only stimulus substitution and not also predictive signalling then the animal would try to eat the conditioned stimulus (CS) (eg, light) rather the unconditioned stimulus (UCS) (eg, food). The predictive signalling component involves differentiated approach and anticipatory responses to the light as opposed to the undifferentiated approach response and consummatory behavior directed towards the primary reinforcer of food.
Similarly, a CSfor a food reward can involve a behavioral approach, avoidance, or orientation responses, distinct from those produced by the unconditioned stimulus itself. The basic idea is that not all unconditioned responses are identical to those conditioned responses elicited by the primary or secondary reinforcer. Moreover, reflexive responses are tied to second signal systems including semantics, operant responses and procedural learning. Under these circumstances, a motor response conditioned to semantic stimulus is not only possible, it is highly probable. Such preconscious responding enables differentiation of salient UCS’s from exceedingly complex background contexts. Finally, CitationGottfried and colleagues (2003) found that the anterior orbitofrontal cortex encoded UCS abstracted de-contextualized value rather than the sensory properties of a stimulus per se (CitationGottfried et al 2003). Related to this delayed de-contextualized encoding of raw value magnitudes, is the finding by CitationTremblay and Schultz (1999) that orbitofrontal neurons increased in spiking responses during the delay of the expectation of rewards. Hence if bipolar disorder is associated with the enhanced connectivity between anterior temporal pole and orbitofrontal cortex brain regions it is still a requirement to determine what are the functional, behavioral, and cognitive manifestations associated with the illness.
Functional neuroanatomy of bipolar disorder
CitationStrakowski and colleagues (2005, p. 105) note in their review of the functional neuroanatomy of bipolar disorder that “…structural measurements may not have clear functional correlates…”. This statement summarizes the concept of supervenience. Supervenience is a term used to describe a broadly physicalist and nonreductive philosophy of mind. In essence it means that two identical brains states cannot then differ in mental qualities, or conversely that something cannot alter in a mental state without altering in some physical respect. However, the reverse scenario is not necessarily true. In other words, psychological properties can be multiply realized in many physical properties or different brains states (CitationDavidson 2001). Thus, it is the configuration of spatially and temporally discontinuous modules distributed throughout the brain and interacting in concert in real-time, that defines a mental state. A mere structural abnormality does not have the effect of causality. For example, in bipolar disorder, until it can be shown that structural difference has reliable and proximate causal effects in the generation of a specific dysphoric trait a single region of interest abnormality hypothesis is untenable.
CitationStrakowski and colleagues (2005, p. 114) in their recent summary of neuroimaging findings pertaining to bipolar disorder argue that it is fundamentally a disorder of effective or functional connectivity (CitationGoebel et al 2004).
“…Imaging studies taken together highlight that it is unlikely that bipolar disorder will localize to abnormalities within a single neuroanatomic structure. The brain is not organized into discrete independent functional packets, but, instead consists of complex, interconnected neural networks. Therefore, dysfunction in any network will echo throughout the brain in complex and, currently, unpredictable activations. Nonetheless, taken together, these studies suggest that, in bipolar disorder, there may be a relatively diminished prefrontal modulation of subcortical and medial temporal structures within the anterior limbic network (eg, amygdala, anterior striatum and thalamus), which results in dysregulation of mood… (p. 114).
In this context, a functional lesion in any pathway is bound to adversely effect the optimal functioning of a pre-frontal mediated neural network, instantiating spatiotemporally distributed higher-order cognitive processes. Indeed,CitationHouenou and colleagues (2007; p. 1002) suggested that rather than focusing on a restricted region of the cortex or the subcortex, an approach more congruent with contemporary models of effective connectivity “…might be more reliable to investigate the characteristics of an entire white matter tract with recently developed techniques such as diffusion tensor tractography rather than exploring a single region of a fiber bundle…”. Quantitative tractography can provide multiple measures with respect to reconstructed fiber tracts such as (i) number of reconstructed fibers, (ii) mean fractional anisotropy, and (iii) mean apparent diffusion coefficient of fibers.
Bipolar disorder and neuropsychological findings
CitationFrantom and colleagues’s elegant 2008 study of euthymic bipolar disorder patients (BP), their unaffected first-degree relative (FD) and healthy controls (HC) is among the most comprehensive in the literature (CitationFrantom et al 2008). The most important finding was that the first-degree relatives scored intermediate between bipolar patients (worst) and normals (best) on 6 of 7 cognitive domains thereby providing strong evidence of a cognitive endophenotype associated with the disorder (CitationGottesman and Gould 2003). Significant differences were found on Block Design (CitationRinge et al 2002) and Benton Judgment of Line Orientation Test (CitationBenton et al 1978) between bipolar disorder and normal healthy controls. Bipolar disorder subjects scored on average 3 scaled score points lower on Block Design than HC’s, while BP’s scored 22.8/30 versus HC’s 26.4/30 on Judgment of Line Orientation. Visuospatial and visuo-constructional deficits have been reported previously in bipolar disorder subjects (CitationSapin et al 1987; CitationGruzeleir et al 1988; CitationCoffman et al 1990; CitationBulbena and Berrios 1993; CitationTham et al 1997; CitationAtre-Vaidya et al 1998; CitationRubinsztein et al 2000; CitationEI Badri et al 2001).
In terms of localizing capability, Judgment of Line Orientation results in increased blood flow in the right temporo-occipital region (CitationHannay et al 1987), whereas the Block Design subtest is a test of constructional praxis, sensitive to right parietal lesions (CitationWarrington et al 1986). CitationFrantom and colleagues (2008) were careful to note that no difference was found on Judgment of Line Orientation performance in discordant monozygotic twins, one of which was afflicted with bipolar disorder (CitationGourovitch et al 1999). The lack of a difference on a test of estimation of the incidental angle between two lines segments in genetically identical subjects, suggests that Judgment of Line Orientation is not part of the core bipolar endophenotype. The two different tasks are clearly not measuring the same abilities (CitationDenes and Pizzamiglio 1999; CitationLezak 1995), and by default then, the core deficit must be constructional praxis.
The bipolar group was found to be impaired significantly on the Wisconsin Card Sort Test perseverative errors score (CitationHeaton et al 1993), and the Trail Making Test Part B (CitationReitan and Wolfson 1985) in Frantom and colleagues’ study, suggesting some loss of mental flexibility. Although the level of general intelligence was similar for the BP, FD, and HC in both the Frantom and Gourovitch twin studies, there were no group differences on either the Wisconsin Card Sort Test or Trails B in the twin study, casting doubt on the claim that a genetically-based executive function deficit is part of the bipolar affective disorder endophenotype. The executive dysfunction hypothesis negation gains support from the finding that no differences were recorded on categories achieved on the WCST or on Category Fluency (CitationReitan and Wolfson 1985) or Letter Fluency Tests (CitationBenton and Hamsher Kde 1989) in the Frantom study.
Of particular interest were the covariate findings that pertain to the hypothesis that some symptomatology might be due to the adverse effects of trait-dependent mood dysregulation. Each subject was tested on the Hamilton Depression Rating Scale (CitationHamilton 1960) and the Young Mania Rating Scale (CitationYoung et al 1978); all BP’s were medicated and no subjects met clinical indicators for mania or depression. However, when these two indicators were used as covariates the Visual Learning and Memory domain was no longer significant, paralleling the nonsignificant findings for any Verbal Learning and Memory measures. This is a helpful clinical finding and suggests that while manic-depressive symptoms may interfere negatively with verbal and visual learning and memory processes, vocational decisions such as return to work or educational initiatives could well be feasible once mood has stabilized.
This recent study’s nonsignificant findings pertaining to Verbal Learning and Memory is contrary to studies showing that deficits in this domain are the most reliable neurocognitive markers of BP (CitationDenicoff et al 1999; CitationAli et al 2000; CitationClark et al 2001; CitationZubieta et al 2001). Many of these latter studies were not as well-controlled as the Frantom study suggesting that the recent demonstration of a lack of verbal and visual memory deficits is a genuine effect. A meta-analysis with bipolar disorder subjects demonstrated an effect size of 0.90 across 10 studies using the California Verbal Learning Test (CitationDelis et al 1987; CitationRobinson et al 2006). However this bipolar sample had a low average of only 2.9 hospitalizations, and an onset of 24 years, with all subjects fully employed or attending university. One conclusion that could be drawn from this collective work is that neuropsychological deficits are by no means the norm, especially for subjects in the upper ability spectrum with less severe and chronic illness features.
CitationFrantom and colleagues (2008) did find an advantage for left, as opposed to right hemisphere cognitive function composite factor, but no such effect for simple motor function. A right hemisphere disadvantage, as well as a specific visuospatial function deficit among the seven neurocognitive domains, was recorded in aggregate variable findings. The results of this well-designed study appear generalizable due to the fact that much of the previous literature related to neuropsychological deficits in bipolar disorder subjects was reviewed and integrated into the study. Finally, this study addressed a number of the limitations of previous studies including: (i) the use of a comprehensive neuropsychological battery, (ii) closely matched participants on all demographic variables, (iii) the requirement for clinical stability, (iv) the use of normal controls and first degree relatives as well as (v) the use of valid research diagnostic strategies. In view of the previous literature and careful systematic review of it, it was found that performance on Block Design was significantly impaired in BP subjects, consistent with a right-hemisphere constructional apraxia.
A large population level study of 47 bipolar disorder families used a battery of laterality and relative hand skill tests, along with a quantitative transmission disequilibrium test (QTDT) and a genetic analysis of blood samples (CitationAbecasis et al 2000). CitationSavitz and colleagues (2007) tested for the degree of handedness as a proxy for atypical lateralization in 55 bipolar disorder I patients and 185 bipolar spectrum patients and unaffected relatives. The hypothesis was that schizophrenia, and to a lesser extent bipolar disorder are caused by a disruption in the normal process of hemispheric specialization (CitationCrow 2000), such that the components of language which are normally localized to the left hemisphere, become bilaterally represented. The end result is that psychotics “…lose the ability to discriminate between what could be said (thought), what is said (speech output) and what is heard (speech input). Thought is confused with linguistic input or output producing feelings of thought insertion, thought broadcast and auditory hallucinations…’(p. 698) (CitationSavitz et al 2007).
However, others such as CitationDewan and colleagues (1987) have found normal structural cerebral asymmetry in bipolar disorder subjects, suggesting an etiological difference between schizophrenia and bipolar disorder. Hence, the hypothesis of deficit lateralization in bipolar disorder subjects has not been supported entirely. The CitationSavitz and colleagues (2007) study is a major forward advance in the understanding of the disorder. First of all the South African group determined that the BP cohort was significantly more lateralized on the Relative Hand Skill Test compared with asymptomatic BP relatives (CitationTapley and Bryden 1985), and on the Waterloo Handedness and Footedness Inventories (CitationElias and Bryden 1998). This finding echoes Dewan’s sentiments that there are important differences between schizophrenia and bipolar disorder. In the past, the general consensus was that the proportion of mixed and left handers, connotating bilateral language representation, was significantly higher in people with schizophrenia (CitationDragovic and Hammond 2005; CitationSommer et al 2001).
However, the putative link between bipolar disorder and schizophrenia is not limited to structural MRI, neuropsychological or psychiatric symptom profile findings. Recent genome-wide scans have shown that the two disorders have linkages to 2q22.1-q23.3, 8p22, 13q31, and 22q11–13, G72/G30, BDNF, and COMT (CitationBadner and Gershon 2002; CitationMaier et al 2005). These genes are widely distributed in the normal population, and hence, important interactions among the various combinations of these alleles have not yet been examined. In the CitationSavitz and colleagues (2007) study, 10 separate genes shown to have an association with bipolar disorder or bipolar disorder/schizophrenia, were assayed for in BP and first-degree relatives. The catechol-O-methyl-transferase (COMT) gene was found to significantly associate with the Relative Hand Skill test. The finding that bipolar disorder I subjects are more strongly lateralized than those with schizophrenia, implies fundamental differences in the nature of the two mental illnesses. A finding that schizophrenia is a disorder of too little hemispheric lateralization of function, and that bipolar disorder is an illness of too much lateralization, still would be congruent with CitationCrow’s (2000) thesis.
The authors speculate that excessive left-lateralized language mechanisms result in bipolar disorder, presumably as a consequence of the rs++ genotype (CitationCorballis 1998). This would be consistent with Bilder and colleague’s finding of greater asymmetry in bipolar as opposed to schizophrenic subjects (CitationBilder et al 1999). However, this hypothesis presumes that only extremely left lateralized configurations are possible and not those of extreme right lateralization. If CitationAnnett’s (1985) hypothesis that rs−− random environmental events will cause one of the hemispheres to become linguistically dominant, then 50% will display left-hemisphere and 50% will display right-hemisphere dominance (CitationAnnett 1985). In this context, the rs++ and the rs−− genotype would both be expected to produce an illness propensity-inducing endophenotype with the former much more prevalent than the latter. To digress, the Relative Hand Skill (RHS) test consists of a series of circles arranged in a repeated pattern (CitationTapley and Bryden 1985). The subject is asked ‘to make a dot in each circle, following the pattern as quickly as you can’. The test is similar in some respects to coding subtests of the Wechsler scales without any elementary letter or symbol manipulatory requirements (CitationNalcaci et al 2001). For this reason the RHS test is a reliable genotypic marker and a neuropsychological test primarily of sensorimotor agility.
The poor performance of bipolar disorder subjects on the Block Design subtest and their superior performance on the RHS test could be consistent with the hypothesis that a motor speed/agility versus constructional praxis trade-off is occurring. COMT is highly localizable in terms of function to the prefrontal cortex (CitationGogos et al 1998), and particularly with regard to regulating dopamine in the striatum (CitationSesack et al 1998). Of interest with respect to neuropsychological function in bipolar disorder, was that the COMT genotype alone accounted for four percent of the variance in Wisconsin Card Sort Test perseverative errors, among individuals with schizophrenia (CitationEgan et al 2001), and it was found to have significant effects on a factor described as speed of processing and attention (CitationBilder et al 2002). It appears, then, that the COMT genotype plays an important role in determining the baseline dopaminergic tone in the prefrontal cortex, and that this relationship, in terms of performance, is an inverted U-shaped curve (CitationMattay et al 2003).
However, there is still a problem with the interpretation of rs−− and rs++ as two sides of the same distribution. Many recent studies have shown extreme rightward lateralization to be associated with normal or even superior intelligence and a mirror reversed pattern of spatial and linguistic functions in some, but not all subjects (CitationKnecht et al 2001; Citation2003 CitationFloel et al 2005). Crowding effects manifest as decreased constructional praxis skills (eg, Block Design); these effects are found in some, but certainly not all, atypically lateralized subjects (CitationKnecht et al 2001, Citation2003; CitationFloel et al 2005). Clearly other complex factors are involved in the etiology of bipolar disorder aside from lateralizing genes, since extreme lateralization is not necessarily associated with either neurological or psychiatric, or intellectual disabilities. Moreover, the recent findings are of theoretical interest; if the archetypal configuration of extreme asymmetry is not by necessity associated with bipolar disorder, this much alone suggests that it cannot be a causal, but can be only a contributing variable in the context of other genotypes or disease processes. Finally, extreme leftward lateralization, although possible to measure in theory with a large enough sample using transcranial doppler or fMRI, has not been systematically studied to date ().
Table 1 An organizational framework for understanding the relationship between cognitive phenotypes encountered with normal and atypical language localization. This is only one potential configuration and many others are possible considering the heterogeneous nature of bipolar disorder
Although complete reversibility in the lateralization of cognitive function has been previously reported, it is, however, extremely rare (CitationFischer et al 1991). Comprehensive single-case studies of such phenomenon in bipolar patients and atypical subjects are nonexistent. These atypical localization cases or unique co-localization scenarios can be instructive with regard to the fundamental and deep principles by which the cerebral cortex is organized. CitationBasso and colleagues (1985) was among the first to note that such subject matter can be highly informative to this line of inquiry. It is no coincidence that the publication of these landmark studies occurred with the widespread dissemination of high resolution computed tomography (CT) scan technology.
Moreover, CitationBasso and her colleagues (1985) noted that no comprehensive theory espousing the organization of language or cognitive function could be considered complete without a theory that accounted for the “anomalies”. CitationFischer and colleagues (1991) took a secondary step in this natural inquiry, and questioned how are different cognitive and behavioral operations related to each other in terms of functional outcome? The derivative and highly pertinent offshoot of this question is whether the intrinsic characteristics or cognitive functions of unexpectedly displaced modules change as a function of either (i) dislocation or (ii) co-localization within an unexpected hemisphere? Single and group case study of anomalous subjects using fMR-adaptation, fMRI, DTI, and cognitive neuropsychological methods could help illuminate both (i) mechanisms responsible for bipolar disorder and atypical localization, as well as those (ii) basic principles of how the brain is organized and lateralized. Thus, genetic alleles conferring some degree of bipolar susceptibility and neuropsychological endophenotypes have been described. However it is not entirely clear why such widespread and disseminated genetic markers of such traits would continue to persist in the general population.
The evolutionary significance of temperamental mood dysphoria
Traits must be explained in terms of the effects of increasing allele frequencies at a population level (CitationNesse 2006), or how vulnerabilities persist in the gene pool (CitationNesse and Williams 1994). Since low mood occurs in all cultures and reliably effects behavior; the question is how do vulnerabilities persist in the gene pool (CitationMorris 1992). The peak onset of depression is early adulthood during the achievement of major life goals (CitationWeissman et al 1996), stressful events that exacerbate low mood (CitationKessler 1997), with a normal distribution from dyphoria to depression (CitationRuscio and Runci 2002) highlighting the usefulness of dimensional models (CitationAngst and Merikangas 2001). The organizing determinant for low mood is the environmental epoch that gave rise to it (CitationTooby and Cosmides 1990), and loss is at the core of most clinical depression. Therefore cognitive modeling of the antecedents of loss provides short-term evolutionary benefits (CitationNesse 2006). These benefits include an impetus to regain lost resources; motivation to do so; avoidance of eliciting situation; protection of other resources; adjustment of strategies; prevention of future losses; or making amends for accrued losses.
Moderate low mood can facilitate superior social judgement (CitationBadcock and Allen 2003), and low mood, characteristic of neurosis has adaptation effects (CitationHartung 1988). One finding is that striving for status and low mood are causally related; when goals are impeded one needs a way to determine if success is possible; or one needs a means to disengage from obsolete goals; and perhaps re-establish new goals (CitationNesse 1999). Mood provides a measure to determine progress towards the achievement of goals (CitationCarver and Scheier 1990), and its homeostatic regulation is designed for careful investment in the allocation of resources (CitationNesse 1999). Key resources include relationships, status, and kinship behavior (CitationKrebs and Davies 1997). Disengagement, associated with low mood, allows for implementation of more productive strategies (CitationKlinger 1975) or re-appraisal of central life goals (CitationGut 1989). It is theorized that dysphoric genes have selective advantages which keep them at high basal allelic frequencies in the population (CitationHoule 1998). Moreover, it appears that absolute mood level is not optimal, but rather, the key is the facility with which a well-designed mood homeostatic mechanism selects the correct behavior at the right time. Sensation-seeking is a good example, since exposure to life events is in part due to shared genetic heritage and therefore kin groups may select the same environment eliciting the same behavioral phenotypes (CitationKendler and Karkowski-Shuman 1997).
Hypomania can result is sustained productivity or enhanced creativity in kin-groups (CitationWilson 1998). Given that the deleterious effects for bipolar are worse than unipolar depression it appears that temperamental disposition effects could be at work. Moreover, it is possible that such traits reliably lead to financial success and/or selection for physically attractive mates (CitationAndreasen 1987: CitationRichards et al 1988). In such a context, if the mood regulator acts as thermostat, then oscillation is naturally to be expected in cycles (CitationNesse 2006). Therefore, bipolar disorder could be conceptualized (i) as a defective regulation of positive feedback cycles or, (ii) more threshold level sensitivity in the servo-control mechanism’s normal variance in fluctuation. The anecdotal evidence that bipolar disorder patients have particular personality and temperamental features and could be perhaps more “sensitive” than others appears congruent with recent theoretical models and empirical findings.
Creativity and bipolar disorder
Creativity requires a minimum level of high general intelligence, a well-developed fund of knowledge both across and within a specific domain, as well as specialized skills. However, these three components of creativity are insufficient. What does appear to be essential is the ability to develop alternative solutions to problems or the ability to use divergent thinking skills. The frontal lobes are known to be crucial in the generation of alternative strategies, just as the temporal and parietal cortices are essential in the storage of specialized knowledge (CitationHeilman et al 2003). Heilman and colleagues hypothesize that finding the “thread that unites” requires the binding of different forms of knowledge stored in different regions and modules of the brain that might not normally be associated. Therefore, atypical localization of cognitive functions, and the attendant co-association of brain regions that are not normally connected through white matter tracts, might be essential for creative innovation (CitationFischer et al 1991). The high co-morbidity between both unipolar and bipolar mood disorder and creative innovation, at least within the upper quartile of the general ability range, suggests that alterations in specific neurotransmitters might be a key explanatory variable to consider.
The threshold theory of IQ and creative innovation have been reviewed elsewhere (CitationBarron and Harrington 1981), and suggest that below an IQ of 120 is important and causally related to creative innovation. However, above a minimum IQ of 120 there is no relation between creativity and intelligence. The threshold theory would explain why William Shockley, who was the inventor of the transistor and a Nobel prize winner, and Pablo Picasso, Albert Einstein, and Charles Darwin were all noted by their primary school teachers to be dull, and in at least one case, scoring low on intelligence tests (CitationHeilman 2005, p. 3). Therefore, general intelligence or IQ is necessary but it is not sufficient to explain creative innovation; moreover it is important to note that the link between creativity, bipolar disorder, and the achievement of eminence within a domain is just as strong in women.
CitationHeilman (2005) notes that a high general level of background knowledge within a domain has been consistently found to be a pre-requisite for creative innovations or works. In the example of our creative geniuses above, all spent formative years perfecting and honing disparate areas of knowledge, expertise and skill before their discoveries were made. Einstein spent years studying math and physics while working at the Swiss patent office, whereas Picasso spent years first learning how to mix paints and draw forms. Finally, Darwin developed an encyclopedic knowledge of the structure and functions of previously uncatalogued species on his two-year voyage around the world.
There are some theoretical and practical reasons to suspect that the largest white matter tract in the brain may have a role in creativity. As Heilman and colleagues (p. 374) note, Rorschach tests of patients who had undergone cerebral commissurotomy revealed that they did not score high on this projective test of creativity (CitationHeilman et al 2003; CitationLewis 1979). This is one of the few positive neuropsychological findings on high level cognitive processes recorded post-commissurotomy, aside from simple divided visual field, dichotic listening, or dichaptic tests. The originators of the commissurotomy for the treatment of intractable epilepsy note that incomplete hemispheric communication enabled lateralized cognition and hemispheric independence, which might be essential for the incubation of ideas in creative innovation (CitationBogen and Bogen 1988). Hence there is reason to speculate that the corpus callosum in the intact state, and in its pathological manifestations, could have an effect on processes associated with creative production.
Writing and bipolar disorder
CitationAndreasen and Glick (1988) noted that bipolar disorder, with its attendant manic episodes and depression, was not causally related to creative work output. For instance, respondents noted that they were unable to work during depressive periods and that work produced during severe mania was of poor quality. Hypomanic episodes were similarly not useful since distractibility, poor concentration, and increased sociability detracted from creative drive. Pickering suggested that periods of illness, particularly depression, may be apt to facilitate the creative process, since it could provide an incubation period during which ideas may develop (CitationPickering 1974). Personality theory is replete with descriptions of creative individuals, and there is a remarkable degree of consistency across studies in the core attributes of: nonconforming, adventuresome, sensitive, introspective, socially detached, self-assertive, intelligent, highly independent and autonomous (CitationMacKinnnon 1965).
These reliable personality variable findings lend support to the hypothesis that it is the temperamental factors, rather than the manic-depressive illness per se, that is responsible for the co-association of creativity and the achievement of eminence, by high ability bipolar probands. Similarly, the affective disorder patient experience of suffering may enhance the creative person’s understanding of the human condition (CitationAndreasen and Glick 1988). CitationAndreason and Glick (1988) note that contrary to mental illness facilitating creativity, what in actual fact occurs is that psychopharmacological treatment of bipolar disorder almost always outweighs the side effects of the medication on the creative process, and that creative acts and works increase and not decrease; particularly with respect to writing, which requires sustained euthymic periods of equilibrium.
Most studies examining the putative relationship between creativity and bipolar disorder have studied eminently creative individuals through either structured interview, semi-structured interview, or purely biographical research methods (CitationAndreasen 1987; CitationJamison 1989; CitationLudwig 1992; CitationLudwig 1994; CitationLudwig 1995; CitationPost 1994). The causal link with eminent samples of bipolar disorder women and men is unequivocal and the effects are large. This sampling method, however, is fraught with technical difficulties. These problems include that such samples are by nature extreme; there no subject interviews, there is routine use of highly subjective biographical review techniques and a failure to use standard instruments. CitationSantosa and colleagues (2007) provide an empirical study to demonstrate the creativity link in bipolar disorder subjects that are noneminent. Santosa’s study is among the first to show an objectively defined causal link between bipolar disorder and enhanced creativity. These findings suggest that the notion of bipolar subjects excellence in the world of arts, literature, and the sciences may have more than a ring of truth. However the question remains, how best to optimally treat the person with bipolar disorder who demonstrates creative and other potential.
Conclusions
The results of this review suggest that for some subtypes of bipolar disorder there may well be aberrant, anomalous, or atypical lateralization of cognitive functions in the brain. The study of bipolar disorder subjects with atypical localization of cognitive functions in addition to the study of healthy subjects with atypical localization of cognitive functions can illuminate some fundamental principles by which the language and other cognitive functions are organized within the cerebral cortex. As noted by several neurolinguists an account of how cognitive function is organized in the human brain cannot be considered complete until these anomalies have been integrated into a comprehensive theory. Biographical, epidemiological, and empirical studies now suggest that creativity is enhanced in bipolar disorder probands and suggests that atypicality and hyperconnectivity of specific brain regions may account for these findings. Several behavioral neurologists have previously hypothesized that such association of brain regions that are not normally connected in the average human brain may be the causal factor involved in enhanced intuition, insight, and the production of original works.
It remains to be determined if the co-association of cognitive functions that are not normally co-localized in such bipolar subjects confers emergent higher-order cognitive functions. This much has been hypothesized previously. Systematic neuropsychological study of high ability bipolar disorder women and men who are eminent artists, writers, and scientists could potentially shed some light on the mechanisms by which the creative brain is structured and how it functions. Comprehensive study of bipolar disorder subjects using DTI tractography, full neuropsychological exams, functional MRI paradigms, systematic psychiatric assessment techniques and controls should help to elucidate the neurophysiologic mechanisms involved in this common mental illness. The effective treatment of higher ability bipolar disorder probands who demonstrate creative potential and works is also a question that should be addressed in future studies given the reliable findings pertaining to personality traits that such people possess as well as their high need for achievement and development of personal goals.
Acknowledgments
Simon M McCrea was a post-doctoral fellow and research associate of the Divisions of Neurology and Neuroophthalmology within the Faculty of Medicine at the University of British Columbia while this manuscript was prepared. The author acknowledges the funding and support of NIMH R01-MH069898 grant to Jason J.S. Barton, MD, PhD during completion of this review. Parts of this manuscript were presented at the VII Annual Meeting of the Vision Sciences Society in Sarasota, Florida from May 11th to May 16th, 2007.
References
- AbecasisGRCardonLRCooksonWO2000A general test of association for quantitative traits in nuclear familiesAm J Hum Genet662799210631157
- AboitizFScheibelABFisherRS1992Fiber composition of the human corpus callosumBrain Res598143531486477
- AlexanderGEDeLongMRStrickPL1986Parallel organization of functionally segregated circuits linking basal ganglia and cortexAnnu Rev Neurosci9357813085570
- AliSODenicoffKDAltshulerLL2000A preliminary study of the relation of neuropsychological performance to neuroanatomic structures in bipolar disorderNeuropsy Neuropsy Be13208
- AndersonAKChristoffKStappenI2003Dissociated neural representations of intensity and valence in human olfactionNat Neurosci619620212536208
- AndreasenNCGlickID1988Bipolar affective disorder and creativity: Implications and clinical managementCompr Psychiatry29207173288437
- AndreasenNC1987Creativity and mental illness: Prevalence rates in writers and first- degree relativesAm J Psychiatry1441288923499088
- AngstJMerikangasKR2001Multi-dimensional criteria for the diagnosis of depressionJ Affect Disorders6271511172869
- AnnettM1985Left, right, hand and brain: The right shift theoryLondonLawrence Erlbaum
- ArnoldSEHymannBTVan HoesenGW1994Neuropathological changes of the temporal pole in Alzheimer’s disease and Pick’s diseaseArch Neurol51145508304839
- Atre-VaidyaNTaylorMASeidenbergMReedRPerrineAGlick-OberwiseF1998Cognitive deficits, psychopathology, and psychosocial functioning in bipolar mood disordersNeuropsy Neuropsy Be111206
- BadcockPBTAllenNB2003Adaptive social reasoning in depressed mood and depression vulnerabilityCognition Emotion1764770
- BadnerJAGershonES2002Meta-analysis of whole-genome linkage scans of bipolar and schizophreniaMol Psychiatryr740511
- Baron-CohenSRingHAWheelwrightS1999Social intelligence in the normal and autistic brain: An fMRI studyEur J Neurosci111891810336657
- BarronFHarringtonDM1981Creativity, intelligence and personalityAnnu Rev Psychol3243976
- BartonJJCherkasovaMO’ConnorM2001Covert recognition in acquired and developmental prosopagnosiaNeurology571161811591830
- BartonJJCherkasovaMHeftnerR2004The covert priming effect of faces in prosopagnosiaNeurology632062815596751
- BassettSSYousemDMCristinzioC2006Familial risk for Alzheimer’s disease alters fMRI activation patternsBrain129112939
- BassoALecoursARMoraschiniS1985Anatomoclinical correlations of the aphasias as defined through computerized tomography: ExceptionsBrain Lang26201292417656
- BaumgaertnerAWeillerCBuchelC2002Event-related fMRI reveals cortical sites involved in contextual sentence integrationNeuroimage167364512169257
- BeaulieuCPlewesCPaulsonLA2004Regional correlation of fractional anisotropy in brain with reading ability in childrenXII Annual Meeting of the International Society for Magnetic Resonance in MedicineMay 15–21Kyoto, Japan
- BecharaADamasioARDamasioHAndersonSW1994Insensitivity to future consequences following damage to human prefrontal cortexCognition507158039375
- BecharaADamasioHTranelDDamasioAR1997Deciding advantageously before knowing the advantageous strategyScience275129359036851
- BeneventoLAFallonJDavisBJRezakM1977Auditory-visual interaction in single cells in the cortex of the superior temporal sulcus and the orbital frontal cortex of the macaque monkeyExp Neurol5784972411682
- BentonALHamsherKVarneyNRSpreenO1978Judgement of line orientationNew YorkUniversity Press
- BentonALHamsherKde S1989Multilingual aphasia examinationIowa CityAJA Associates
- BerrittiniWSteinDJKupferDJSchatzbergAF2006Genetics of bipolar and unipolar disorderTextbook of mood disordersWashingtonAmerican Psychiatric Publishing23547
- BerthozSArmonyJLBlairRJRDolanRJ2002An fMRI study of intentional and unintentional (embarrassing) violations of social normsBrain125169670812135962
- BilderRMVolavkaJCzoborP2002Neurocognitive correlates of the COMT Val(158)Met polymorphism in chronic schizophreniaBiol Psychiatry52701712372660
- BilderRMWuHBogertsB1999Cerebral volume asymmetries in schizophrenia and mood disorders: A quantitative magnetic resonance imaging studyInt J Psychophysiol34199205
- BinderJRWestburyCFMcKiernanKA2005Distinct brain systems for processing concrete and abstract conceptsJ Cognitive Neurosci1790517
- BogenJEBogenGM1988Creativity and the corpus callosumPsychiatr Clin N Am11293301
- BottiniGCorcoranRSterziR1994The role of the right hemisphere in the interpretation of figurative aspects of language. A positron emission tomography studyBrain1171241537820563
- BraakHBraakEBohlJ1993Staging of Alzheimer-related cortical destructionEur Neurol3340388307060
- BreiterHCAharonIKahnemanDDaleAShizgalP2001Functional imaging of neural responses to expectancy and experiences of monetary gains and lossesNeuron306193911395019
- BrightPMossHTylerLK2004Unitary vs multiple semantics: PET studies of word and picture processingBrain Lang894173215120534
- BrooksJOHobylnJ2005Secondary mania in older adultsAm J Psychiatry1622033816263839
- BrunetESarfatiYHardy-BayleMCDecetyJ2000A PET investigation of the attribution of intentions with a nonverbal taskNeuroimage111576610679187
- BulbenaABerriosGE1993Cognitive function in the affective disorders: A prospective studyPsychopathology266128511231
- CalargeCAndreasenNCO’LearyDS2003Visualizing how one brain understands another: A PET study of theory of mindAm J Psychiatry16019546414594741
- CarmichaelSTPriceJL1995Limbic connections of the orbital and medial prefrontal cortex in macaque monkeysJ Comp Neurol363615418847421
- CarranMAKohlerCGO’ConnorMJBilkerWBSperlingMR2003Mania following temporal lobectomyNeurology61770414504319
- CarverCSScheierMF1990Origins and functions of positive and negative affect: A control-process viewPsychol Rev971935
- CastelliFHappeFFrithUFrithCD2000Movement and mind: A functional imaging study of perception and interpretation of complex intentional movement patternsNeuroimage123142510944414
- CataniMffytcheDH2005The rises and falls of disconnection syndromesBrain12822243916141282
- ChamberlainSRBlackwellADFinebergNARobbinsTWSahakianBJ2005The neuropsychology of obsessive compulsive disorder: The importance of failures in cognitive and behavioural inhibition as candidate endophenotype markersNeurosci Biobehav R29399419
- ClarkLIversonSGoodwinG2001A neuropsychological investigation of prefrontal cortex involvement of acute maniaAm J Psychiatry15816051111578991
- ClarkeJMZaidelE1994Anatomical-behavioral relationships: Corpus callosum morphometry and hemispheric specializationBehav Brain Res641852027840886
- CoffmanJABornsteinRAOlsonSCSchwarzkopfSNasrallahHA1990Cognitive impairment and cerebral structure by MRI in bipolar disorderBiol Psychiatry271188962354225
- CorballisMC1998Cerebral asymmetry: Motoring onTrends Cogn Sci21527
- CrinionJTLambon-RalphMAWarburtonEAHowardDWiseRJ2003Temporal lobe regions engaged during normal speech comprehensionBrain126119320112690058
- CrinionJTWarburtonEALambdon-RalphMAHowardDWiseRJ2006Listening to narrative speech after aphasic stroke: The role of the left anterior temporal lobeCereb Cortex1611162516251507
- CritchleyHD2004The human cortex responds to an interoceptive challengeP Natl Acad Sci U S A10163334
- CrowTJMarnerosAAngstJ2000Bipolar shifts as disorders of the bi-hemispheric integration of language: Implications for the genetic origins of the psychotic continuumBipolar disorders: 100 years after manic depressive insanityDordrechtKluwer33548
- DamasioHGrabowskiTJTranelDPontoLLHichwaRDDamasioAR2001Neural correlates of naming actions and of naming spatial relationsNeuroimage1310536411352611
- DavidsonD2001Mental eventsEssays on actions and events: Philosophical essaysNew YorkOxford University Press20728
- DavisM1992The role of the amygdala in fear and anxietyAnn Rev Neurosci15353751575447
- DelisDCKramerJHKaplanEOberBA1987The California Verbal Learning TestNew YorkPsychological Corporation
- DenesGPizzamiglioL1999Handbook of clinical and experimental neuropsychologyEast Sussex, UKPsychology Press
- DenicoffKDAliSOMirskyAFSmith-JacksonEELeverichGSDuncanCC1999Relationship between prior course of illness and neuropsychological functioning in patients with bipolar disorderJ Affect Disorders56677310626782
- DewanMJHaldipurCVLaneEDonnellyMPBoucherMMajorLF1987Normal cerebral asymmetry in bipolar patientsBiol Psychiatry221058663651527
- DivacIRosvoldHESzwarcbartMK1967Behavioral effects of selective ablation of the caudate nucleusJ Comp Physiol Psych6318490
- DragovicMHammondG2005Handedness in schizophrenia: A quantitative review of the evidenceActa Psychiatr Scand111410915877707
- DuffauHGatignolPMandonnetE2005New insights into the anatomofunctional connectivity of the semantic system: A study using cortico-cortical electrostimulationsBrain12879781015705610
- DuffauH2008The anatomofunctional connectivity of language revisited: New insights provided by electrostimulation and tractographyNeuropsychologia469273418093622
- EbelingUvon CramonD1992Topography of the uncinate fascicle and adjacent temporal fiber tractsActa Neurochir1151438
- EganMFGoldbergTEKolachanaBS2001Effect of COMT val105/108 met genotype on frontal lobe function and risk for schizophreniaP Natl Acad Sci U S A98691722
- El BadriSMAshtonCHMoorePBMarschVRFerrierIN2001Electro- physiological and cognitive function in young euthymic patients with bipolar affective disorderBipolar Disord3798711333067
- EliasLJBrydenMP1998Footedness is a better predictor of language lateralization than handednessLaterality3415115513074
- ElliottRFrithCDDolanRJ1997Differential neural response to positive and negative feedback in planning and guessing tasksNeuropsychologia3513954049347486
- FellowsLKFarahMJ2005Different underlying impairments in decision-making following ventromedial and dorsolateral frontal lobe damage in humansCereb Cortex15586315217900
- FellowsLK2007The role of orbitofrontal cortex in decision making: A component process accountAnn NY Acad Sci11214213017846161
- FerstlECvon CramonDY2002What does the frontomedian cortex contribute to language processing: Coherence or theory of mind?Neuroimage17159961212414298
- FischerRSAlexanderMPGabrielCGouldEMilioneJ1991Reversed lateralization of cognitive functions in right handers: Exceptions to classical aphasiologyBrain114245611998885
- FletcherPCFrithCDBakerSCShallliceTFrackowiakRSJDolanRJ1995bThe mind’s eye precuneus activation in memory-related imageryNeuroimage21952009343602
- FletcherPCHappeFFrithUBakerSC1995aOther minds in the brain: A functional imaging study of “theory of mind” in story comprehensionCognition57109218556839
- FloelAJansenADeppeM2005Atypical hemispheric dominance for attention: Functional MRI topographyJ Cerebr Blood F Met251197208
- FlynnDDMashDC1986Characterization of L-[3H] nicotine binding in human cerebral cortex: Comparison between Alzheimer’s disease and the normalJ Neurochem471948543772387
- FrantomLVAllenDNCrossCL2008Neurocognitive endophenotypes for bipolar disorderBipolar Disord103879918402627
- FrithUFrithCD2003Development and neurophysiology of mentalizingPhilos T Roy Soc B35845973
- FrithU2001Mind blindness and the brain in autismNeuron329697911754830
- GainottiGBarbierAMarraC2003Slowly progressive defect in recognition of familiar people in a patient with right anterior temporal atrophyBrain12679280312615639
- GallagherHLHappeFBrunswickNFletcherPCFrithUFrithCD2000Reading the mind in cartoons and stories: An fMRI study of ‘theory of mind’ in verbal and nonverbal tasksNeuropsychologia28112110617288
- GallagherHLJackAIRoepstorffAFrithCD2002Imaging the intentional stance in a competitive gameNeuroimage168142112169265
- GentileschiVSperberSSpinnlerH2001Crossmodal agnosia for familiar people as a consequence of right infero-polar temporal atrophyCognitive Neuropsych1843963
- GermanTPNiehausJLRoartyMPGiesbrechtBMillerMB2004Neural correlates of detecting pretense: Automatic engagement of the intentional stance under covert conditionsJ Cognitive Neurosci16180517
- GlosserGZwilASGlosserDSO’ConnorMMSperlingMR2000Psychiatric aspects of temporal lobe epilepsy before and after anterior temporal lobectomyJ Neurol Neurosur Ps68538
- GoebelRRoebroeckAKimD-SKanwisherNDuncanJ2004A framework for the investigation of directed cortical interactions: Theoretical background and application to dynamic sensorimotor mappingFunctional neuroimaging of visual cognition: Attention and performance XXNew YorkOxford University Press43962
- GoelVGrafmanJSadatoNHalletM1995Modeling other mindsNeuroreport6174168541472
- GogosJAMorganMLuineV1998Catechol-O-methyltransferase-deficient mice exhibit sexually dimorphic changes in catecholamine levels and behaviorP Natl Acad Sci U S A9599916
- Goldman-RakicPSPorrinoLJ1985The primate mediodorsal (MD) nucleus and its projection to the frontal lobeJ Comp Neurol242535602418080
- GottesmanIIGouldTJ2003The endophenotype concept in psychiatry. Etymology and strategic intentionsAm J Psychiatry1606364512668349
- GottfriedJAO’DohertyJDolanRJ2003Encoding predictive reward value in human amygdala and orbitofrontal cortexScience3101104712934011
- GottfriedJAO’DohertyJPDolanRJ2002Appetitive and aversive olfactory learning in humans studied using event-related functional magnetic resonance imagingJ Neurosci22108293712486176
- GourovitchMTorreyEFGoldJMRandolphCWeinbergerDGoldbergT1999Neuropsychological performance of monozygotic twins discordant for bipolar disorderBiol Psychiatry456394610088052
- GruzeleirJSeymourKWilsonLJolleyAHirschS1988Impairments on neuropsychiatric tests of temporohippocampal and frontohippocampal functions and word fluency in remitting schizophrenia and affective disordersArch Gen Psychiatry4562393382322
- GutE1989Productive and unproductive depressionNew YorkBasic Books
- HamiltonM1960A rating scale for depressionJ Neurol Neurosur Ps235362
- HannayHJFalgoutJCLeliDAKathaliCRHalseyJHJrWillsEL1987Focal right temporo-occipital blood flow changes associated with judgement of line orientationNeuropsychologia25755633431672
- HartungJLockardJSPaulhusDDeceiving down1988Self-deception: An adaptive mechanismEnglewood Cliffs, NJPrentice-Hall17085
- HeatonRKCheluneGJTalleyJLKayGGCurtisG1993Wisconsin Card Sorting Test (WCST) Manual – Revised and ExpandedOdessa, FLPsychological Assessment
- HeilmanKMNadeauSEBeversdorfDO2003Creative innovation: Possible brain mechanismsNeurocase93697914972752
- HeilmanKMValensteinE2003Clinical NeuropsychologyFourth EdNew YorkOxford University Press
- HeilmanKMCreativity and the brainNew YorkPsychology Press2005
- HornakJO’DohertyJBramhamJ2004Reward-related reversal learning after surgical excisions in orbito-frontal or dorsolateral prefrontal cortex in humansJ Cognitive Neurosci1646378
- HouenouJWessaMDouaudG2007Increased white matter connectivity in euthymic bipolar patients: Diffusion tensor tractography between the subgenual cingulate and the amygdalo-hippocampal complexMol Psychiatry1210011017471288
- HouleD1998How should we explain variation in genetic variation in traits?Genetica102/103241539720283
- IacoboniMLiebermanMDKnowltonBJ2004Watching social interactions produces dorsomedial prefrontal and medial parietal BOLD fMRI signal increases compared to a resting baselineNeuroimage2111677315006683
- JamesTWGauthierI2004Brain areas engaged during visual judgements by involuntary access to novel semantic informationVision Res444293914680768
- JamisonKR1989Mood disorders and patterns of creativity in British writers and artistsPsychiatry52125342734415
- JohnsonTNRosvoldHEMishkinM1968Projections from behaviourally defined sectors of the prefrontal cortex to the basal ganglia, septum, and diencephalon of the monkeyExp Neurol2120344970775
- JonesDKCataniMPierpaoliC2006Age effects on diffusion tensor magnetic resonance imaging tractography measures of frontal cortex connections in schizophreniaHum Brain Mapp27230816082656
- JorgeERRobinsonRGStarksteinSEArndtSVForresterAWWeislerFH1993Secondary mania following traumatic brain injuryAm J Psychiatry150916218494069
- KanaanRAShergillSSBarkerGJ2006Tract-specific anisotropy measurements in diffusion tensor imagingPsychiatr Res1467382
- KendlerKSKarkowski-ShumanL1997Stressful life events and genetic liability to major depression: Genetic control of exposure to the environment?Psychol Med27539479153675
- KesslerRC1997The effects of stressful life events on depressionAnnu Rev Psychol481912149046559
- KimHShimojoSO’DohertyJP2006Is avoiding an aversive outcome rewarding? Neural substrates of avoidance learning in the human brainPLoS Biol4e23316802856
- KlingASSteklisHD1976A neural substrate for affiliative behaviour in nonhuman primatesBrain Behav Evolut1321638
- KlingerE1975Consequences of commitment to and disengagement from incentivesPsychol Rev82125
- KnechtSDragerBFloelA2001Behavioral relevance of atypical language lateralization in healthy subjectsBrain12416576511459756
- KnechtSJansenAFrankA2003How atypical is atypical language dominance?Neuroimage189172712725767
- KrebsJRDaviesNB1997Behavioral ecology: An evolutionary approachOxford, UKBlackwell Science
- KubickiMWestinC-FMaierSE2002Uncinate fasciculus findings in schizophrenia: A magnetic resonance diffusion tensor imaging studyAm J Psychiatry1598132011986136
- LewisRT1979Organic signs, creativity, and personality characteristics of patients following cerebral commissurotomyClin Neuropsychology12933
- LezakMD1995Neuropsychological assessmentThird editionNew YorkOxford University Press
- LudwigAM1992Creative achievement and psychopathology: Comparison among professionsAm J Psychother46330561530096
- LudwigAM1994Mental illness and creative activity in female writersAm J Psychiatry151165067943456
- LudwigAM1995The Price of GreatnessNew YorkThe Guilford Press
- LukeKKLiuHLWaiYYWanYLTanLH2002Functional anatomy of syntactic and semantic processing in language comprehensionHum Brain Mapp161334512112767
- MacKinnnonDW1965Personality and the realization of creative potentialAm Psychol202738114268806
- MaguireEAFrithCDMorrisRG1999The functional neuroanatomy of comprehension and memory: The importance of prior knowledgeBrain12218395010506087
- MaierWHofgenBZobelARietschelM2005Genetic models of schizophrenia and bipolar disorder: Overlapping inheritance or discrete genotypes?Eur Arch Psy Clin N25515966
- MandonnetENouetAGatignolP2007Does the left inferior longitudinal fasciculus play a role in language? A brain stimulation study?Brain130623917264096
- MattayVSGoldbergTEFeraF2003Catechol O-methyltransferase val158- met genotype and individual variation in the brain response to amphetamineP Natl Acad Sci U S A100618691
- MaybergHSSteinDJKupferDJSchatzbergAF2006Brain imagingThe textbook of mood disordersWashingtonAmerican Psychiatric Publishing21934
- MishkinMManningFJ1978Nonspatial memory after selective prefrontal lesions in monkeysBrain Res14331323415803
- MitchellJPMacraeCNBanajiMR2006Dissociable medial prefrontal contributions to judgements of similar and dissimilar othersNeuron506556316701214
- MoranMAMufsonEJMesulamMM1987Neural inputs to the temporopolar cortex of the rhesus monkeyJ Comp Neurol256881033819040
- MorecraftRJGeulaCMesulamMM1992Cytoarchitecture and neural afferents of orbitofrontal cortex in the brain of the monkeyJ Comp Neurol323341581460107
- MorrisWN1992A functional analysis of the role of mood in affective systemsRev Pers Soc Psychol2173646
- MunariCKahanePTassiL1993Intracerebral low frequency electrical stimulation: A new tool for the definition of the epileptogenic area?Acta Neurochir581815
- MuraiTFuijimotoS2003Rapid cycling bipolar disorder after temporal polar damageBrain Injury17355812637187
- NakamuraKKubotaK1996The primate temporal pole: Its putative role in object recognition and memoryBehav Brain Res7753778762159
- NalcaciEKalayciogluCCicekMGencY2001The relationship between handedness and fine motor performanceCortex3749350011721861
- NesseRMWilliamsGC1994Why we get sick: The new science of Darwinian medicineNew YorkVintage
- NesseRMSteinDJKupferDJSchatzbergAF2006Evolutionary explanations for mood and mood disordersTextbook of mood disordersWashingtonAmerican Psychiatric Publishing15975
- NesseRM1999The evolution of hope and despairJ Soc Issues6642969
- NoppeneyUPriceCJ2004Retrieval of abstract semanticsNeuroimage221647015110006
- O’DohertyJKringelbachMLRollsETHornackJAndrewsC2001bAbstract reward and punishment representations in the human orbitofrontal cortexNat Neurosci495102
- O’DohertyJRollsETFrancisSBowtellRMcGloneF2001aRepresentation of pleasant and aversive taste in the human brainJ Neurophysiol85131521
- O’DohertyJPDeichmannRCritchleyHDDolanRJ2002Neural responses during anticipation of a primary taste rewardNeuron338152611879657
- O’DohertyJP2007Lights, camembert, action! The role of the human orbitofrontal cortex in encoding stimuli, rewards, and choicesAnn NY Acad Sci11212547217872386
- OhnishiTMoriguchiYMatsudaH2004The neural network for the mirror system and mentalizing in normally developed children: An fMRI studyNeuroreport151483715194879
- OlsonIRPlotzkerAEzzyatY2007The enigmatic temporal pole: A review of findings on social and emotional processingBrain13017183117392317
- Padoa-SchioppaC2007Orbitofrontal cortex and the computation of economic valueAnn NY Acad Sci11212325317698992
- PeraniDCappaSFSchnurT1999The neural correlates of verb and noun processing – A PET studyBrain12223374410581226
- PickeringG1974Creative maladyNew YorkOxford University Press
- PostF1994Creativity and psychopathology: A study of 291 world-famous menBrit J Psychiatry16522347953036
- PriceCJWiseRJSWarburtonEA1996Hearing and saying: The functional neuro-anatomy of auditory and word processingBrain119919318673502
- PurvesPSeltzerBLeporeFPittoMJasperHH1986The topography of commissural fibersTwo hemispheres – one brain: Functions of the corpus callosumNew YorkAlan R. Liss773
- RayJPPriceJL1993The organization of projections from the mediodorsal nucleus of the thalamus to orbital and medial prefrontal cortex in macaque monkeysJ Comp Neurol3371317506270
- ReitanRMWolfsonD1985The Halstead-Reitan neuropsychological test battery: Theory and clinical interpretationTucson, AZNeuropsychology Press
- RichardsRLKinnerDKLundeIBenetMMerzelAPC1988Creativity in manic- depressives, cyclothymes and their normal first-degree relatives: A preliminary reportJ Abnorm Psychol9728183192819
- RingeWKSaineKCLacritzLHHynanLSCullumCM2002Dyadic short forms of the Wechsler Adult Intelligence Scale – IIIPsychol Assessment925460
- RobertsAC2006Primate orbitofrontal cortex and adaptive behaviourTrends Cogn Sci10839016380289
- RobinsonLThompsonJGallagherP2006A meta-analysis of cognitive deficits in euthymic patients with bipolar disorderJ Affect Disorders931051516677713
- RollsETHornakJWadeDMcGrathJ1994Emotion-related learning in patients with social and emotional changes associated with frontal lobe damageJ Neurol Neurosur Ps57151824
- RubinszteinJSMichaelAPaykelESSahakianBJ2000Cognitive impairment in remission in bipolar affective disorderPsychol Med2010253612027040
- RuscioJRunciA2002A structure-based approach to psychological assessment: Matching measurement models to latent structureAssessment941611911233
- SakaiKL2005Language acquisition and brain developmentScience3108151916272114
- SantosaCMStrongCMNowakowskaCWangPWRennickeCMKetterTA2007Enhanced creativity in bipolar patients: A controlled studyJ Affect Disorders10031917126406
- SapinLRBerrittiniWHNurnbergerJIJrRothblatLA1987Mediational factors underlying cognitive changes and laterality in affective illnessBiol Psychiatry22979863607138
- SavitzJvan der MerweLSolmsMRamesarR2007Lateralization of hand skill in bipolar affective disorderGenes Brain Behav669870517309660
- SaxeRPowellLJ2006It’s the thought that counts: Specific brain regions for one component of theory of mindPsychol Sci17692916913952
- SchultzRTGrelottiDJKlinA2003The role of the fusiform face area in social cognition: Implications for the pathophysiology of autismPhilos T Roy Soc B35841527
- ScottSKBlankCCRosenSWiseRJ2000Identification of a pathway for intelligible speech in the left temporal lobeBrain1232400611099443
- ScottSKLeffAPWiseRJ2003Going beyond the information given: A neural system supporting semantic interpretationNeuroimage19870612880815
- SesackSRHawrylakVAMatusCGuidoMALeveyAI1998Dopamine axon varicosities in the prelimbic division of the rat prefrontal cortex exhibit sparse immunoreactivity for the dopamine transporterJ Neurosci1826977089502827
- SevostianovAHorwitzBNechaevVWilliamsRFrommSBraunAR2002fMRI study comparing names versus pictures of objectsHum Brain Mapp161687512112770
- SingerWKanwisherNDuncanJ2004Time as a coding space in the cerebral cortexFunctional neuroimaging of visual cognition: Attention and performance XXNew YorkOxford University Press99123
- SmallDMGregoryMDMakYEGitelmanDMesulamMMParrishT2003Dissociation of neural representation of intensity and affective valuation in human gustationNeuron397011112925283
- SnookLPaulsonLARoyD2004Diffusion tensor imaging of neurodevelopment in adolescents and young adultsXII Annual Meeting of the International Society for Magnetic Resonance in MedicineMay 15–21Kyoto, Japan
- SnowdenJSThompsonJCNearyD2004Knowledge of famous faces and names in semantic dementiaBrain1278607214985259
- SommerIAlemanARamseyNBoumaAKahnR2001Handedness, language lateralization and anatomical asymmetry in schizophreniaBrit J Psychiatry1783445111282814
- StoweLABroereCAPaansAM1998Localizing components of a complex task: Sentence processing and working memoryNeuroreport9299599804304
- StrakowskiSMDelbelloMPAdlerCM2005The functional neuroanatomy of bipolar disorder: A review of neuroimaging findingsMol Psychiatry101051615340357
- StrakowskiSMWoodsBTTohenM1993MRI subcortical signal hyperintensities in mania at first hospitalizationBiol Psychiatry3320468448268
- TapleySMBrydenMP1985A group test for the assessment of performance between the handsNeuropsychologia23215214000456
- ThamAEngelbrektsonKMatheAAJohnsonLOlssonEAberg-WistedtA1997Impaired neuropsychological performance in euthymic patients with recurring mood disordersJ Clin Psychiatry582699055834
- ToobyJCosmidesL1990The past explains the present: Emotional adaptations and the structure of ancestral environmentsEthol Sociobiol11375424
- TremblayLSchultzW1999Relative reward preference in primate orbitofrontal cortexNature398704810227292
- TusaRJUngerleiderLG1985The inferior longitudinal fasciculus: A re-examination in humans and monkeysAnn Neurol18583914073852
- UrsuSCarterCS2005Outcome representations, counterfactual comparisons and the human orbitofrontal cortex: Implications for neuroimaging studies of decision- makingCognitive Brain Res235160
- Van HoesenGPandyaDNButtersN1975Some connections of the entorhinal (area 28) and perirhinal (area 35) cortices of the rhesus monkey II: Frontal lobe afferentsBrain Res9525381156867
- VandenbergheRPriceCWiseRJosephsOFrackowiakRSJ1996Functional anatomy of a common semantic system for words and picturesNature38325468805700
- VigneauMBeaucousinVHervePY2006Meta-analyzing left hemisphere language areas: Phonology, semantics, and sentence processingNeuroimage3014143216413796
- VingerhoetsGVan BorselJTesinkC2003Multilingualism: An fMRI studyNeuroimage2021819614683721
- VogeleyKBussfeldPNewenA2001Mind reading: Neural mechanisms of theory of mind and self-perspectiveNeuroimage141708111525326
- VogtBAPandyaDN1987Cingulate cortex of the rhesus monkey II: Cortical afferentsJ Comp Neurol262271893624555
- VollmBATaylorANWRichardsonP2006Neuronal correlates of theory of mind and empathy: A functional magnetic resonance imaging study in a nonverbal taskNeuroimage2990816122944
- von KriegsteinKEgerEKleinschmidtAGiraudAL2003Modulation of neural responses to speech by directing attention to voices or verbal contentCognitive Brain Res174855
- WalterHAdenzatoMCiaramidaroAEnriciIPiaLBaraBG2004Understanding intentions in social interaction: The role of the anterior paracingulate cortexJ Cogn Neurosci1618546315701234
- WangFJackowskiMKalmarJH2008Abnormal anterior cingulum integrity in bipolar disorder determined through diffusion tensor imagingBrit J Psychiatry193126918669996
- WangFKalmerJHEdmistonE2008Abnormal corpus callosum integrity in bipolar disorder: A diffusion tensor imaging studyBiol Psychiatry64730318620337
- WarringtonEKJamesMMaciejewskiC1986The WAIS as a lateralizing and localizing diagnostic instrument: A study of 656 patients with unilateral cerebral lesionsNeuropsychologia24223393714027
- WeisbergRWSternbergRJ1999Creativity and knowledge: A challenge to theoriesHandbook of creativityCambridge, MACambridge University Press22650
- WeissmanMMBlandRCCaninoGJ1996Cross-national epidemiology of major depression and bipolar disorderJAMA27629368656541
- WilsonDR1998Evolutionary epidemiology and manic depressionBr J Med Psychol71375959875952
- WilsonFAScalaidheSPGoldman-RakicPS1993Dissociation of object and spatial processing domains in primate prefrontal cortexScience260195588316836
- YoungRCBiggsJTZieglerVEMeyerDA1978A rating scale for mania: Reliability, validity, and sensitivityBrit J Psychiatry13342935728692
- Yurgelun-ToddDASilveriMMGruberSA2007White matter abnormalities observed in bipolar disorder: A diffusion tensor imaging studyBipolar Disord95041217680921
- ZaldDHKimSW1996Anatomy and function of the orbital frontal cortex. I: Anatomy, neurocircuitry, and obsessive-compulsive disorderJ Neuropsychiatry Clin Neurosci8125389081547
- ZubietaJHugueletPO’NeillRLGiordaniBJ2001Cognitive function in euthymic bipolar I disorderPsychiat Res102920